Chapter 39 The Mammary Gland
Anatomical Aspects of the Mammary Gland
1. The milk-secreting cells of the mammary gland develop through the proliferation of epithelium into hollow structures called alveoli.
2. Most of the milk that accumulates before suckling or milking is stored in the alveoli, even though animals have enlarged milk-storage areas called cisterns.
3. A suspensory system involving the udder of the cow allows the animal to carry a large amount of milk.
1. Initial development of the mammary gland is programmed by embryonic mesenchyme.
2. Proliferation of the mammary duct system begins at puberty, with ducts under the control of estrogens, growth hormone, and adrenal steroids, and alveoli under the control of progesterone and prolactin.
1. Prepartum milk secretion (without removal) results in the formation of colostrum.
2. The ingestion of colostrum is important because of the passive immunity it confers through the presence of high concentrations of immunoglobulins.
3. The time immunoglobulins can be absorbed through the neonatal gut is limited to the first 24 to 36 hours of life.
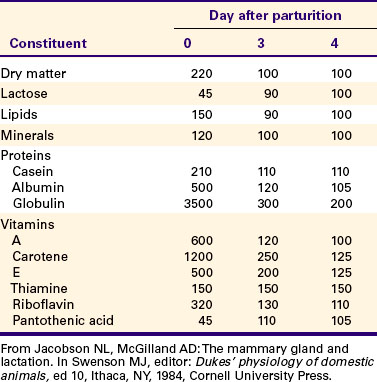
4. Lipids (particularly vitamin A) and proteins (caseins and albumins) are high in concentration in colostrum; carbohydrates (lactose) are low.
1. Prolactin, inhibited by dopamine and stimulated by vasoactive intestinal peptide, is the most important hormone involved in the process of milk synthesis, or lactogenesis; growth hormone is also important for lactogenesis.
2. The release of fat into milk from the alveolar cell involves constriction of the plasma membrane around the fat droplet; fats are dispersed in milk in droplet form.
3. Milk proteins and lactose are released from alveolar cells by the process of exocytosis.
1. Efficient milk removal requires the release of oxytocin, which causes contraction of muscle cells that surround the alveoli (myoepithelial cells), and movement of milk into the ducts and cisterns.
1. Carbohydrate stores are good in neonates born as singles or twins, whereas carbohydrate stores are low in neonates born in litters; consequently the former can stand a longer interval to first suckling than can the latter.
1. Fats are the most important energy source in milk.
2. Lactose, composed of glucose and galactose, is the main carbohydrate of mammalian milk.
3. The main proteins in milk are called caseins and are found in curd.
1. Milk production peaks at 1 month postpartum in dairy cattle, followed by a slow decline in production; milking usually stops at 305 days of lactation so that the animal can prepare the mammary gland for the next lactation.
2. Lactation can be induced by hormone administration (estrogen and progesterone) and enhanced by growth hormone and increased photoperiod exposure.
Diseases Associated with the Mammary Gland
1. The main diseases that affect the mammary gland directly are mastitis (prevalent in dairy cattle and dogs) and neoplasia (prevalent in intact dogs and cats).
2. The main conditions that involve the mammary gland indirectly are passive transfer of red blood cell agglutinating antibodies by the ingestion of colostrum (mare) and hypocalcemia caused by the transient drain of calcium that occurs with initiation of lactation (dairy cattle) or during the perinatal period (dog).
Animals that belong to the class Mammalia are characterized as having bodies that are basically covered with hair, delivering live young instead of eggs (the monotremes are an exception), and, pertinent to this chapter, nurturing their young through the use of structures called mammary glands. The ability of mammals to nurture their young through milk secretion by mammary glands during the early part of postfetal life has given these animals survival advantages. Because the reproductive strategy of mammals involves the production of far fewer young, compared with reptiles, amphibians, and birds, mammary glands have allowed mammals to be much more efficient in the nurture of their young. Egg-laying classes of animals, such as fish, reptiles, and amphibians, depend on favorable environmental factors for the nurture of their young; the offspring are often vulnerable to the vagaries of nature. Mammalian young do not require teeth for the suckling process and thus can be delivered with immature maxillae and mandibles, which facilitates the delivery of the head. The development of teeth coincides with the need to consume food other than milk.
ANATOMICAL ASPECTS OF THE MAMMARY GLAND
The Milk-Secreting Cells of the Mammary Gland Develop Through the Proliferation of Epithelium into Hollow Structures Called Alveoli
Embryonic ectoderm is the source of the mammary glands. The mammary ectoderm is first represented by parallel linear thickenings on the ventral belly wall. The continuity of the ridge that is formed is broken into the appropriate number of mammary buds, from which the functional part of the mammary gland will be derived.
The parenchyma, or milk-secreting cells, of the mammary gland develops through the proliferation of epithelial cells that arise from the primary mammary cord. The epithelial cells eventually form hollow, circular structures called alveoli, which are the fundamental milk-secreting units of the mammary gland (Figure 39-1). In concert with this development, an enlarged area of epithelium, the nipple, which is the external connection to the internal milk-secreting system, develops on the surface. In males, although nipples often develop, the underlying primary mammary cord does not develop into substantial glandular tissue.
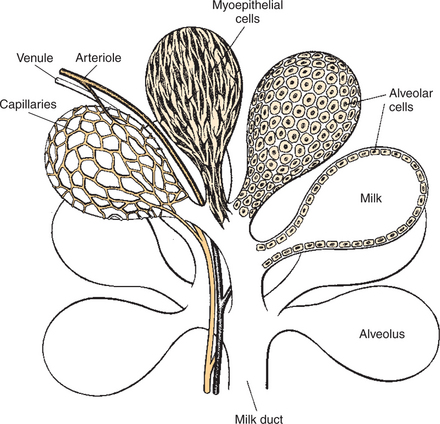
Most of the Milk That Accumulates Before Suckling or Milking Is Stored in the Alveoli, Even Though Animals Have Enlarged Milk-Storage Areas Called Cisterns
Duct systems connect alveoli with the nipple, or teat, enabling milk to pass from the area of formation to the area of delivery (nipple). The ducts may come together so that there is only one final duct per gland, which has one opening through the nipple, or teat, such as occurs in cattle, goats, and sheep. Two main ducts and associated openings occur in the mare and sow, whereas the cat and dog can have 10 or more openings in the nipple, with each opening representing separate glands (Figure 39-2). Both the cow and the doe (goat) have specialized areas for holding milk, called cisterns, which are located in the ventral part of the gland and into which all main ducts empty (Figure 39-3). This has enabled the cow, for example, to synthesize and store larger amounts of milk than would otherwise be possible. Despite this adaptation, it is important to realize that a majority of the milk present at the time of milking is stored in the duct system of the mammary glands.
