CHAPTER 1 Equine dental evolution
perspective from the fossil record
Introduction
It is generally believed that horses are native to the Old World and were first brought to North America by the Spanish explorers during the 16th century. While this is correct for historical times, the prehistoric fossil record of horses and their extinct relatives indicates that the Equidae underwent the majority of its evolutionary history in North America from about 55 million years ago (early Eocene) until this family became extinct about 10 000 years ago at the end of the last Ice Age (Pleistocene). The fossil record of horses in North America is a classic and compelling example of long-term (i.e., macro-) evolution.1,2 Fossil horses were exceedingly widespread and abundant in North America. Their teeth are highly durable and readily fossilize, and therefore figure prominently in our understanding of the evolutionary history of this group. This chapter will review what is known about fossil horse teeth and related morphological adaptations from the rich time sequence in North America to provide the framework within which teeth of modern Equus can be understood.
Equid interrelationships and phylogeny
Extant equids (horses, zebras, and asses) and fossil horses are classified in the family Equidae as part of the Order Perissodactyla, or ‘odd-toed ungulates.’ Other perissodactyl families include tapirs (Tapiridae), rhinoceroses (Rhinocerotidae), and several extinct families. So far as is known, all perissodactyls are united by a suite of unique characters including a concave, saddle-shaped navicular (central tarsal) facet on the astragalus (talus3), axis of symmetry through the central metapodial (III), hind-gut fermentation, and particular cheek tooth cusp morphology.2 Likewise, so far as is known, all perissodactyls living and extinct have been herbivores. With the exception of the extinct clawed chalicotheres, all perissodactyls have a foot terminating with an ungual phalanx that is either padded or hooved.
The 7–10 (i.e., depending upon classification) extant equine species can all be conservatively classified within the single modern genus Equus.4 In contrast to this single genus, about 32 extinct genera and more than 150 species of fossil horses are recognized over the past 55 million years,2,5 and these also represent a far greater diversity of morphology and adaptations than is seen in modern Equus. Fossil horses are first known 55 million years ago during the early Eocene throughout the northern continents (Fig. 1.1).6 These are represented by Hyracotherium (or ‘eohippus,’ the dawn horse) and a solely Old World group, the palaeotheres (family Palaeotheriidae).5 Horses persisted in North America after the Eocene, but this family and the horse-like palaeotheres became extinct in the Old World by the early Oligocene, 29 million years ago. During the Oligocene and later times, the major evolutionary diversification of horses occurred in North America. Ancient dispersal events resulted in three-toed (tridactyl) horses immigrating into the Old World during the Miocene 23 million years ago (Anchitherium), 15 million years ago (Sinohippus), and after 12 million years ago (hipparions; Fig. 1.1). Extinct species of one-toed (monodactyl) Equus, which first originated in North America 4.5 million years ago during the Pliocene, subsequently dispersed into the Old World across the Bering Land Bridge 3.5 million years ago.7 During the Pleistocene after about 2 million years ago, Equus species also dispersed into South America after the formation of the Isthmus of Panama. The genus Equus subsequently became extinct 10 000 years ago throughout the New World at the end of the last Ice Age (Pleistocene).
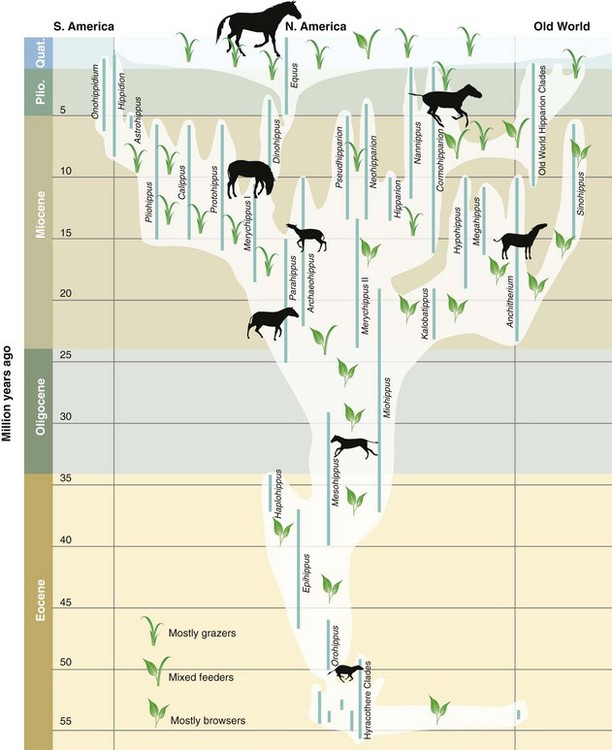
Fig. 1.1 Phylogeny, geographic distribution, diet, and body sizes of the Equidae over the past 55 million years.
