CHAPTER 37 Pulmonary Ventilation
The respiratory system supplies oxygen to the tissues and removes carbon dioxide. The major functional events of respiration include (1) pulmonary ventilation, which is the movement of air in and out of the alveoli; (2) diffusion of oxygen and carbon dioxide between the blood and alveoli; (3) transport of oxygen and carbon dioxide to and from the peripheral tissues; and (4) regulation of respiration. This chapter provides a discussion of pulmonary ventilation.
Mechanics of Pulmonary Ventilation (p. 465)
Muscles That Cause Lung Expansion and Contraction
Lung Volume Increases and Decreases as the Thoracic Cavity Expands and Contracts
Any time the length or thickness of the thoracic cavity increases or decreases, simultaneous changes in lung volume occur.
Raising and Lowering the Rib Cage Causes the Lungs to Expand and Contract
When the rib cage is elevated, the ribs project almost directly forward so the sternum also moves forward and away from the spine, increasing the anteroposterior thickness of the chest.
Movement of Air In and Out of the Lungs and the Pressures That Cause the Movement (p. 465)
Pleural Pressure Is the Pressure of the Fluid in the Space between the Lung Pleura and Chest Wall Pleura
The normal pleural pressure at the beginning of inspiration is about −5 cm of water, which is the amount of suction required to hold the lungs at their resting volume. During inspiration, expansion of the chest cage pulls the surface of the lungs with still greater force and creates a still more negative pressure, averaging about −7.5 cm of water.
Alveolar Pressure Is the Air Pressure Inside the Lung Alveoli
When the glottis is open and there is no movement of air, the pressures in all parts of the respiratory tree are equal to the atmospheric pressure, which is considered to be 0 cm of water.
Lung Compliance Is the Change in Lung Volume for each Unit Change in Transpulmonary Pressure
Transpulmonary pressure is the difference between the alveolar and pleural pressures. The normal total compliance of both lungs together in the average adult is about 200 mL/cm of water. Compliance is dependent on the following elastic forces:
Surfactant, Surface Tension, and Collapse of the Lungs (p. 467)
Water Molecules Are Attracted to One Another
The water surface lining the alveoli attempts to contract as the water molecules pull toward one another. This attempts to force air out of the alveoli, causing the alveoli to attempt to collapse. The net effect is to cause an elastic contractile force of the entire lung, called the surface tension elastic force.
Surfactant Reduces the Work of Breathing (Increases Compliance) by Decreasing Alveolar Surface Tension
Surfactant is secreted by type II alveolar epithelial cells. Its most important component is phospholipid dipalmitoylphosphatidylcholine. The presence of surfactant on the alveolar surface reduces the surface tension to one twelfth to one half of the surface tension of a pure water surface.
Smaller Alveoli Have a Greater Tendency to Collapse
Note from the following formula (Law of Laplace) that the collapse pressure generated in the alveoli is inversely related to the radius of the alveolus. This means that the smaller the alveolus the greater is the collapse pressure:
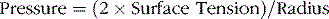
Surfactant, “Interdependence,” and Lung Fibrous Tissue Are Important for “Stabilizing” the Size of the Alveoli
If some of the alveoli were small and others were large, theoretically the smaller alveoli would tend to collapse and cause expansion of the larger alveoli. This instability of alveoli does not occur normally for the following reasons:
Pulmonary Volumes and Capacities (p. 469)
Most pulmonary volumes and capacities can be measured with a spirometer. The total lung capacity, functional residual capacity, and residual volume cannot be measured with a spirometer. Figure 37–1 shows a recording for successive breath cycles at various depths of inspiration and expiration. The recording was made using an apparatus called a spirometer.
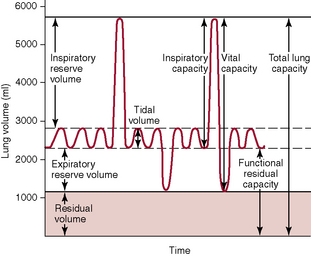
Figure 37–1 Respiratory excursions during normal breathing and during maximum inspiration and maximum expiration.
The Pulmonary Volumes Added Together Equal the Maximum Volume to Which the Lungs Can Be Expanded
The four pulmonary volumes are listed on the left in Figure 37–1.
Pulmonary Capacities Are Combinations of Two or More Pulmonary Volumes
The pulmonary capacities are listed in Figure 37–1 and can be described as follows:
Minute Respiratory Volume and Alveolar Ventilation (p. 471)
The Minute Respiratory Volume Is the Total Amount of New Air That Is Moved Into the Respiratory Passages Each Minute
It is equal to the tidal volume multiplied by the respiratory rate. The normal tidal volume is about 500 mL, and the normal respiratory rate is about 12 breaths per minute; therefore the minute respiratory volume normally averages about 6 L/min.