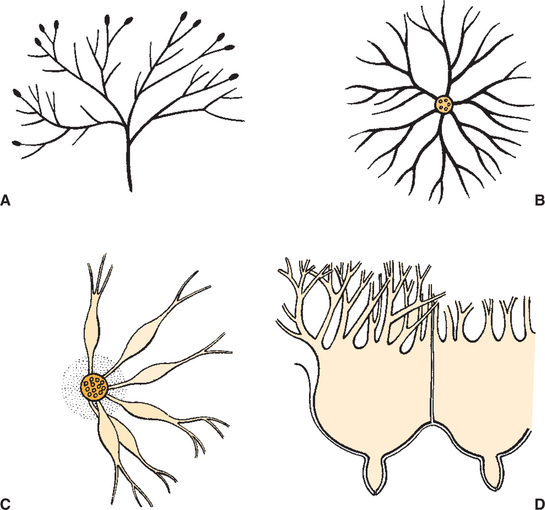
FIGURE 39-2 Diagram showing different arrangements of the mammary duct system. A, Cow, goat, sheep. B, Horse, pig. C, Cat, dog. D, Cow, goat cistern.
(From Cowie AT: Lactation. In Austin CR, Short RV, editors: Reproduction in mammals, ed 2, vol 3, Hormonal control of reproduction, Cambridge, UK, 1984, Cambridge University Press.)
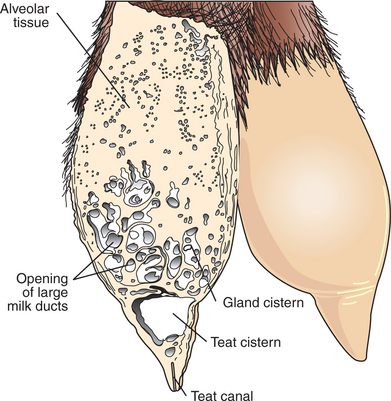
FIGURE 39-3 Depiction of the udder of a goat in which a section of the left mammary gland shows the dense alveolar tissues, the gland cistern with the large ducts opening into it, the teat cistern, and the teat canal.
(From Cowie AT: Lactation. In Austin CR, Short RV, editors: Reproduction in mammals, ed 2, vol 3, Hormonal control of reproduction, Cambridge, UK, 1984, Cambridge University Press.)
Mammary glands develop typically as paired structures. The number of pairs in domestic animals vary from one in goats, horses, and sheep; two in cattle; to seven to nine in the sow and seven to ten in the bitch and queen. The position of mammary glands varies in animals, being thoracic in primates; extending the length of the thorax and abdomen in cats, dogs, and pigs; and being inguinal in cattle, goats, and horses. In domestic species, such as cattle, goats, horses, and sheep, pairs of mammary glands are closely apposed to each other; the resulting structure is called an udder. In the cow, for example, two pairs of glands (four quarters) compose the udder.
A Suspensory System Involving the Udder of the Cow Allows the Animal to Carry a Large Amount of Milk
One of the important anatomical adaptations of the udder that allows dairy cows to carry large amounts of milk is the development of a suspension system for the udder. This system is formed by the median suspensory ligament (formed between pairs of mammary glands) composed of elastic connective tissue that originates from the abdominal tunic. The lateral (nonelastic) suspensory ligament, which originates from prepubic and subpubic ligaments, enters the glands laterally at various levels to become part of the interstitial connective tissue framework of the udder. It is not unusual for heavy-producing dairy cows to have 25 kg (55 lb) of milk in their udder immediately before milking. If the suspensory support system were not in place, the mammary gland system would soon break down from the weight of the milk.
CONTROL OF MAMMOGENESIS
Initial Development of the Mammary Gland Is Programmed by Embryonic Mesenchyme
The fetal development of the mammary gland is under both genetic and endocrine control. The initial development of the mammary bud is under control of embryonic mesenchyme (connective tissue). If mammary mesenchyme is transplanted to another area, mammary bud formation will occur at the site of transplantation. Although little is known about fetal mammary development, it is not thought to be driven by hormones. However, actively secreting mammary glands may be present at birth as a result of exogenous administration of certain hormones to the mother.
Proliferation of the Mammary Duct System Begins at Puberty, with Ducts Under the Control of Estrogens, Growth Hormone, and Adrenal Steroids, and Alveoli Under the Control of Progesterone and Prolactin
Development of the mammary gland in postfetal life usually starts in concert with puberty. Cyclical ovarian activity results in the production of estrogen and progesterone. Estrogen, with growth hormone and adrenal steroids, is responsible for proliferation of the duct system. The development of alveoli from the terminal ends of the ducts requires the addition of progesterone and prolactin (Figure 39-4).
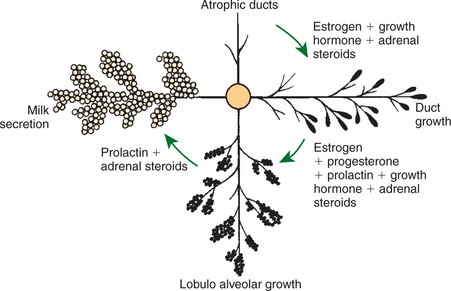
FIGURE 39-4 Hormones involved in the growth of the mammary gland and in the initiation of milk secretion in the hypophysectomized-ovariectomized-adrenalectomized rat.
(From Cowie AT: Lactation. In Austin CR, Short RV, editors: Reproduction in mammals, ed 2, vol 3, Hormonal control of reproduction, Cambridge, UK, 1984, Cambridge University Press.)
