Chapter 13 Birds
The birds are a unique group of vertebrates. There are about 8500 known species in the world, all belonging to the class Aves. They are considered to have evolved about 150 million years ago from the thickly feathered Archaeopteryx, which links reptiles to birds. Birds and reptiles have much in common, such as the ability to lay eggs, but no other class of animal has feathers. The possession of feathers, and in the majority of species the ability to fly, is the reason for their success. Flight allows them to colonise new habitats, find new sources of food and escape predators. Much of their anatomy and physiology has evolved to facilitate life in the air.
Musculoskeletal system
The skeleton of the bird (Fig. 13.1) is unlike that of any other group of animals and it has developed from the evolution of powered flight. A combination of a reduction in the total number of bones and the fusion of many joints has resulted in a skeleton that provides a strong base for the attachment of the flight muscles. Although the skeleton must be light enough to enable the downward force of the wing to lift the bird into the air and to keep it airborne, when compared to mammals of a similar size birds are not exceptionally light.
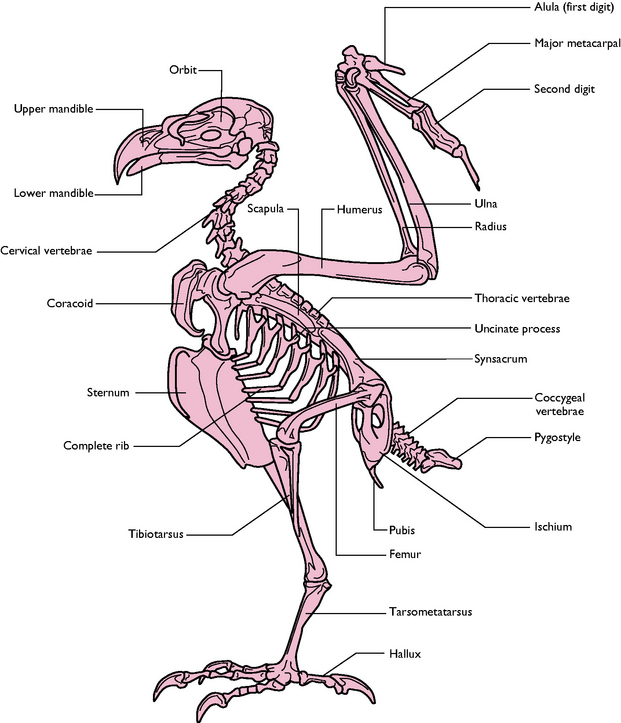
Fig. 13.1 Skeleton of a typical bird (hawk).
(With permission from Colville T, Bassett JM 2001 Clinical anatomy and physiology for veterinary technicians. Mosby, St Louis, MO, p 355.)
Main features
Bone structure

Skeletal modifications
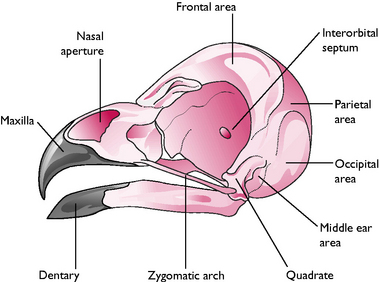
Head (Fig. 13.3)
Leg and foot
Wing
Feathers
Feathers are the distinctive feature of members of the class Aves. They develop from epidermal cells in a similar way to the hairs of mammals and the scales of reptiles. Feathers are made of keratin and create a strong but lightweight covering over the wing and the body.
All feathers have a similar structure (Fig. 13.6). The central shaft or rachis is filled with blood capillaries during growth but later, as the feather matures, it becomes hollow. On either side of the shaft, the vane consists of barbs and interlocking barbules. These hook together to form a flattened, wind-resistant surface.
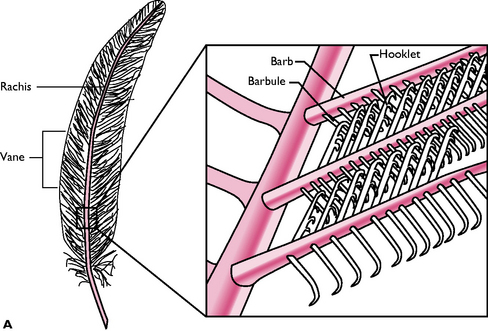
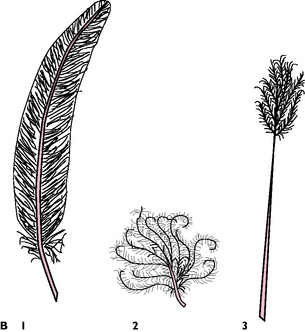
Fig. 13.6 A General structure of a feather. B Four types of feather: 1 Primary and contour; 2 Down; 3 Filoplume.
