DEVELOPMENT AND STRUCTURE OF CELLS AND TISSUES
After reading this chapter the student will be able to:
• describe the cell and how it divides
• discuss the origin of tissue, the ovarian cycle, and development of the embryonic disk
• describe the various tissues of the human body and some of the adverse factors such as environmental stress and hereditary and dietary factors that may affect development of these tissues
OVERVIEW
The smallest unit of structure in the human body is the cell, composed of a nucleus and cytoplasm. The nucleus contains deoxyribonucleic acid (DNA) and ribonucleic acid (RNA), the fundamental structures of life. The cytoplasm functions in absorption and cell duplication, in which organelles perform specific actions. The cell cycle is the time required for the DNA to duplicate before mitosis. This chapter discusses the four stages of mitosis: prophase, metaphase, anaphase, and telophase. Also described are the three periods of prenatal development: proliferative, embryonic, and fetal. The fertilization of the ovum in the distal uterine tube, zygote migration, and the zygote’s implantation in the uterine wall are discussed. In addition, the origin of human tissues—ectoderm, mesoderm, and endoderm—is presented, followed by the differentiation of tissue types, such as those of ectodermal origin, epithelium and skin with its derivatives, and the central and peripheral nervous systems. This chapter also delineates development of the mesodermal components involving connective tissues of the body, such as fibrous tissue, three types of cartilage, two types of bone, three kinds of muscles, and the cardiovascular system. The reader will better comprehend the origin, development, organization, and structure of the various cells and tissues of the human body.
CELL STRUCTURE AND FUNCTION
The human body is composed of cells, intercellular substance (the products of these cells), and fluid that bathes these tissues. Cells are the smallest living units capable of independent existence. They carry out the vital processes of absorption, assimilation, respiration, irritability, conductivity, growth, reproduction, and excretion. Cells vary in size, shape, structure, and function. Regardless of function, each cell has a number of characteristics in common with other cells, such as cytoplasm and a nucleus, which contains a nucleolus. However, some cell characteristics are related to function. A cell on the surface of the skin, for example, serves best as a thin, flattened disk, whereas a respiratory cell functions best as a cuboidal or columnar cell to facilitate adsorption with mobile cilia to move fluid from the lung to the oropharynx. Surrounding each cell is the intercellular material that provides the cell with nutrition, takes up waste products, and provides the body with form. It may be as soft as loose connective tissue or as hard as bone cartilage or teeth. Fluid, the third component of the body, is the blood and lymph that travel throughout the body in vessels or the tissue fluid that bathes each cell and fiber of the body.
Cell Nucleus
A nucleus is found in all cells except mature red blood cells and blood platelets. The nucleus is usually round to ovoid, depending on the cell’s shape. Ordinarily a cell has a single nucleus; however, it may be binucleate, as are cardiac muscle cells or parenchymal liver cells, or multinucleate, as are osteoclasts and skeletal muscle cells. The nucleus is important in the production of DNA and RNA. DNA contains the genetic information in the cell, and RNA carries information from the DNA to sites of actual protein synthesis, which are located in the cell cytoplasm. The nucleus is bound by a membrane, the nuclear envelope, which has openings at the nuclear pores. This envelope is composed of two phospholipids layers similar to the plasma membrane of the cell. The pores are associated with the endoplasmic reticulum that forms at the end of each cell division. The nucleus contains from one to four nucleoli, which are round, dense bodies constituting the RNA contained in the nucleus. Nucleoli have no limiting membrane (Fig. 1-1).
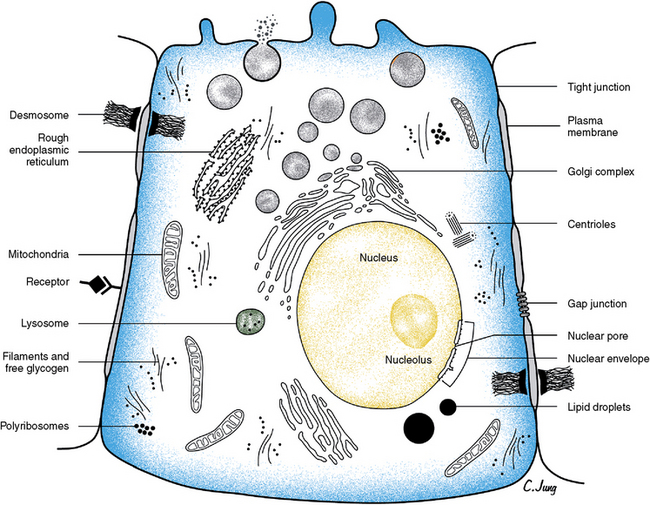
Cell Cytoplasm
Cytoplasm contains structures necessary for adsorption and for creation of cell products. The cytosol is the part of the cytoplasm that contains the organelles and solutes. The cytosol uses the raw materials brought into the cell to produce energy. It also functions in the excretion of waste products. These functions are carried out by the endoplasmic reticulum (ER)— parallel membrane-bound cavities in the cytoplasm that contain newly acquired and synthesized protein. Two types of ER, smooth surfaced and granular or rough surfaced, can be found in the same cell. Rough-surfaced ER is caused by ribosomes on the surface of the reticulum and is the site at which protein production is initiated. Proteins are vital to the cell’s metabolic processes, and each type of protein is composed of a number of different amino acids linked in a specific sequence. Amino acids form protein-containing groups, which, in turn, form acids or bases.
Ribosomes are particles that translate genetic codes for proteins and activate mechanisms for their production. They can be found as separate particles in the cytoplasm, clustered, or attached to the ER membranes. Ribosomes are nonspecific as to what type of protein they synthesize. The type is dependent on the messenger RNA (mRNA), which carries the message directly from the DNA of the nucleus to the RNA in the ER. This molecule attaches to the ribosomes and gives orders about the formation of the amino acids.
The ER transports substances in the cytoplasm. The ER is connected to the Golgi apparatus via small vesicles. The Golgi apparatus or complex helps sort, condense, package, and deliver proteins arriving from the ER. The Golgi apparatus is composed of cisternae (flat plates) or saccules, small vesicles, and large vacuoles. From here the secretory vesicles move or flow to the cell surface, where they fuse with the cell membrane and the plasmalemma and release their contents by exocytosis.
Lysosomes are small, membrane-bound bodies that contain a variety of acid hydrolase and digestive enzymes to help break down substances both inside and outside the cell. They are in all cells except red blood cells but are prominent in macrophages and leukocytes.