Although the development of the mammary gland begins with the onset of puberty, the gland remains relatively undeveloped until the occurrence of pregnancy. In most domestic animals, udder development usually becomes evident by the middle of gestation; the secretion of milk often begins during the latter part of gestation (mainly from increasing prolactin secretion) and results in the formation of colostrum, as discussed later. By the end of pregnancy, the mammary gland has been transformed from a structure involving mostly stromal (connective tissue) elements to a structure that is filled with alveolar cells that are actively synthesizing and secreting milk. Groups of adjacent alveoli form lobules that further combine into larger structures called lobes. Connective tissue bands delineate the lobules and the lobes (Figure 39-5).
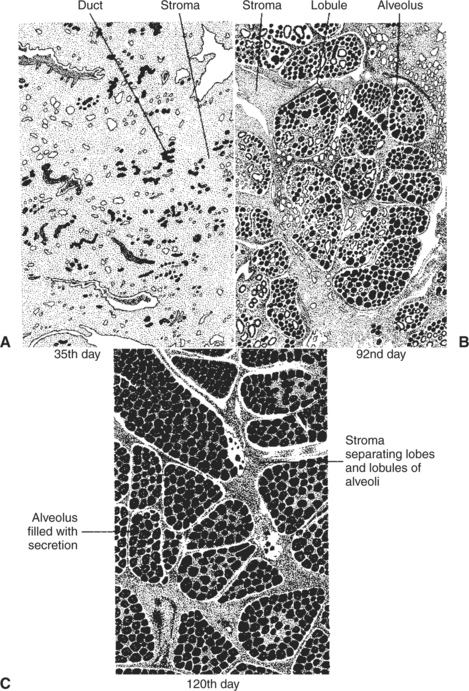
FIGURE 39-5 Drawings of sections of the mammary gland of the goat at three different times during pregnancy (which lasts approximately 150 days). A, Note the small collections of ducts scattered throughout the stroma on the 35th day. B, On the 92nd day the lobules of alveoli are forming in groups known as lobes; secretion is present in some of the alveolar lumina, and there is still considerable stromal tissue. C, On the 120th day the lobules of alveoli are almost fully developed; the alveoli are full of secretion, and the stromal tissue is reduced to thin bands separating lobules and thicker strands between lobes.
(From Falconer IR, editor: Lactation, London, 1970, Butterworths.)
COLOSTRUM
Prepartum Milk Secretion (Without Removal) Results in the Formation of Colostrum
The milk formed before parturition is called colostrum. Its formation represents a secretory process in which lactogenesis occurs in the absence of milk removal. Lactation cannot fully blossom until pregnancy is terminated, however, because of the inhibitory effects of progesterone and estrogen on milk secretion, inhibitory factors that are removed at or just before delivery.
The Ingestion of Colostrum Is Important Because of the Passive Immunity It Confers Through the Presence of High Concentrations of Immunoglobulins
When colostrum is formed before parturition, certain substances are concentrated in the process. Ingestion of colostrum is important for the well-being of the neonate. In addition to nutrition, colostrum has an important function in temporary, or passive, protection against infectious agents. Immunoglobulins (e.g., immunoglobulin A, or IgA) are produced in the mammary gland by plasma cells (derived from B lymphocytes originating in the gut) as a result of exposure of the mother to certain microorganisms. The immunoglobulins gain access to the milk system through the migration of the plasma cells from adjacent tissue sites. The immunoglobulins are highly concentrated in colostrum, and through the consumption of colostrum, the neonate can receive passive immunity against pathogens experienced by the mother. This allows the young to receive immediate protection from environmental organisms. The neonates of all domestic animals acquire antibodies through the ingestion of colostrum. The absorption of antibodies through milk in domestic animals contrasts with the situation in other species, including humans, rabbits, and guinea pigs, in which a more substantial amount of antibody is passed to the fetus through the placenta.
The Time Immunoglobulins Can Be Absorbed Through the Gut Is Limited to the First 24 to 36 Hours of Life
Neonates usually have a limited time (24-36 hours) in which immunoglobulins (proteins) can be absorbed through the gut. Thus the feeding of colostrum within this period is important to ensure the presence of immunoglobulins in the newborn. Other antimicrobial factors found in milk that are important for protection against the development of pathogenic enteric bacterial flora include lysozymes, lactoferrin, and the lactoperoxidase system.
Lipids (Particularly Vitamin A) and Proteins (Caseins and Albumins) Are High in Concentration in Colostrum; Carbohydrates (Lactose) Are Low
Colostrum is a rich source of nutrients, especially vitamin A, in addition to immunoglobulins. Placental transfer of vitamin A is limited in domestic animals, with calves and piglets being particularly low in vitamin A at birth. This deficiency is corrected by the ingestion of colostrum. Lipids and proteins, including caseins and albumins, are also present in relatively high concentration in colostrum. One exception is lactose; its synthesis is significantly inhibited by progesterone until about the time of delivery. Nevertheless, at the moment of delivery, the newborn’s milk supply is nutritive (high protein, fat, and vitamin A content) and protective (immunoglobulins) (Table 39-1).
LACTOGENESIS
Prolactin, Inhibited by Dopamine and Stimulated by Vasoactive Intestinal Peptide, Is the Most Important Hormone Involved in the Process of Milk Synthesis, or Lactogenesis; Growth Hormone Is Also Important for Lactogenesis
Prolactin plays an important role in the secretion of milk, or lactogenesis. Prolactin is released in conjunction with manipulation of the teat through either the suckling or the milking process. Sensory stimuli are carried into the hypothalamus, and the synthesis and release of dopamine, a major inhibitor of prolactin secretion, is blocked while neurons in the paraventricular nucleus are stimulated to produce and release vasoactive intestinal peptide, a stimulator of prolactin release (Figure 39-6). A short-lived surge of prolactin secretion occurs immediately after the onset of milk removal; peak values are usually reached within 30 minutes after the initial stimulus. Major surges of prolactin apparently do not need to be elicited on an hourly basis to maintain lactation because 12-hour release intervals, as occur in association with the milking of dairy cows, are sufficient to maintain lactogenesis. Prolactin responses, as judged by the amount of hormone release after mammary gland stimulation, decrease as the lactation period progresses.
