Chapter 53 Thermoregulation
1. Temperature is a major factor affecting tissue function.
2. Homeotherms and poikilotherms use different strategies to regulate body temperature.
3. Body temperature depends on the balance between heat input and heat output.
1. Heat is a byproduct of all metabolic processes.
2. Shivering produces heat by muscle contraction.
3. Nonshivering thermogenesis is an increase in basal metabolic rate, caused especially by the oxidation of fats, to produce heat.
1. Because tissues are poor conductors, heat is most effectively transferred in the blood.
2. Countercurrent heat exchange mechanisms are used both to conserve and to lose heat.
Heat Exchange with the Environment
1. Heat loss by convection occurs when the body warms air or water.
2. Heat loss by conduction occurs when the body is in contact with a cooler surface.
3. Heat loss by radiation occurs when infrared radiation emitted by the body is absorbed by cooler objects.
4. Heat loss by evaporation occurs when the water in sweat, saliva, and respiratory secretions is converted into water vapor.
1. Mammals and birds regulate the input and output of heat to maintain body temperature within a narrow limit.
2. Temperature-sensitive receptors are located in the central nervous system, the skin, and some internal organs.
3. Information from central and peripheral heat-sensitive neurons is integrated in the hypothalamus to regulate heat-losing or heat-conserving mechanisms.
1. The responses to heat stress are peripheral vasodilation and increased evaporative cooling.
2. The responses to cold stress are peripheral vasoconstriction, piloerection, and increased metabolic heat production by shivering and nonshivering thermogenesis.
3. Fever is an elevation of body temperature that results from an increase in the thermoregulatory set point.
Heat Stroke, Hypothermia, and Frostbite
1. Heat stroke occurs when heat production or input exceeds heat output, so body temperature rises to dangerous levels.
2. Hypothermia occurs when heat output exceeds heat production, so body temperature decreases to dangerous levels.
3. Frostbite occurs when ice crystals form in the tissues of the extremities.
Temperature Is a Major Factor Affecting Tissue Function
Because body function is the result of chemical and physical processes that are sensitive to changes in temperature, animals use a variety of strategies to regulate the temperature of their tissues. If body temperature is allowed to decrease too much below its normal value of about 38° C (~100° F), metabolic processes are slowed to such an extent that body function ceases. Below 34° C (93.2° F), animals become unable to regu-late their own temperature, and at 27° to 29° C (80.6°–84.2° F), cardiac fibrillation and death occur. At the other extreme, an increase in temperature to 45° C (113° F) can cause fatal brain lesions.
Homeotherms and Poikilotherms Use Different Strategies to Regulate Body Temperature
Fish, reptiles, and amphibians are called “cold-blooded animals,” or poikilotherms, because their body temperature varies with the temperature of the environment. However, this does not mean that these animals have no control over their body temperature. They use behavioral methods to prevent major changes in their temperature. For example, the lizard basks on a sun-baked rock to increase its temperature early in the morning and hides beneath the rock later in the day to prevent overheating. Veterinarians are sometimes asked to advise on the management of captive poikilotherms; it is important to remind owners to provide supplemental heat if they want their animals to be active at the cooler times of the year.
Mammals and birds are homeotherms; they maintain a constant body temperature in the presence of considerable changes in environmental temperature. Although the maintenance of a constant temperature allows mammals to live in a wide variety of environments and to remain active during the cold times of the year, it is not without cost. Homeotherms must maintain a high metabolic rate just to provide the heat necessary to maintain body temperature. This requires a high energy intake and therefore almost constant foraging for food. Poikilotherms require much less energy and are better able to survive times of food shortage. Because most veterinarians are primarily concerned with mammals and birds, this chapter focuses on the maintenance of a normal body temperature by homeotherms.
Body Temperature Depends on the Balance Between Heat Input and Heat Output
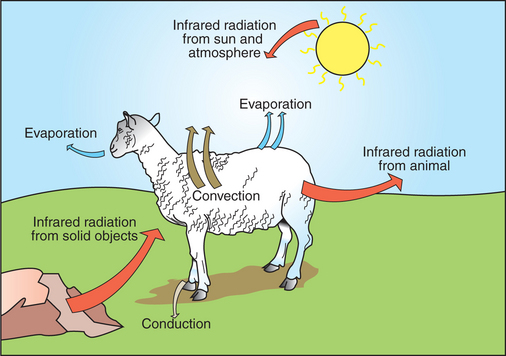
Heat inputs to the body are from metabolism and from external sources (Figure 53-1). Once food energy is ingested, heat is produced at all stages of the metabolic process. Eventually, all food energy is converted into heat, which is dissipated into the environment and radiated into space.
Heat production by the body is related to metabolic rate. A basal metabolic rate is necessary to maintain the function of cells. During exercise, metabolic heat production can increase more than 10-fold. If this heat is not dissipated to the environment, body temperature can increase to dangerous levels. Furthermore, this increase in body temperature increases metabolic rate, which further increases heat production.
Animals gain heat from the environment when ambient temperature exceeds body temperature and when they are exposed to radiant heat sources. The latter occurs when an animal is exposed to sunlight or is placed close to solid objects that are warmer than its body temperature.