Mitochondria are membrane-bound organelles that lie free in the cytoplasm and are present in all cells. They are important in generating energy, are a major source of adenosine triphosphate (ATP), and therefore are the site of many metabolic reactions. These organelles appear as spheres, rods, ovoids, or threadlike bodies. Usually the inner layer of their trilaminar bounding membrane inflects to form transverse-appearing plates, the cristae (see Fig. 1-1). Mitochondria lie adjacent to the area that requires their energy production.
Microtubules are small tubular structures in the cytoplasm that are composed of the protein tubulin. These structures may appear as singles, as doublets, or as triplets. They probably function as structural and force-generating elements and relate to cilia (motile cell processes) and to centrioles in relation to mitosis. They have cytoskeletal functions in maintaining cell shape. Centrioles are short cylinders appearing near the nucleus. Their walls are composed of nine triplets of microtubules. Centrioles are microtubule-generating centers and are important in mitosis, self-replicating before mitosis begins.
Surrounding the cell is the plasma membrane or plasmalemma, which envelops the cell and provides a selective barrier that regulates transport of substances into and out of the cell. All membranes are composed mainly of lipids and proteins with a small amount of carbohydrates. The plasma membrane also receives signals from hormones and neurotransmitters. In addition, cells contain proteins, lipids, or fatty substances that provide energy in the cell and are important components of cell membranes and permeability. Carbohydrates are also important in cells as the most available energy component in the body. These carbohydrates may exist as polysaccharide-protein complexes, glycoprotein complexes, glycoproteins, and glycolipids. Carbohydrate compounds are important in cell function and for development of cell products, such as supportive tissues and body lubricants.
Genetic mechanisms help a cell to develop and maintain a high degree of order. The ability is dependent on the genetic information that is expressed within the cell. The basic genetic processes in the cell are RNA and protein synthesis, DNA repair, and replication and genetic recombination. These processes produce type proteins and nucleic acids of a cell. These genetic events are relatively simple compared with other cell processes.
CELL DIVISION
Cell division is a continuous series of discrete steps by which the cell component divides. This function is related to the need for growth or replacement of tissues and is partly dependent on the length of the cell’s life. Continually renewing cells line the gastrointestinal tract and compose the epidermis and the bone marrow. A second type of cell is part of an expanding population—the cells of the kidney, liver, and some glands. The third type of cell does not undergo cell division or DNA synthesis. An example is the neurons of the adult nervous system. For a somatic cell to undergo cell division, it must pass through a cell cycle, which ensures time for DNA genetic material in the daughter cells to duplicate that of the parent cell. However, in a sex cell, ovum, or spermatozoon, the process of meiosis occurs, in which a reduction division of chromosomes in the daughter cell takes place. The result is that half as many chromosomes are in the daughter cell as are in the parent cell. Through meiosis, after fertilization of the ovum by the male chromosomes, the original (diploid) number of chromosomes is regained. The duration of the cell cycle in somatic cells is now known (Fig. 1-2). After mitosis, the cells enter the reduplication or G1 phase of the interphase, the initial resting stage. This is followed by the S phase, in which DNA synthesis is completed. Next the cell enters the G2 phase or quiescent phase of the post-DNA duplication and proceeds into the mitotic stages of prophase, metaphase, anaphase, and telophase (Fig. 1-3). The cell then reenters and remains in the interphase stage until duplication resumes the mitotic process of developing two daughter cells identical to the parent cells.
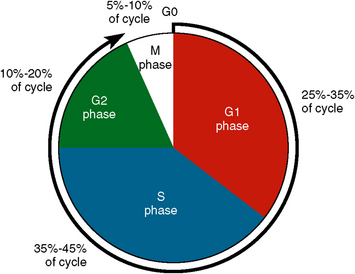
Fig. 1-2 Periods of cell cycle indicate relative time needed for each phase. G1 is the reduplication phase, or resting phase, which takes about 6 to 8 hours. In the S phase, DNA duplication takes place in 8 to 10 hours. The G2 phase is the postduplication phase, which takes about 4 to 6 hours. In the M phase, mitosis takes about 35 to 40 minutes. These figures are for cultured mammalian cells. The total is 18 to 24 hours for these four stages of cytokinesis. Other types of cells can have a longer or shorter cell cycle.
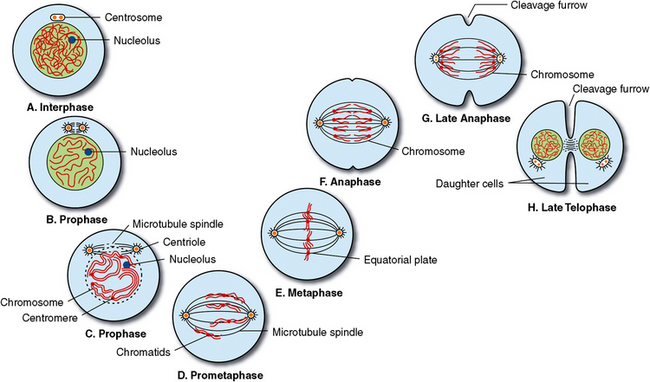
Fig. 1-3 Mitosis of somatic cell. The continuous process of cell division is shown. Mitosis is replication of parent chromosomes and distribution of two sets of chromosomes into two separate and equal nuclei. Stages are as follows: A, Interphase, resting cell. B and C, During prophase, chromatin thread shortens and thickens and becomes chromosomes, which then split into pairs of chromatids. Nuclear membrane disappears, and centrioles appear and begin migration to opposite poles of cell. D, In prometaphase, or early metaphase, chromatid pairs attach to centromere and line up in equatorial plate of cell. E, Metaphase occurs when centromeres and chromatids line up in middle of cell. Centrioles are at opposite ends of cell and attach to chromosomes by mitotic spindles. F, Anaphase is a division and movement of completed identical sets of chromatids (chromosomes) to opposite ends of cells. G, In late anaphase, identical sets of chromosomes have reached opposite ends of the cells as cleavage begins. H, In telophase, a nuclear membrane reappears, nucleoli appear, and chromosomes lengthen and form chromatin thread. Mitotic spindles disappear, and centrioles duplicate so that each cell has completely identical properties.