(From ref.6 and reproduced with permission of the American Association for the Advancement of Science.)
Fossil horse dental adaptations
The earliest equid, Hyracotherium, is characterized by the primitive placental mammalian dental formula of three incisors, one canine, four premolars, and three molars (3 : 1 : 4 : 3), both upper and lower. The canine is large and sexually dimorphic.8 The premolars are primitive in structure, and roughly triangular in shape, whereas the molars are relatively square and have a greater surface area for trituration. During the Eocene and into the Oligocene, fossil horses in North America are characterized by progressive ‘molarization’ of the premolars (Fig. 1.2), resulting in a functional dental battery consisting of six principal teeth (P2/p2 through M3/m3) for mastication of foodstuffs. The cheek teeth of Hyracotherium and other early horses are short-crowned (brachydont). The preorbital cheek region is relatively unexpanded and the mandible is shallow (Fig. 1.3). Studies of dental structure and wear patterns suggest that these early horses were browsers, probably feeding on soft leafy vegetation and groundcover (e.g., including perhaps ferns) in ancient woodlands.8 This overall dental bauplan and inferred diet continued through the first half of equid evolution from 55 to 20 million years ago. (It also should be noted that grasslands had not yet evolved as principal biome types in North America.9)
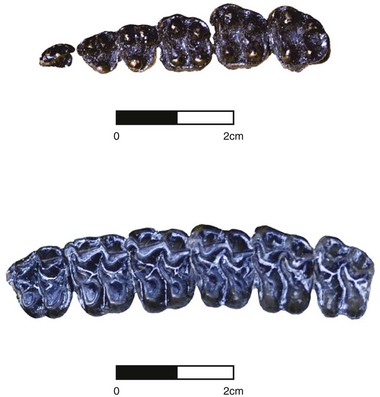
Fig. 1.2 Upper cheek tooth dentitions (excluding anterior-most P1) of Eocene Hyracotherium (top) compared with Oligocene Mesohippus (bottom). Note that relative to the triangular-shaped premolars (P2–P4; i.e., left three teeth in row) in Hyracotherium, those of Mesohippus are more square, or ‘molarized.’
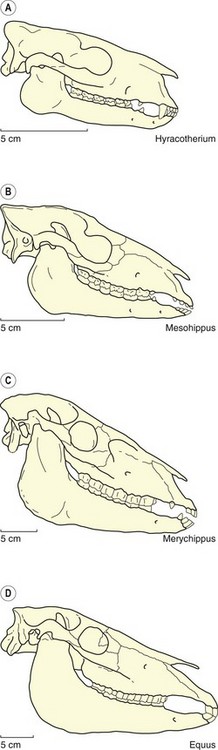
Fig. 1.3 Changes in the cranial proportions of the family Equidae as represented in Eocene Hyracotherium (top), Oligocene Mesohippus, Miocene Merychippus, and Pliocene – modern Equus (bottom).10,11
(From ref.2 and reproduced with permission of Cambridge University Press.)
The major morphological evolution of the equid skull and dentition occurred during the middle Miocene, between 20 and 15 million years ago.10–12 This evolution resulted in a morphology adapted for grazing, including a relatively longer cheek tooth row and deeper skull and jaws accommodating high-crowned (hypsodont) teeth. Miocene and later horses with hypsodont teeth are principally interpreted to have been grazers, although there are exceptions to this rule. Hypsodont teeth are well adapted to increased wear resulting from eating abrasive grasses (in contrast to soft browse), as well as ingesting contaminant grit from plants growing close to the soil substrate. Evidence from the fossil plant record indicates that grasslands became a dominant biome in North America during the middle Cenozoic9 and horses soon thereafter exploited this newly available food resource as they invaded the ‘grazing adaptive zone,’1 i.e., this is when they became grazers (Fig. 1.4).13,14 The maximum diversity of horses occurred during the middle Miocene when some dozen genera coexisted at some North American fossil localities.

Fig. 1.4 Reconstruction of a Miocene savanna grassland in North America showing a diversity of horse species, as they might have existed in a local community.
(From ref.13 and reproduced with permission of the American Museum of Natural History.)
