Visual Pathway
The visual pathway consists of the series of cells and synapses that carry visual information from the environment to the brain for processing. It includes the retina, optic nerve, optic chiasm, optic tract, lateral geniculate nucleus (LGN), optic radiations, and striate cortex (Fig. 15.1). The first cell in the pathway—a special sensory cell, the photoreceptor—converts light energy into a neuronal signal that is passed to the bipolar cell and the amacrine cell and then to the ganglion cell. All these cells and synapses lie within the retina. The axons of the ganglion cells exit the retina via the optic nerve, with the nasal fibers from each eye crossing in the optic chiasm and terminating in the opposite side of the brain. The optic tract carries these fibers from the chiasm to the LGN, where the next synapse occurs. The fibers leave the LGN as the optic radiations that terminate in the visual cortex of the occipital lobe. From various points in this pathway, information about the visual environment is transferred to related neurological centers and to visual association areas.
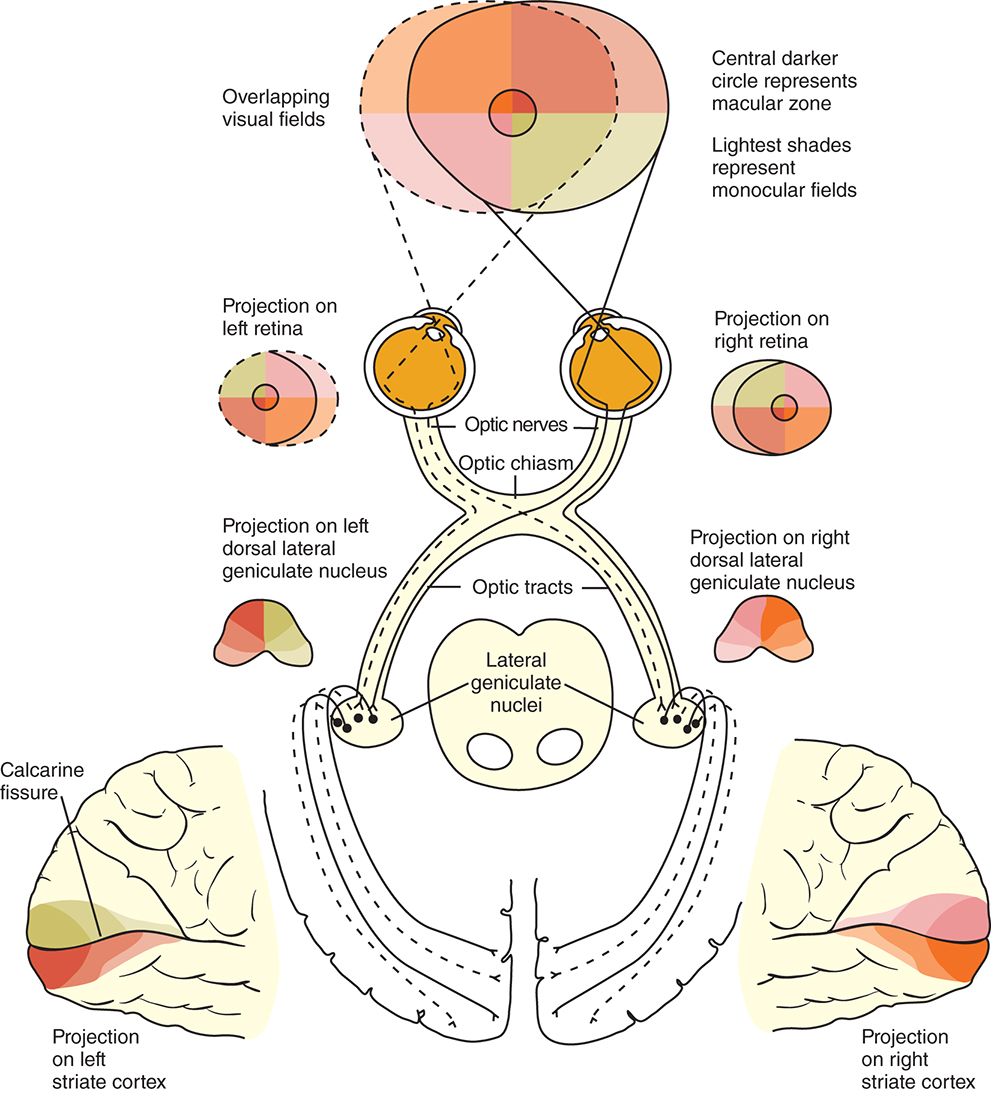
This chapter discusses the structures of the visual pathway and orientation of the fibers within each structure, then it briefly reviews characteristic visual field defects associated with specific locations in the visual pathway. The anatomy of the retina and optic disc are discussed in Chapter 8.
Anatomy of Visual Pathway Structures
Optic Nerve: Cranial Nerve II
The retinal nerve fibers make a 90-degree turn at the optic disc and exit the globe as the optic nerve. This nerve consists of visual fibers, 90% of which will terminate in the LGN. The rest of the fibers, approximately 10%, project to areas controlling pupil responses, circadian rhythm, or the orientation of the head and eyes toward stimuli. Various counts of the optic nerve fibers range from 1 million to 2.22 million, with their size ranging from small-diameter macular fibers to larger-caliber extramacular fibers.1–5
The optic nerve is 5 to 6 cm long and can be divided into four segments on the basis of location: intraocular (0.7–1 mm), intraorbital (30 mm), intracanalicular (6–10 mm), and intracranial (10–16 mm).4,6,7
The intraocular section of the optic nerve can be divided into prelaminar and laminar sections depending on the location relative to the lamina cribrosa. In the prelaminar optic nerve, a glial tissue network provides structural support for the delicate nerve fibers, with sheaths of astrocytes bundling the nerve fibers into fascicles. The orbital portion of the optic nerve contains approximately 928 fascicles.8 That number decreases slightly within the intracranial section.
The border tissue of Elschnig consists of fibrous tissue that extends anteriorly from the edge of the sclera and fuses with Bruch membrane to separate the choroid from ganglion cell axons as they pass through the optic nerve.9–11 The length of this border tissue may correlate with the lamina cribrosa defects and microvascular dropout associated with glaucoma.12 The astrocyte and pia mater form a tissue, the border tissue of Jocoby, that separates the choroid from the optic nerve fibers. As that tissue extends to the outer edge of the retinal nerve fiber layer, it separates the outer retinal layers from the optic nerve fibers and is called the intermediary tissue of Kuhnt.6,13,14 Tight junctions within the glial border tissue may prevent leakage from the adjacent choriocapillaris into the optic nerve head.15 The border tissue is shown in Fig. 15.2.
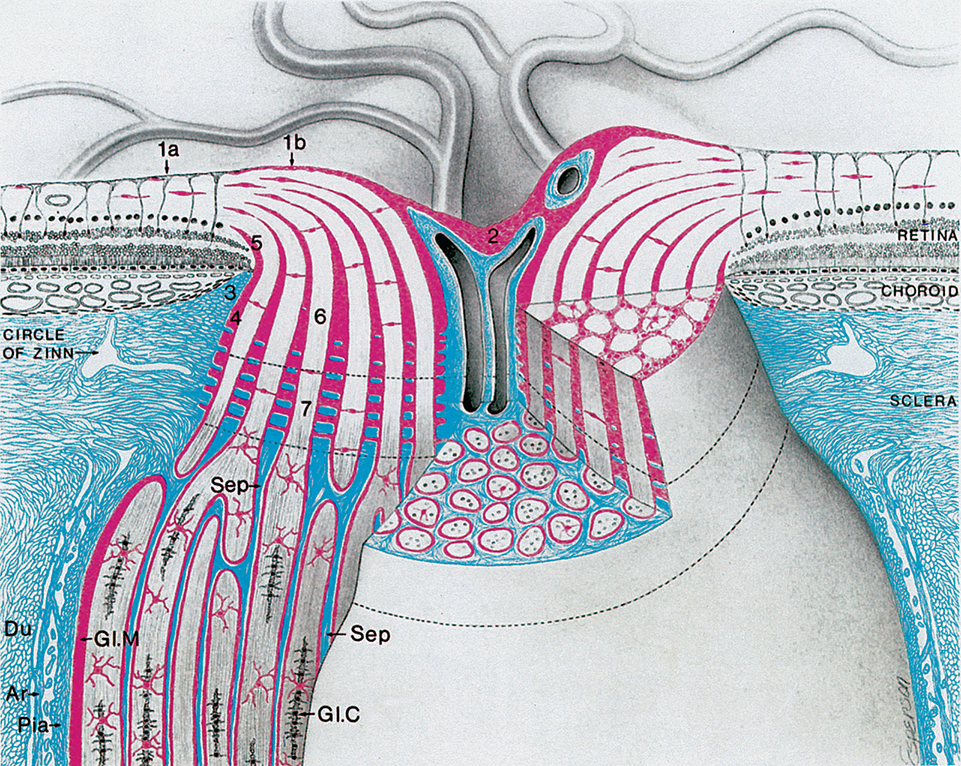
The intraorbital (postlaminar) optic nerve length exceeds the distance from the globe to the apex of the orbit, giving the nerve a slight sine wave shaped curve, allowing for full eye excursions without stretching the nerve. Within the orbit, the nerve is surrounded by the rectus muscles. The sheaths of the superior and medial rectus muscles are adherent to the sheath of the optic nerve (which explains the pain associated with eye movements associated with optic neuritis).
The intraorbital optic nerve is surrounded by three meningeal sheaths continuous with the meningeal coverings of the cranial contents. The outermost sheath, the dura mater, is tough, dense connective tissue containing numerous elastic fibers. Inner to this, a thin collagenous membrane of arachnoid sends a fine network of trabeculae through the subarachnoid space to connect to the innermost layer, the pia mater. The subarachnoid space around the optic nerve is continuous with the intracranial subarachnoid space and contains cerebrospinal fluid. The subarachnoid space is larger directly behind the globe compared with the remainder of its course.16 There is evidence that relatively low cerebrospinal fluid pressure within the orbit compared with the intraocular pressure may play a role in glaucomatous damage.17,18 The loose, vascular connective tissue of the pia mater branches, sending blood vessels and connective tissue septa into the nerve (see Fig. 15.2). Of these sheaths, only the pia continues along the intracranial optic nerve where it runs through the subarachnoid space to the optic chiasm.15,19 The arachnoid does not continue through the optic canal but merges with the pia mater within the canal.20 The dura is continuous with the sclera anteriorly and the periosteum and tendons of the extraocular muscles posteriorly.20
As the unmyelinated retinal fibers pass through the scleral perforations of the lamina cribrosa, they become myelinated by oligodendrocytes, the myelin-producing cells of the central nervous system. It is postulated that the lamina cribrosa is a barrier to oligodendrocytes because myelination does not normally occur in the retina. The sheath of connective tissue, branching from and continuous with the pia mater meningeal covering, is added to the glial sheath of each fascicle posterior to the lamina cribrosa. These additional tissues double the diameter of the optic nerve as it leaves the eye. The nerve is approximately 1.5 to 1.8 mm in diameter at the level of the retina and 3 mm after its exit from the globe, increasing to 4 to 5 mm with the inclusion of the optic nerve sheaths.21–24 The septa that separate the fiber fascicles end near the chiasm.6 Astrocytes present in the optic nerve probably function similar to Müller cells of the retina. They provide structure, store glycogen, and regulate the extracellular concentration of certain ions.