 CLINICAL COMMENT
CLINICAL COMMENTMitosis
Before mitosis the cell exists in the interphase, as seen in Figure 1-3, A. The first step of mitosis is prophase, in which four structural changes occur (Fig. 1-3, B). The chromatin thread of the nucleus thickens into rodlike structures called chromosomes. Each chromosome then splits, forming two chromatids. These chromatids line up along the central area of the cell, called the equatorial plate. Each chromatid pair is attached to a spherical body called a centromere. The centriole pair duplicates, and the chromatids accompany the centrioles’ migration to the opposite ends of the cell. Those fibers not formed between the migrating centrioles are spindle fibers and those that form around the centrioles are astral rays or asters (Fig. 1-3, C). At this time the nucleolus disappears and its components become attached to the chromatids. Finally, the nuclear envelope breaks down and changes into granular elements, such as the ER (Fig. 1-3, D).
Chromatids have moved to the cell center by the metaphase stage. They are arranged along an equatorial plate at right angles to the long axis of the spindle (Fig. 1-3, E). The two chromatids of each chromosome become attached centrally at the equatorial plate to a centromere. These chromatids then split at the centromere into two sets of chromosomes.
In anaphase, the daughter chromosomes move to the opposite poles of the cell with the full complement of 46 at each end (Fig. 1-3, F and G). This is thought to occur by movement of the chromosomal microtubules that attract the chromatids toward the poles. A constriction begins to appear around the midbody of the cell (Fig. 1-3, G).
In telophase, the chromosomes detach from the chromosomal microtubules and the microtubules disintegrate. The chromosomes next elongate and disperse, losing their identity and regaining the chromatin thread appearance. Both the nucleoli within the nucleus and the nuclear envelope then reappear. As each nucleus matures, the cleavage furrow deepens in the midcell until the two daughter cells separate (Fig. 1-3, H).
Meiosis
Meiosis is the process of reduction of the number of chromosomes to half the normal number in the germ cells to allow fusion of the male and female germ cells. There are two cell divisions in meiosis. In the first meiotic division, the chromosomes divide equally with pairing of the homologous chromosomes and the appropriate synthesis of DNA. In the second meiotic division, the DNA is not synthesized, and three of the daughter cells divide into polar bodies that become inactive; the one remaining germ cell containing half the amount of DNA pairs with the germ cell of the opposite sex. This pairing of the XY chromosomes of the male and female germ cells provides the needed mature somatic cell.
Apoptosis
Apoptosis, or programmed cell death, is the fragmentation of a cell into membrane bound particles, which are then eliminated by phagocytosis by specialized cells. Cell death is the usual accompaniment of embryonic growth and differentiation. It is a means of eliminating transient and obsolete tissues. Thus, cell death, as well as histogenesis and morphogenetic movement, accomplishes the final form of the structure. Cell death typically occurs at sites during folding or invagination of tissues. Cell death is a useful way of eliminating tissues or organs that provided a function during early embryonic life, for example the tadpole tail and gills.
Adult stem cells (Fig. 1-4) are found in hematopoietic cells in bone marrow and have the multipotent capacity to form a number of cell types. Stem cells have been found in the dental pulp as well as the brain, muscle, skin, intestinal tract, and blood vessels. It is the hope of the future that these cells will be able to replace damaged, dead, or malfunctioning tissue. It has recently been reported that damaged corneal cells of the eye can be replaced with bits of oral epithelium utilizing the patient’s own stem cells to aid in the healing process and in restoring vision.
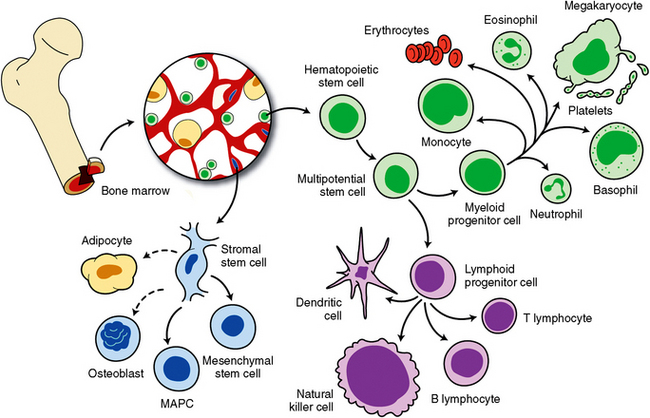
Fig. 1-4 Stem cells in the bone marrow (hematopoietic) have been studied extensively. These cells can differentiate into blood and immune cell lines. Other stem cells in the bone marrow are stromal stem cells, and they have been reported to be able to differentiate into fat and bone cell precursors. Other stem cells have been discovered in the brain, eyes, skin, muscle, dental pulp, blood vessels, and gastrointestinal tract.
All cells have a limited lifetime. For example, the life span of a white blood cell is only a few hours to a few days. Red blood cells live approximately 120 days before they are ingested by macrophages. Surface-covering cells—such as those of the skin, hair, or nails—renew as they are replaced as do cells lining the respiratory, urinary, and gastrointestinal tracts. Other cells in the body—such as those of the liver, kidneys, and thyroid gland—do not normally renew after maturity unless they are injured.
ORIGIN OF HUMAN TISSUE
Epithelial Mesenchymal Interaction
The following are several definitions that are important to understanding the basic processes of early development.
Induction
Induction is the process in which an undifferentiated cell is instructed by specific organizers to produce a morphogenic effect.
Cell Differentiation
The organizer is the part of an embryo that influences another part to direct histologic and morphologic differentiation. Chemical substances called growth factors induce cells to initiate specific cellular processes including DNA synthesis in a specific temporal and spatial manner.
Periods of Prenatal Development
Implantation and enlargement of the blastocyst, which contains the embryonic tissue, occur rapidly in the proliferative period, which lasts for 2 weeks. During this time, fertilization, implantation, and formation of the embryonic disk take place. After the second week, this mass of cells begins to take the form of an embryo, so the period of 2 to 8 weeks is termed the embryonic period. During this period, the different types of tissue develop and organize to form organ systems. The heart forms and begins to beat by the fourth week, and the face and oral structures develop during weeks 4 to 7. The embryo takes on a more human appearance in the eighth week and moves into the fetal period, which extends until birth (Fig. 1-5). During this period, the tissues that developed in the embryonic stage enlarge, differentiate, and become capable of function.