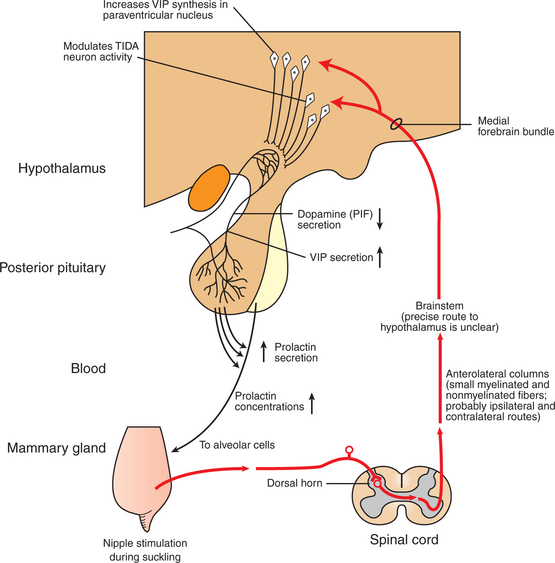
FIGURE 39-6 Somatosensory pathways in the suckling-induced reflex release of prolactin. The exact route taken by sensory information between brainstem and hypothalamus is speculative. Although tuberoinfundibular (TIDA) dopamine neuron activity is modulated as a result of the arrival of this somatosensory-derived input (prolactin inhibiting factor, PIF), the increased secretory activity of neurons containing vasoactive intestinal peptide (VIP) in the paraventricular nucleus is probably also crucial in driving prolactin secretion during suckling.
(Modified from Johnson M, Everitt B: Essential reproduction, ed 3, London, 1988, Blackwell Scientific.)
Another major hormone required for milk production in ruminants is growth hormone (GH). There is now considerable interest in the use of GH to promote additional milk production from cows through exogenous administration of the hormone.
The Release of Fat into Milk from the Alveolar Cell Involves Constriction of the Plasma Membrane Around the Fat Droplet; Fats Are Dispersed in Milk in Droplet Form
The synthesis and release of milk by alveolar epithelial cells is a remarkable physiological process (Figure 39-7). Alveolar cells synthesize fats, proteins, and carbohydrates and extrude the products into the lumen of the alveolus. Fat droplets first accumulate in the basal cytoplasm of the cell and then move to the apex, where the droplet protrudes into the alveolar lumen. The cell membrane constricts about the base of the fat droplet, so fat is dispersed in milk in small droplets, surrounded by cell membranes; the droplet often contains portions of cell cytoplasm.
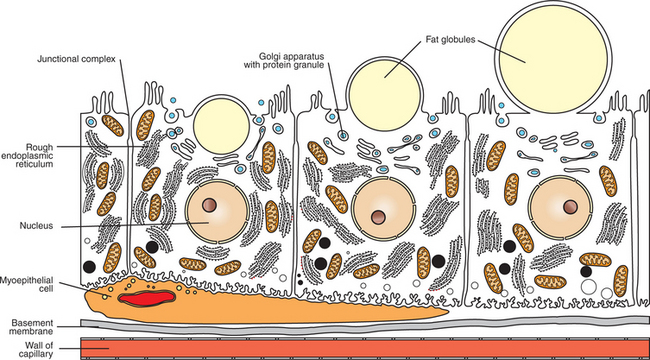
Milk Proteins and Lactose Are Released from Alveolar Cells by the Process of Exocytosis
Milk proteins are synthesized on the endoplasmic reticulum; the casein molecules pass to the Golgi apparatus, where they are phosphorylated and formed into micelles within the Golgi vesicles. Lactose is also synthesized within the Golgi vesicles and is released in conjunction with milk proteins. The process of extrusion of proteins and carbohydrates is different from that of fat; the Golgi vesicles fuse with the cell membrane, and the release of proteins and carbohydrates occurs by exocytosis. Although it is not certain how often cells go through a synthesis and extrusion cycle, it may occur twice daily, particularly in dairy cows that are milked two times per day.
MILK REMOVAL
In order for lactogenesis to be maintained, milk must be removed from the mammary gland by suckling or milking. If milk is not removed within about 16 hours in dairy cows, the synthesis of milk begins to be suppressed. As indicated previously, most of the milk in the udder of a dairy cow at the time of milking is located in the ducts and alveoli. The movement of milk into the gland cistern at suckling or milking would be slow, and less milk would be obtained during the milking of a cow, if the drainage of milk were a passive process.
Efficient Milk Removal Requires the Release of Oxytocin, Which Causes Contraction of Muscle Cells That Surround the Alveoli (Myoepithelial Cells), and Movement of Milk into the Ducts and Cisterns
To facilitate the process of milk removal, myoepithelial cells surround the alveoli and ducts (see Figures 39-1 and 39-7). The myoepithelial cells are particularly responsive to oxytocin and, in fact, contract when exposed to the hormone. The synthesis and release of oxytocin from the posterior pituitary is elicited by a neuroendocrine reflex involving tactile stimulation of the udder by suckling by the young, or the manual stimulation of washing before milking. The sensory stimuli from the udder are carried through the spinal cord into the hypothalamus. Neurons in the paraventricular and supraoptic nuclei are stimulated to synthesize oxytocin and release it from nerve terminals that impinge on the median eminence (Figure 39-8). Other sensory stimuli that elicit oxytocin release include auditory, visual, and olfactory stimuli that occur near or within the kennel, cattery, or milking parlor. Past societies used various deceptions to have earlier breeds of cattle release their milk. They often allowed the calf to suckle one teat while they milked the other glands. They also knew about the Ferguson reflex, if not in name, in which stimulation of the cervix (and release of oxytocin) was elicited by blowing air into the vagina using hollow tubes.
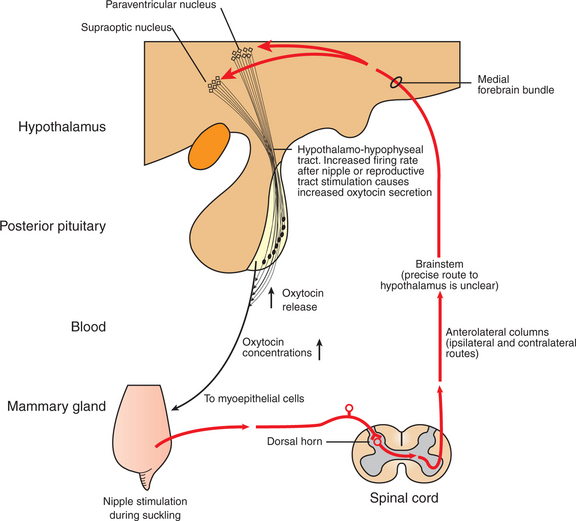
FIGURE 39-8 Somatosensory pathways in the suckling-induced reflex release of oxytocin. The actual pathway of sensory input in the hypothalamus is unknown, but it probably involves the medial forebrain bundle.