Heat is lost to the environment by radiation from the body surface to a cooler object; by convection as the surrounding air or water is warmed by the body; by evaporation of respiratory secretions, sweat, or saliva; and by conduction to cooler surfaces with which the animal is in contact. A small amount of heat is also lost with urine and feces.
Many of the metabolic heat sources, such as the liver, heart, and limb muscles, are remote from the skin, which is the site of heat loss. Therefore, it is necessary to transfer heat among these sites. Body tissues are poor conductors, so heat is transferred mainly by convection in the circulation.
HEAT PRODUCTION
Heat Is a Byproduct of All Metabolic Processes
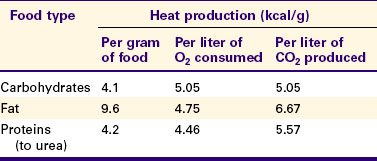
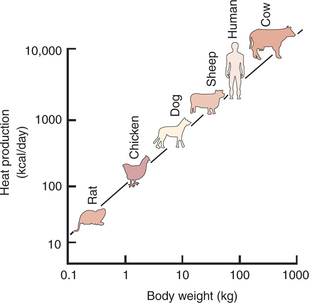
Table 53-1 shows the amount of heat produced by the metabolism of carbohydrates, fats, and proteins. The basal metabolic rate (BMR) is the rate of energy metabolism measured under minimal stress while the animal is fasting. BMR is greater in homeotherms than in poikilotherms because homeotherms need to generate heat to maintain body temperature. The BMR per kilogram of body weight is greater in smaller than in larger mammals (Figure 53-2). This is necessitated partly by the greater surface/volume ratio of smaller animals. The relatively greater surface area per kilogram body weight of small animals provides a larger area for heat loss.
Shivering Produces Heat by Muscle Contraction
Shivering is one method of increasing the metabolic production of heat. Antagonistic groups of limb muscles are activated so that they produce no useful work. The chemical energy used in shivering is transferred to the body core as heat. If necessary, shivering can go on for many hours and can double heat production. Short bursts of shivering can even increase heat production up to fourfold.
Nonshivering Thermogenesis Is an Increase in Basal Metabolic Rate, Caused Especially by the Oxidation of Fats, to Produce Heat
When animals are chronically exposed to cold, they develop the ability to increase metabolic heat production without shivering (nonshivering thermogenesis). This increase in metabolism is mediated through an increase in thyroxine secretion and the calorigenic effects of catecholamines on lipids. Table 53-1 shows that fat metabolism is an effective way to produce heat. Brown fat is a specialized vascular, mitochondria-rich fat found between the scapulae of newborn small mammals. Catecholamines increase metabolism in all fats, but particularly in brown fat, and the heat produced is distributed around the body through the bloodstream.
HEAT TRANSFER IN THE BODY
Because Tissues Are Poor Conductors, Heat Is Most Effectively Transferred in the Blood
Because heat is produced primarily in muscles of the limbs and in the liver and is eliminated through the skin and the respiratory tract, it is necessary to transfer heat around the body. Tissues have a thermal conductivity similar to that of cork; therefore, conduction is not an efficient means of heat redistribution.
The blood perfusing a metabolically active organ collects heat and transfers it to cooler parts of the body by circulatory convection. Redistribution of blood flow can deliver heat preferentially to certain body regions, or it can allow regions to cool when the maintenance of the temperature of the brain and major viscera (core temperature) is threatened.
Under conditions of heat stress, circulatory transfer of heat to the skin can be increased dramatically by two mechanisms. First, the arterioles of skin vascular beds dilate, which results in increased capillary blood flow. Second, arteriovenous anastomoses open in the limbs, ears, and muzzles. These two actions greatly increase the total blood flow to the periphery, and the increased heat delivery increases the temperature of the skin, which facilitates heat loss. Conversely, under cold stress, skin vascular beds vasoconstrict, and arteriovenous anastomoses close; thus skin and limb temperatures decrease. This results in reduced heat loss from the skin and in a gradient of temperatures along the limb (Figure 53-3). Under severe cold stress, the skin temperature of the extremities can approach ambient temperature. Interestingly, the lipids in the limb extremities have a lower melting point than those in the core, so fats do not solidify in extreme cold stress.
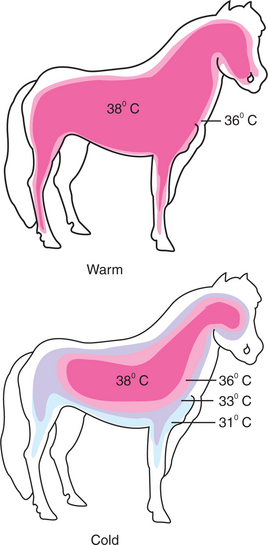
FIGURE 53-3 Representation of the distribution of temperatures in a pony under warm and cold environmental conditions. Under warm conditions, the core body temperature extends into the limbs and close to the skin surface of the animal. Under cold conditions, vasoconstriction in the peripheral blood vessels results in a gradient of temperatures between the core and the extremities. The core temperature is maintained only in the abdomen, thorax, and brain of the animal. The more peripheral tissues are allowed to cool considerably.