(Adapted with permission from Colville T, Bassett JM 2001 Clinical anatomy and physiology for veterinary technicians. Mosby, St Louis, MO, p 347.)
The feathers must be kept clean in order to function effectively. Birds constantly groom themselves to ‘zip up’ the barbules and to apply the secretions from the preen gland at the base of the tail, which keeps the feathers waterproof.
There are several types of feather (Fig. 13.6):
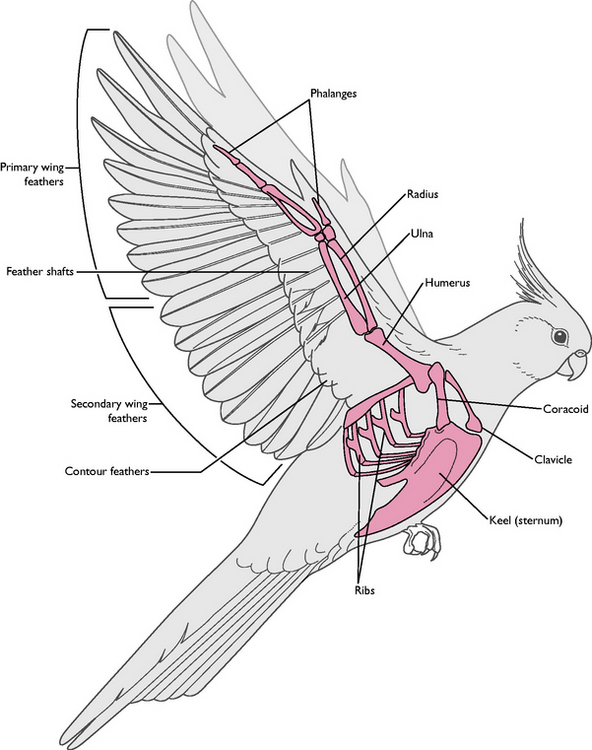
 Primaries: attached to digit 3 and to the fused metacarpal bones (Fig. 13.5). Usually 11 feathers but number varies with species; provide the major thrust during flight
Primaries: attached to digit 3 and to the fused metacarpal bones (Fig. 13.5). Usually 11 feathers but number varies with species; provide the major thrust during flightMoulting
Adult birds moult once a year, usually after the breeding season. The old feathers are lost over a period and during this time the birds are vulnerable: flight is affected and processes such as egg laying in domestic hens may cease. Production of new feathers requires energy so food intake, particularly the protein requirement, increases. Any nutritional deficiency may result in stunted feather growth or poor colouration. Feathers are used for flight, for insulation and, in some species, for display. Captive birds may be kept for their beauty and for showing.
When handling birds it is essential that the feathers are not damaged. They are only replaced at the next moult so the ability to fly may be seriously impaired, while the owner may be annoyed that their appearance has been affected.
When preparing birds for surgery, avoid plucking the feathers as this will affect the bird’s ability to keep warm until the next moult. Damp the feathers with water or spirit and part them to reveal the skin.
Flight
Most of the surface of the wing is covered with flight feathers, forming a light but strong structure. It is slightly curved, the dorsal surface being longer and convex and the ventral surface shorter and more concave. This aerofoil shape generates maximum lift and minimum drag as the wing moves through the air. Air passing over the dorsal surface has to travel faster than the air passing under the wing, resulting in lower pressure over the dorsal surface, which generates lift. The lift force must equal the weight of the bird if flight is to occur.
Control of flight is brought about by tilting the leading edge of the wing and the formation of slots between the feathers, e.g. separation of the primary feathers and the alula allows some air through, which maintains a smooth air flow on the dorsal surface, thus increasing lift. If the angle of tilt is greater than 15 ° then lift is reduced and the bird stalls or falls out of the sky.