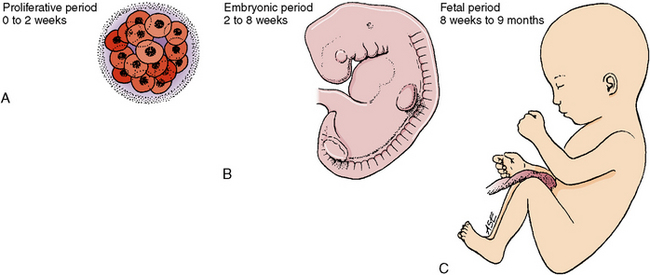
Fig. 1-5 The developing human passes through three periods of growth. A, Proliferative period: the first 2 weeks when cell division is prevalent. B, Embryonic period: from the second to the eighth weeks. C, Fetal period: from the eighth week to birth.
 Consider the Patient
Consider the PatientOvarian Cycle, Fertilization, Implantation, and Development of the Embryonic Disk
The origin of tissue begins with fertilization of the egg, or ovum, which occurs when sperm contact the egg in the distal part of the uterine tube (Fig. 1-6). The fertilized egg then grows and is termed the zygote. The cell mass produces a ball of cells (the morula) in the uterine tube. The morula grows and begins migration medially to the uterus, which it reaches at the end of the first week. The uterine cavity meanwhile prepares for the arrival of the fertilized ovum. The uterine lining (endometrium) thickens, and capillaries and glands develop to nourish the ovum. Estrogen and progesterone control this cyclical event (Fig. 1-7). The morula increases in size and is termed a blastocyst. As the blastocyst swells, it becomes hollow and develops a small inner cell mass. When this blastocyst or zygote reaches the uterine cavity, it attaches to the sticky wall of the uterus and becomes embedded in its surface. The cells of the zygote digest the uterine endometrium, permitting deeper penetration. This process is known as implantation. If no fertilized ovum reaches the uterine cavity, the development of capillaries and glands is terminated by menstruation (Fig. 1-8).
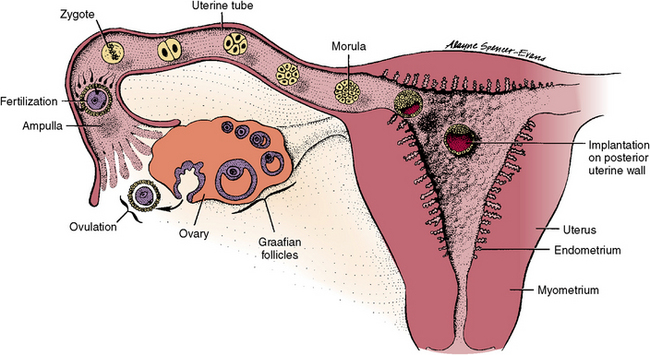
Fig. 1-6 Schematic diagram of the uterus and uterine tubes reveals the path of sperm to the distal tube, in which fertilization of the newly appearing ovum from the adjacent ovary occurs. The resultant zygote travels to uterus while undergoing cleavage, and implantation occurs on seventh day after conception.
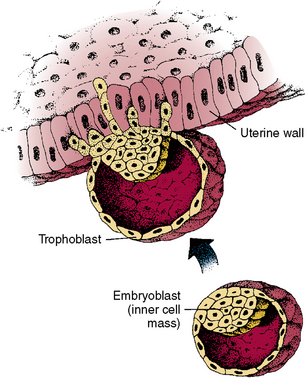
Fig. 1-7 Implantation of a fertilized ovum (zygote) in wall of uterus. Outer cells of trophoblast digest uterine cells to implant. An embryoblast develops within cell mass. As the mass expands, a surrounding cavity is formed.
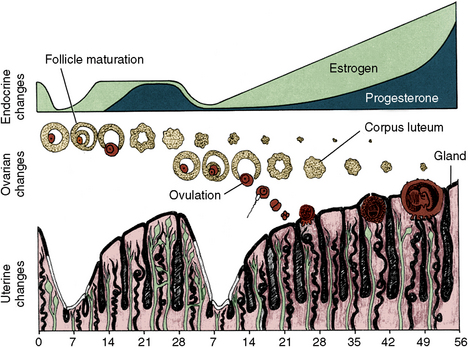
Fig. 1-8 Cyclical events of ovulatory cycle. Top, Endocrine changes: ovulation is controlled by estrogen and progesterone. Center, Ovarian changes: the ovum matures, is expelled from ovary on fourteenth day, and if fertilized, becomes implanted in uterine wall 7 days later. Bottom, Uterine changes: uterine wall thickens and prepares for implantation each month. If implantation does not occur, uterine wall erodes with loss of blood vessels and gland ducts (menstruation).
Two small cavities develop on either side of the inner cell mass. They reach each other in the center, where a small disk (the embryonic disk) is formed (Fig. 1-9). The embryonic disk becomes the embryo, composed of the common walls of the two adjacent sacs. One sac is lined with ectodermal cells, which will form the outer body covering (epithelium). The other sac is lined with endodermal cells. On the dorsal surface of the embryonic disk, the ectoderm forms the neural plate, whose lateral boundaries elevate to form a neural tube that will become the brain and spinal cord (Fig. 1-10). The endodermal cells also form a tube, which will become the gastrointestinal tract. As this tube elongates, it anteriorly develops outpouchings that form the pharyngeal pouches, lung buds, liver, gallbladder, pancreas, and urinary bladder (Fig. 1-11).
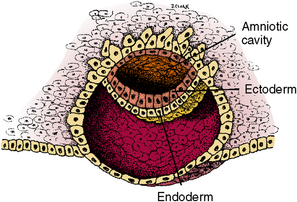
Fig. 1-9 Second small cavity lined with ectoderm develops (amniotic cavity). The other cavity (yolk sac) is lined with endoderm. The two cell layers contact in the center to form an area of ectoderm and endoderm for embryonic disk.
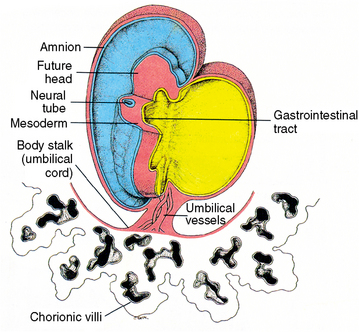
Fig. 1-10 A 3-week human embryo, viewed from the ventral-lateral aspect, illustrating an elongating gastrointestinal tube and a dorsally located neural tube.
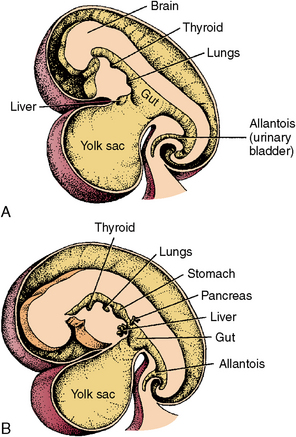
Fig. 1-11 Further development of the gastrointestinal tract. A, At 4½ weeks and B, at 5 weeks. Outpouchings of the intestinal tube form gastrointestinal organs.