The anterior perforated substance, the root of the olfactory tract, and the anterior cerebral artery lie superior to the optic nerve in its intracranial path. The sphenoid sinus is medial, with only a thin plate of bone separating it from the nerve. The internal carotid artery is below and then lateral to the nerve, and the ophthalmic artery enters the dural sheath of the optic nerve as it passes through the optic canal.
Optic Chiasm
The optic chiasm is roughly rectangular, approximately 15 mm horizontally, 8 mm anterior to posterior, and 4 mm high.4,7,25 As with the intracranial optic nerve, the optic chiasm lies in the subarachnoid space and is surrounded by cerebrospinal fluid.
The chiasm lies within the circle of Willis, a circle of blood vessels that is a common location for aneurysms. The circle of Willis is an anastomotic group of anterior and posterior arteries that join the anterior circulation of the internal carotid arteries with the posterior circulation of the vertebral and basilar arteries (Fig. 15.4). The internal carotid arteries supply the anterior cranial regions, including most of the cerebral hemispheres and orbital and ocular structures. Branches of the vertebral arteries and basilar artery supply the posterior regions, including the brainstem, occipital lobes, and inferomedial temporal lobes, thus supplying most of the ocular motor centers and the cortical visual areas. If the circle of Willis is complete, the anterior cerebral arteries are joined via the anterior communicating artery, and each internal carotid artery is joined to the ipsilateral posterior cerebral artery by a posterior communicating artery. The anterior cerebral and anterior communicating arteries are anterior and superior to the chiasm. An internal carotid artery lies on each lateral side of the chiasm.
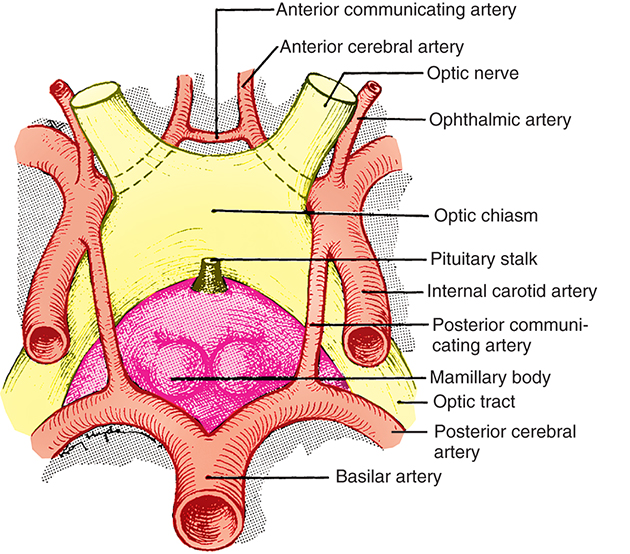
Above the optic chiasm is the hypothalamus and floor of the third ventricle. Approximately 1 cm below the chiasm is the pituitary gland, and the infundibulum lies immediately posterior to the chiasm (Fig. 15.5). The position of the optic chiasm above the sella turcica (the fossa in which the pituitary gland sits) can vary from being directly above it (in 75% of the population) to a position referred to as prefixed (if the optic nerves are short and the chiasm lies above the anterior part of the pituitary gland) or postfixed (if the optic nerves are long and the chiasm is situated toward the posterior part of the pituitary gland). The chiasm is anteriorly displaced in approximately 10% of individuals and posteriorly displaced in 15%.15
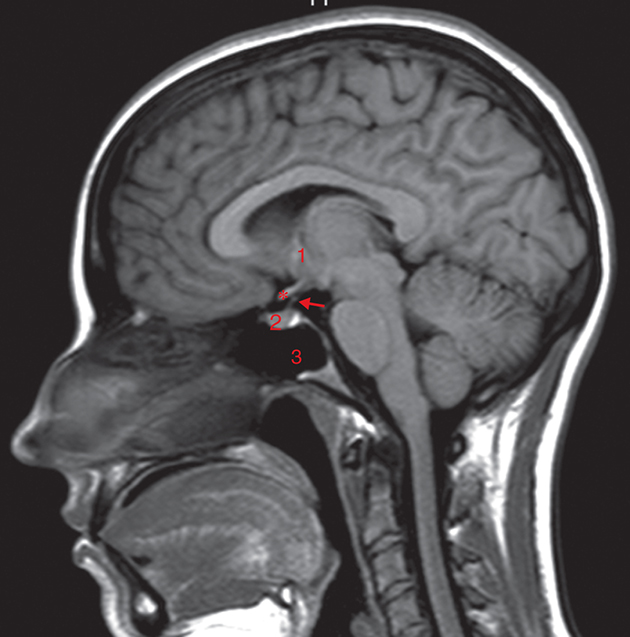
Posterior to the optic chiasm, the visual pathway continues into both the right and the left sides of the brain (the structures on only one side are described here).
Optic Tract
The optic tract is a cylindric, slightly flattened band of fibers approximately 3.5 mm high and 5.1 mm long that runs from the posterolateral corner of the optic chiasm to the LGN.7 Most of the fibers (which are still the axons of retinal ganglion cells) terminate in the LGN. Fibers from the retinal ganglion cells may branch so that the same cell sends fibers to various target structures, or alternatively, some retinal ganglion cell axons may be destined for a specific structure. The afferent fibers of the pupillomotor reflex leave the optic tract before reaching the LGN and pass by way of the superior brachium to the pretectal nucleus in the midbrain. Other fibers project to the suprachiasmatic nucleus in the hypothalamus and to the superior colliculus.
The optic tract lies along the upper anterior and then the lateral surface of the cerebral peduncle and runs parallel to the posterior cerebral artery. The globus pallidus is above, the internal capsule is medial, and the hippocampus is below the optic tract.
Lateral Geniculate Nucleus
Information from all the sensory systems except the olfactory system passes through the thalamus before being transferred to the cerebral cortex. Visual information is processed in the LGN, located on the dorsolateral aspect of the thalamus, before being relayed to higher cortical centers. The LGN resembles an asymmetric cone, the rounded apex of which is oriented laterally. The retinal axons terminate here. Most of the fibers that leave the LGN project to the visual cortex.
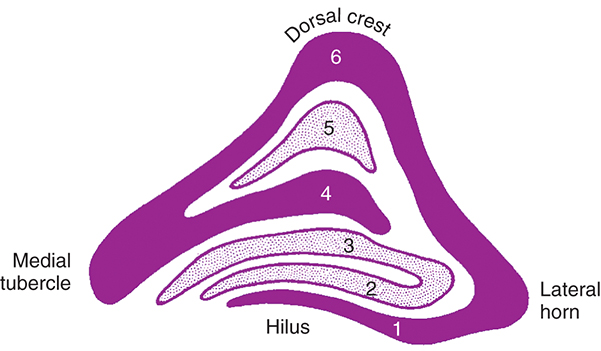
The LGN is a layered structure. The layers are piled on each other, with the larger ones draping over smaller ones, and some layers becoming fragmented and irregular. The cells within a layer are all of the same type, and three types have been identified according to size. Magnocellular layers contain large cells, parvocellular layers contain medium-sized cells, and koniocellular layers contain small cells. The number of layers present depends on the location of the plane through the structure. In the classic textbook presentation of the LGN, six layers are seen. Two magnocellular layers are located inferiorly and numbered 1 and 2, and the four parvocellular layers above them are numbered 3, 4, 5, and 6 (Fig. 15.6). Below each of these six layers lies a koniocellular layer which receives information from short wavelength cones (Fig. 15.7).26 Using functional magnetic resonance imaging (fMRI) on humans, magnocellular layers were found more ventral and medial. Parvocellular layers were found dorsally and laterally.27
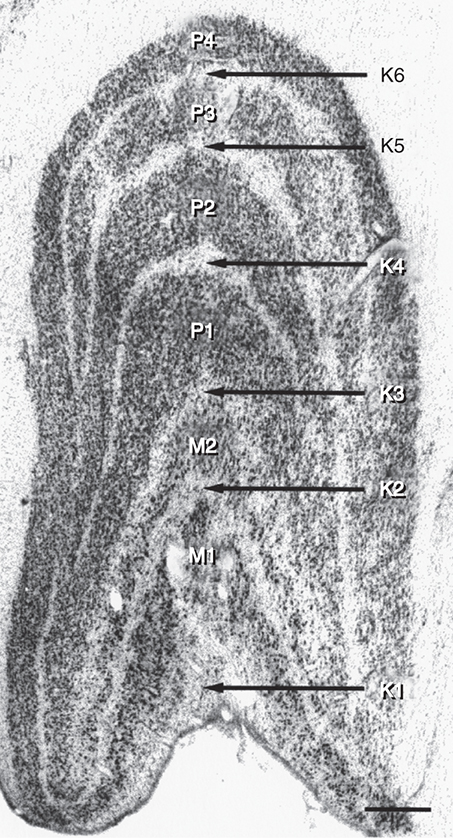
The LGN is not a simple relay station. It also receives input from cortical and subcortical centers and reciprocal innervation from the visual cortex, becoming a center of complex processing. It regulates the flow of visual information, ensuring that the most important information is sent to the cortex.28
The optic tract enters the LGN anteriorly. The internal capsule is lateral, the medial geniculate nucleus is medial, and the inferior horn of the lateral ventricle is posterolateral to the LGN. The axons leave the LGN as the optic radiations.