Alveolar Ventilation Is the Rate at which New Air Reaches the Gas Exchange Areas of the Lungs
During inspiration, some of the air never reaches the gas exchange areas but, instead, fills respiratory passages; this air is called dead space air. Because alveolar ventilation is the total volume of new air that enters the alveoli, it is equal to the respiratory rate multiplied by the amount of new air that enters the alveoli with each breath:
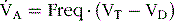
where 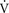 A is the volume of alveolar ventilation per minute, Freq is the frequency of respiration per minute, Vt is the tidal volume, and Vd is the dead space volume. Thus with a normal tidal volume of 500 mL, a normal dead space of 150 mL, and a respiratory rate of 12 breaths per minute, alveolar ventilation equals 12 × (500 − 150), or 4200 mL/min.
A is the volume of alveolar ventilation per minute, Freq is the frequency of respiration per minute, Vt is the tidal volume, and Vd is the dead space volume. Thus with a normal tidal volume of 500 mL, a normal dead space of 150 mL, and a respiratory rate of 12 breaths per minute, alveolar ventilation equals 12 × (500 − 150), or 4200 mL/min.
Functions of the Respiratory Passageways (p. 472)
Trachea, Bronchi, and Bronchioles
Air Is Distributed to the Lungs by Way of the Trachea, Bronchi, and Bronchioles
The trachea is the first-generation passageway, and two main right and left bronchi are the second-generation passageways. Each division thereafter is an additional generation. There are between 20 and 25 generations before the air reaches the alveoli.
The Walls of the Bronchi and Bronchioles Are Muscular
The walls are composed mainly of smooth muscle in all areas of the trachea and bronchi not occupied by cartilage plates. The walls of the bronchioles are almost entirely smooth muscle, except for the most terminal bronchioles (respiratory bronchioles), which have only a few smooth muscle fibers. Many obstructive diseases of the lung result from narrowing of the smaller bronchi and bronchioles, often because of excessive contraction of the smooth muscle itself.
The Greatest Resistance to Air Flow Occurs in the Larger Bronchi, Not in the Small, Terminal Bronchioles
The reason for this high resistance is that there are relatively few bronchi in comparison with about 65,000 parallel terminal bronchioles, each of which pass only a minute amount of air. However, under disease conditions, the smaller bronchioles often play a greater role in determining air flow resistance for two reasons: (1) they are easily occluded because of their small size and (2) they constrict easily because they have a greater proportion of smooth muscle fibers in their walls.
Epinephrine and Norepinephrine Cause Dilation of the Bronchiole Tree
Direct control of the bronchioles by sympathetic nerve fibers is relatively weak because few of these fibers penetrate as far as the central portions of the lung. The bronchial tree, however, is exposed to circulating norepinephrine and epinephrine released from the adrenal gland medullae. Both of these hormones, especially epinephrine because of its greater stimulation of β-adrenergic receptors, cause dilation of the bronchial tree.
The Parasympathetic Nervous System Constricts the Bronchioles
A few parasympathetic nerve fibers derived from the vagus nerve penetrate the lung parenchyma. These nerves secrete acetylcholine, which causes mild to moderate constriction of the bronchioles. When a disease process such as asthma has already caused some constriction, parasympathetic nervous stimulation often worsens the condition. When this occurs, administration of drugs that block the effects of acetylcholine, such as atropine, can sometimes be used to relax the respiratory passages sufficiently to relieve the obstruction.
Mucus Lining the Respiratory Passageways; Action of Cilia to Clear the Passageways (p. 473)
All the Respiratory Passages Are Kept Moist with a Layer of Mucus
The mucus is secreted in part by individual goblet cells in the epithelial lining of the passages and in part by small submucosal glands. In addition to keeping the surfaces moist, the mucus traps small particles from the inspired air. The mucus itself is removed from the passages by the actions of ciliated epithelial cells.
The Entire Surface of the Respiratory Passages Is Lined with Ciliated Epithelium
Included in these passageways are the nose and lower passages down as far as the terminal bronchioles. The cilia beat continually, and the direction of their “power stroke” is toward the pharynx (i.e., the cilia in the lungs beat upward, whereas those in the nose beat downward). This continual beating causes the coat of mucus to flow toward the pharynx. The mucus and its entrapped particles are then swallowed or coughed to the exterior.