Next, cells develop between the ectodermal and endodermal layers in the embryonic disk. This area becomes the mesodermal layer. These cells will develop into the muscles, skeleton, and blood cells of the embryo (Fig. 1-12). Mesodermal cells also accompany the elongating digestive tube and support its walls with muscle growth. This enables function and assists in the formation of organs arising from the developing gastrointestinal tube. From these three layers—ectoderm, mesoderm, and endoderm—develop all tissues of the body, as well as the complex organs (see Fig. 1-12).
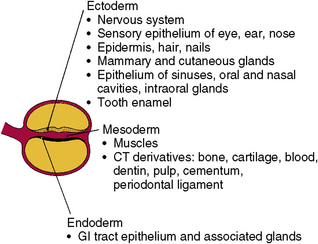
Environmental teratogens may affect the development of normal cells, tissues, organs, or organ systems. A defect in the development of a group of cells is considerably less damaging than a defect in an organ or organ system. The smaller and less complex the development, the less extensive the problem created. Development is also related to timing. Tissues are most susceptible to defective development when they begin to differentiate in the embryonic period (2 to 8 weeks).
DEVELOPMENT OF HUMAN TISSUES
The skin is a dual organ that has an epidermis, a surface cell layer that develops from the surface of ectodermal cells, and a dermis, which arises from the underlying mesoderm. The dermis originates in the somites, the masses of mesoderm that lie on either side of the neural tube. From this mesoderm come both the dermis of the epithelium and the visceral mesoderm that covers the yolk sac and later becomes the gastrointestinal tract (Fig. 1-13). Therefore, all the muscles functioning in peristalsis of the intestines arise from this mesoderm.
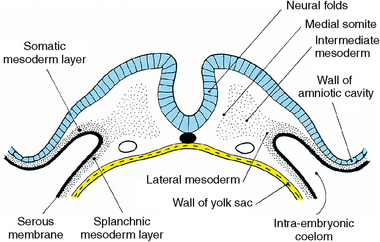
Fig. 1-13 Neural folds and somites in transverse section at approximately 20 days after conception. Medial somite (mesoderm) forms the axial skeleton that surrounds neural tube. Intermediate mesoderm forms striated muscle of body, and lateral mesoderm forms dermis of the epithelium of the body wall (somatic) and gastrointestinal tract (splanchnic).
Initially, the embryo is covered with a single layer of ectodermal cells (Fig. 1-14, A). By 11 to 12 weeks, this ectodermal layer of epithelium thickens into four layers. From the basal layer of cells come the more superficial cells of the epithelium (Fig. 1-14, B). Later, melanocytes invade and pigment the skin. At birth, the skin may show varying degrees of keratinization. Hair, teeth, nails, and mammary, sebaceous, and salivary glands all develop from a combination of epidermal and dermal cells. This development occurs when epithelial cells proliferate, invade the underlying dermis, and finally differentiate into glands or teeth, with both the epidermis and dermis contributing to each of these structures.
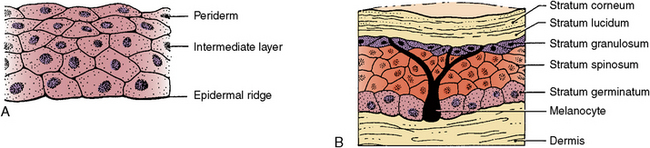
Fig. 1-14 Development of the skin. A, At 4 weeks and B, at 36 weeks. Initial layer of epithelial cells thickens into multiple layers, and underlying connective tissue becomes dermis. Dermis and epithelium combine to become skin.
Epithelial-mesenchymal interactions are the necessary interactions of an epithelium and underlying mesenchyme that determine the terminally differentiated tissue. There are many examples of this process including tooth and salivary gland induction and differentiation during development.
Nervous System
The neural folds appear during the third prenatal week. The lateral edges of the neural plate begin to elevate as folds arising dorsally (see Fig. 1-9). These folds represent the first change in shape of the embryo’s body from the flat sheet of cells (see Fig. 1-8). These folds reach the midline, first in the cervical region, and then the neural tube closes both anteriorly and posteriorly (Fig. 1-15). When the anterior tube closes, it shows three dilations that form the primary brain vesicles, the forebrain, midbrain, and hindbrain (Fig. 1-16, A). The neural tube bends forward just behind the midbrain and backward behind the hindbrain (Fig. 1-16, C and D). The cerebral hemispheres develop from the forebrain vesicles. The midbrain is a pathway from the cerebral cortex to centers in the pons and cerebellum of the hindbrain. The fifth cranial nerve develops in the midbrain (Fig. 1-16, B to D). The cerebral hemispheres of the forebrain develop into the frontal, temporal, and occipital lobes.
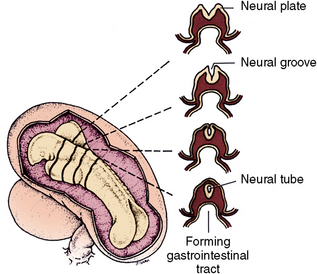
Fig. 1-15 Left, Dorsal view of closing neural tube of 3-week human embryo. Closure occurs initially in the dorsal area, then anteriorly and posteriorly. Right, Transverse sections of neural folds appear anteriorly, and those of closed neural tube are in the midbrain region.
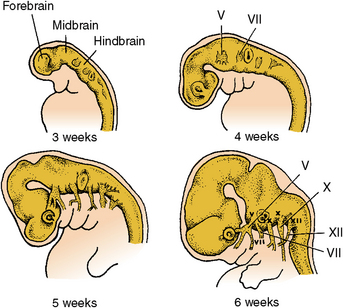
Fig. 1-16 Development of cranial nerves. A, 3 weeks; B, 4 weeks; C, 5 weeks; D, 6 weeks. At 3 weeks, the forebrain has enlarged, and sensory vesicles are laterally located. At 4 and 5 weeks, the forebrain has bent forward, and cranial nerves have grown into tissues they innervate. At 6 weeks, the anterior brain has enlarged and bent back on the posteriorly located cerebellum.