Optic Radiations (Geniculocalcarine Tract)
The optic radiations spread out fanwise as they leave the LGN, deep in the white matter of the cerebral hemisphere. The anterior bundle sweeps anteriorly and laterally around the anterior tip of the temporal horn of the lateral ventricle before turning posteriorly (Meyer loop) to travel in the temporal lobe en route to the occipital lobe (Fig. 15.8).29,30 The middle bundle travels superior to the temporal horn of the lateral ventricle. The posterior bundle travels within the parietal lobe lateral to the occipital horn of the lateral ventricle before terminating in the striate cortex. The majority of optic radiation fibers terminate in the primary visual cortex; however, there are some fibers that have direct connections with the extrastriate cortex areas.29,31
Visual Cortex
The primary visual cortex (striate cortex or V1), is located almost entirely on the medial surface of the occipital lobe. Just a small portion (perhaps 1 cm long) extends around the posterior pole onto the lateral surface. The visual cortex also is called the striate cortex because a white myelinated fiber layer, the white stria of Gennari, is characteristic of this area. The calcarine fissure extends from the parietooccipital sulcus to the posterior pole, dividing the visual cortex into an upper portion (the cuneus gyrus) and a lower part (the lingual gyrus) (Fig. 15.9). Most of the primary visual cortex is buried in the tissue within the calcarine fissure.
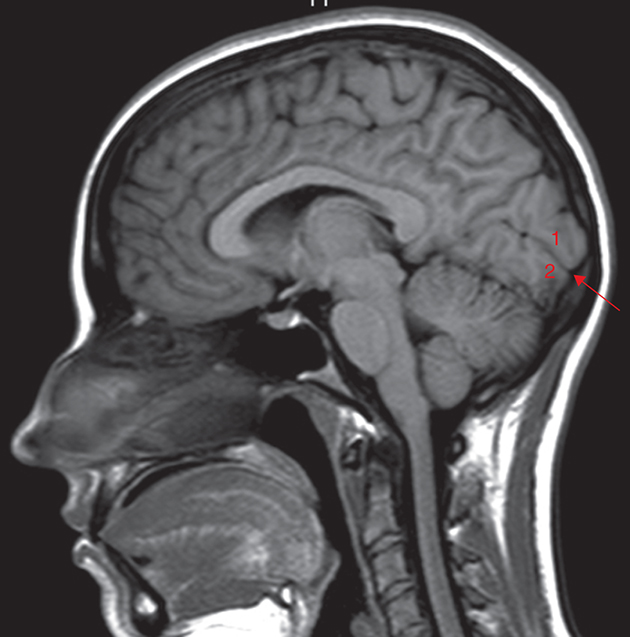
The primary visual cortex has a thickness of about 2 mm and is organized into horizontal layers and vertical columns. Layer I, the most superficial layer, contains a few scattered neurons. Layer II contains neurons that send axons only to deeper cortical layers. Layer III contains neurons that communicate with both near and far cortical locations. Layer IV contains the stria of Gennari and is subdivided into strata, one of which receives information from the magnocellular layers of the LGN and another receives information from the parvocellular layers of the LGN. Layer IV sends axons to more superficial areas of the primary visual cortex, as well as other visual cortical areas. Layer V sends axons to the superior colliculus and other areas in the brainstem. Layer VI sends projections back to the LGN.28
Certain cortical regions are active during motion stimulation, whereas others are active during color vision stimulation.32 The magnocellular areas mediate movement detection and low-spatial-frequency contrast sensitivity, and the parvocellular areas mediate color and high-spatial-frequency contrast sensitivity, although this generalization oversimplifies the properties.33–36
Cells are also distributed in a vertical organization, according to the eye of origin, forming alternating parallel ocular dominance columns. These columns are lacking in the area of the cortex that represents the physiologic blind spot because this region receives information exclusively from one eye. A second system of columns, specific for stimulus orientation, responds on the basis of the direction of a light slit or edge.37 Contour analysis and binocular vision are two functions of the visual cortex, and such processing is a function of both its horizontal and its vertical organization. The cells within the striate cortex are activated only by input from the LGN, although other cortical areas have input into the striate cortex.38–40
The striate cortex communicates with the superior colliculus and the frontal eye fields. The superior colliculus, which has a complete retinotopic map of the contralateral field of vision, receives communication from fibers exiting the posterior optic tract, as well as the striate cortex. It does not analyze sensory information for perception but is important for visual orientation, foveation, and the control of saccadic eye movements with input from the frontal eye fields.41,42 The frontal eye fields, in the frontal lobe, receive fibers from the striate cortex that contribute to the control of conjugate eye movements. Both voluntary and reflex ocular movements are mediated in this area, as are pupillary responses to near objects (see Ch. 14).
The striate cortex combines and analyzes the visual information relayed from the LGN and transmits this information to the higher visual association areas (the extrastriate cortex), which provide further interpretation. These areas (V2, V3, V4, and V5) surround the striate cortex and are arranged in a nesting pattern, progressively more lateral and anterior, within the occipital cortex. The visual and visual association areas in one hemisphere are connected to the corresponding areas in the other hemisphere through the posterior portion of the corpus callosum.4
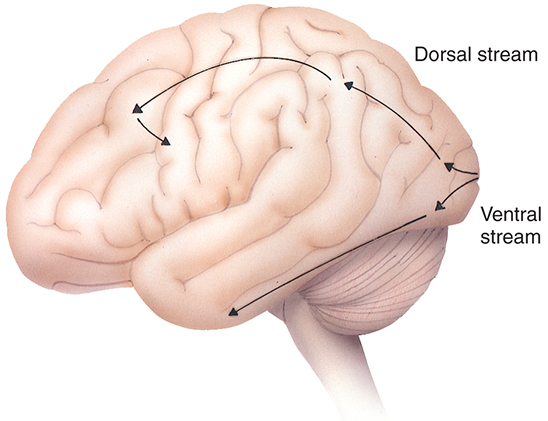
Extrastriate areas allow object recognition even among similar objects and when objects are transformed, such as a change in size, rotation, or illumination. Two pathways, the ventral stream and dorsal stream, are involved in processing information (Fig. 15.10). The ventral stream travels through the occipitotemporal cortex and includes V2, V4, and the inferior temporal cortex. This pathway aids in processing object qualities, such as size, color, and shape.43,44 The dorsal stream courses through the occipitoparietal cortex and involves V5 (the medial temporal area). The dorsal stream processes spatial information and visually guided actions, including position, motion, depth perception, and relationships between objects.43,44
Blood Supply to the Visual Pathway
The structures of the visual pathway have an extensive blood supply. Fig. 15.11 shows many of the involved vessels. The outer retinal layers receive nutrition from the choroid, whereas the inner retina is supplied by the central retinal artery. The circle of Zinn, the anastomotic ring of branches of the short ciliary arteries, and peripapillary vessels supply the intralaminar optic disc (see Fig. 12.3).4,45 Capillaries within the optic nerve are composed of nonfenestrated endothelium joined by zonula occludens, thus the vessels perfusing the nerve head are part of the blood-brain barrier.15,46 Pial vessels supply the optic nerve throughout its length. The intraorbital and canalicular pial vessels are supplied by branches from the ophthalmic artery. The superior hypophysial artery, a branch of the internal carotid artery, is the main blood supply to the intracranial optic nerve.19 Branches of the ophthalmic and anterior cerebral arteries may also contribute to the distal and proximal vascular supplies of the intracranial optic nerve, respectively.
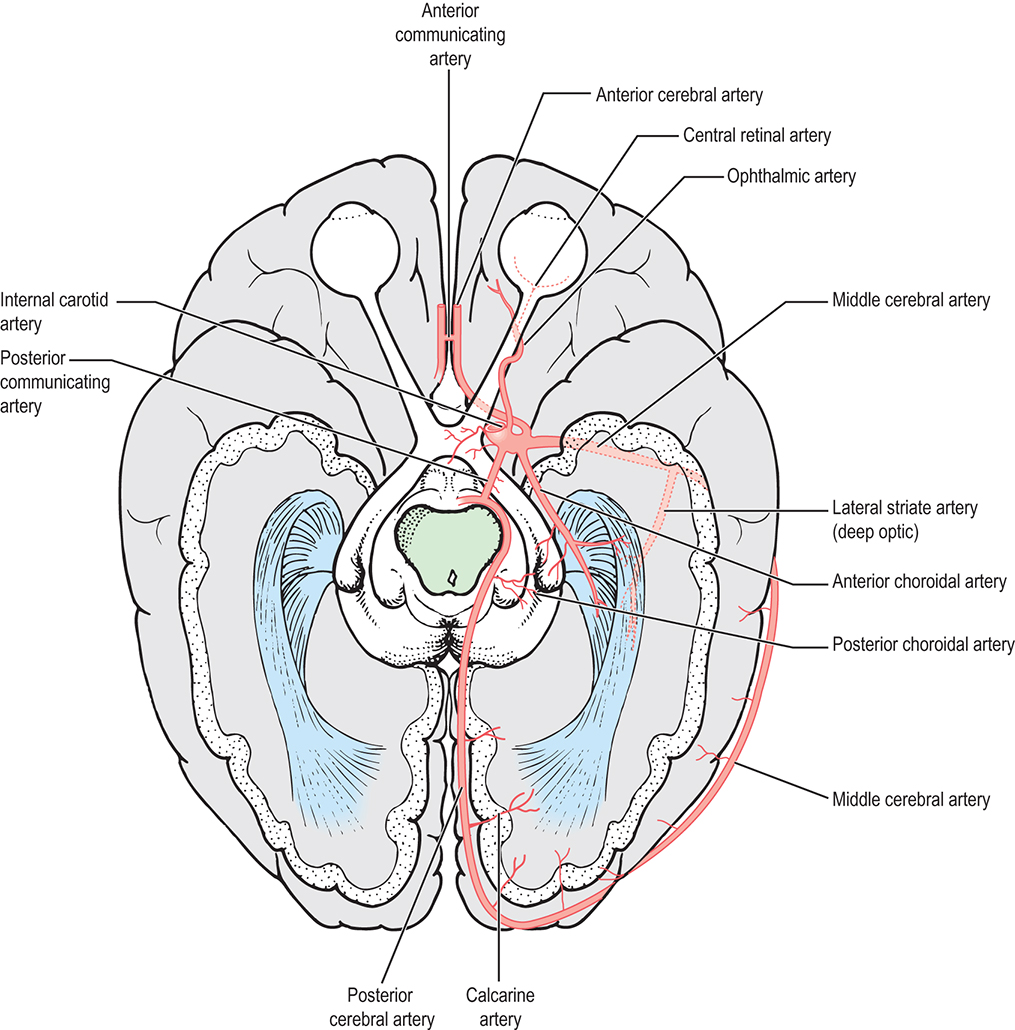
The blood supply to the optic chiasm is rich and anastomotic, with arterioles from the circle of Willis forming capillary beds at two levels.47,48 The superior network is supplied by the anterior cerebral, anterior communicating, posterior communicating, and superior hypophyseal arteries, whereas the inferior network is supplied by the superior hypophyseal and posterior communicating arteries.49 The anterior choroidal artery, a branch of the internal carotid, is a primary supplier of the optic tract, although small branches from the middle cerebral artery also contribute.4,6,25 The blood supply to the LGN is derived from the anterior choroidal artery and the lateral choroidal and posterior choroidal branches of the posterior cerebral artery.6,50
The anterior optic radiations are supplied by the anterior choroidal artery and the middle cerebral artery. The middle group of fibers is supplied by the lateral striate (deep optic) branch of the middle cerebral artery. Branches of the posterior cerebral artery, including the calcarine branch, supply the posterior radiations. Branches from the middle cerebral artery also contribute. The calcarine branch of the posterior cerebral artery is the major blood supply for the striate cortex, often supplemented by the posterior temporal or parietooccipital branch of the posterior cerebral artery. The occipital pole, corresponding to the central visual field, may have a dual blood supply as the temperooccipital branch of the middle cerebral artery anastomoses with branches of the posterior cerebral artery.