(Modified from Johnson M, Everitt B: Essential reproduction, ed 3, London, 1988, Blackwell Scientific.)
The release of oxytocin occurs within seconds after the stimulus arrives in the hypothalamus; increased pressure within the mammary gland is evident within a minute of stimulation as milk is forced out of the alveoli and ducts because of contraction of the myoepithelial cells. The term used in mammals to describe this phenomenon is milk letdown. Increased pressure within the udder is often obvious within a minute of the stimulation. The release of oxytocin lasts only a few minutes, and it is important that the milking process begin soon after milk letdown is complete (Figure 39-9). The milking process, as done by machine or by hand in earlier times, is often completed within 4 to 5 minutes.
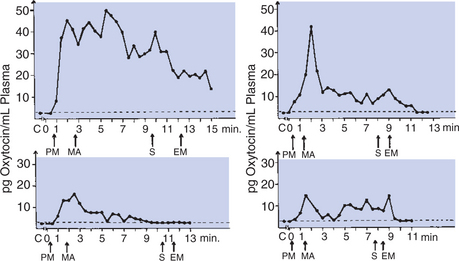
FIGURE 39-9 Oxytocin in the blood of cows before, during, and after milking. Abscissae show time in minutes. C, Control level; EM, end of machine milking; PM, preparation for milking; MA, application of teat cups; S, stripping.
(From Cowie AT: Lactation. In Austin CR, Short RV, editors: Reproduction in mammals, ed 2, vol 3, Hormonal control of reproduction, Cambridge, UK, 1984, Cambridge University Press.)
It is interesting to compare stimuli that release oxytocin, which initiates the passive part of lactogenesis, with stimuli that release prolactin, which directly influences lactogenesis. Any sensory stimulus that a cow associates with milking has the potential for releasing oxytocin. The neuroendocrine reflex is elicited in the expectation of milk removal because of the environment (kennel, cattery, or milking parlor) to which the animal is exposed. Prolactin, on the other hand, is released only by tactile stimulation of the udder. The latter makes sense, because there is no need to stimulate milk synthesis and release unless the evidence for milk removal (udder stimulation) is strong. Milk removed during hand milking is trapped in the teat and forced out, whereas milk removed by milking machines moves by suction.
FIRST NURSING
Carbohydrate Stores Are Good in Neonates Born as Singles or Twins, Whereas Carbohydrate Stores Are Low in Neonates Born in Litters; the Former Can Stand a Longer Interval to First Suckling Than Can the Latter
In domestic animals that have one or two offspring, such as cattle, horses, sheep, and goats, the young have to be able to stand in order to suckle. In this situation, neonates have reasonably good carbohydrate stores, and suckling may not occur for 1 to 2 hours without adverse effect as the young gain the ability to stand and locate the mammary gland. Young that are part of litters (cats, dogs, and pigs) are usually immediately nestled toward the mammary glands and often will be sucking in less than 30 minutes. This is important for animals born in litters because they tend to be immature at birth and susceptible to hypoglycemia, and suckling delays are often detrimental to their survival. Hypoglycemia results in stasis of the bowel (ileus) and can promote neonatal sepsis (overwhelming infection).
The suckling interval during the neonatal period varies considerably among domestic animals. Species nursing litters, such as cats, dogs, and pigs, often nurse at intervals of 1 hour or less. Goats, horses, and sheep nurse at slightly longer intervals, often up to 2 hours. Rabbits are an exception regarding the time between suckling periods; their young nurse at 24-hour intervals. As can be imagined, baby rabbits are engorged after each suckling period.
COMPOSITION OF MILK
Fats Are the Most Important Energy Source in Milk
Of the components of milk, fat is the most important energy source. Milk fat is composed of a number of lipids, including monoglycerides, diglycerides, triglycerides, free fatty acids, phospholipids, and steroids; triglycerides are the main component of milk fat. The types of lipid synthesized are complex, with great variations in both chain length and saturation of fatty acids observed on the basis of species. The amount of fat produced varies greatly both within and among species (Table 39-2). Marine mammal milk has a high fat content, with values of about 40% to 50% in seals, 40% in dolphins, and 30% in whales. In these species the high energy content of the milk through fat helps offset the heat loss of the young.
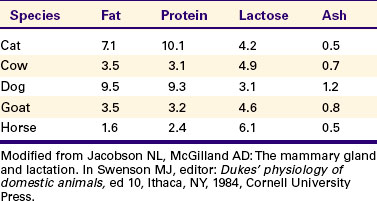
In domestic animals, sheep, swine, dogs, and cats have milk that ranges from 7% to 10% in fat content. Dairy cattle have values that range from 3.5% to 5.5%; goats are similar to cows (3.5%), and mares have lower values (1.6%). In the past, milk was sold on a butterfat basis, and breeds that had a relatively high butterfat content of milk (e.g., the Jersey with 5% butterfat) found more acceptance in dairy operations than is currently the case. Small farms produced mainly cream (for butter manufacture); the fat-concentrated portion of milk was produced by use of a separator that separated cream on the basis of specific gravity and centrifugal force. Because milk is now sold on a solids, not fat, basis, breeds that produce more milk (and protein) are favored, even though the fat content of the favored breed, Holstein-Friesian, is lower (3.5%).
Lactose, Composed of Glucose and Galactose, Is the Main Carbohydrate of Mammalian Milk
Lactose is the main carbohydrate of most mammals. It is composed of glucose and galactose. Blood glucose is the main precursor molecule for lactose, with propionate an important precursor for glucose in ruminants. Lactose is formed under the direction of lactose synthetase, an enzyme composed of α-lactalbumin (a milk protein) and galactosyl transferase. Lactose synthesis is held in abeyance until immediately before term because progesterone is inhibitory for the formation of α-lactalbumin. Prolactin, on the other hand, is stimulatory for the formation of lactose synthetase. Animals must have the enzyme lactase present in the jejunum for lactose to be cleaved (to glucose and galactose) and used. Lactase is present in most mammalian young but is sometimes not present in adult animals, including humans. In the absence of lactase, lactose can have an osmotic effect in the gastrointestinal tract, which can lead to diarrhea.