Cranial nerves
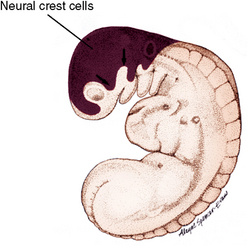
The ventricles of the brain are continuous and connect posteriorly with the spinal cord. The walls of the neural tube are lined with neuroepithelium. As these cells proliferate, they differentiate into neuroblasts and become the white and gray matter of the spinal cord. Neuroblasts are primitive nerve cells that develop into adult nerve cells called the neurons. These cells do not divide further. Along the surface of the developing brain and spinal cord, neural crest cells form the sensory system of the dorsal root ganglia of the cranial and spinal nerves (Fig. 1-17). The neural crest cells also contribute to tissues of the face, such as cartilage, muscles, teeth, and ligaments.
Connective Tissue
Connective tissue develops from the somites as fibro-blasts migrating from either side of the neural tube (see Fig. 1-13). Early in formation, the ventromedial part of the somite differentiates into the sclerotome, the dorsolateral part becomes the dermatome, and a third division becomes the intermediate mesoderm or myotome. The medial sclerotome differentiates into mesenchymal cells, which become osteoblasts, chondroblasts, and fibroblasts. A large part of the embryonic skeleton develops from these cells. Dermatome cells form the dermis, the subcutaneous tissue, and the visceral mesoderm, which supports the endoderm of the gastrointestinal tract, as well as a system of mesenteries that stabilize and support the gastrointestinal tract (Fig. 1-18). Also, connective tissue arises from the somites, providing supporting connective tissues, bones, cartilage, tendons, and ligaments. The tendons connect the muscles to the skeleton as they develop. Connective tissue also functions as capsules of glands and the supporting tissues within them.
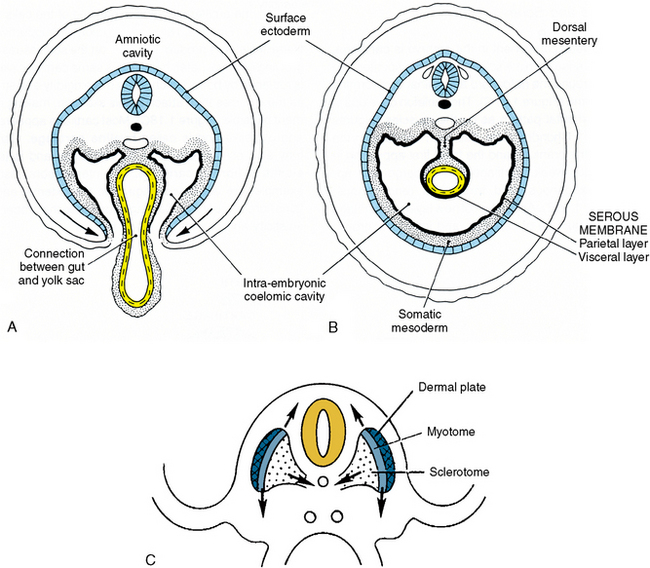
Fig. 1-18 Cross sections of embryo. A and B illustrate the yolk sac’s role in development of the gastrointestinal tube. The developing body wall is growing ventrally, closing the ventral opening. C, Contributions of somite to skin, muscles, and cartilage. Cartilage forms a support for the spinal column (sclerotome), which surrounds neural tube. Contribution of somitic mesoderm (dermal plate) to the body wall seen in B. Muscles arise from intermediate mesoderm (myotome).
Blood and lymphatic tissues
Blood is a specialized connective tissue that is composed of seven liters of fluid and cells in the body. The blood contains formed elements that are the red blood cells or erythrocytes, white blood cells or leukocytes, and blood platelets suspended in a liquid termed plasma. The red blood cells are most numerous (5 × 103 per mm3), and they carry oxygen from the lungs by means of a substance termed hemoglobin and also carry carbon dioxide from the cells of the tissue to the lungs by both the hemoglobin of the red blood cell and the plasma of the blood. Thus, blood is a pathway for conducting blood cells throughout the body. The white blood cells or leukocytes are few compared with the red (6,500 to 10,000/ml) and function in defending the body against bacteria. The leukocytes only travel in the blood vessels from their site of origin to the area of infection where they leave the blood vessel, migrating between the endothelial cells to travel in tissue spaces to the site of infection. Three types of granulocytes exist: neutrophils, eosinophils, and basophils, and two types of granulocytes and agranulocytes: lymphocytes and monocytes. The neutrophils (polymorphonuclear leukocytes) are the most numerous of the white blood cells, representing 60% to 70%, and function in destroying bacteria that invade the tissue spaces. The platelets are small, disk-shaped cell fragments carried in the blood and originate from megakaryocytes in the marrow spaces. There are 300,000 to 350,000 platelets in 1 mm3 of blood; they function to limit hemorrhage to the endothelium of the vessel.
The lymphatic system is composed of the lymph nodes, thymus and the spleen as well as the vessels that carry the lymph throughout the body. The lymphatic system is a protective mechanism as the immunologic defense of the body. The lymphoid system destroys bacteria, viruses, and invasive microorganisms. The lymphatics are made up of the innate and the adaptive immune systems. The cells that constitute the innate and adaptive immune systems are the B cells, the T cells, the NK cells, and macrophages, and they are all formed in the bone marrow. The T cells migrate to the thymus to become immunocompetent. The thymus consists of a cortex and medulla and is composed of epithelia and reticular cells and macrophages. The medulla consists primarily of thymocytes that are immunocompetent T cells. Throughout the lymphatic vascular system are lymph nodes that act as filters for all bacteria or substances foreign to the body. The lymph nodes are composed of a cortex and a medulla, the cortex is composed of lymph nodules and the medulla is composed of lymph sinuses interposed with cords of lymph cells. The spleen is the other lymphatic organ and is composed of a cortex and the hilum where the blood vessels enter and exit. The spleen functions in T- and B-cell formation and also in blood formation if the need arises.
Cartilage and bone
The initial skeletal component in the embryo is cartilage. Cartilage cells arise from the sclerotome and migrate to surround the notochord and spinal cord, which form the spinal column (see Fig. 1-18, C). The skeleton develops in the same segmental pattern as the muscles do (Figs. 1-19 and 1-20). Chondroblasts also form cartilage in the appendages, the cranium, and the face, which first appear in the fifth prenatal week. Cartilage cells undergo both appositional (exogenous) and interstitial (endogenous) growth (Fig. 1-19, B).