Fiber Orientation and Visual Fields
With the eye looking straight ahead and fixating on an object, one is able to detect other objects around the point of regard, although the details may not be discernible. This entire visible area is termed the visual field. Information from the visual field is taken in by the retina and processed through the afferent visual sensory pathway. The location and orderly arrangement of the fibers throughout this pathway have been extensively studied. Damage in this afferent visual pathway will cause a defect in the visual field. Knowledge of the fiber patterns in the pathway can help to identify the location of a lesion on the basis of the resultant visual field defect.
Retina
The axons of the retinal ganglion cells form characteristic patterns in the nerve fiber layer. The group of fibers that course from the macular area to the optic disc is called the papillomacular bundle (Fig. 15.12). The superior and inferior temporal fibers, separated by a horizontal line extending through the center of the fovea called the horizontal retinal raphe, must arch superiorly and inferiorly around the macular area, forming characteristic arcuate patterns in their course to the optic disc. The temporal retinal vessels usually do not cross the horizontal raphe either. The nasal fibers can travel directly to the optic disc and are described as radiating. Nasal and temporal fibers are separated by a theoretic vertical line passing through the center of the fovea. The long nerve fibers, from the peripheral retina, are more vitread in location than are the short peripapillary fibers, with extensive intermingling in the prelaminar optic nerve.51
Optic Disc
All of the axons in the nerve fiber layer come together at the optic disc, creating a specific pattern. The nasal fibers radiate directly to the nasal side of the disc, whereas the papillomacular bundle courses directly to the temporal side of the disc (see Fig. 15.12). The fibers from the superior temporal retina arch around the papillomacular bundle to enter the superior pole of the disc. Fibers from the inferior temporal retina curve below the papillomacular bundle to the inferior pole. The macular fibers take up approximately one-third of the disc, although the macular area encompasses only one-twentieth of the retinal area. The temporal fibers occupy approximately one-third of the disc, as do the nasal fibers (Fig. 15.13A). The boundaries between each set of fibers are not always clear-cut in all parts of the pathway. The fibers from the peripheral retina are more superficial than those coming from the central retina.52
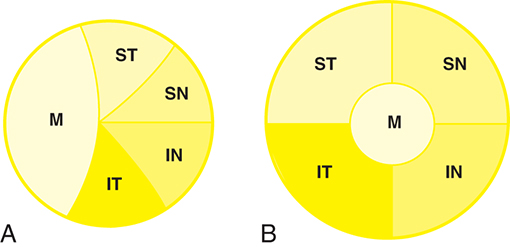
Optic Nerve
Near the lamina cribrosa, the fibers have the same orientation as they do at the disc, but within a short distance the macular fibers move to the center of the nerve. The rest of the fibers take up their logical positions: superior temporal fibers in the superior temporal optic nerve, inferior temporal fibers in the inferior temporal nerve, superior nasal fibers in the superior nasal nerve, and inferior nasal fibers in the inferior nasal optic nerve (Fig. 15.13B).
Optic Chiasm
In the optic chiasm, the nasal fibers cross (decussate). The ratio of crossed to uncrossed fibers in the chiasm is approximately 53 to 47.53 The crossing pattern depends on processes that occur during embryological development, with certain molecular guides directing the path taken by nerve fibers. The majority of the nasal fibers cross in the paracentral rather than the central chiasm.54 The inferior fibers cross more anteriorly and then travel back through the chiasm into the contralateral optic tract (Fig. 15.14).54 Traditionally, it was thought that the inferior nasal fibers looped 1 to 2 mm forward into the terminal part of the opposite optic nerve before turning to run back through the chiasm. The existence of these anterior loops (anterior knees of Wilbrand) is controversial. Some studies show that the anterior knees of Wilbrand are artifacts caused by the prior enucleation in those studies cited by Wilbrand.55 In addition, some studies found no junctional scotoma in the visual field after surgical sectioning of the anterior chiasmal junction.55,56 Other clinical findings in patients demonstrating a junctional scotoma support the existence of Wilbrand knees;57–59 however, in some of the cases, the association has been questioned.60 The fibers which cross in the inferior anterior chiasm make a large, almost 90-degree crossing angle, and it is thought that it is this sharp angle that makes them more suspectable to compression.54 It is also possible that a compressive lesion could affect the inferior fibers as they travel anteriorly in the chiasm despite them not traveling into the contralateral optic nerve.57 Furthering the controversy, a recent study using the anisotropic light reflecting properties of myelinated axons revealed inferior, but not superior, fibers arching toward the contralateral optic nerve before reversing direction.61
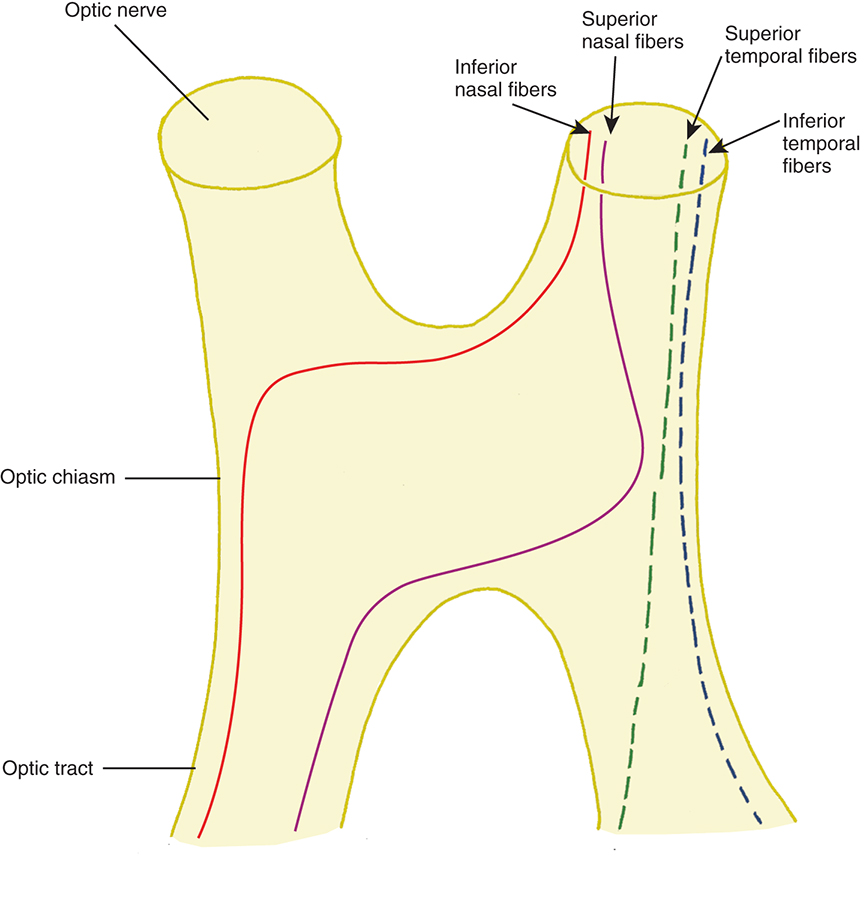
The superior nasal fibers enter the superior chiasm, where they cross in the more posterior chiasm and then leave the chiasm in the contralateral optic tract. These fibers make shallower crossing angles than do the inferior crossing fibers.54 Fibers from the temporal retina course directly back through the chiasm into the optic tract. Temporal fibers are located laterally in the chiasm, whereas nasal fibers, even after crossing are more centrally located.54 Nasal macular fibers also cross and are spread throughout most of the chiasm.
A small number of fibers have been identified that exit the posterior of the chiasm and enter the suprachiasmatic nucleus in the hypothalamus. These fibers have a role in synchronization of circadian rhythm.62–64
Optic Tract
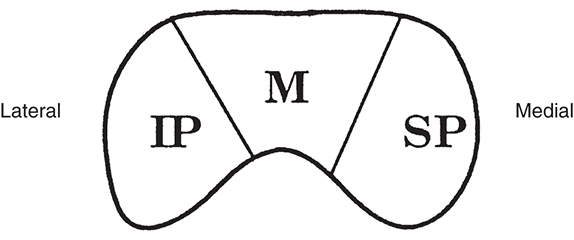
As the fibers leave the chiasm in the optic tract, the crossed and uncrossed fibers intermingle. The superior fibers (the fibers from both the ipsilateral superior temporal retina and the contralateral superior nasal retina) move to the medial side of the tract. Fibers from the inferior retina (ipsilateral inferior temporal retinal fibers and contralateral inferior nasal retinal fibers) occupy the lateral area of the tract (Fig. 15.15). The macular fibers, crossed and uncrossed, are located between these two groups.