Wing shape affects the type and speed of flight. Soaring birds such as the albatross or buzzard have broad wings that maximise lift while the wings of many garden birds are short and allow rapid bursts of flapping flight. The wings tips of the buzzard are slotted to reduce turbulence while the feathers on the edge of a barn owl’s wing are fringed to reduce the noise of the wingbeat as the bird approaches its prey.
In gliding flight the wing is held still and at a low angle so that the bird gradually loses height. Birds such as the albatross actually lose very little height because of the size and shape of the wing and because they make constant adjustments to the angle of the leading edge. Species such as the vulture and the buzzard make use of thermals or upwards warm air currents which allow them to soar and gain height.
In flapping or powered flight the bird’s body is held almost stationary and the wings are moved rhythmically up and down to generate forward thrust as well as lift. The main power is provided by the large pectoral muscles, which extend from the keel to the humerus and make up to 20% of the body mass. In strong fliers such as the pigeon the muscles are deep red because of their high myoglobin content and their good blood supply. In species such as the domestic fowl and the turkey the muscles are almost white and are capable of producing powerful bursts of flight that only last for a short time.
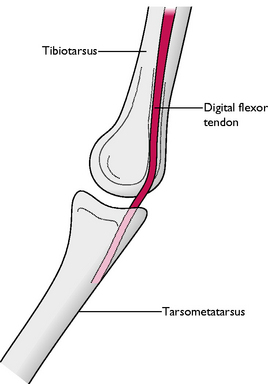
At take-off maximum lift is helped by running or by launching from a branch. To land the tail is used as a brake and the wings are extended into the stall position. The legs are extended to absorb the force of the impact and in perching birds the flexor tendons contract as the foot grasps the branch.
Special senses
The avian brain is larger in proportion to its body than that of all other vertebrates except mammals (Fig. 13.7). The basic division of the brain into fore-, mid- and hindbrain can be identified and the hind- and midbrains are similar to those of mammals. The parts of the forebrain differ because the bird needs a different range of senses in order to survive. Within the brain, the control centres for sight (optic lobes) and hearing (cerebellum) are well developed while those for touch, smell (olfactory bulbs) and taste are small and underdeveloped.
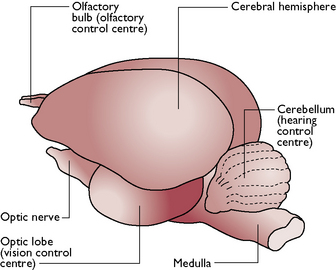
Fig. 13.7 Lateral view of a bird’s brain.
(With permission from Colville T, Bassett JM 2001 Clinical anatomy and physiology for veterinary technicians. Mosby, St Louis, MO, p 359.)
Main features
Sight
This is a highly developed sense and is essential if the bird is going to be able to fly at high speed and/or altitude, search for food, escape from predators and find a mate. The optic lobes are large and occupy the majority of the midbrain (Fig. 13.7) and much of the skull is adapted to housing and protecting the large eyes. The orbits determine the shape of the eye, which in diurnal species can be round, e.g. in hawks, or flat, e.g. in swans. In nocturnal species, the eye may be tubular and the pupil is larger in relation to the retina, enabling more light to be gathered at night.
The eyes fill the orbits, leaving room for few eye muscles associated with movement. Birds have to move their heads rather than their eyes to pick up an image. The position of the eyes on the head varies according to feeding habits: predators such as owls and hawks have forward-pointing eyes producing binocular vision but a reduced size of visual field, while seed-eaters have laterally placed eyes that provide monocular vision but a wide visual field in which to locate predators.
The structure of the eye (Fig. 13.8) is similar to that of mammals but with a few differences:
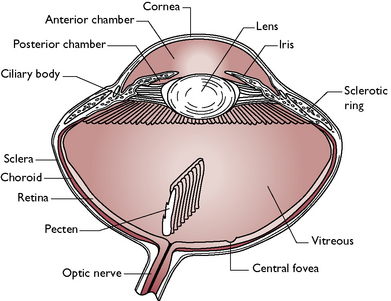
Fig. 13.8 Structure of the avian eye.
(With permission from Colville T, Bassett JM 2001 Clinical anatomy and physiology for veterinary technicians. Mosby, St Louis, MO, p 360.)