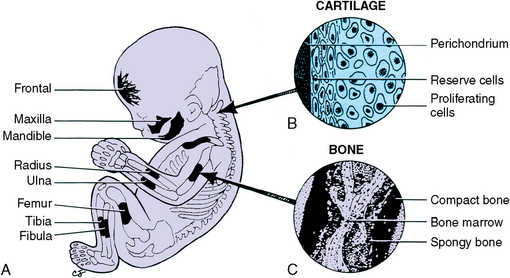
Fig. 1-19 Embryo’s skeleton (A) illustrating development of cartilage and bones. B, Cartilage development by both surface apposition and internal interstitial growth. C, Endochondral bone development in the shaft of a long bone.
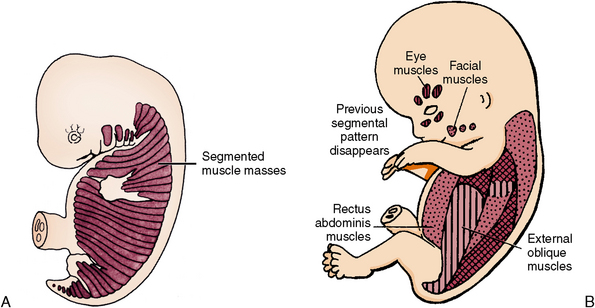
Fig. 1-20 A, Primitive myotome in skeletal muscle formation in an embryo. B, Differentiation of skeletal muscle by enlargement of fibers and attachment to bony skeleton to become functional units. The previous segmental pattern disappears.
Apposition of new layers of cartilage occurs on the surface of cartilage, and interstitial growth involves the proliferation and expansion of the cells within the matrix (Fig. 1-19, B). A supportive cartilage skeleton is produced rapidly to support the soft tissues of the growing embryo. Later, most of this same cartilage skeleton is replaced by bone, which offers more rigidity and strength as muscles attach to it, making movement possible (Fig. 1-19, C). Most cartilage appears clear and glasslike and is called hyaline cartilage. Cartilage may also contain elastic fibers and be termed elastic or fibrous cartilage. The intervertebral disks, for example, are fibrous cartilage, but the external ear contains elastic cartilage. Cartilage combines the properties of elasticity and strength.
Bone replaces cartilage by a process termed endochondral bone development (Fig. 1-21). In this case a small blood vessel enters the cartilage shaft (diaphysis), the cartilage calcifies and disintegrates in the center, and a marrow space is formed (Fig. 1-21, B). New bone develops on the surface of cartilage spicules that border the marrow space (Fig. 1-21, C). Small blood vessels enter the head of the long bones, and secondary ossification centers appear, repeating the process that took place in the shaft of the long bone (Fig. 1-21, D). During the growth period a developing cartilage disk remains in the neck of each long bone and bone forms on either side. This disk is known as the epiphyseal line and will remain as long as the bone is forming. The wider part of the diaphysis adjacent to the epiphyseal line is known as the metaphysis (Fig. 1-21, D). Cartilage develops and expands by interstitial growth, which is growth within the cartilage matrix by each cartilage cell enlarging and forming matrix around each cell. New bone forms along the cartilage margins of the epiphyseal line. After bone replaces the epiphysis, cartilage is limited to covering the heads of long bones, the nasal septum, the ears, and a few other sites.
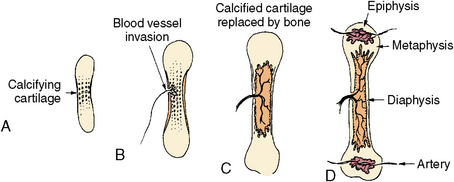
Fig. 1-21 Schematic diagram of endochondral ossification as seen in developing long bones of the body. A, Original hyaline cartilage is calcified in the center of the diaphysis. B, A blood vessel invades the center of the shaft. C, Marrow space appears in the center of the shaft, and bone forms around the diaphysis. D, Bone formation continues in the shaft, and secondary ossification sites appear in the heads (epiphysis) of the bones. A disk of cartilage remains between bone forming in the head and the shaft (epiphyseal line).
Direct transformation of connective tissue into bone may also take place. In this case, collagen fibers of connective tissue organize into closely knit meshwork, and this matrix gradually calcifies into bone by a process termed intramembranous bone formation or membranous bone formation (Fig. 1-22). It is much simpler for bone cells to organize in this manner and to form spicules of bone through coalescence with neighboring spicules until a bony plate is formed. The bones of the face and cranium develop in this manner.
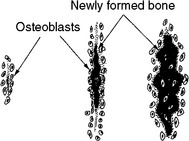
Fig. 1-22 Membranous bone formation that takes place in connective tissue. Initial membranous sites grow by apposition of new bone on their surfaces.
DNA transcription is an example of gene expression. Transcription generates mRNAs that carry information for protein synthesis, as well as transferring ribosomal and other RNA molecules that have structural and catalytic functions. RNA molecules synthesize RNA polymerase enzymes, which make an RNA copy of a DNA sequence.
Muscle
By the tenth prenatal week, muscle cells (myoblasts) have begun migrating from the myotome, following a segmental pattern similar to that of the bony skeleton (see Figs. 1-19 and 1-20). They gradually differentiate into elongated, multinucleated muscle fibers, which are specialized cells with the property of contractility. In this manner, muscle is able to provide motion on the basis of structural and functional characteristics.
Muscle is divided into three types: skeletal, smooth, and cardiac. Later, these skeletal muscles lose their segmental pattern of development as they acquire insertion on skeletal elements. These muscle fibers become the striated voluntary muscles, which divide into groups that supply the dorsal and ventral parts of the limbs and provide both the deep and superficial muscle fibers (see Fig. 1-20, B). These muscles are called striated because they have lines across them that are the contraction sites that cause the muscles to function.
Muscle cells also migrate to the gastrointestinal tract and support the trachea, bronchi, urogenital tract, and larger blood vessels. These muscle cells develop and become oriented in the direction in which their contractility will be exerted. They are termed smooth muscle cells and are under the control of the autonomic nervous system, not under conscious control as are skeletal muscles. The blood vessels that develop in the head region, limbs, and body wall gain their muscular coat from local mesenchyme.
Cardiovascular System
The cardiovascular system originates from cells termed angioblasts, which arise from angiogenic clusters from the visceral mesoderm located in the walls of the yolk sac during the third week of prenatal life (Fig. 1-23). As these cells separate into clusters, the outer cells organize into a series of elongating tubes and the inner cells become blood cells (Fig. 1-24). For the first few weeks, nutrition moves from the yolk sac to the embryo through the developing vitelline vascular system (Fig. 1-25). The entire blood vascular system within the embryo is created in the same manner with longitudinal growth of vessels and the appearance of blood cells within them. As vessels begin to develop in the embryo, they, in turn, form a vascular network connected to the placenta. Because it traverses the umbilical cord, this network is termed the umbilical system (see Fig. 1-25). Through this umbilical system, nutrition and oxygen are conducted to the embryo, and carbon dioxide and wastes to the placenta. By the fourth week, the heart begins to beat. This vascular system takes over the functions as the vitelline system expires because the yolk sac has nothing more to contribute (see Fig. 1-25).