Countercurrent Heat Exchange Mechanisms Are Used Both to Conserve and to Lose Heat
When the environmental temperature is high, the blood perfusing the skin vascular beds returns to the core through superficial veins from which heat is lost to the skin and air. Under cold conditions, limb blood flow returns to the core through deep veins that accompany arteries (Figure 53-4). Heat is transferred by countercurrent exchange from the warm arterial blood to the cooler venous blood and thereby returned to the core of the body.
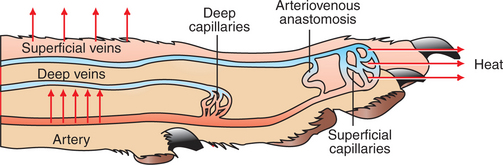
FIGURE 53-4 Representation of a limb showing the arterial supply and venous drainage by deep and superficial veins. Under warm conditions, blood perfuses the more superficial capillary beds, and heat is lost to the environment through the skin. Blood returns from these superficial vascular beds through the superficial veins, and this provides an additional source of heat loss. Under cold conditions, peripheral vasoconstriction occurs, and the blood flow to the limb is directed to the deeper vascular beds and returns to the trunk through the deep veins. Countercurrent heat exchange between the arteries and veins conserves body heat.
A similar countercurrent exchange of heat occurs in a carotid rete in sheep and some other ungulates. In this system the carotid artery forms a rete bathed in a sinus of venous blood that has drained the nasal cavity. The colder venous blood from the nose cools the arterial blood supplying the brain and protects the temperature of the brain. This mechanism becomes important during exercise, when the increase in ventilation aids in cooling the blood that drains from the nose. As a result, the arterial blood carrying heat from the exercising muscles is cooled before it enters the brain.
Some mammals, including humans and horses, do not possess a carotid rete and must rely on other thermoregulatory mechanisms to cool their brains during exercise. In the horse the guttural pouches may serve as such a mechanism. The guttural pouches contain air that is cooler than the arterial blood carried in the internal carotid artery. Because anatomically these guttural pouches surround the internal carotid arteries, heat is transferred from the blood to the air in the guttural pouches, thus protecting the brain from hyperthermia (Figure 53-5). In addition, the intracranial cavernous venous sinuses may assist in cooling the horse’s brain during exercise. This mechanism is thought to function in the same manner as the carotid rete, but less efficiently.
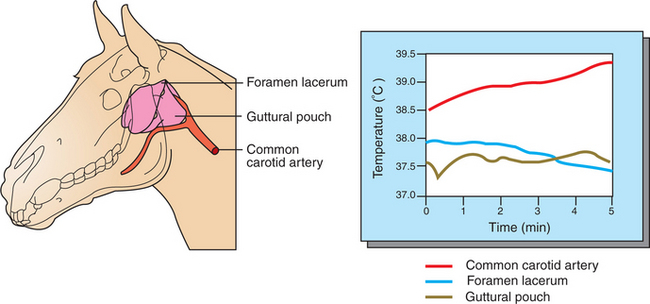
FIGURE 53-5 Guttural pouches cool the blood passing through the internal carotid artery on its way to the brain. Left, Anatomic arrangement of the guttural pouches and carotid arteries in the skull. The location of the temperature probes used to measure blood temperature is indicated. Right, Graph of blood and guttural pouch temperatures during a period of cantering. Note that even though the temperature of blood entering the guttural pouch in the common carotid artery increases with the duration of exercise, temperature at the foramen lacerum (where the internal carotid artery enters the cranium) decreases slightly.
HEAT EXCHANGE WITH THE ENVIRONMENT
Heat Loss by Convection Occurs When the Body Warms Air or Water
When the air or water in contact with the skin is heated, it flows away, thereby exposing the skin to cooler fluids. Because it takes more heat to warm water than it does to warm an equivalent mass of air, water-dwelling animals lose more heat by convection than do terrestrial mammals. The amount of heat lost by convection depends on the thermal gradient (temperature difference) between the skin of the animal and the fluid overlying the skin; a greater thermal gradient results in more heat loss. In natural convection the warmed air or water rises from the surface of the animal because it is less dense than the cooler fluid. In forced convection, cooler fluid is moved over the skin surface by a breeze or current or simply because the limbs and animal are moving. Forced convection is more effective than natural convection as a mode of heat loss because the thermal gradient is maintained by the constant renewal of the cooler air or water that blankets the surface of the skin. Young or small animals left in a cool draft can quickly lose much body heat by convection and should be protected from such situations.
The thermal gradient for heat loss can be altered by changes in skin blood flow and the amount of insulation separating the animal from the environment. Increasing blood flow to the skin raises skin temperature and therefore heat loss, whereas a reduction in skin blood flow reduces heat loss. Hair traps air and impairs convection. The thickness of the layer of hair can be altered by piloerection (making the hair stand up) and by growing a thicker hair coat in preparation for winter. The thick layer of blubber in sea mammals also provides a layer of insulation. Reducing the area of the exposed body surface area also reduces convective heat loss. Animals accomplish this by curling up in a ball or by huddling with littermates.
