Physiologic Anatomy of the Male Sexual Organs and Spermatogenesis
 Describe the physiologic anatomy of the male sexual organs.
Describe the physiologic anatomy of the male sexual organs.
Describe the steps of spermatogenesis.
List the hormonal factors that stimulate spermatogenesis.
Describe the structure of the mature sperm.
Describe the role of the prostate gland and seminal vesicle.
Describe the composition of semen.
Define the following terms: cryptorchidism, oligospermia, and azoospermia.
The proliferation and differentiation of spermatogonia through definite stages of development to form sperm.
Immature germ cells in the testes.
It is the process by which spermatids mature into spermatozoa.
It is failure of testis to descend from the abdomen into the scrotum at or near the time of birth.
Low sperm count in the ejaculate.
Absence of sperm in the ejaculate.
Figure 94-1A shows the various portions of the male reproductive system and Figure 94-1B gives a more detailed structure of the testis and epididymis. The testis is composed of up to 900 coiled seminiferous tubules, each averaging more than one-half meter long, in which the sperms are formed. The sperms then empty into the epididymis, another coiled tube about 6 meters long. The epididymis leads into the vas deferens, which enlarges into the ampulla of the vas deferens immediately before the vas enters the body of the prostate gland.
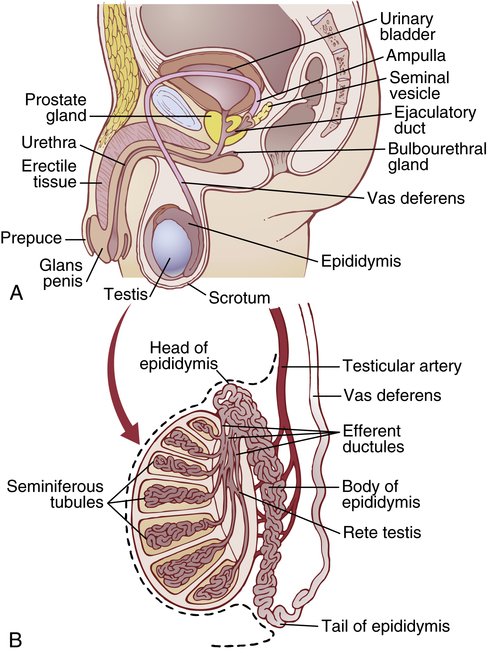
Two seminal vesicles, one located on each side of the prostate, empty into the prostatic end of the ampulla and the contents from both the ampulla and the seminal vesicles pass into an ejaculatory duct leading through the body of the prostate gland and then emptying into the internal urethra. Prostatic ducts also empty from the prostate gland into the ejaculatory duct and from there into the prostatic urethra.
Finally, the urethra is the last connecting link from the testis to the exterior. The urethra is supplied with mucus derived from a large number of minute urethral glands located along its entire extent and even more so from bilateral bulbourethral glands (Cowper glands) located near the origin of the urethra.
Spermatogenesis
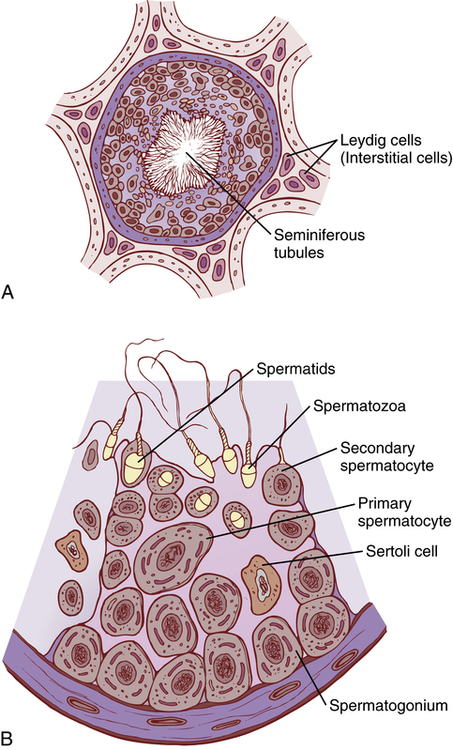
During formation of the embryo, the primordial germ cells migrate into the testes and become immature germ cells called spermatogonia, which lie in two or three layers of the inner surfaces of the seminiferous tubules (a cross section of one is shown in Figure 94-2A). The spermatogonia begin to undergo mitotic division, beginning at puberty, and continually proliferate and differentiate through definite stages of development to form sperms, as shown in Figure 94-2B.