Lateral Geniculate Nucleus
Fibers from the superior retinal quadrants terminate in the medial aspect of the LGN, whereas fibers from the inferior retinal quadrants terminate in the lateral aspect. A dorsal wedge, composing two-thirds to three-fourths of the LGN, represents the macula.65,66 Each of the magnocellular and parvocellular layers receives input from just one eye: layers 1, 4, and 6 receive fibers from the contralateral nasal retina, whereas layers 2, 3, and 5 receive ipsilateral temporal retinal fibers (Fig. 15.16).53 Most of the structure, including the wedge representing the macula, contains all layers, although in the far medial and lateral aspects, some of the layers merge.4,65
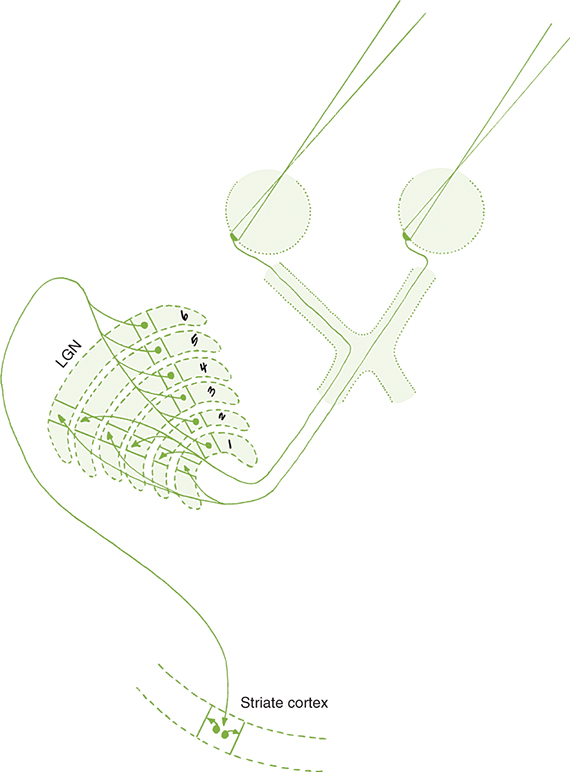
The anatomic structure of the human LGN is similar to that of the monkey, so detailed maps of the monkey LGN have been applied to the human structure.67 Each layer of the LGN contains a retinotopic map or representation of the contralateral hemifield of vision. A retinotopic map is a point-to-point localization of the retina. These maps are stacked on one another, such that if a line (called a line of projection) were passed through all six layers, perpendicular to the surface, the intercepted cells all would be carrying information about the same point in the visual field. This alignment is so precise that there is a gap in each contralateral layer along the line of projection that corresponds to the location of the optic disc.68 Thus the fibers that carry information from the same site in the visual field of each eye terminate in adjacent layers of the LGN, right next to one another (see Fig. 15.16). The fibers course through the posterior limb of the internal capsule as they leave the LGN to form the optic radiations.
Optic Radiations
The fibers leaving the lateral aspect of the LGN, representing inferior retina, follow an indirect route to the occipital lobe. They pass into the temporal lobe and loop around the tip of the temporal horn of the lateral ventricle, forming Meyer loops; these fibers form the inferior radiations (Fig. 15.17). Fibers from the medial aspect of the LGN, representing superior retina, lie superiorly as they pass through the parietal lobe. The fibers from the macula are generally situated between the superior and inferior fibers.
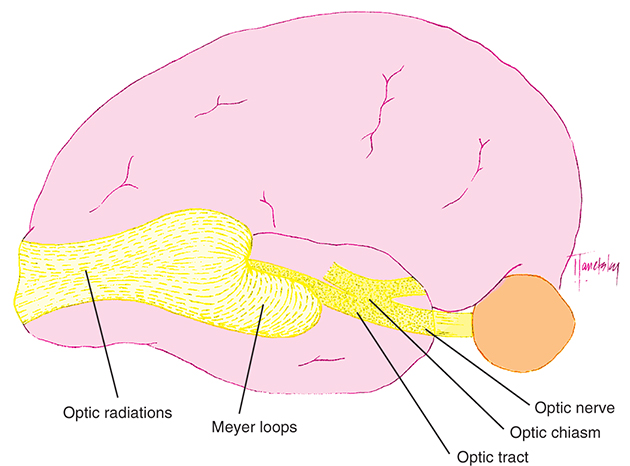
Striate Cortex
The superior radiations terminate in the area of the striate cortex above the calcarine fissure, called the cuneus gyrus. The inferior radiations terminate in the region below the calcarine fissure—the lingual gyrus. Thus the cuneus gyrus receives projections from the superior retina and the lingual gyrus from the inferior retina. Only one-third of the striate cortex is on the surface of the occipital lobe. The majority is buried within the calcarine fissure, and only a small portion is on the posterolateral aspect of the occipital posterior pole.
Fibers from the macular area terminate in the most posterior part of the striate cortex, with the superior macular area represented in the cuneus gyrus and the inferior macula represented in the lingual gyrus. The macular projection might extend onto the posterolateral surface of the occipital cortex. The macular area representation occupies a relatively large portion of striate cortex compared with the small macular area in the retina. The macular cells are densely packed, and macular fibers are small caliber. Because macular function involves sharp, detailed vision, the macular representation in the striate cortex is more extensive than the representation of peripheral retinal areas. The most anterior part of the striate cortex, the part adjacent to the parietal lobe, represents the periphery of the nasal retina, corresponding to an area of visual field, the temporal crescent, that is seen by the contralateral eye only.
Retinotopic representation is present in the striate cortex. Those fibers that are adjacent to one another in the layers of the LGN project to the same area in the visual cortex (see Fig. 15.16). That is, corresponding points from the two retinas (ipsilateral temporal and contralateral nasal) that represent the same target in the visual field will project to neighboring locations in the primary visual cortex. All the cells in a column correspond to a stimulus presented at the same point in the visual field, and cells in an adjacent column correspond to an adjacent point in the visual field.
Clinical Comment: Visual Field Testing
The visual field is tested monocularly, with the patient looking straight ahead at a fixation point and responding when a target is seen anywhere in the area surrounding that fixation point. The field can be divided into four quadrants by a vertical line and a horizontal line that intersect at the point of fixation. The point of fixation is seen by the fovea. The temporal field is slightly larger than the nasal field.
Inversion and reversal of the field are caused by the optical system of the eye. The superior field is imaged on the inferior retina and the inferior field on the superior retina. The nasal field is imaged on the temporal retina and the temporal field on the nasal retina (Fig. 15.18). This orientation is maintained in the cortex, where the superior field is projected onto the visual cortex inferior to the calcarine fissure and where the inferior visual field is projected onto the cortex superior to the calcarine fissure.
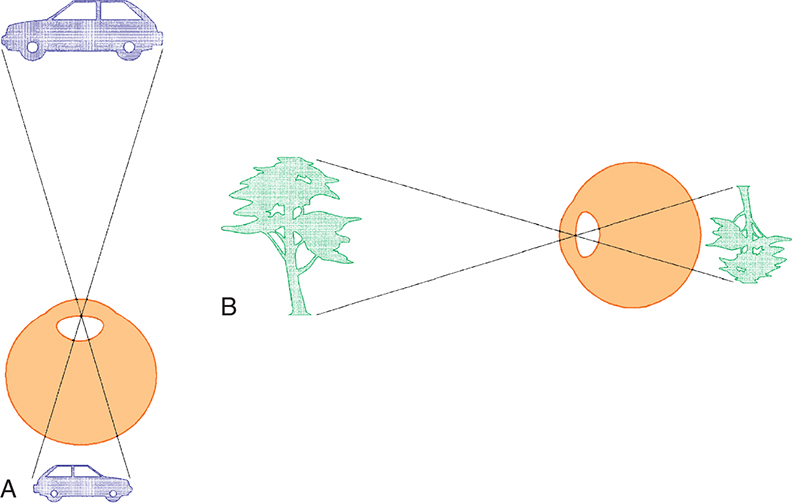
The reader is cautioned to be aware of the difference between visual fibers and visual fields. Both can be described as nasal, temporal, superior, and inferior.
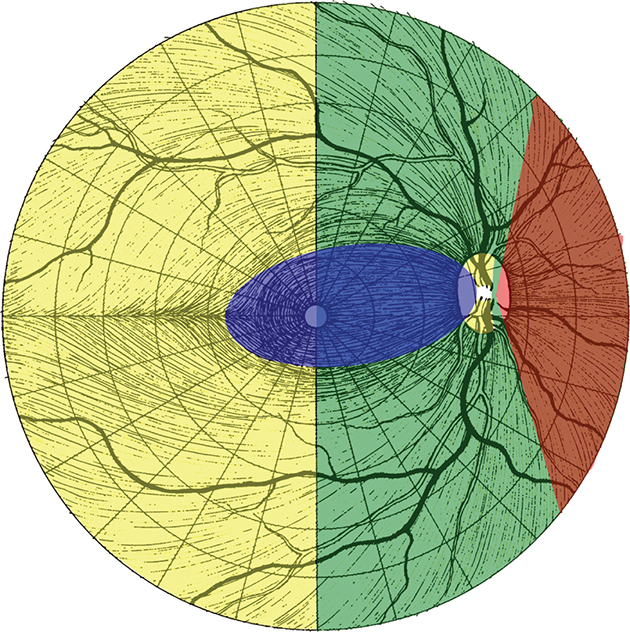
The visual field seen by the right eye is nearly the same as that seen by the left eye. The nasal part of the field for one eye is the same as the temporal part of the field seen by the other eye, with the exception of the far temporal peripheral field, which is called the temporal crescent. The temporal crescent is imaged on the nasal retina of one eye but not on the temporal retina of the other because the depth of the orbit and the prominence of the nose blocks the periphery of the field from imaging on the temporal retina. Within each temporal field is an absolute scotoma, the physiologic blind spot, a result of the lack of photoreceptors on the optic disc (Fig. 15.19).
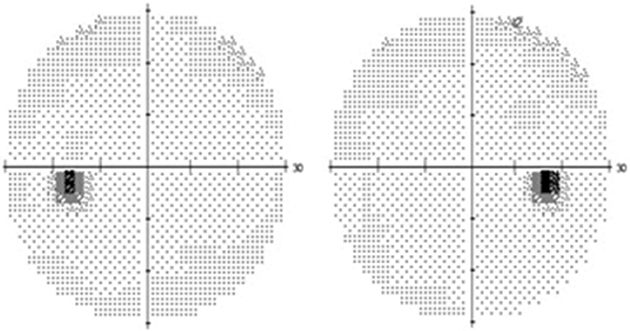
Because the fibers that emanate from the nasal retina cross in the chiasm, the postchiasmal pathway carries information from the contralateral temporal field and the ipsilateral nasal field. These combined areas can be described as the contralateral hemifield (i.e., the right postchiasmal pathway carries information from the left side of the visual field for both eyes). Thus the left side of the field is “seen” by the right striate cortex, paralleling the involvement of the right hemisphere in the motor and sensory activities of the left side of the body. Similarly, objects in the right side of the field are “seen” by the left striate cortex (see Fig. 15.1). A defect that affects the nasal field of one eye and the temporal field of the other eye is described as homonymous.