The direct correlation between high-crowned teeth and grazing in horses is not absolute.15 Recent studies of the carbon content preserved in fossil hypsodont horse teeth indicate that some coexisting equid species secondarily acquired partial browsing diets.16 The extant genus Equus is first known 4.5 million years ago during the Pliocene from North America. It has a hypsodont dental battery and elongated and deepened skull and jaws, all of which are characters adapted for grazing (Fig. 1.3). Similar studies of the carbon content in extinct Equus teeth indicate that these horses were primarily grazers. However, depending upon available food resources and competing species, extinct Equus sometimes was a mixed feeder, incorporating some browse into its diet.
Trends in dental evolution
Number of teeth
Primitive equids from the Eocene have a dental formula of 3 I/i, 1 C/c, 4 P/p, and 3 M/m. The cheek teeth, consisting of the premolars and molars, represent the functional dental battery for post-cropping mastication. During equid evolution the rostal-most cheek teeth, P1/p1, were either reduced to small, relatively functionless teeth, or lost completely. In Equus the P1, or wolf tooth, is rudimentary, or often absent. The corresponding p1 is characteristically absent.3,17 Like most other mammalian families in which there is little evolutionary variation in the dental formula, other than the variable presence of the first premolar, equids are relatively constant in the dental formula throughout their phylogeny.
Histology
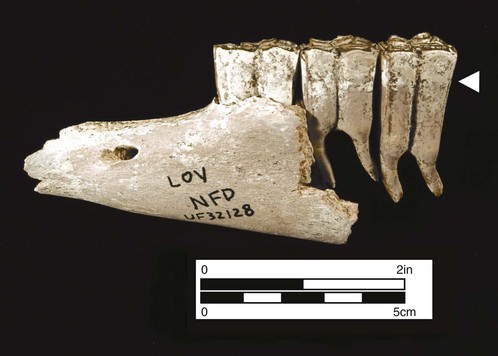
The teeth of primitive horses demonstrate three primary dental tissues: pulp, dentin, and enamel. The composition of each of these dental tissues is developmentally very conservative, i.e., there is little variation in mammals, including equids.18 Composed of collagen, connective tissue, and reticulin fibers, pulp is the relatively soft tissue located in the center of the tooth,19 but is not normally exposed on the occlusal surface unless the tooth is heavily worn. Enamel and dentin are characterized by an inorganic component consisting of the mineral hydroxyapatite (the primary constituent of vertebrate bone). Enamel is more than 95 % hydroxyapatite, whereas dentin is about 75 % mineral, the remaining portion consisting of organic compounds, mostly collagen.20 Minor chemical variations in fossil teeth result primarily from changes in diet, difference in climate, and the source elements available in the animals’ environments. Considerable infolding of the enamel occurs in later, hypsodont horses, resulting in a more durable tooth surface. Cementum, the external dental tissue in extant horses, first appeared during the Miocene in advanced species of Parahippus, and thereafter it was characteristically developed in hypsodont species (Fig. 1.5). Cementum is seen in numerous herbivorous mammalian groups and functions to provide an additional occlusal surface for mastication of abrasive foodstuffs, i.e., principally grasses.21
Dental ontogeny and wear
Most ungulates, including horses, are characterized by determinant dental growth of two sets of premolars and one molar series. Likewise, the individual teeth are characterized by growth that is completed during the lifetime of the individual when crown enamel mineralization ends and the roots form. Despite the fact that some mammals, e.g., elephants and manatees, have supernumerary tooth sets, and other mammals, e.g., rodents and lagomorphs, possess teeth that are ever-growing, the dental ontogeny in the family Equidae is conservative. A fixed set of premolars and molars and determinant tooth mineralization during an individual’s lifetime is pervasive in fossil horses and Equus, with one notable exception. One species of tiny three-toed horse, Pseudhipparion simpsoni, from the 4.5-million-year-old Pliocene of Florida, had teeth that were partially ever-growing,22 thus providing an effective dental battery for feeding on abrasive foodstuffs and potentially increasing individual longevity.
Like modern horses, individuals of fossil equid species can be aged by the relative wear on teeth as represented in large quarry samples presumed to be ancient populations. It also can be determined if breeding was synchronized, thus implying a relatively seasonal ancient environment, or occurred year-round as in more equable climates. In seasonal climates, tooth wear was discontinuous within the population because births occurred in annual cohorts, i.e., a group of individuals that all started to wear their teeth about the same time (Fig. 1.6). In contrast, species that lived in equable climates will demonstrate continuous wear because individuals were born at different times during the year.