Heat Loss by Conduction Occurs When the Body Is in Contact with a Cooler Surface
Because animals usually do not lie on cool surfaces for long periods, conduction is not usually a major form of heat loss. In some situations, however, conductive heat loss can lead to hypothermia. A cold, stainless steel surgery table can form a heat sink for a small, anesthetized bird or mammal. Insulation or a heat source should be provided for such animals. Similarly, newborn piglets can lose much heat by lying on a cold concrete floor. Adult pigs cool themselves by conduction when they wallow in cool mud puddles.
Heat Loss by Radiation Occurs When Infrared Radiation Emitted by the Body Is Absorbed by Cooler Objects
All solid objects emit invisible electromagnetic radiation in the infrared range. Warm objects emit on a shorter wavelength and emit more emissions per unit time than do cool objects. When these emissions strike another object, some are absorbed and thus transfer heat. Although all objects emit radiant heat, the net heat transfer is from warm to cool objects. It is important to realize that radiant heat loss can occur even when a thermally neutral or warm environment surrounds the animal. Heat can be lost from an animal to the uninsulated walls of a building even though the intervening air is warm.
Heat Loss by Evaporation Occurs When the Water in Sweat, Saliva, and Respiratory Secretions Is Converted into Water Vapor
The evaporation of 1 L of water into water vapor requires 580 kilocalories (kcal). If the body provides this heat, evaporation can be a major form of heat loss. Some evaporative heat loss occurs continuously by the diffusion of water through the skin and by loss of water vapor from the respiratory tract. This water loss is obligatory, but under thermal stress, evaporative cooling can be increased greatly because sweat glands are activated or the animal begins to pant. Evaporative heat loss becomes increasingly important as the ambient temperature approaches body temperature; it is the only form of heat loss available once ambient temperature exceeds body temperature. The effectiveness of evaporation is reduced as the relative humidity increases, that is, as the air becomes more saturated with water vapor.
Sweating occurs from two types of coiled, tubular sweat glands located in the dermis. Apocrine glands produce a protein-containing secretion, whereas eccrine glands produce an aqueous secretion. All placental mammals except rodents and lagomorphs have sweat glands, but in the dog and pig these glands are poorly developed and of little use in thermoregulation. Apocrine glands produce the thermoregulatory sweat of hoofed animals, whereas in primates, sweat is produced by the eccrine glands. Secreted sweat has an ionic composition similar to that of plasma. As it passes to the skin surface along the duct, its composition is altered by the reabsorption of ions. If secretion rates are low, almost all the sodium and chloride, along with water, is absorbed. Therefore the sweat reaching the skin is a concentrated solution of urea, lactic acid, potassium ions, and, in the case of hoofed mammals, protein. When secretion rates are high, less sodium and chloride are absorbed, more water is lost, and the other constituents are consequently diluted. In hot environments, acclimatization increases the sweating rates, and because of increased secretion of aldosterone, most of the sodium and chloride is reabsorbed before the sweat reaches the skin. In most species, sweating is under the control of sympathetic cholinergic nerve fibers, but in the horse, control is through β2 adrenoceptors activated by catecholamines originating from sympathetic nerves or the adrenal medulla.
Panting is one mode of increasing evaporation from the respiratory tract. Small tidal volumes are moved at rapid frequency (200 breaths/min) over the respiratory dead-space. The rate of panting is close to the resonant frequency of the respiratory system, and thus the work of breathing is minimized and does not add to the heat load. In the panting animal, two mechanisms act to elevate heat loss through evaporation: (1) vascular engorgement of the respiratory and oral mucosa and (2) increased salivation. By ventilating primarily dead-space, severe hyperventilation and respiratory alkalosis are avoided. In birds, gular flutter is another method of increasing airflow over the respiratory dead-space. Even in mammals that do not pant, such as the horse, evaporative heat loss from the respiratory tract probably increases during prolonged exercise because dead-space ventilation increases.
Mammals vary in the relative importance of different modes of evaporative heat loss. In horses and cattle, sweating is the major form of evaporative heat loss. Sheep sweat, but panting is also of considerable importance. The dog relies almost totally on panting. Even small rodents, which neither pant nor sweat, increase evaporative heat loss by smearing saliva or water on their fur.
TEMPERATURE REGULATION
Mammals and Birds Regulate the Input and Output of Heat to Maintain Body Temperature Within a Narrow Limit
It is customary to measure body temperature as a first part of the clinical examination of mammals. This is because body temperature is maintained within fairly narrow limits despite large variations in ambient conditions. In diseased animals the ability to regulate temperature can be impaired, for example, by dehydration. In addition, infectious and other agents produce pyrogens that can cause an increase in body temperature. Table 53-2 lists the normal ranges of rectal temperatures in some common domestic mammals. The rectal temperature is somewhat lower than the core temperature of the animal, and changes in rectal temperature lag behind changes in core temperature. However, rectal temperature is a convenient measure in domestic mammals and provides a useful indication of core temperature.