Note that reference to the left side of the visual field is not the same as the visual field of the left eye. Clinicians will refer to the right visual field (meaning the right side of the field) and the left visual field (meaning the left side of the field).
A visual field defect of just one eye must be caused by a disruption anterior to the chiasm. If there is a defect in the fields of both eyes, there are two lesions, one in each prechiasmal pathway, or there is a single lesion in the chiasm or the postchiasmal pathway, where the fibers for the two eyes are brought together. The pattern of the defect, as well as associated signs or symptoms, might aid in determining the location of the damage.
Clinical Comment: Characteristic Visual Field Defects
Fig. 15.20 depicts examples of various visual field defects.

The regular fiber orientation in each structure of the visual pathway can be correlated with a specific pattern of visual field loss. A lesion of the choroid or outer retina will cause a field defect that is similar in shape to the lesion and is in the corresponding location in the field (e.g., if the lesion is in the inferior temporal retina, the defect will be in the superior nasal field). These lesions can cross the horizontal or vertical midlines (Fig. 15.21).
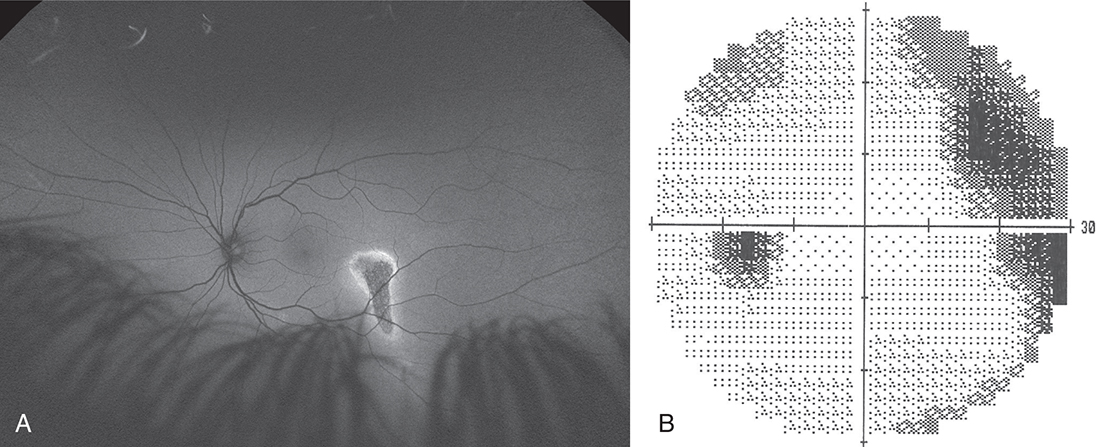
A lesion in the nerve fiber layer will cause a field defect corresponding to the location and configuration of the affected nerve fiber bundle. One of the disease processes that affects the nerve fiber layer is glaucoma. It first affects the temporal nerve fibers as they exit the globe at the superior and inferior optic disc. If temporal retinal fibers are affected, an arcuate defect can be produced that curves around the point of fixation, starting at the blind spot and terminating at the horizontal nasal meridian (Fig. 15.22). This abrupt edge at the horizontal meridian is called a nasal step and results from the configuration of the fibers at the temporal retinal raphe. Less often, a lesion affects a nasal bundle of nerves, producing a wedge-shaped defect emanating from the physiologic blind spot into the temporal field.
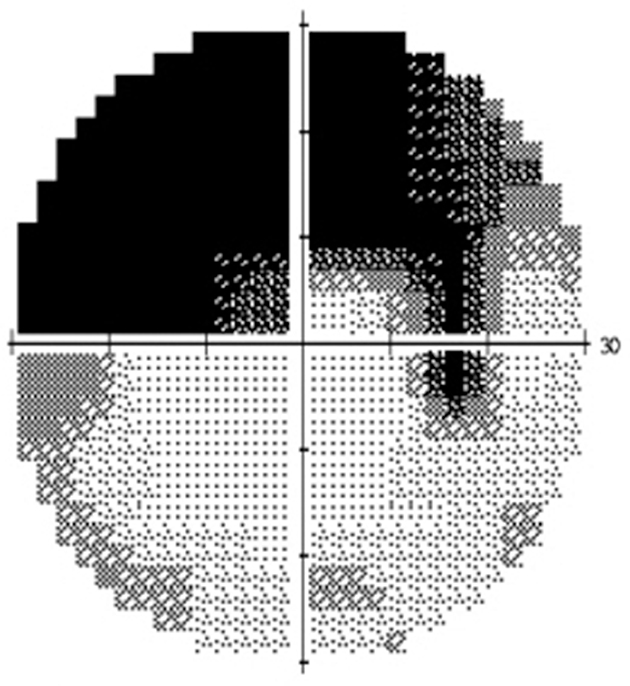
Injury to the optic nerve is accompanied by a visual field defect, a relative afferent pupillary defect, and atrophy of the affected nerve fibers, which eventually is manifested at the optic disc.
The optic chiasm brings all the visual fibers together. Lesions of the chiasm usually will show bitemporal defects. The most common cause of a bitemporal field defect is a pituitary gland tumor, and a visual field defect is often the first clinical sign (Fig. 15.23). The crossed fibers are generally damaged first in compressive lesions, such as a tumor, because of the large crossing angle of the fibers.54 This susceptibility to damage might also be attributable to the purported weak blood supply of the medial portion of the chiasm. Consequently, the crossed fibers also are more susceptible to ischemia in a vascular event.48
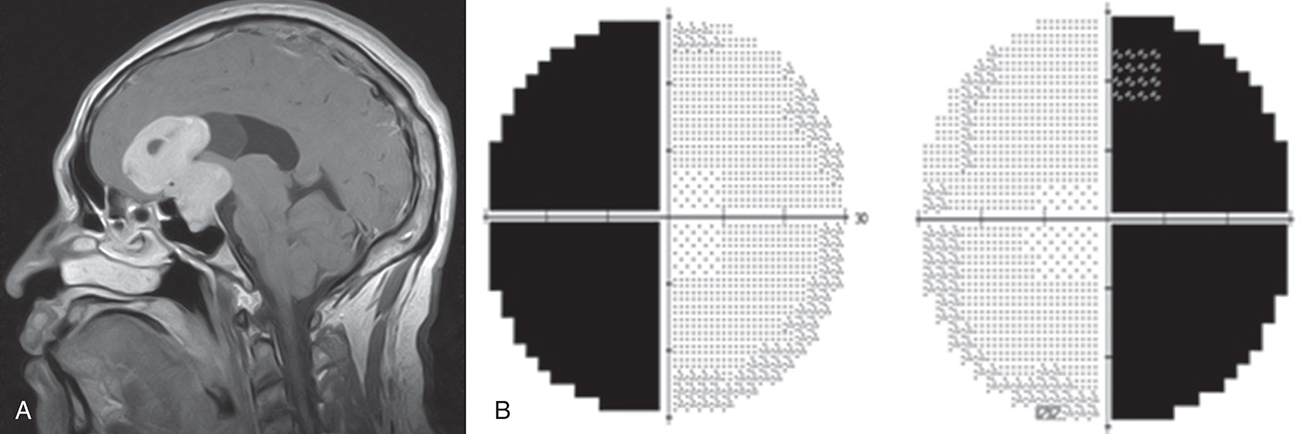
A single lesion at the optic chiasm and its junction with the optic nerve might be characterized by an overall depression in the field of the eye on the same side as the lesion, as well as a superior temporal defect in the field of the opposite eye. This is known as a junctional scotoma. It may occur because the inferior nasal fibers are located anteriorly after crossing in the chiasm and because the nasal fibers have a large crossing angle relative to fibers entering the chiasm from the ipsilateral optic nerve.
A homonymous field defect will be produced by a single lesion in the postchiasmal pathway, as the nasal fibers of the contralateral eye join the temporal fibers of the ipsilateral eye. Visual acuity usually is not affected because one-half the fovea is sufficient for 20/20 Snellen acuity. In a postchiasmal lesion, the field loss is present on the side of the field contralateral to the lesion. Other signs or symptoms accompanying a homonymous defect can help the diagnostician determine more exactly the site of the lesion.
A lesion involving the optic tract eventually will produce optic nerve atrophy, which usually becomes evident as optic disc pallor. Because the optic tract is relatively small in cross section, a lesion often damages all of the fibers, causing a homonymous field defect that affects the entire half of the field. If a partial hemianopia results, the defects will often be incongruent. Defects in a homonymous field are congruent if the two defects are similarly shaped and are incongruent if the defect shapes are dissimilar (Figs. 15.24 and 15.25). Because crossed fibers outnumber uncrossed fibers, a lesion of the complete optic tract may be accompanied by a relative afferent pupillary defect of the contralateral eye.

A lesion in the LGN will affect the contralateral field and eventually cause optic atrophy. Because of the point-to-point localization in the LGN, lesions here produce moderate to complete congruent field defects.69
Damage to the optic radiations or cortex does not normally cause atrophy of the optic nerve or a pupillary defect because it does not involve the fibers of the retinal ganglion cells. A lesion of the optic radiations causes a contralateral homonymous field defect. Because the fibers are so spread out, the defect is generally incongruent and may affect only one quadrant. If a lesion of the temporal lobe involves the Meyer loop, a superior field defect will result. Parietal lobe lesions more commonly cause inferior field defects (see Fig. 15.24).41
The characteristic feature of a defect in the occipital lobe is congruency. Congruency depends on how closely fibers from corresponding points of each eye (carrying the same visual field information) are positioned to one another at the site of the lesion. As the fibers reach the occipital lobe and finally the striate cortex, fibers emanating from corresponding points in the field come together to form a point-to-point representation of the field. Therefore a lesion here will cause a congruent defect. Injury to the lingual gyrus will cause a superior visual field defect. Involvement of the cuneus gyrus will cause an inferior visual field defect (see Fig. 15.25). Lesions more anterior in the occipital lobe will affect more peripheral visual field, whereas lesions more posterior will affect macular fibers.