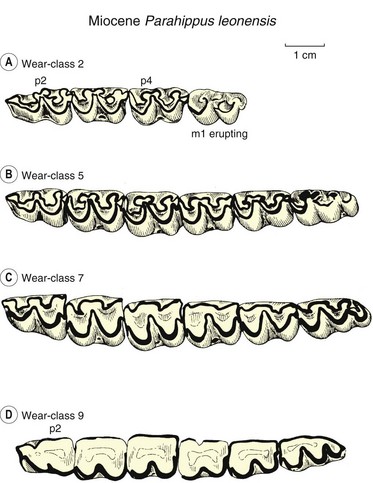
Fig. 1.6 Progressive dental wear on the lower cheek teeth of the three-toed horse Parahippus leonensis from the 18-million-year-old Thomas Farm locality, Miocene of Florida. The different wear stages shown are interpreted to represent individuals that died at different ages within the same population. The top dentition (A) probably represents an individual about 2 years old, whereas that at the bottom (D) was probably about 9–10 years old when it died. The occlusal enamel pattern is indicated in black. Pulp is exposed in the center of each tooth in Wear-class 9.
(Modified from ref.2 and reproduced with permission of Cambridge University Press.)
When horses are aged from fossil sites by the amount of wear on their teeth, we can see that potential individual longevity has evolved since the Eocene (Fig. 1.7). Eocene and Oligocene horses from 55 to 30 million years ago indicate a maximum potential longevity of 4–5 years per individual based on tooth wear and population analysis of Hyracotherium and Mesohippus. Beginning about 20 million years ago during the Miocene, cohort analyses indicate an increase in potential longevity from 5–15 years depending upon taxon,2 and thereafter up to 20–25 years per individual during the Pliocene and Pleistocene, as also has been reported for wild populations of Equus.4 As longevity is generally correlated with adult body size in modern mammals,23 it is not surprising that longevity increased in fossil horses over the past 20 million years because this also was the time of dramatic increases in body size.24
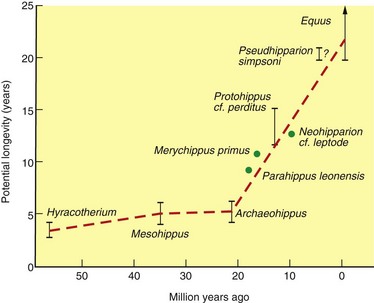
Fig. 1.7 Evolution of individual potential longevity in selected species of fossil Equidae based on analysis of the population dynamics of well-preserved quarry samples.
(From ref.2 and reproduced with permission of Cambridge University Press.)
Sexual dimorphism
Relative to certain modern mammalian species in which the males can be as much as 30–40 % larger than females within a population,4 the degree and expression of sexual dimorphism as represented in skeletal hard parts is relatively minor in living Equus. While male equids are generally larger23 and have relatively more robust canines, these sexually dimorphic characteristics are much less distinctive than in fossil equids.
A quarry sample of 24 individuals of Hyracotherium tapirinum from a 53-million-year-old (early Eocene) locality from Colorado gives insight into the sexual dimorphism in cranial and tooth size in this early horse.8 The males are on average 15 % larger than females, and have markedly robust canines relative to females (Fig. 1.8). Thereafter, during the Eocene through early Miocene, size and canine dimorphism are characteristic of more primitive species for which there are sufficient samples for statistical discrimination. With the evolution of open-country grazing forms during the Miocene, cheek teeth are essentially monomorphic,25 but sexual discrimination can be seen in the relative canine size (Fig. 1.9). Likewise, in an extraordinary quarry accumulation interpreted to represent an ancient population of Equus (E. simplicidens), the species close to the origin of the modern genus, from 3.5-million-year-old Pliocene sediments of Idaho,26 males and females can be distinguished based on relative canine size.
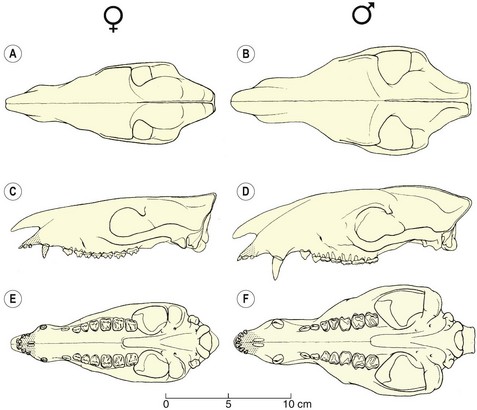
Fig. 1.8 Dorsal, left lateral, and ventral views of female (left: A, C, E) and male (right: B, D, F) crania of Hyracotherium tapirinum from the 53-million-year-old Huerfano Quarry, Eocene of Colorado. These are from the same locality and therefore interpreted to represent individuals within the same ancient population. Note the larger cranium and canine in the male. Shading indicates reconstruction.