Table 53-2 Rectal Temperature (in °C) of Domestic Mammals
| Species | Average | Range |
|---|---|---|
| Cat | 38.6 | 38.1-39.2 |
| Cattle (beef) | 38.3 | 36.7-39.1 |
| Cattle (dairy) | 38.6 | 38.0-39.3 |
| Dog | 38.9 | 37.9-39.9 |
| Donkey | 37.4 | 36.4-38.4 |
| Goat | 39.1 | 38.5-39.7 |
| Horse | 37.7 | 37.2-38.2 |
| Pig | 39.2 | 38.7-39.8 |
| Sheep | 39.1 | 38.5-39.9 |
In well-hydrated animals living in temperate climates, the range of normal temperature is quite narrow. Mammals living in hot, arid climates tolerate a wider range of temperature, allowing body temperature to decrease during the cool nights so that more heat can be absorbed during the ensuing hot day.
To maintain temperature within narrow limits, the animal must regulate its heat inputs and outputs. The inputs and outputs clearly cannot be equal at all times. During exercise, for example, heat production exceeds heat loss. Heat is stored in the body and then dissipates when exercise ceases. The specific heat of body tissues is similar to that of water; therefore, large amounts of heat can be stored without a potentially lethal increase in temperature.
Temperature-Sensitive Receptors Are Located in the Central Nervous System, the Skin, and Some Internal Organs
To regulate body temperature, the animal needs a variety of temperature sensors at various locations within the body. These sensors relay information to the brain, which then initiates mechanisms either to increase or to decrease heat loss or production.
Numerous heat-sensitive neurons are located in the preoptic area of the hypothalamus. These neurons increase their firing rate in response to minor increases in local temperature. In addition, experimentally warming this area immediately initiates heat-losing mechanisms, such as peripheral vasodilation and sweating. These observations suggest that this region of the brain may be the main center for temperature regulation. Other hypothalamic and midbrain neurons decrease their firing in response to heat, and still others increase firing in response to cold. All these temperature-sensitive neurons are monitoring brain or core temperature.
When an animal is exposed to a change in temperature, considerable heat loss or heat gain can occur before a change in core temperature occurs. Therefore, it is advantageous to have temperature-sensitive neurons located in the skin so that environmental temperature changes are detected before they threaten core temperature. The most numerous temperature-sensitive neurons in the skin respond to cold. If these receptors are activated, the body can initiate mechanisms for heat conservation and heat production before the core temperature decreases. Skin cold receptors are particularly sensitive to the rate of decrease in temperature. For this reason, shivering can occur after exercise as the skin is rapidly cooled by sweat evaporation, despite that the core temperature may be normal or slightly elevated. Skin receptors sensitive to heat also exist and can initiate heat loss when the skin temperature rises.
Temperature-sensitive neurons are also present at various locations in the viscera. Drinking large volumes of cold fluids may stimulate cold receptors in the gastrointestinal system, initiating body heat–conserving mechanisms.
Information from Central and Peripheral Heat-Sensitive Neurons Is Integrated in the Hypothalamus to Regulate Heat-Losing or Heat-Conserving Mechanisms
Figure 53-6 shows the feedback control mechanisms for the regulation of body temperature. Central integration of the information from various receptors occurs in the anterior hypothalamus. Information from central temperature receptors seems to predominate over information from skin and visceral receptors. For this reason, a rise in core temperature of only 0.5°C causes a sevenfold increase in the amount of skin blood flow; similarly, a modest decrease in core temperature initiates vasoconstriction and shivering. The effect of central receptors is about 20-fold greater than the effect of peripheral receptors.
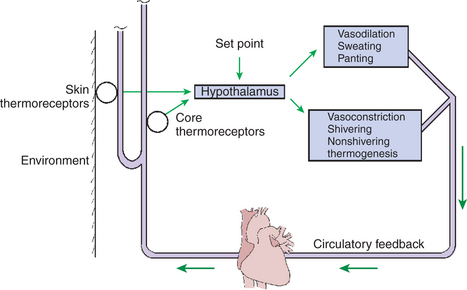
FIGURE 53-6 Feedback control mechanisms for the regulation of body temperature. Temperature receptors in the skin and the core relay information to the hypothalamus, which adjusts the responses either to conserve and produce or to lose heat. The results of these responses are relayed to the receptors through the circulatory feedback.
In the regulation of body temperature, the hypothalamus behaves as if it has a normal set point. When the core temperature rises above the set point, heat-losing mechanisms are initiated; when temperature decreases, heat conservation or production begins. Information from peripheral receptors modifies the set point, and thus shivering begins at a higher core temperature when the skin is cool than when it is warm. Similarly, sweating is initiated at a higher core temperature when the skin is cool than when it is warm.