The photoreceptor cells are much thicker than those of any other vertebrate and result in a much more highly defined imageHearing
The structure of the ear is simpler than in the mammal. The ears are found on the sides of the head just behind and slightly below the eyes. There are no external ear pinnae in any species – birds such as the eagle owl that appear to have ears actually only have tufts of feathers.
The ear consists of the external ear, which collects sound and carries it to the middle ear. This is a single bone, the columella, which transmits sound to the inner ear. The inner ear is similar to that of mammals and comprises the membranous canals responsible for balance and the cochlea responsible for hearing.
Nocturnal species have an extremely well-developed sense of hearing which enables them to locate and catch prey in darkness. Barn owls, Tyto alba, in particular, have asymmetrical external ear openings (which help in the vertical location of sound), large eardrums, columellae and cochleae and a large acoustic centre in the hindbrain.
Taste
This is a poorly developed sense in birds and experiments have shown that the perception of tastes such as bitter, salty and sour is species-specific. There are relatively few taste buds and they are distributed around the sides of the tongue and soft palate.
Smell
Little is known about the avian sense of smell but it is thought to vary between species. In passerines and birds of prey it is very poor but the females of some species, e.g. mallards, secrete an odour that stimulates the males to breed. Other species, e.g. quail and albatross, use the sense of smell to locate food.
Touch
The skin is supplied with sensory nerve endings that are sensitive to heat, cold, pain and touch. Touch may be used to find food and touch-sensitive endings are found around the beak and mouth. Others are found at the base of the feathers and enable the bird to respond to the smallest movement of its feathers.
Respiratory system
The respiratory system of birds (Fig. 13.9) is very different from that of mammals and is thought to be 10 times more efficient. It is adapted to meet the needs of the bird when flying at speed and at high altitudes, where oxygen levels are low, without the bird losing consciousness. There are three main features:
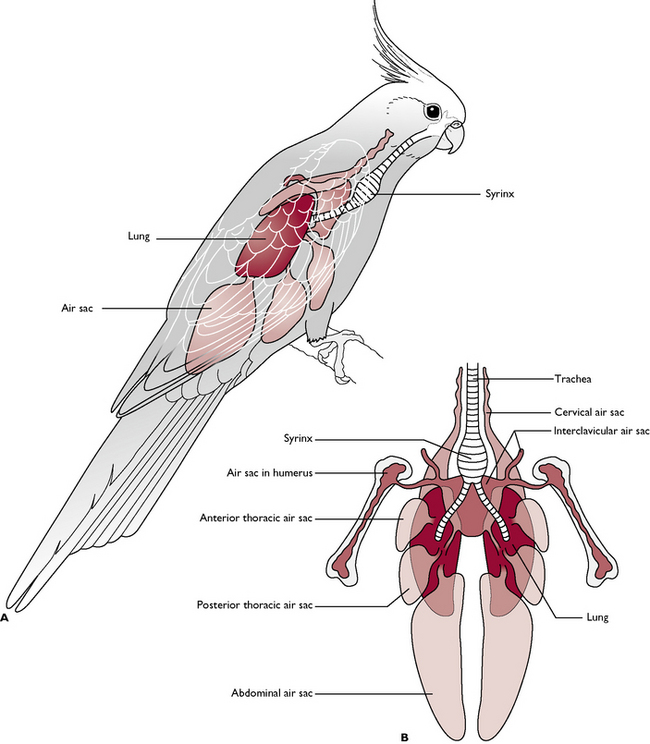
Air enters the system via a pair of nostrils or nares leading into the nasal cavity, and also through the mouth. Some birds have a cleft in the hard palate known as the choana connecting the oral and nasal cavities. The nasal cavities contain several mucosa-covered conchae, which help to warm and filter the inspired air.
Some diving birds do not have external nares as this would allow water to enter the airways during diving. They normally rely on mouth breathing. If the beak is tied to prevent the bird from pecking during treatment, there is a risk of asphyxiation.
Air passes through the glottis on the floor of the oral cavity and into a complex larynx. It travels down the trachea to the point where the trachea divides into right and left primary bronchi. At this point there is a swelling known as the syrinx, whose size and shape varies with the species. The combined effect of air passing through the larynx and the syrinx produces the characteristic sounds associated with the species.