Steps of Spermatogenesis
Spermatogenesis occurs in the seminiferous tubules during active sexual life as the result of stimulation by anterior pituitary gonadotropic hormones, beginning at an average age of 13 years and continuing throughout most of the remainder of life but decreasing markedly in old age.
In the first stage of spermatogenesis, the spermatogonia migrate among Sertoli cells toward the central lumen of the seminiferous tubule. The Sertoli cells are large with overflowing cytoplasmic envelopes that surround the developing spermatogonia all the way to the central lumen of the tubule.
Meiosis
Spermatogonia that cross the barrier into the Sertoli cell layer become progressively modified and enlarged to form large primary spermatocytes (Figure 94-3). Each of these, in turn, undergoes meiotic division to form two secondary spermatocytes. After another few days, these two divide to form spermatids that are eventually modified to become spermatozoa (sperms).
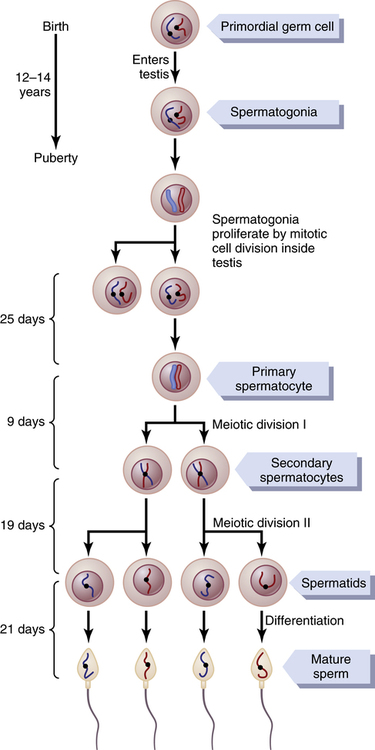
During the change from the spermatocyte stage to the spermatid stage, the 46 chromosomes (23 pairs of chromosomes) of the spermatocyte are divided, so 23 chromosomes go to one spermatid and the other 23 to the second spermatid. This also divides the chromosomal genes so that only one-half of the genetic characteristics of the eventual fetus are provided by the father, whereas the other half are derived from the oocyte provided by the mother.
The entire period of spermatogenesis, from spermatogonia to spermatozoa, takes about 74 days.
Sex Chromosomes
In each spermatogonium, one of the 23 pairs of chromosomes carries the genetic information that determines the sex of each eventual offspring. This pair is composed of one X chromosome, which is called the female chromosome, and one Y chromosome, the male chromosome. During meiotic division, the male Y chromosome goes to one spermatid that then becomes a male sperm, and the female X chromosome goes to another spermatid that becomes a female sperm. The sex of the eventual offspring is determined by which of these two types of sperm fertilizes the ovum. Fertilization is discussed further in Chapter 98.
Formation of Sperm
When the spermatids are first formed, they still have the usual characteristics of epithelioid cells, but soon they begin to differentiate and elongate into spermatozoa (spermiogenesis). As shown in Figure 94-4, each spermatozoon is composed of a head and a tail. The head comprises the condensed nucleus of the cell with only a thin cytoplasmic and cell membrane layer around its surface. On the outside of the anterior two-thirds of the head is a thick cap called the acrosome that is formed mainly from the Golgi apparatus. This contains a number of enzymes similar to those found in lysosomes of the typical cell, including hyaluronidase (which can digest proteoglycan filaments of tissues) and powerful proteolytic enzymes (which can digest proteins). These enzymes play important roles in allowing the sperm to enter the ovum and fertilize it.

The tail of the sperm, called the flagellum, has three major components: (1) a central skeleton constructed of 11 microtubules, collectively called the axoneme—the structure of this is similar to that of cilia found on the surfaces of other types of cells; (2) a thin cell membrane covering the axoneme; and (3) a collection of mitochondria surrounding the axoneme in the proximal portion of the tail (called the body of the tail).