INTEGRATED RESPONSES
The Responses to Heat Stress Are Peripheral Vasodilation and Increased Evaporative Cooling
For all mammals and birds, an environmental temperature exists at which body temperature can be maintained in a normal range, primarily by vasomotor mechanisms (Figure 53-7). This zone of thermoneutrality varies with the metabolic rate and the amount of insulation. The pig, which lacks fur, clearly has a higher thermoneutral temperature than does the sheep, which has thick wool. Dairy cattle that have high milk production produce so much metabolic heat that their thermoneutral zone is surprisingly low: 4° to 15°C (39°–59°F). In the thermoneutral zone, body temperature can be regulated by vasomotor mechanisms that increase or decrease skin blood flow and therefore change the amount of heat loss by convection and radiation.
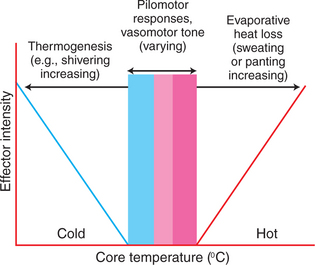
FIGURE 53-7 Relationship between the intensity of thermoregulatory responses and core temperature. The set point for temperature regulation is indicated by the pink bar zone. On either side of this set point is a zone where temperature can be maintained by vasomotor responses (blue and red zones). As the core temperature deviates more dramatically from the set point, there is a need either to increase thermogenesis during cold stress or to increase evaporative heat loss during heat stress.
(Modified from Bligh J: Temperature regulation in environmental physiology of animals. In Bligh J, Cloudsley-Thompson JL, MacDonald AG, editors: Environmental physiology of animals, Oxford, UK, 1976, Blackwell Scientific.)
When a homeotherm is exposed to heat stress, the initial response is vasodilation, which increases skin and limb blood flow. The resulting increase in skin temperature and the extension of core temperature down the limbs increase the temperature gradient between the skin and the environment, resulting in more heat loss by radiation and convection (see Figure 53-3).
If vasodilation alone is ineffective in maintaining a normal temperature, evaporative cooling is increased by sweating, panting, or both. Evaporative cooling is the only method of heat loss available once the environmental temperature exceeds the skin temperature and is most effective when relative humidity is low. Figure 53-8 shows that at −10°C (14°F), cows lose about 10% of their heat by evaporation, but as the ambient temperature rises to about 32°C (90°F), they lose 80% by evaporation. As relative humidity rises, animals have increasing difficulty losing heat; therefore, exercise in hot, humid conditions is likely to cause heat exhaustion. This was a major concern for horses competing in the 1996 Olympics in Athens, Georgia, where the high July temperatures were combined with high humidity.
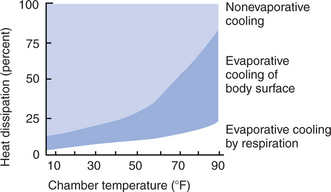
FIGURE 53-8 Methods of heat loss used by a cow as the environmental temperature increases. At low temperatures, most heat loss is by nonevaporative cooling (pale blue shading), but as the environmental temperature increases, the cow becomes increasingly dependent on evaporation (darker blue shading).
Animals also use behavioral methods to resist heat stress. These methods, which include seeking shade, standing in water, and wallowing in mud, are not available to intensively managed livestock. The producer must assume increased responsibility for the animals’ comfort and survival. Because high-producing dairy cattle have such a low thermoneutral temperature, their primary requirement is shade in hot climates, which is a greater concern than a source of heat or insulation in cold climates.
The Responses to Cold Stress Are Peripheral Vasoconstriction, Piloerection, and Increased Metabolic Heat Production by Shivering and Nonshivering Thermogenesis
As the ambient temperature decreases, homeotherms initially conserve heat by peripheral vasoconstriction. This sets up a temperature gradient along the limbs and reduces skin temperature, so there is only a narrow temperature gradient for radiation and convective heat loss (see Figure 53-3). Piloerection provides insulation and also decreases heat loss. Further cold stress initiates increases in metabolic heat production by shivering or nonshivering thermogenesis. All adult mammals can shiver, and neonates born in an advanced state of development, such as lambs and foals, can also shiver. Puppies and other less-developed neonates cannot shiver and rely on the warmth of mother and the nest to protect them from cooling. Brown fat is present in some of the latter neonates and in other small mammals and provides a source of nonshivering thermogenesis.
Chronic exposure of animals to cold results in increased secretion of thyroxine and an increase in basal metabolism, which increases basal heat production. When animals are housed where they receive natural light, the thickness of the hair coat increases at cold times of the year. Hair growth is the result of decreasing daylight as cold weather approaches.
Fever Is an Elevation of Body Temperature That Results from an Increase in the Thermoregulatory Set Point
Fever, also known as pyrexia, occurs in response to the elevation of an animal’s thermoregulatory set point and most often accompanies infectious diseases. Fever is believed to be an evolutionary adaptation to fight off infection and can be induced in some of the oldest species, such as reptiles and amphibians. Studies indicate that during infection, an increase in body temperature enhances leukocyte activity. This results in a decrease in animal morbidity and mortality from infections.
