Chapter 3 The body cavities and the diaphragm
The embryo takes on a three-dimensional shape during the fourth week when the edges of the embryonic disc are brought to the ventral surface of the embryo by lateral and longitudinal folding. Folding of the embryo results in the formation of the gut tube and converts the intra-embryonic coelom into a closed cavity. The process of folding is described in Chapter 1.
The body cavities arise from the three parts of the coelomic cavity, and become the future pericardial, pleural and peritoneal cavities. Delicate serous membranes, derived from the lateral plate mesoderm, line the walls of those cavities and cover the organs. The main function of the body cavities is to provide space for development, expansion and movement of organs such as heart, lungs and liver. The body cavities are lined by two serous layers: the somatic mesoderm, which is in contact with the ectoderm, and the splanchnic mesoderm which adheres to the endoderm (see Fig. 1.10C). The term ‘somatic’ refers to the body wall; therefore the somatic mesoderm will give rise to the parietal layer of the serous membranes. The term ‘splanchnic’ is used for organs, and the splanchnic mesoderm will form the visceral layer of serous membranes.
Septum transversum and intra-embryonic coelom
Before folding
The septum transversum is a sheet of mesoderm that appears on day 22 rostral to the developing heart (Fig. 3.1). Before folding, the intra-embryonic coelom appears as a horseshoe- or U-shaped cavity (Fig. 3.2). The bend of the U lies anteriorly, and represents the part of the coelomic cavity from which the pericardial cavity will develop. Each limb of the U consists of two cavities: (1) a pericardioperitoneal canal, from which the pleural cavity will develop, and (2) a peritoneal cavity which lies in the future abdomen. In the umbilical region, the peritoneal cavities in each limb open into the chorionic cavity or the extra-embryonic coelom. This communication allows the herniation of midgut loops into the umbilical cord (see Chapter 7).
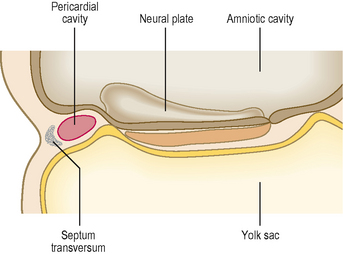
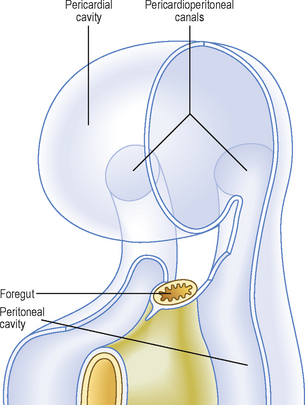
After folding
As a result of the enlarging head fold, the heart, with its pericardial cavity, swings ventral to the foregut. The septum transversum, initially cranial to the pericardial cavity, finally is wedged between the heart and the neck of the yolk sac. The pericardial cavity now opens into the paired pericardioperitoneal canals, which run on either side of the foregut to reach the peritoneal cavity. These canals end in two openings on the dorsolateral aspect of the foregut; therefore, at this stage the septum transversum is an incomplete partition between the thorax and abdomen (Fig. 3.3). The septum transversum forms the central tendon of the diaphragm.
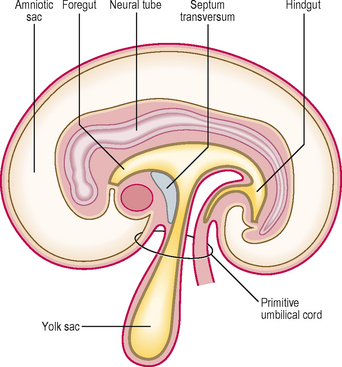
Division of the intra-embryonic coelom into four cavities
Paired mesenchymal ridges grow from the lateral body wall into the pericardioperitoneal canals and separate these paired cavities from the pericardial and peritoneal cavities (Fig. 3.4). When invaginated by the lung buds, the pericardioperitoneal canals become the pleural cavities. The pleuropericardial folds separate the pleural cavity from the pericardial cavity, and the more substantial caudal partitions, which separate the pleural cavity from the peritoneal cavity, are known as the pleuroperitoneal membranes. The pleuroperitoneal membranes are at the level of the septum transversum.
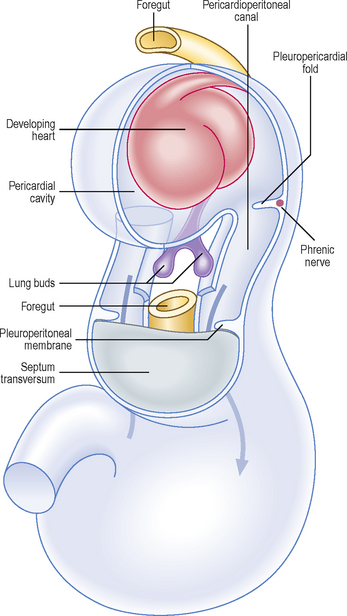
Fig. 3.4 The ventrolateral view of the embryo showing the pericardial cavity, pericardioperitoneal canals and their associated folds and membranes with surrounding tissues removed. The arrows indicate communication between the pericardioperitoneal canals and the peritoneal cavity.