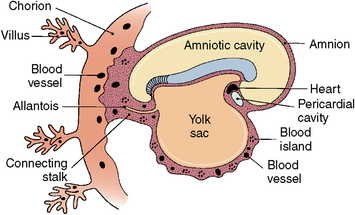
Fig. 1-23 Origin of blood cells and blood vessels in walls of yolk sac, placenta, and body stalk in a 2½-week-old embryo.
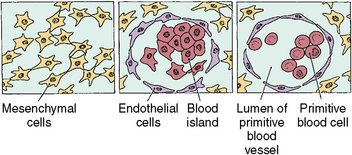
Fig. 1-24 Appearance of blood islands from mesenchymal cells in the location noted in Figure 1-22. The more peripheral cells form capillary walls, and the inner cells form red blood cells. The tubes or capillaries then lengthen.
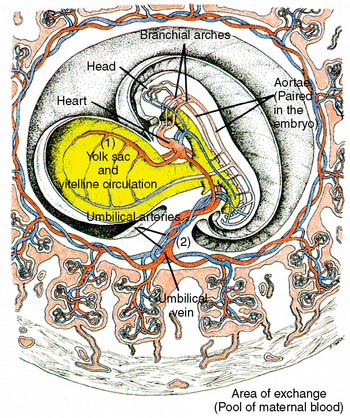
Fig. 1-25 Development of blood vascular system in an embryo. 1, In the yolk sac, vitelline circulation develops, persisting for only a few weeks until this nutritional source is exhausted. 2, The umbilical system develops in the umbilical cord, supplying the embryo and fetus with oxygen and nutrients until birth.
Other mesenchyme cells migrate into the pericardial area to function in the development of heart tubes, and these cells later differentiate into cardiac muscle. Two angiogenic cell clusters initially form the straight bilateral endocardial heart tubes, which fuse during the third week. They then enlarge and bend back on themselves (Fig. 1-26). As the great vessels that bring blood to the heart enlarge and become more extensive, the heart grows and internal partitioning begins. An opening persists between the right and left atria (foramen ovale) until birth. As the heart tube enlarges and twists during development, strands of muscle take on the arrangement of parallel fibers. Like striated muscle, cardiac muscle fibers are also striated and have an array of specialized junctional complexes between adjacent cells called intercalated disks. The myofibrils on either side of these disks exert contraction through the interaction of these many cells. Cardiac muscle is thus not under conscious control and begins to beat during the fourth week. Umbilical circulation then becomes active in transporting oxygen and nutrition from the placenta.
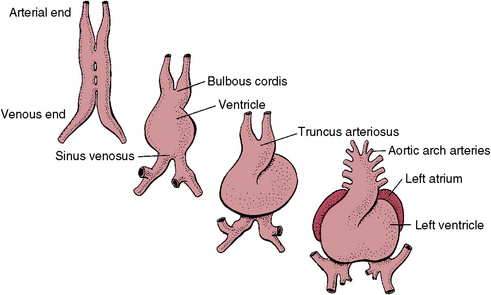
Fig. 1-26 Development of the four-chamber heart from fusion of two bilateral endocardiac heart tubes. Tubes fold laterally into a single tube, which is next divided by internal septa into a four-chamber heart.
The human placenta is often considered in terms of its function in exchanging fetal oxygen and carbon dioxide. It also exchanges nutrients and electrolytes, such as proteins and carbohydrates. The placenta produces hormones, such as progesterone and estrogen, which can help maintain pregnancy. It also produces a lactogenic hormone that gives the fetus first priority on circulating maternal blood glucose.
Developmental Abnormalities
Developmental defects may be environmental or hereditary. Most developmental defects are usually an interaction between environmental and hereditary factors. Not much can be done to reduce hereditary factors in humans. However, we have learned a great deal about dietary and stress factors and when they may affect development. For example, we know the developing human is least susceptible to teratogens during the proliferative period, which is the first 2 to 3 weeks after conception. Because of multiple cell mitosis, compensation may occur. However, the third through the eighth weeks are the most critical time in development because this is the period of differentiation. During this time, the embryo tissues and organs are developing into specific structures. Serious malformations may arise during this period. The fetal period from 8 weeks until birth is a declining period of susceptibility. Only minor defects may occur during this period.
Hereditary causes of abnormalities may result from either genetic or chromosomal abnormalities. Many chromosomal abnormalities may result from an increase or a decrease in number from the normal number of chromosomes (46) in humans.
Genetic abnormalities can perpetuate from one generation to the next. Abnormal development may be caused by expression of defective genes, which may be dominant or recessive. A dominant gene expresses itself whether it is on one member of the pair of homologous chromosomes or both pairs. A recessive gene expresses itself only when it is present on both members of the homozygous chromosomes. An example of a dominant genetic abnormality is dentinogenesis imperfecta, which results in defective dentin formation. Some examples of autosomal recessive genetic disorders include sickle cell disease and cystic fibrosis. There are also sex (X-linked) recessive defects including hemophilia and Duchenne’s muscular dystrophy.
SELF-EVALUATION QUESTIONS
1. What is the smallest unit of structure, and what are its eight functions in the body?
2. Name the structures found in cell cytoplasm, and describe their functions.
3. Name the cells that do not undergo division.
4. Describe changes in the embryonic disk during the third and fourth prenatal weeks.
5. Define the cell cycle and describe the activities that occur in the G1 and G2 phases.
6. What is the significance of the angiogenic clusters found in the vitelline and umbilical vascular systems?
7. What develops from the gastrointestinal tract?
8. Describe the characteristics of the three prenatal periods.
9. Name three types of cartilage, and describe where they are in the human body.
Discussion: Two diagnostic tests are available. Amniocentesis is the withdrawal of a small amount of amniotic fluid; it reveals genetic disorders and age of the fetus. Fetal ultrasound reflects body tissues to the video monitor; it reveals abnormal or normal development, vitality, sex, and fetal age. Neither test causes tissue damage. Ultrasound would be the choice in this case.