The Main Proteins in Milk Are Called Caseins and Are Found in Curd
The main proteins produced by the alveolar cells are called caseins. Caseins can be removed (as a curd) from milk through a process called curdling or coagulation, with other milk proteins, such as albumins and globulins, remaining in the fluid part of the milk (whey).
THE LACTATION CYCLE
The time required for changeover from colostrum to normal milk secretion varies with each species. In cattle, colostral milk tends to be stringy and yellow for several days postpartum. The complex bovine udder needs time for all areas to be flushed of colostrum. The milk of cattle is withheld from the milk supply for several days because of its unacceptable aesthetic quality, not because of the basic quality of the milk.
Milk Production Peaks at 1 Month Postpartum in Dairy Cattle, Followed by a Slow Decline in Production; Milking Usually Stops at 305 Days of Lactation So That the Animal Can Prepare the Mammary Gland for the Next Lactation
Milk production tends to increase for the first 3 to 4 weeks of lactation and then begins to slowly decline through the end of lactation (Figure 39-10). Cows are usually “dried up” after 305-day lactational periods; pounds of milk and butterfat production rates are calculated on this basis. Dairy animals are forced to stop lactating in order to prepare for the next lactation. The usual procedure is to stop milking. The back pressure of milk within the alveoli gradually inhibits the secretion of milk by the alveolar epithelial cells, with a resultant regression of the alveolar cells and small ducts. The process, called involution, often requires at least a month, with a 6-week period usually desired as the minimal interval from drying off to the onset of the next lactational period. Within 1 to 2 months, the secretory (alveoli) and excretory (duct) systems regress and are once again replaced. The process by which epithelial structures regress, yet retain coding for the renewal of duct and alveolar systems, is truly remarkable.
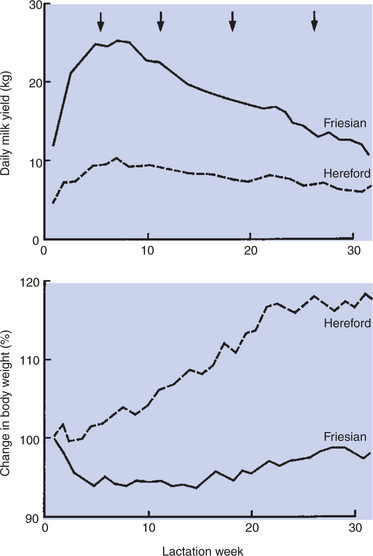
FIGURE 39-10 Average daily milk yield (top) and average percentage change in body weight (bottom) in seven low-yielding cows (broken line) and eight high-yielding cows (solid line). Arrows indicate times of blood sampling.
(Courtesy Dr IC Hart. From Cowie AT: Lactation. In Austin CR, Short RV, editors: Reproduction in mammals, ed 2, vol 3, Hormonal control of reproduction, Cambridge, UK, 1984, Cambridge University Press.)
Lactation Can Be Induced by Hormone Administration (Estrogen and Progesterone) and Enhanced by Growth Hormone and Increased Photoperiod Exposure
The induction of lactation by hormone treatment is sometimes desired, especially in dairy animals with high-lactation records but poor reproductive performance. The use of a combined treatment of estrogen and progesterone over a relatively short period (1 week) has induced alveolar development sufficiently to result in milk production. Although the amount of milk produced is less than normal, the cows can be maintained in the milking string while efforts to impregnate them continue. To induce lactogenesis by hormonal means, animals should not be lactating at treatment and should have mammary glands free of infection.
Growth hormone, which is important to the normal lactational process, can be used for the enhancement of lactation when administered over a rather wide range of concentrations (Figure 39-11). The ability to synthesize GH is relatively recent; its availability has expanded interest in its use for increasing the amount of milk produced by dairy cows. In general, GH acts on the postabsorptive use of nutrients so that protein, fat, and carbohydrate metabolism in the whole body are changed, and the nutrients are directed toward milk synthesis. If cows are in early lactation and in a negative energy balance, GH administration results in the mobilization of body fats that are used for milk formation. If cows are in positive energy balance, GH has no effect on the metabolism of body fat. Initially, GH treatment decreases the energy balance of cows; however, this is adjusted by a voluntary increase in feed consumption. Despite the increased feed consumption, GH administration increases the gross efficiency of lactation by as much as 19%. In essence, the effects of exogenous GH do not depend on gross alterations in nutrient digestibility or on body maintenance requirements. The use of GH may be economically viable, with the increased milk production justifying the expense of the hormone.
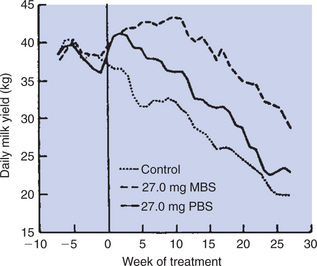
FIGURE 39-11 Average weekly milk yield of cows injected daily with diluent (control), 27 mg of methionyl bovine somatotropin (MBS), or 27 mg of pituitary bovine somatotropin (PBS). Treatments began at week 0 at an average of 84 ±10 days after parturition.
(From Tucker HA: Lactation and its hormonal control. In Knobil E, Neill J, Ewing LL, et al, editors: The physiology of reproduction, vol 2, New York, 1988, Raven Press.)
An interesting controversy has arisen from the fact that cows treated with GH do not produce “organically” derived milk, despite that synthetic GH is almost identical to endogenously derived GH. Although there is no evidence that increased concentrations of GH occur in the milk as a result of its administration, some consider the resultant milk to be abnormal.
The results with GH are in contrast to the studies in which thyroid hormone administration, in the form of iodinated casein (thyroprotein), was used to increase lactation in cows. Although the administration of thyroprotein increased lactation, extra feed was necessary to prevent excessive body weight loss, and milk production declined abruptly when thyroprotein was removed from the diet. In essence, the use of thyroprotein does not affect the efficiency of the lactational process as GH does. In dogs, one differential for gynecomastia (mammary enlargement) is profound hypothyroidism, causing elevated thyrotropin-releasing hormone (TRH) levels, which in turn stimulate prolactin secretion.