The induction of fever begins with production by many cell types of small polypeptides known as pyrogens (Figure 53-9). This occurs when infectious agents, toxins, or the lipopolysaccharide complexes in gram-negative bacteria invade the body. Pyrogens include interleukin-1 (IL-1; considered to be the most important), tumor necrosis factor (TNF), interleukin-6 (IL-6), interferon (IFN), and macrophage inflammatory protein (MIP). In addition, prostaglandins (PGs), products of the arachidonic acid cascade that are produced by endothelial cells, are major participants in the pathogenesis of fever. When released into the blood, pyrogens reach a part of the hypothalamus called the organa vasculosum of the lamina terminalis (OVLT). This region of the hypothalamus is highly vascularized, and the blood-brain barrier here is almost nonexistent, so endogenous pyrogens and PGs enter the brain easily from the bloodstream. Once in the hypothalamus, the endogenous pyrogens act on the endothelial cells to produce additional prostaglandin E2 (PGE2) and other arachidonic acid metabolites. These PGs cause the set point to rise.
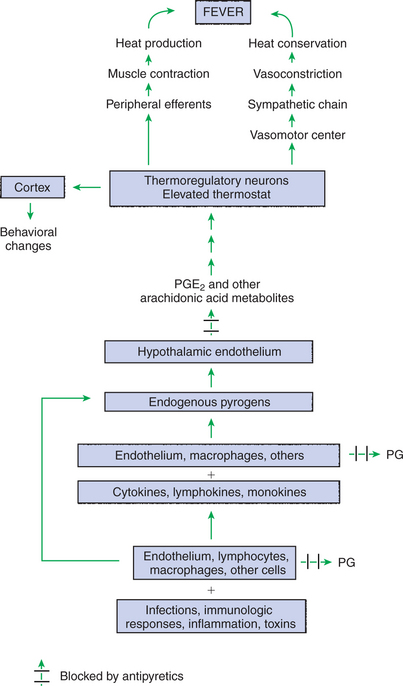
FIGURE 53-9 Peripheral and central mechanisms involved in the pathogenesis of fever. PGE2, Prostaglandin E2; PG, prostaglandins.
Once the set point increases, the animal initiates responses to conserve and produce heat until the body temperature reaches the new set point (Figure 53-10). Shivering, peripheral vasoconstriction, piloerection, and huddling behavior are all characteristic of the onset of fever. Once the new set point is reached, the animal maintains its body at the new temperature until the pyrogen is metabolized and production ceases. When this occurs, the set point decreases back to normal, and the animal initiates heat-losing mechanisms such as vasodilation and sweating to decrease body temperature. Because the local production of PGE2 in the hypothalamus is involved in increasing the set point, nonsteroidal antiinflammatory drugs (NSAIDs; e.g., aspirin, flunixin, ibuprofen) are used to treat fever. These antipyretic drugs block the enzyme cyclooxygenase, an integral enzyme in the arachidonic acid cascade, thus blocking prostaglandin production (see Figure 53-9).
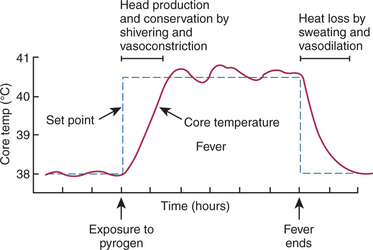
FIGURE 53-10 Events involved in fever. Exposure to a pyrogen increases the set point for the temperature-regulating system. This results in heat production and conservation to elevate body temperature, which in turn results in fever. When the fever ends, the set point decreases, and heat must be lost from the body through sweating and vasodilation.
HEAT STROKE, HYPOTHERMIA, AND FROSTBITE
Heat Stroke Occurs When Heat Production or Input Exceeds Heat Output, so Body Temperature Rises to Dangerous Levels
In hot, humid weather it is difficult for animals to lose heat because evaporative cooling cannot occur effectively. Strenuous exercise under these conditions can lead to a dangerous increase in body temperature. Similarly, when dogs are closed in cars in the sun, their panting saturates the air with water vapor, so further heat loss is impossible. As the body temperature rises, the metabolic rate increases, and more heat is produced. In addition, panting or sweating (or both) leads to dehydration and circulatory collapse, so it is more difficult to transfer heat to the skin. Once the body temperature exceeds 41.5° to 42.5°C (~107° −109° F), cellular function is seriously impaired and consciousness is lost.
Hypothermia Occurs When Heat Output Exceeds Heat Production, so Body Temperature Decreases to Dangerous Levels
Small or sick animals exposed to a cold environment may lose more heat than they can generate, and body temperature may decrease to a point at which heat-regulating mechanisms no longer work. The ability of the hypothalamus to regulate body temperature is greatly impaired below a temperature of 29° C (84° F). Cardiac arrest occurs at around 20° C (68° F). Neonates seem to be able to withstand cooling more than adults, and apparently comatose lambs, piglets, and puppies can be revived through warming.
Frostbite Occurs When Ice Crystals Form in the Tissues of the Extremities
In extremely cold conditions, when the extremities are vasoconstricted to conserve heat, the tissues may cool below the freezing point of tissue water. Ice crystals disrupt the tissue integrity, and gangrene can result. Normally, frostbite is prevented because vascular smooth muscle dilates in extreme cold, causing an inflow of warm blood. This mechanism apparently works adequately in animals that winter outdoors in northern climates.