(Modified from ref.8 and reproduced with permission of the Paleontological Society.)
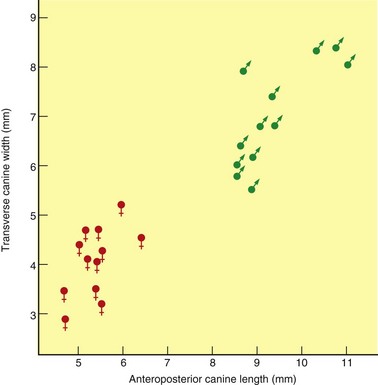
Fig. 1.9 Bivariate plot of canine length versus width in a late Miocene quarry sample of the three-toed horse Hipparion tehonense from MacAdams Quarry, Texas. The distinctly bimodal populations represent individuals interpreted to represent females (lower left) and males (upper right).
(Modified from ref.27 and reproduced with permission of the American Museum of Natural History.)
Cranial adaptations
The 55-million-year evolutionary history of the family Equidae is characterized by profound changes in cranial morphology. Primitively, Hyracotherium had a skull in which the orbit was centrally located, a postcanine diastema, and a relatively shallow mandible that accommodated short-crowned teeth (Fig. 1.8). In contrast, Equus has a preorbital region that is much longer than the postorbital region, a relatively more elongated diastema, and the mandible, which accommodates high-crowned teeth, is very deep. These trends all relate to the fundamental change in diet that occurred from the morphology seen in Hyracotherium to that of Equus. This evolution, however, was not gradual, and a major morphological reorganization occurred in equid skulls during the Miocene related to the adaptation to grazing.10,11
Although not directly related to diet and feeding adaptations, fossil horses show a fundamental evolution in the cheek region over the past 20 million years during the middle Cenozoic. Primitively, Hyracotherium has a smooth preorbital cheek region (junction of nasal, maxillary, and lacrimal bones), but during the Miocene there was an adaptive radiation resulting in an elaboration of a pit, or multiple pits, in the facial region. These are collectively termed preorbital fossae, of which the dorsal preorbital fossa is most widespread (Fig. 1.10). Preorbital fossae are absent in living Equus, so the function of this structure cannot be based on a modern closely related analog, and has, therefore, engendered much discussion in the literature. One theory suggests that preorbital fossae housed an organ complex that could have been used for vocalization. The time of maximum morphological diversity of facial fossae is seen at the time of maximum equid diversity during the Miocene. During the Pliocene and Pleistocene, when equid diversity declined, facial fossae became reduced and were ultimately lost in Equus.2
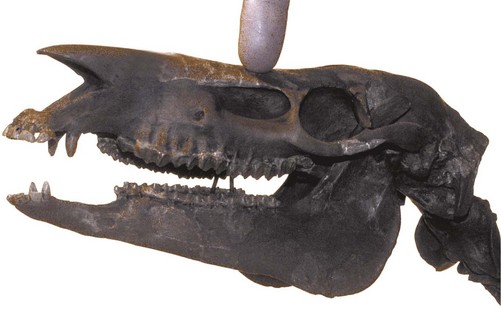
Summary: modern Equus
The cranial and dental adaptations of modern Equus, in particular the elongated preorbital region, high-crowned molarized cheek teeth, and deep mandible, represent an integrated character complex related to feeding on abrasive foodstuffs. These morphological adaptations are first seen 20 million years ago during the Miocene when equids exploited the grazing niche during the expansion of grasslands. The 55-million-year fossil record, particularly the ubiquitous and abundant horse teeth, provides fundamental evidence for macroevolution within the family Equidae in North America.
Acknowledgments
Jeff Gage, Lee Seabrook, and Tammy Johnson for preparing some of the graphic images in the text.
The US National Science Foundation supported aspects of the research presented in this chapter.
This is University of Florida Contribution to Paleobiology number 631.
1 Simpson GG. Horses: the study of the horse family in the modern world and through sixty million years of history. Oxford: Oxford University Press; 1951.
2 MacFadden BJ. Fossil horses: systematics, paleobiology, and evolution of the family Equidae. New York: Cambridge University Press; 1992.