Another interesting finding of the 1980s concerning the manipulation of lactation has been that the milk yield in cows can be increased by exposing them to increased light. Cows under a photoperiod regimen of 16 hours of light (8 hours of dark) produced 6% to 10% more milk than animals under the reverse photoperiod (8 hours of light and 16 hours of dark) (Figure 39-12). Although the mechanism by which light affects lactation is not known, it likely involves prolactin secretion, at least to some extent, in that increased light exposure results in increased prolactin secretion. Similarly, the queen’s estrous cycle is affected by photoperiod, mediated by melatonin and prolactin levels. Melatonin and prolactin secretion may play a role in ovarian function in the cat, with lower levels of both hormones present during estrus than during the interestrous period. Protocols exist for inducing improved lactation in postpartum bitches using low-dose oxytocin and metoclopramide (a dopamine D-2 receptor antagonist).
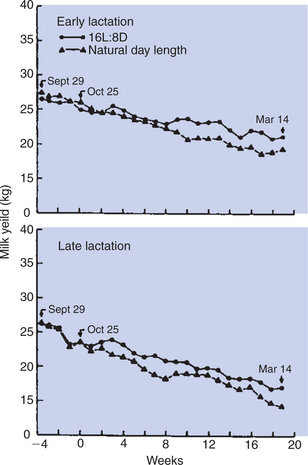
FIGURE 39-12 Influence of day length on milk production of Holstein cows. Between September 29 and October 24, cows at 37 to 74 days (early lactation) or 94 to 204 days (late lactation) after parturition were exposed to natural photoperiods of 12 hours of light per day and standardized diets. Between October 25 and March 14, cows were exposed to natural photoperiod (9-12 hours of light daily) or to 16 hours of fluorescent lighting superimposed on the natural photoperiod. L, Light; D, dark.
(From Tucker HA: Lactation and its hormonal control. In Knobil E, Neill J, Ewing LL, et al, editors: The physiology of reproduction, vol 2, New York, 1988, Raven Press.)
DISEASES ASSOCIATED WITH THE MAMMARY GLAND
The Main Diseases That Affect the Mammary Gland Directly Are Mastitis (Prevalent in Dairy Cattle and Dogs) and Neoplasia (Prevalent in Intact Dogs and Cats)
The most important problems involved in the production of milk are those caused by inflammation of the gland (mastitis). One fundamental cause of mastitis is injury to the teat canal from the repeated stretching that occurs with the milking process. Organisms that ordinarily would be excluded from the gland are able to make their way past the barrier located within the teat canal, and with repeated microorganism exposure, an infection is established.
One of the adverse consequences of mastitis is the formation of connective tissue within the udder as a result of the attempt of the gland to wall off the infection. The presence of connective tissue limits the area into which ducts and alveoli can proliferate, thus reducing the milk-producing potential of the gland. The mammary gland is an example of an organ (the eye is another example) where the elicitation of an inflammatory response is often detrimental to the function of the organ. Thus, therapies directed toward the treatment of mastitis often combine antiinflammatory and antibacterial agents.
Another process that disturbs the structure of the mammary gland is neoplasia. In domestic animals, the dog is most susceptible to the occurrence of mammary tumors. The exposure of the mammae to the ovarian hormones estrogen and progesterone greatly increases the chance of neoplasia. The incidence of mammary tumors is relatively low if the dog is ovariectomized before the first estrous cycle, but it increases progressively through exposure to two ovarian cycles; ovariectomy has little effect on neoplasia if done after the third or fourth cycle. Some owners want their dogs to have one or two cycles before they are ovariectomized. It is important for veterinarians to point out the beneficial aspects of ovariectomy before the onset of puberty because of the incidence of mammary neoplasia, as well as the usual benefits of fertility and behavioral control.
The Main Conditions That Involve the Mammary Gland Indirectly Are Passive Transfer of Red Blood Cell Agglutinating Antibodies by the Ingestion of Colostrum (Mare) and Hypocalcemia Caused by the Transient Drain of Calcium That Occurs with Initiation of Lactation (Dairy Cattle) or During the Perinatal Period (Dog)
An immunological disease associated with the mammary gland involves the transfer of red blood cell agglutinating antibodies to the fetus through the milk. The situation is most common in the horse, in which fetal red blood cells (RBCs) pass into the maternal system and elicit antibody formation against the fetal RBCs. These antibodies tend to be concentrated in the colostrum along with other immunoglobulins. At birth the foal is able to absorb the RBC agglutinating antibodies (as well as other beneficial immunoglobulins) for up to 48 hours. Foals often go into a hemolytic crisis between 24 and 48 hours after delivery and can die unless given vigorous therapy, including blood transfusions. If fetal RBC antibody formation is suspected in a mare, the disease can be handled by muzzling the foal at birth through 48 hours and feeding with colostrum saved (frozen) from other preparturient mares. A similar condition has been reported in kittens born to queens sensitized by a previous litter with dissimilar blood type.
A disease associated with the mammary gland and life threatening to the dam is hypocalcemia. At parturition the acceleration of lactogenesis causes a great increase in the movement of calcium from the blood into the milk. Both cows and dogs are particularly susceptible, with some dams unable to respond immediately to the calcium drain from the blood by the mobilization of calcium. As a result, the animals lose their ability to maintain normal muscle activity, are often unable to stand, and become prostrate with the appearance of being comatose. The syndrome occurs in cows at parturition and in dogs during the last weeks of pregnancy or the first few weeks postpartum, when lactation reaches its peak. Inappropriate prenatal nutrition, often with calcium supplementation, sets bitches up for this condition by inhibiting normal parathyroid gland development, necessary to meet the demands for mobilization of calcium by lactation. The systemic administration of calcium to hypocalcemic cows often produces a dramatic recovery in 10 to 20 minutes.
CLINICAL CORRELATIONS
Pregnant Mare That Does Not Have Sufficient Milk or Udder Development
History.
A 13-year-old pregnant Arab mare is due to foal in the next week, based on breeding dates. This is her second foal; the first foal did well. She has had limited udder development. The mare is current on vaccinations and deworming; she has no previous medical problems. She has been on pasture with all the other horses. She is supplemented with 2 pounds of 14% mare and foal feed per day. She is given grass hay, and she is in good body condition. The farm is located in eastern Tennessee.