CLINICAL CORRELATIONS
Influenza in Pigs
History.
You are called to examine a group of 3-month-old pigs in an intensively managed fattening house. The group of 20 pigs is in a pen, and there are multiple similar pens within the barn. In the last 2 days in this particular pen of pigs, the animals have been reluctant to eat and have started huddling together. The owner has observed that the outer pigs in the huddle continually try to burrow toward the center of the pile of pigs and that they appear to be trembling. At this time the remaining pigs in the barn are not affected. When you enter the barn and the pigs are disturbed, they begin sneezing and coughing, and some are reluctant to move.
Clinical Examination.
Three pigs are caught, and the rectal temperature is found to be 41°C (normal, 39.2°C). There is a nasal discharge, and the conjunctiva and nasal mucosa are congested. You treat the pigs with antibiotics, and over several days, the pigs recover; however, the disease spreads progressively through the remaining pens in the fattening house. All pigs show the same clinical signs, and no pigs die from the disease. Blood samples are taken for virus neutralization tests from the acutely affected pigs 2 weeks after they have recovered.
The diagnosis from the viral neutralization test is swine influenza, which has a high morbidity rate but a low mortality rate.
Comment.
The clinical signs produced by this disease are caused largely by the development of fever. The pigs that were examined had an elevated body temperature because the infection had raised the set point of their thermoregulatory centers to a high value. To raise body temperature to this new value, the pigs huddled together, and the pigs on the outside shivered to generate metabolic body heat. Once the infection is overcome and pyrogens are metabolized, the behavior of the pigs changes; they need to lose heat, so they separate and move around the pens more freely.
Heat Stroke in a Boston Terrier
History.
At 3 PM on a 95°F (35°C) day in August, you receive a frantic phone call from a client. The client went to a shopping mall and left her car parked in the lot. She had her Boston terrier with her, but because she thought she would only be a few minutes, she left the dog in the car. While in the mall, she was delayed by an uncooperative clerk when her debit card was rejected. When she returned to the car, her dog was prostrate with its tongue hanging out of its mouth and was unresponsive. You instruct the owner to bring the dog over to your practice immediately and to drive with the windows open for the half-mile trip.
Clinical Examination.
On arrival at the clinic, the dog is laid on the examining table, where it fails to respond to its name. Its mouth is open, its tongue is distended, and its mucous membranes are dry. Body temperature is 42.2°C (normal, 38.5°C).
From the history, the animal’s body temperature, and its lack of response, you diagnose heat stroke. The dog is placed in a bath of cool water, and fluids are administered intravenously. Within 5 to 10 minutes, the dog begins to look around and recognizes its owner. The water bath treatments are continued for 2 hours, at which time the body temperature is close to normal. The dog remains in the hospital overnight and is then discharged to the relieved owner the next day.
Comment.
The temperature inside a car parked in the hot sun rises rapidly to above body temperature. At this time, the only mechanism available for losing heat is evaporation of water from the respiratory tract, which the dog attempts by panting and salivating. For a short time this is an effective means of losing heat, but water vapor is transferred to the air in the car and progressively saturates the atmosphere with water. As the percentage saturation of the air increases, the animal has more and more difficulty achieving evaporation and therefore heat loss. Eventually the animal cannot lose heat, and the body temperature begins to rise. Once the body temperature exceeds 41.5° to 42.5°C, the animal loses consciousness. In addition, the panting results in dehydration and reduces the dog’s ability to deliver heat from the core of the body to the extremities. Brachycephalic dogs, such as Boston terriers, have an added disadvantage in temperature regulation: the short nose and convolutions in the wall of the pharynx increase the work of breathing, especially when the dogs pant. This increased work is an additional source of body heat, and the anatomy of the upper airway probably makes evaporative cooling less effective.
Therapy for this condition is to reduce body temperature and to restore circulatory function as rapidly as possible. For this reason, the dog is placed in a cool water bath to reduce body temperature and also receives intravenous fluids to rehydrate it by expanding its circulatory volume and restoring the ability of the circulation to redistribute heat within the body.
Eckert R, Randall D. Animal physiology: mechanisms and adaptations. New York: Freeman, 1983.
Hales JRS. The partition of respiratory ventilation of the panting ox. J Physiol. 1966;188:45.
Maugham RJ, Lindinger MI. Preparing for and competing in the heat: the human perspective. Equine Vet J Suppl. 1995;20:8.
Nadel ER. Regulation of body temperature. In: Boron WF, Boulpaep EL. Medical physiology: a cellular and molecular approach, updated edition. Philadelphia: Saunders, 2005.
Schmidt-Nielsen K. Animal physiology: adaptation and environment. Cambridge, UK: Cambridge University Press, 1997.
PRACTICE QUESTIONS
1. Sweating is an effective cooling mechanism because:
2. In the cold, animals both conserve and produce heat. Which of the following is a method of heat conservation?
3. Which of the following methods of heat loss can occur in an animal (body temperature = 38°C) standing in a room (temperature = 40° C) with relative humidity of zero? The walls of the room have a temperature of 30° C.
4. Which of the following describes thermoregulation?