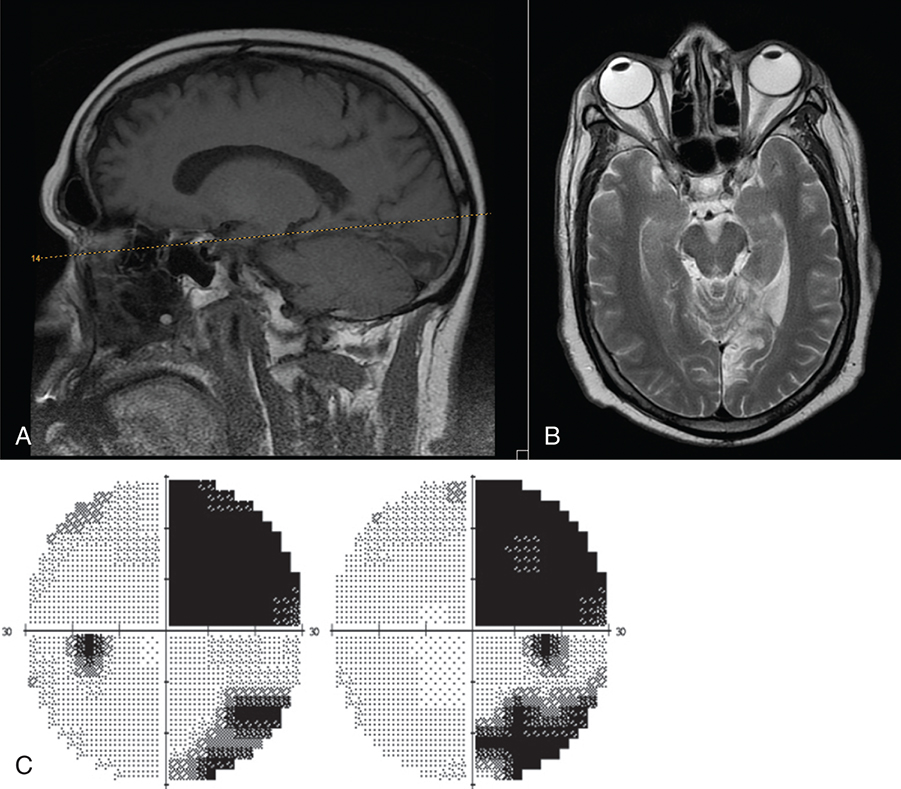
When visual association areas within the occipital, temporal, or parietal lobes are involved, higher cortical visual processes may be affected. Lesions of the parietal lobe can cause agnosia (inability to recognize objects), apraxia (inability to carry out movements), or aphasia (difficulty with speech). Temporal lobe lesions can cause memory impairment, seizures, or aphasia. Injury involving the occipitotemporal cortex can affect object and facial recognition.70 Blindsight occurs when there seems to be some sight in a hemifield but there is no conscious awareness of the sight. That is, a motor reflex response can be elicited with the presentation of an unexpected stimulus in the affected field, but the patient has no awareness of the vision. Connections between the LGN and the human motion area in the extrastriate middle temporal cortex (V5), which bypass the visual cortex, are thought to be involved in blindsight.71,72
Striate Cortex Maps
An early study correlating the visual field to the striate cortex was done by Holmes and Lister.73 They studied injured soldiers from World War I and attempted to match visual field defects with injuries from shrapnel to the occipital lobe. The Holmes map provided a detailed source showing the representation of the visual field in human striate cortex. The macular portion extended from the posterior pole forward, with the periphery of the field represented in the anterior occipital lobe and the uniocular temporal crescent in the most anterior aspect of the striate cortex adjacent to the parietooccipital sulcus. Detailed mapping of a monkey striate cortex using electrophysiological methods revealed discrepancies between monkey and human data. These findings suggested that either the monkey cortex and human cortex are not as alike as believed or that the Holmes map required some modification.
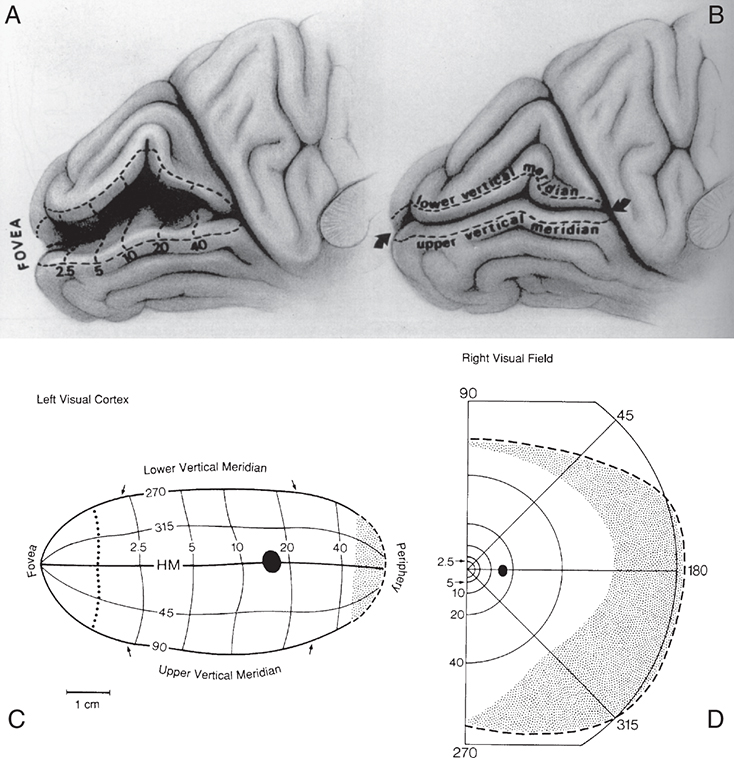
Technologies, such as MRI, have been used to study the human cortex, allowing more direct correlation of a lesion with a field defect. Some investigators suggest revision of the Holmes map.74 The primary change concerns the extent of the area depicting macular representation. A much greater area of the visual cortex is thought to be taken up by macular projection, with the central 30 degrees of the visual field represented in approximately 83% of the striate cortex (Fig. 15.26).74 Other imaging studies more closely agree with the Holmes map and show that the central 15 degrees of vision occupies 37% of the surface area of the striate cortex.75 Some discrepancies may result from the nature of the lesion because an MRI may overestimate the actual area involved when edema is present.75
Macular Sparing
Macular sparing occurs when an area of central vision remains within a homonymous field defect. Because fixational eye movements of 1 to 2 degrees do occur during the visual field examination, the area spared within the defect should involve at least 3 degrees for macular sparing to be confirmed clinically. Even in the presence of an extensive lesion, some of the macular projection area might remain unaffected, either because the posterior pole of the occipital lobe has such an extensive blood supply or because the macular projection covers a very large area. Macular sparing can also be explained by the size and overlap of the receptive field of the retinal ganglion cells.76
Aging Within the Visual Pathway
Neural cell death occurs throughout all structures of the visual pathway, although the extent varies significantly within the population.77,78 Age is accompanied by a decrease in the extent of the visual field, caused both by loss of cells and by a decrease in the transparency of the ocular media.25,79 The ability to perceive accurately the speed of moving objects declines with age, and animal studies have identified an age-related difference in temporal processing speed at the level of the visual cortex.80 This decline in accurately perceiving the speed of moving objects may contribute to the higher incidence of automobile accidents among the elderly population.
References
1. Hoyt WF, Osman L.Visual fiber anatomy in the infrageniculate pathway of the primate. Arch Ophthalmol. 1962;68:124.
2. Polyak S.The Vertebrate Visual System. Chicago: University of Chicago Press; 1957.
3. Jonas JB, Schmidt AM, Muller-Bergh JA, et al. Human optic nerve fiber count and optic disc size. Invest Ophthalmol Vis Sci. 2012;33(6):1992.
4. Sadun AA, Glaser JS.Anatomy of the visual sensory system. In: Tasman W, Jaeger EA, editors. Duane's Foundations of Clinical Ophthalmology, vol 1. Philadelphia: Lippincott; 1994.
5. Mikelberg FS, Drance SM, Schulzer M, et al. The normal human optic nerve. Axon count and axon diameter distribution. Ophthalmology. 1989;96:1325.
6. Warwick R.Visual pathway. In: Eugene Wolff's Anatomy of the Eye and Orbit, ed 7. Philadelphia: Saunders; 1976: p. 325-395.
7. Parravano JG, Toledo A, Kucharczyk W.Dimensions of the optic nerves, chiasm, and tracts: MR quantitative comparison between patients with optic atrophy and normal. J Comput Assist Tomograph. 1993;17(5):688.
8. Radunović M, Vitosević Z, Cetković M, et al. Morphometric analysis of the fascicular organisation of the optic nerve. Vojnosanitetski Pregled. 2015;72(2):132-135.
9. Kim, M, Kim, T-W, Weinreb, RN, et al. Differentiation of parapapillary atrophy using spectral-domain optical coherence tomography. Ophthalmology. 2013;120(9):1790-1797.
10. Reis ASC, Sharpe GP, Yang H, et al. Optic disc margin anatomy in patients with glaucoma and normal controls with spectral domain optical coherence tomography. Ophthalmology. 2012;119(4):738-747.
11. Sanfilippo PG, Huynh E, Yazar S, et al. Spectral-domain optical coherence tomography-derived characteristics of Bruch membrane opening in a young adult Australian population. Am J Ophthalmol. 2016;165:154-163.
12. Han JC, Choi JH, Park DY, et al. Border tissue morphology is spatially associated with focal lamina cribrosa defect and deep-layer microvasculature dropout in open-angle glaucoma. Am J Ophthalmol.2019;203:89-102.
13. Jonas JB, Holbach L, Panda-Jonas S.Peripapillary ring: histology and correlations, Acta Ophthalmol. 2014;92(4):e273-279.
14. Okinami S, Ohkuma M, Tsukahara I.Kuhnt intermediary tissue as a barrier between the optic nerve and retina. Graefe Arch Clin Exp Ophthalmol. 1976;201(1):57-67.
15. Levin LA.Optic nerve. In: Kaufman PL, Alm A, editors. Adler's Physiology of the Eye, ed 10. St Louis: Elsevier; 2003: p. 603.
16. Hayreh SS.Pathogenesis of optic disc edema in raised intracranial pressure. Prog Retinal Eye Res. 2016;50:108-144.
17. Jonas JB, Xu L.Histological changes of high axial myopia. Eye (London, England). 2014;28(2):113-117.
18. Jonas JB, Wang N, Yang D, et al. Facts and myths of cerebrospinal fluid pressure for the physiology of the eye. Prog Retinal Eye Res. 2015;46:67-83.
19. Caporlingua A, Prior A, Cavagnaro MJ, et al. The intracranial and intracanalicular optic nerve as seen through different surgical windows: endoscopic versus transcranial. World Neurosurg. 2019;124:522-538.
20. Liugan M, Xu Z, Zhang M.Reduced free communication of the subarachnoid space within the optic canal in the human. Am J Ophthalmol. 2017;179:25-31.