Clinical Examination.
The mare is pregnant. The foal is viable based on movement and heartbeat. The foal is in the pelvic inlet, indicating that the mare will foal soon. All other parameters of the examination are normal. The mare is in good body condition.
Comment.
With a mare close to parturition that has had limited udder development, one concern is whether the mare has been fed fescue. The owners are asked about this and say that she has been receiving a grass hay with fescue in it. They did not know that fescue could cause problems. The clients are informed that fescue can contain an endophytic fungus, Neotyphodium coemophialum, which produces alkaloid toxins. These toxins are dopaminergic and inhibit prolactin. Additionally, the increase in dopamine activity directly decreases prolactin. Normally, neurons in the proventricular nucleus would release prolactin, which would stimulate lactogenesis. In some animals, progesterone levels are also decreased.
Treatment.
Domperidone can be given because it inhibits the dopaminergic effects. If domperidone is started 5 to 10 days before parturition, this is often sufficient time for the mare to develop sufficient milk for the foal. If domperidone is not started until up to 24 hours after foaling, it must be given for 10 to 14 days. Some mares will respond and will increase the milk production, whereas some mares do not respond. Although this treatment is often useful in stimulating milk production by the mare, it does not necessarily reverse other complications associated with fescue. Mares that have been given fescue may have foals that appear dysmature, are weak, or have prolonged gestation. The placenta from these affected mares is often thickened. To prevent these effects of fescue, clients are encouraged to keep the mares off fescue-containing feeds for at least 30 to 60 days before foaling. In addition, endophyte-free fescue grasses are available, but expensive.
Neonatal Isoerythrolysis
History.
You are called to examine a mare, 7 months pregnant, that has a previous history of having conceived and delivered a normal foal after her first pregnancy; the foal was subsequently suckled and was sold as a weanling. The mare had no trouble conceiving and carrying the next two pregnancies, but the foals died within 2 to 4 days of birth, even though they were healthy and vigorous at birth and the mare had colostrum and milk. The previous owner became discouraged because of these deaths and sold the mare to the current owner at a bargain price.
Clinical Examination.
You perform a general physical examination of the mare and find all organ systems to be normal. Palpation of the uterus per rectum reveals the presence of a viable fetus that appears to be of the correct size for a pregnancy of the purported duration. Both the external genitalia and the mammary glands are normal in appearance.
Comment.
From the history, and because the mare appears to be undergoing a normal pregnancy, you conclude that there is nothing wrong with the reproductive process per se. The fact that the previous two foals were healthy at birth and yet weakened rapidly and died within 4 days indicates that something likely happened to them after delivery. If the deaths were caused by an issue associated with the mare, the most likely cause of these deaths would be neonatal isoerythrolysis. In this situation the mare becomes exposed to the red blood cells (RBCs, erythrocytes) of the fetus during pregnancy, or the mare could have been exposed to RBCs from a stallion, whose erythrocytes are recognized as foreign. If the mare is exposed to the RBCs from the stallion or the fetus, those RBCs enter the circulation of the dam. She responds by making antibodies to the RBCs because of the presence of foreign antigen on the fetal erythrocytes that were inherited from the sire. In the mare, these antibodies do not pass through the placental barrier, so the fetus is protected from these antibodies during pregnancy. The antibodies do pass into the colostrum and are concentrated during the process of colostrum formation. Therefore, when the foal suckles the colostrum, it acquires the antibodies that will react to its own RBCs. The foal develops a type II hypersensitivity reaction, in which the antibodies destroy the foal’s RBCs through different mechanisms.
Treatment.
The foal needs to be prevented from suckling the mare for the first 2 to 3 days of life. During the first 1 to 2 days, the foal is able to absorb large protein molecules, including the important immunoglobulins that enable the foal to ward off infections, as well as, in this case, antibodies against fetal RBC antigens. The gut epithelium closes to the passage of large protein molecules by 36 to 48 hours of life; at this time, or shortly thereafter, the foal can be allowed to suckle without risk of absorbing the antibodies. The key is to prevent the foal from suckling during the first 2 to 3 days of life to prevent absorption of the antibodies that will react to its own RBCs. The mare needs to be monitored closely before parturition so that the foal can be muzzled shortly after delivery. The foal does need nourishment during the first 2 to 3 days of life; thus it is important that the foal be fed colostrum obtained from other mares (usually maintained frozen). If the foal does receive the mare’s colostrum containing antibodies that will react to its own RBCs, the foal can still be treated. The foal should not nurse the mare for the first 3 to 5 days of life, and the foal can be given packed RBCs from a donor if necessary. To anticipate the potential risk, blood typing of stallions and mares is now available.
Bogaerts P: Clinical approach to genital and mammary pathologies in cats. Presented at 5th Biannual Congress, European Veterinary Society for Small Animal Reproduction (EVSSAR), Budapest, Hungary, 2006.
Cowie T. Lactation. In: Austin CR, Short RV. Reproduction in mammals, ed 2, vol 3, Hormonal control of reproduction. Cambridge, UK: Cambridge University Press, 1984.
Feldman EC, Nelson RW. Canine and feline endocrinology and reproduction. Philadelphia: Saunders, 2004.
Leyva H, Madley T, Stabenfeldt GH. Effect of light manipulation on ovarian activity and melatonin and prolactin secretion in the domestic cat. J Reprod Fertil Suppl. 1989;39:125.
Neill JD, ed. Knobil and Neill’s physiology of reproduction, ed 3, Philadelphia: Elsevier, 2005. vols 1 and 2
Park CS, Lindberg GL. The mammary gland and lactation. Reece WO, ed. Dukes’ physiology of domestic animals, ed 12, Ithaca, NY: Cornell University Press, 2004.
PRACTICE QUESTIONS
1. The development of the duct system in the mammary gland is under the control of estrogens, growth hormone, and adrenal steroids. If the duct system is to develop functional milk-secreting units, called alveoli, which of the following hormone(s) are essential to this development?
2. The hormone that is most important for the maintenance of lactation (lactogenesis) is:
3. Sensory inputs, including sound, sight, and smell, but not necessarily touch, elicit the release of what important hormone required for the lactation process in the cow?
4. The contraction of what anatomical structure is of fundamental importance for the release of milk from the udder of the cow?