Birds are easily intubated for anaesthesia as the glottis at the base of the tongue is easy to see. Always use an uncuffed tube to prevent rupture of the incomplete tracheal rings. The caudal air sacs may also be intubated after induction by injection or by mask, in cases where oral or beak surgery is to be performed.
The primary bronchi lead into the relatively dense lungs, closely applied to the dorsal body wall. Within the lung tissue the bronchi divide into further smaller bronchi and into cylindrical parallel tubes known as parabronchi. Air capillaries surrounded by pulmonary blood capillaries perforate the walls of the parabronchi and it is here that gaseous exchange takes place. This process is the same as occurs in mammals.
Leading from various bronchi within the lungs are thin-walled air sacs. Most birds have nine air sacs, which penetrate the spaces of the body cavity and the inside of many bones (Fig. 13.9). They are not involved in gaseous exchange and they are thought to act as a reservoir of air and to have a bellows-effect, pushing air back through the lungs. They also lighten the weight of the skeleton and so aid flight.
Respiration
During respiration air circulates continuously and passes through the lungs twice (Fig. 13.10). Most gaseous exchange occurs during the second passage. This system ensures that the removal of oxygen from the inspired air occurs with maximum efficiency.
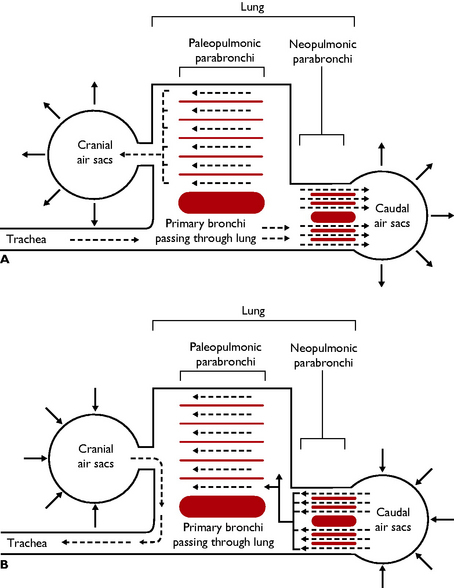
Fig. 13.10 Diagram to show the passage of air through the lungs and air sacs during respiration. A Inspiration. B Expiration.
As their respiratory system is so efficient, birds react rapidly to inhalation anaesthetics and anaesthesia may lighten or deepen much more quickly than is expected.
Circulatory system
The circulatory system follows a similar plan to that of the mammal. In order to provide for the high metabolic rate of the bird, the heart in particular must be able to pump the blood to deliver oxygen and nutrients to the tissues quickly and efficiently. In a resting chicken blood takes only 6 seconds to travel around the body.
Main features
Heart
This is a four-chambered pump lying in the cranial part of the thoracoabdominal cavity. It is covered in a pericardial sac that sticks to some of the internal surfaces to hold the heart in place.
Circulation
The arrangement of the arteries, veins and capillaries (Fig. 13.11) is similar to that of mammals with the following differences:
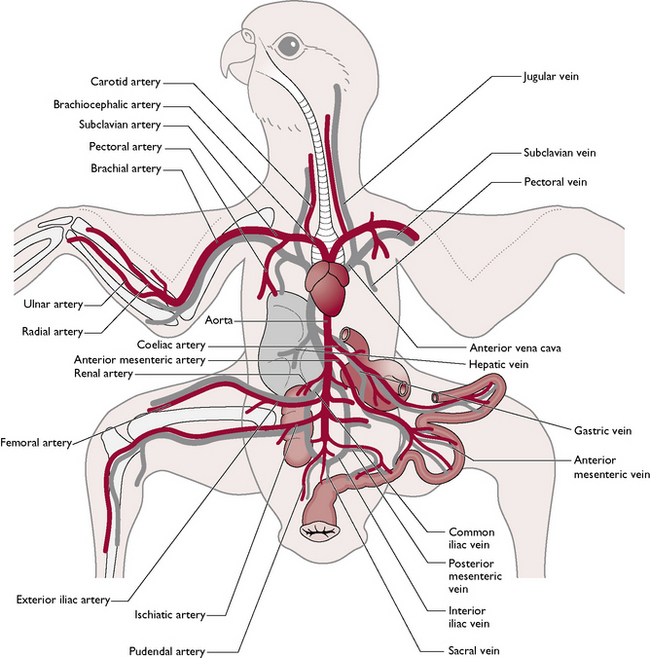
Intravenous injections or blood sampling can be carried out using the brachial or basilic vein on the medial side of the wing close to the elbow joint, the jugular vein in the neck and the medial metatarsal vein on the caudal aspect of the leg (Fig. 13.12).