As the heart descends, the pleuropericardial folds migrate ventrally and fuse with each other forming the fibrous pericardium. The thoracic cavity is then divided into a definitive pericardial cavity and two lateral pleural cavities. The phrenic nerves, which have been located within the pleuropericardial folds, run through the fibrous pericardium in the adult to supply the diaphragm.
The diaphragm
Initially, the septum transversum forms an incomplete partition between the thoracic and abdominal cavities, due to the presence of the pericardioperitoneal canals. Soon a number of mesodermal structures fuse with each other at the level of the septum transversum to form the definitive diaphragm. Five structures, described below, contribute to the development of the diaphragm (Fig. 3.5).
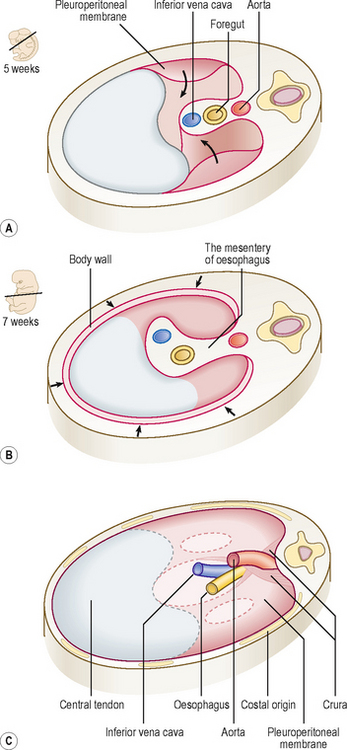
Fig. 3.5 Formation of the diaphragm as seen from above. (A) At week 5 the pleuroperitoneal membranes are growing towards the septum transversum. (B) At week 7 the membranes have fused with the septum transversum. Note the arrows indicating ingrowth of the body wall. (C) The components of the diaphragm in a newborn. The outlines of the pleuroperitoneal canals may still be seen, and are the potential sites of congenital defects.
The septum transversum
The septum transversum, composed of mesoderm, gives rise to the central tendon of the diaphragm.
The body wall
As the pleural cavities enlarge and extend into the lateral body walls, the somatic mesoderm contributes to the outer rim of the diaphragm. Extensions of the pleural cavities form the costodiaphragmatic recesses. The peripheral part of the diaphragm attached to the ribs, therefore, receives its sensory innervation from the lower six intercostal nerves.
The mesentery of the oesophagus
The mesenchyme around the oesophagus and its mesentery contribute to the connective tissues of the diaphragm. During further development, the muscular crura of the diaphragm are formed as mesenchymal condensations within the mesentery of the oesophagus.
The pleuroperitoneal membranes
These membranes fuse with the mesentery of the oesophagus and with the septum transversum, thus sealing off the pleural cavities from the peritoneal cavity. The pleuroperitoneal membranes contribute a small portion to the fully developed diaphragm.
The cervical somites
During the fourth week, the heart and septum transversum lie in the cervical region opposite the third, fourth and fifth cervical somites. At this stage the myoblasts from these somites migrate into the septum transversum and differentiate into the diaphragmatic musculature. Because of its origin the muscle of the diaphragm is innervated by the nerves of third, fourth and fifth segments of the cervical spinal cord. The nerve fibres from these segments join to form the phrenic nerve. The final thoracic position of the diaphragm is reached during the sixth week after descent of the heart and formation of the neck. This accounts for the cervical origin of the nerves supplying the diaphragm.
 Clinical box
Clinical boxDiaphragmatic herniae
If one of the pleuroperitoneal canals fails to close, the abdominal viscera may enter the pleural cavity (Fig. 3.6). This condition, called a diaphragmatic hernia, is usually on the left side. Because of the presence of the abdominal viscera in the chest, the heart is displaced, and the lungs are reduced in size. Consequently, the affected child may have severe respiratory problems even after the hernia is surgically repaired.
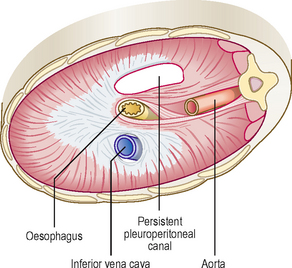
Sometimes, congenital herniae occur through small gaps in the anterior part of the diaphragm near its sternal attachment or through the oesophageal opening. These herniae, known as the parasternal and oesophageal hernia respectively, do not usually have any clinical significance. Acquired diaphragmatic herniae usually occur through the oesophageal opening and are termed hiatal herniae. Such herniae may have a peritoneal covering, and be associated with digestive disturbances.
 The space formed within the lateral plate mesoderm becomes the intra-embryonic coelom lined by the somatic and visceral layers of lateral plate mesoderm. The somatic mesodermal layer becomes the parietal serous membrane and the splanchnic mesodermal layer becomes the visceral serous membrane. After folding of the embryo, partitions divide this coelom into pericardial, pleural and peritoneal cavities. The pleuropericardial folds separate the pleural from the pericardial cavities and form the fibrous pericardium.
The space formed within the lateral plate mesoderm becomes the intra-embryonic coelom lined by the somatic and visceral layers of lateral plate mesoderm. The somatic mesodermal layer becomes the parietal serous membrane and the splanchnic mesodermal layer becomes the visceral serous membrane. After folding of the embryo, partitions divide this coelom into pericardial, pleural and peritoneal cavities. The pleuropericardial folds separate the pleural from the pericardial cavities and form the fibrous pericardium.