21. Kim DH, Jun J-S, Kim R.Ultrasonographic measurement of the optic nerve sheath diameter and its association with eyeball transverse diameter in 585 healthy volunteers. Sci Rep. 2017;7(1):15906.
22. Kuang T-M, Liu CJ-L, Ko Y-C, et al. Distribution and associated factors of optic disc diameter and cup-to-disc ratio in an elderly Chinese population. J Chin Med Assoc. 2014;77(4), 203-208.
23. Shofty B, Ben-Sira L, Constantini S, et al. Optic nerve sheath diameter on MR imaging: Establishment of norms and comparison of pediatric patients with idiopathic intracranial hypertension with healthy controls. AJNR. 2012;(2):366-369.
24. Siam AL H, El-Mamoun TA, Ali MH. A restudy of the surgical anatomy of the posterior aspect of the globe: an essential topography for exact macular buckling. Retina (Philadelphia, Pa). 2011;31(7):1405-1411.
25. Harrington DO.The Visual Fields, ed 5. St Louis: Mosby; 1981.
26. D'Souza DV, Auer T, Strasburger H, et al. Temporal frequency and chromatic processing in humans: An fMRI study of the cortical visual areas. J Vision. 2011;11(8).
27. Denison RN, Vu AT, Yacoub E, et al. Functional mapping of the magnocellular and parvocellular subdivisions of human LGN. NeuroImage. 2014;102 Pt2:358-369.
28. Casagrande VA, Ichida JM.The primary visual cortex. In: Kaufman PL, Alm A, editors. Adler's Physiology of the Eye ed 10. St Louis: Elsevier; 2003: p. 669.
29. Arrigo A, Calamuneri A, Mormina E, et al. New insights in the optic radiations connectivity in the human brain. Invest Ophthalmol Vis Sci. 2016;57(1):1-5.
30. Párraga RG, Ribas GC, Welling LC, et al. Microsurgical anatomy of the optic radiation and related fibers in 3-dimensional images, Neurosurgery. 2012;71(1 Suppl Operative):160–171; discussion 171-172.
31. Alvarez I, Schwarzkopf DS, Clark CA. Extrastriate projections in human optic radiation revealed by fMRI-informed tractography. Brain Struct Funct. 2015;220(5):2519-2532.
32. Zeki S, Watson JD, Lueck CJ, et al. A direct demonstration of functional specialization in human visual cortex. J Neurosci. 1991;11(3):641.
33. Livingstone MS, Hubel DH.Segregation of form, color, movement, and depth: anatomy, physiology, and perception. Science. 1988;240:740.
34. Hockfield S, Tootell RB, Zaremba S. Molecular differences among neurons reveal an organization of human visual cortex. Proc Natl Acad Sci USA. 1990;87(8):3027.
35. Hubel DH, Livingstone MS.Color and contrast sensitivity in the lateral geniculate body and primary visual cortex in the macaque monkey. J Neurosci. 1990;10(7):2223.
36. Silverman SE, Trick GL, Hart WM Jr. Motion perception is abnormal in primary open-angle glaucoma and ocular hypertension. Invest Ophthalmol Vis Sci. 1990;31(4):722.
37. Horton JC, Dagi LR, McCrane EP, et al. Arrangement of ocular dominance columns in human visual cortex. Arch Ophthalmol. 1990;108(7):1025.
38. Lachica EA, Casagrande VA.The morphology of collicular and retinal axons ending on small relay (W-like) cells of the primate lateral geniculate nucleus. Vis Neurosci. 1993;10(3):403.
39. Boyd JD, Matsubara JA.Extrastriate Cortex. In: Kaufman PL, Alm A, editors. Adler's Physiology of the Eye, ed 10. St Louis: Elsevier; 2003: p. 686.
40. Catani M, Jones DK, Donato R, et al. Occipito-temporal connections in the human brain. Brain. 2003;126(pt 9):2093.
41. Horton JC.The central visual pathways. In: Hart WM Jr. editor. Adler's Physiology of the Eye, ed 9. St Louis: Mosby; 1992: p. 728.
42. Wurtz RH.Vision for the control of movement: the Friedenwald Lecture. Invest Ophthalmol Vis Sci. 1996;37:2310.
43. Blumberg J, Kreiman G.How cortical neurons help us see: visual recognition in the human brain. J Clin Invest. 2010;120(9):3054-3063.
44. Kravitz DJ, Saleem KS, Baker CI, et al. The ventral visual pathway: an expanded neural framework for the processing of object quality. Trends Cognitive Sci. 2013;17(1):26-49.
45. Jonas JB, Jonas SB.Histomorphometry of the circular peripapillary arterial ring of Zinn-Haller in normal eyes and eyes with secondary angle-closure glaucoma, Acta Ophthalmol. 2010;88(8):e317-322.
46. MacKenzie PJ, Cioffi G.Vascular anatomy of the optic nerve head. Can J Ophthalmol. 2008;43:308-312.
47. Francoisa J, Neetens A, Collette JM. Vascularization of the optic pathway. Brit J Ophthalmol. 1958;42:80.
48. Lao Y, Gao H, Zhong Y. Vascular architecture of the human optic chiasma and bitemporal hemianopia. Chin Med Sci J. 1994;9(1):38.
49. Salaud C, Ploteau S, Blery P, et al. Extrinsic and intrinsic blood supply to the optic chiasm. Clin Anat (New York, N.Y.).2018;31(3):432-440.
50. Luco C, Hoppe A, Schweitzer M, et al. Visual field defects in vascular lesions of the lateral geniculate body. J Neurol Neurosurg Psychiatry. 1992;55(1):12.
51. Ogden TE.Nerve fiber layer of the macaque retina: retinotopic organization. Invest Ophthalmol Vis Sci. 1983;24:85.
52. Ballantyne AJ.The nerve fiber pattern of the human retina. Trans Ophthalmol Soc UK. 1946;66:179.
53. Kupfer C, Chumbley L, Downer J, et al. Quantitative histology of optic nerve, optic tract, and lateral geniculate nucleus of man. J Anat. 1967;101:393.
54. Jain NS, Jain SV, Wang X, et al. Visualization of nerve fiber orientations in the human optic chiasm using photomicrographic image analysis. Invest Ophthalmol Vis Sci. 2015;56(11):6734-6739.
55. Lee JH, Tobias S, Kwon J-T, et al. Wilbrand's knee: does it exist?Surg Neurol. 2006;66(1), 11-17; discussion 17.
56. Zweckberger K, Unterberg AW, Schick U. Pre-chiasmatic transection of the optic nerve can save contralateral vision in patients with optic nerve sheath meningioma. Clin NeuroNeurosurg. 2017;115(12):2426-2431.
57. Alvarez-Fernandez D, Rodriguez-Balsera C, Shehadeh-Mahmalat S, et al. Junctional scotoma. A case report. Arch Soc Espanola Oftalmol. 2019;94(9):445-448.
58. Karanjia N, Jacobson DM.Compression of the prechiasmatic optic nerve produces a junctional scotoma. Am J Ophthalmol. 1999;128(2):256-258.
59. Pellegrini F, Lee AG, Cercato C. Multicentric glioblastoma multiforme mimicking optic neuritis. Neuro-Ophthalmology (Aeolus Press). 2018; 42(2):112-116.
60. Horton JC.Compression of the prechiasmatic optic nerve produces a junctional scotoma. Am J Ophthalmol. 2000;129(6):826-828.
61. Shin RK, Qureshi RA, Harris NR, et al. Wilbrand knee. Neurology. 2014;82(5):459-460.
62. Moore RY.Retinohypothalamic projection in mammals: a comparative study. Brain Res. 1973;49:403.
63. Berson M.Phototransduction in ganglion-cell photoreceptors. Euro J Physiol. 2007;454:849-855.
64. La Cour M, Ehinger B. The retina. In: Fischbarg J, editor.The Biology of the Eye. Amsterdam: Elsevier; 2006: p. 195-252.
65. Kupfer C.The projection of the macula in the lateral geniculate nucleus of man. Am J Ophthalmol. 1962;54:597.
66. Hickey TL, Guillery RW.Variability of laminar patterns in the human lateral geniculate body. J Comparat Neurol. 1979;183:221.
67. Malpeli JG, Baker FH.The representation of the visual fields in the lateral geniculate body of Macacamulatta. J Comparat Neurol. 1975;161:569.
68. Casagrande VA, Ichida JM.The lateral geniculate nucleus. In: Kaufman PL, Alm A, editors. Adler's Physiology of the Eye, ed 10. St Louis: Elsevier; 2003: p. 655.
69. Ferreira A, Braga FM.Microsurgical anatomy of the anterior choroidal artery. Arqu Neuro-Psi. 1990;48(4):448(abstract).
70. Balcer LJ. Anatomic review and topographic diagnosis. Ophthalmol Clin North Am. 2001;4:1.
71. Ajina S, Bridge H.Blindsight relies on a functional connection between hMT+ and the lateral geniculate nucleus, not the pulvinar. PLoS Biol. 2018; 16(7):e2005769.
72. Ajina S, Pestilli F, Rokem A, et al. Human blindsight is mediated by an intact geniculo-extrastriate pathway. ELife4. 2015.
73. Holmes G, Lister WT.Disturbances of vision from cerebral lesions with special reference to the cortical representation of the macula. Brain. 1916;39:34.
74. Horton JC, Hoyt WF.The representation of the visual field in human striate cortex. Arch Ophthalmol. 1991;109:816.
75. Wong AM, Sharpe JA.Representation of the visual field in the human occipital cortex: a magnetic resonance imaging and perimetric correlation. Arch Ophthalmol. 1999;117(2):208.
76. Reinhard J, Trauzettel-Klonsinski S.Nasotemporal overlap of retinal ganglion cells in humans: a functional study. Invest Ophthalmol Vis Sci. 2003;44(4):1568.
77. Samarawickrama C, Hong T, Jonas JB, et al. Measurement of normal optic nerve head parameters. Surv Ophthalmol. 2012;57(4):317-336.
78. Hirose T, Katsumi O.Functional changes: psychophysical and electrophysiologic measurements. In: Albert DM, Jakobiec FA, editors. Principles and Practice of Ophthalmology. Philadelphia: Saunders; 1994: p. 728.
79. Trobe JD, Glasser JS. The Visual Field Manual: A Practical Guide to Testing and Interpretation. Gainesville, Fla: Triad; 1983.
80. Mendelson JR, Wells EF.Age-related changes in the visual cortex. Vision Res. 2002;42:695.