Digestive system
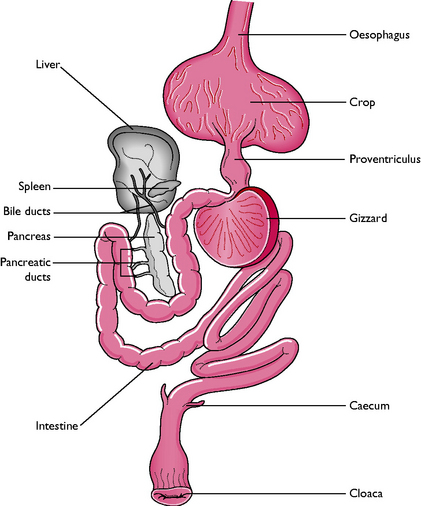
The basic pattern of the digestive tract is much the same between species of birds and the upper part shows adaptations for flight (Fig. 13.13). The majority of the tract is suspended between the wings in the body cavity to centralise the weight, enabling the bird to keep its balance and remain stable in flight without the use of a long tail.
Main features
Oral cavity
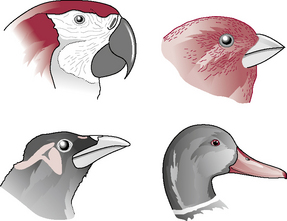
The teeth, heavy jaw bones and muscles have been replaced by a beak and lighter bones and muscles. Birds are unable to chew their food but can manipulate it and break it into small pieces using the beak and the tongue. The shape of the beak varies according to species and is suited to the particular food type (Fig. 13.14). Salivary glands are present in most species and the saliva contains mucus. In some species, such as the sparrow, the saliva is also rich in the enzyme amylase. Taste buds are found towards the back of the oral cavity and their structure is similar to those in other species.
Oesophagus and crop
Food passes down the oesophagus on the right side of the neck into the crop (Fig. 13.13). The oesophagus is thin-walled and distensible to allow the passage of relatively large pieces of food. The crop is a diverticulum of the oesophagus, lying outside the body cavity on the right side of the cranial thoracic inlet. It varies in size and shape according to the diet of the species – grain-eating birds have large bilobed crops while in owls and insectivores the crop may be rudimentary or absent.
The crop is principally a storage organ but in some species, e.g. doves and pigeons, the epithelial lining proliferates and sloughs under the influence of the hormone prolactin to produce ‘crop milk’. This is rich in proteins and fat and is used to feed the young for the first few days after hatching.
Stomach
Food passes into the stomach, which has two parts:
Small intestine
Food leaves the gizzard by the pylorus and enters the small intestine, which consists of a duodenum and ileum (Fig. 13.13). Beyond this there is no clear delineation into different parts. The pancreas, consisting of three lobes, lies in the duodenal loop and pours its secretions into the duodenum via three ducts. The liver is bilobular and relatively large and some species, e.g. chickens, ducks and geese, have a gallbladder.
Large intestine
This consists of a pair of blind-ending caeca, originating at the junction of the small and large intestines, a rectum and a cloaca. Bacterial digestion occurs in the caeca, which are large and prominent in herbivorous and granivorous species and owls but are rudimentary or absent in carnivorous and nectivorous species, e.g. hawks and parrots.
The rectum is short and terminates at the cloaca – the common exit from the body cavity shared with the urinary and reproductive systems.
Urinary system
This consists of a pair of symmetrical kidneys lying in a depression of the fused pelvic bones (Figs 13.15, 13.16). A pair of ureters carries urine to the outside via the cloaca. There is no bladder in the bird. The kidneys are relatively larger than those of mammals and occupy about 2% of body weight.