3 Getty R. Sisson and Grossman’s The anatomy of domesticated animals. Philadelphia: WB Saunders; 1975.
4 Nowak RM, Walker’s Mammals of the world, 5.1. Online, Johns Hopkins, Baltimore, 1997.
5 McKenna MC, Bell SK. Classification of mammals above the species level. New York: Columbia; 1997.
6 MacFadden BJ. Fossil horses–Evidence for evolution. Science. 2005;307:1728–1730.
7 Lindsay EH, Opdyke ND, Johnson ND. Pliocene dispersal of the horse Equus and late Cenozoic mammalian dispersal events. Nature. 1980;287:135–138.
8 Gingerich PD. Variation, sexual dimorphism, and social structure in the early Eocene horse Hyracotherium (Mammalia, Perissodactyla). Paleobiology. 1981;7:443–455.
9 Jacobs BF, Kingston JD, Jacobs LL. The origin of grass-dominated ecosystems. Annals Missouri Botanical Garden. 1999;86:590–643.
10 Radinsky LB. Allometry and reorganization in horse skull proportions. Science. 1983;221:1189–1191.
11 Radinsky LB. Ontogeny and phylogeny in horse skull evolution. Evolution. 1984;38:1–15.
12 MacFadden BJ, Hulbert RC, Jr. Explosive speciation at the base of the adaptive radiation of Miocene grazing horses. Nature. 1988;336:466–468.
13 MacFadden BJ. The heyday of horses. Natural History. 1994;103(4):63–65.
14 MacFadden BJ. Origin and evolution of the grazing guild in New World terrestrial mammals. Trends in Ecology and Evolution. 1997;12:182–187.
15 Janis CM. An estimation of tooth volume and hypsodonty indices in ungulate mammals, and the correlation of these factors with dietary preference. In: Russell DE, Santoro JP, Sigogneau-Russell D, eds. Teeth revisited: Proceedings of the VIIth International Symposium on Dental Morphology, Paris 1986, 53. Paris (série C): Mémoires Musée National Histoire Naturelle; 1988:367–387.
16 MacFadden BJ, Solounias N, Cerling TE. Ancient diets, ecology, and extinction of 5-million-year-old horses from Florida. Science. 1999;283:824–827.
17 Sach WO, Habel RE. Rooney’s Guide to the dissection of the horse. Ithaca: Veterinary Textbooks; 1976.
18 Janis CM, Fortelius M. On the means whereby mammals achieve increased functional durability of their dentitions, with special reference to limiting factors. Biological Reviews. 1988;63:197–230.
19 Dixon PM. Dental anatomy. In: Baker GJ, Easley J. Equine dentistry. Philadelphia: WB Saunders; 1999:3–28.
20 Carlson S. Chapter 21. Vertebrate dental structures. In: Carter JS, ed. Skeletal biomineralization: patterns, processes and evolutionary trends, Vol 1. New York: Van Nostrand Reinhold; 1990:531–556.
21 White TE. The endocrine glands and evolution, no. 3: os cementum, hypsodonty, and diet. Contributions from the Museum of Paleontology, University of Michigan. 1959;13:211–265.
22 Webb SD, Hulbert RC, Jr., Systematics and evolution of Pseudhipparion (Mammalia, Equidae) from the late Neogene of the Gulf Coastal Plain and the Great Plains. Contributions to Geology, University of Wyoming, Special Paper 3. Flanagan KM, Lillegraven JA. Vertebrates, phylogeny, and philosophy, 1986:237–272.
23 Eisenberg JF. The mammalian radiations: an analysis of trends in evolution, adaptation, and behavior. Chicago: University of Chicago Press; 1981.
24 MacFadden BJ. Fossil horses from ‘Eohippus’ (Hyracotherium) to Equus: scaling, Cope’s Law, and the evolution of body size. Paleobiology. 1987;12:355–369.
25 MacFadden BJ. Dental character variation in paleopopulations and morphospecies of fossil horses and extant analogues. In: Prothero DR, Schoch RM. The evolution of perissodactyls. Oxford: Clarendon Press; 1989:128–141.
26 Gazin CL. A study of the fossil horse remains from the Upper Pliocene of Idaho. Proceedings US National Museum. 1936;83:281–319.
27 MacFadden BJ. Systematics and phylogeny of Hipparion, Neohipparion, Nannippus, and Cormohipparion (Mammalia, Equidae) from the Miocene and Pliocene of the New World. Bulletin of the American Museum of Natural History. 1984;179:1–196.