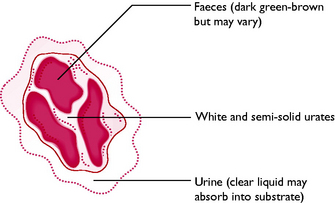
Birds excrete nitrogenous waste resulting from protein metabolism in the form of uric acid and urates. This is similar to reptiles but different from mammals, which excrete nitrogenous waste as urea. The waste materials are suspended in urinary water resulting from glomerular filtration and the resulting semi-solid urine leaves the kidneys via the ureters. In the cloaca it mixes with faecal material from the digestive tract. The material then moves by retroperistalsis into the rectum where further reabsorption of water takes place, eventually producing a small volume of ‘droppings’. Normal bird droppings (Fig. 13.17) consist of white urates and greeny-brown faeces surrounded by clear urine.
Many species of marine bird have a salt gland that enables them to deal with a salt water environment. In most birds it is located above the eye and secretes large quantities of sodium chloride. In this way the osmotic concentration of the body fluids is maintained within normal limits. Terrestrial species rely on the kidneys to regulate salt levels.
Reproductive system
Female
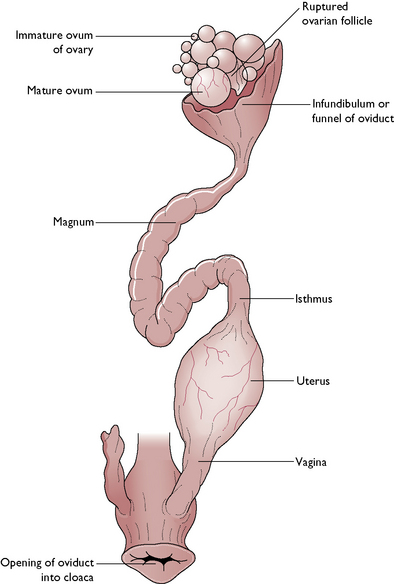
The tract comprises a pair of ovaries and oviducts leading to the cloaca (Fig. 13.15). However, in many species it is only the left side that is fully developed and functional, the right side being vestigial. Ova develop within the ovarian tissue and consist of an oocyte and a yolk surrounded by several layers of cells. During the breeding season one ovum is released at approximately 24 hour intervals until the clutch is complete.
Domestic poultry have been selectively bred to lay eggs over long periods – usually for 40 weeks per year. Wild birds will lay a clutch of eggs of a size appropriate to their species and then begin incubation.
The ovum is carried away down the left oviduct by peristaltic contractions. The oviduct is divided into distinct regions, each one of which contributes to the final egg:
The completed egg passes through a sphincter muscle into the vagina and then to the outside via the cloaca.
Male
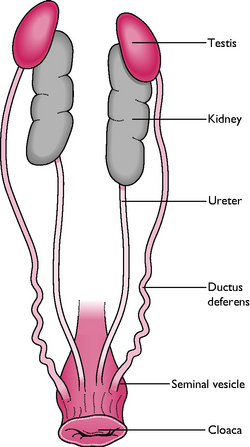
The tract comprises a pair of testes, each one connected to the cloaca by a vas deferens (Fig. 13.16). The testes lie inside the body cavity and are attached to the body wall cranial to the kidneys. In seasonal breeders, the size and weight of the testes may increase during the active breeding season.
Spermatozoa are produced within the seminiferous tubules of the testes and pass through a series of convoluted tubules to the ductus deferens, where they are stored. Mature sperm are transported to the cloaca by a vas deferens leading from each testis.
Within the cloaca is a modified area of tissue forming a phallus or rudimentary penis, which becomes engorged with lymph during mating. In many species semen is transferred into the vagina of the female by positioning the engorged phallus against the everted cloaca of the female. Species such as ducks, geese, ostrich and rhea have a well-developed phallus that is capable of erection to introduce sperm directly into the female cloaca.
Sexual differentiation
It is not always easy to differentiate between the sexes of birds but in some cases, e.g. captive breeding programmes for endangered species, it may be important to be able to do this. The value of some species, e.g. large parrots, may be increased by knowing the sex of an individual.
There are several methods available, which depend on the species:
Sexually mature female budgerigars have a pinky-brown cere above their beaks while the cere of the male is blue Female cockatiels have horizontal bars on the underside of the tail; the male loses these at about 1 year oldIt is the female bird that determines the sex of the offspring rather than the male as in mammals.