Back-and-forth movement of the tail (flagellar movement) provides motility for the sperm. This movement results from a rhythmical longitudinal sliding motion between the anterior and posterior tubules that make up the axoneme. The energy for this process is supplied in the form of adenosine triphosphate, which is synthesized by the mitochondria in the body of the tail.
Normal sperm move in a fluid medium at a velocity of 1 to 4 mm/min. This allows them to move through the female genital tract in quest of the ovum.
Hormonal Factors That Stimulate Spermatogenesis
The role of hormones in reproduction is discussed later, but at this point, let us note that several hormones play essential roles in spermatogenesis. Some of these are as follows:
1. Testosterone, secreted by the Leydig cells located in the interstitium of the testis (see Figure 94-2), is essential for growth and division of the testicular germinal cells, which is the first stage in forming sperm.
2. Luteinizing hormone, secreted by the anterior pituitary gland, stimulates the Leydig cells to secrete testosterone.
3. Follicle-stimulating hormone, also secreted by the anterior pituitary gland, stimulates the Sertoli cells; without this stimulation, the conversion of the spermatids to sperm (the process of spermiogenesis) will not occur.
4. Estrogens, formed from testosterone by the Sertoli cells when they are stimulated by follicle-stimulating hormone, are probably also essential for spermiogenesis.
5. Growth hormone (as well as most of the other body hormones) is necessary for controlling background metabolic functions of the testes. Growth hormone specifically promotes early division of the spermatogonia themselves; in its absence, as in pituitary dwarfs, spermatogenesis is severely deficient or absent, thus causing infertility.
Maturation of Sperm in the Epididymis
After formation in the seminiferous tubules, the sperm require several days to pass through the 6-meter-long tubule of the epididymis. Sperms removed from the seminiferous tubules and from the early portions of the epididymis are nonmotile, and they cannot fertilize an ovum. However, after the sperm have been in the epididymis for 18 to 24 hours, they develop the capability of motility, even though several inhibitory proteins in the epididymal fluid still prevent final motility until after ejaculation.
Storage of Sperms in the Testes
The two testes of the human adult form up to 120 million sperms each day. A small quantity of these can be stored in the epididymis, but most are stored in the vas deferens. They can remain stored, maintaining their fertility, for at least a month. During this time, they are kept in a deeply suppressed, inactive state by multiple inhibitory substances in the secretions of the ducts. Conversely, with a high level of sexual activity and ejaculations, storage may be no longer than a few days.
After ejaculation, the sperms become motile and they also become capable of fertilizing the ovum, a process called maturation. The Sertoli cells and the epithelium of the epididymis secrete a special nutrient fluid that is ejaculated along with the sperm. This fluid contains hormones (including both testosterone and estrogens), enzymes, and special nutrients that are essential for sperm maturation.
Physiology of the Mature Sperm
The normal motile, fertile sperms are capable of flagellated movement through the fluid medium at velocities of 1 to 4 mm/min. The activity of sperms is greatly enhanced in a neutral and slightly alkaline medium, as exists in the ejaculated semen, but it is greatly depressed in a mildly acidic medium. A strong acidic medium can cause rapid death of sperm.
The activity of sperms increases markedly with increasing temperature, but so does the rate of metabolism, causing the life of the sperms to be considerably shortened. Although sperms can live for many weeks in the suppressed state in the genital ducts of the testes, life expectancy of ejaculated sperm in the female genital tract is only 1 to 2 days.
Function of the Seminal Vesicles
Each seminal vesicle is a tortuous, loculated tube lined with a secretory epithelium that secretes a mucoid material containing an abundance of fructose, citric acid, and other nutrient substances, as well as large quantities of prostaglandins and fibrinogen. During the process of emission and ejaculation, each seminal vesicle empties its contents into the ejaculatory duct shortly after the vas deferens empties the sperms. This adds greatly to the bulk of the ejaculated semen, and the fructose and other substances in the seminal fluid are of considerable nutrient value for the ejaculated sperms until one of the sperm fertilizes the ovum.
Prostaglandins are believed to aid fertilization in two ways: (1) by reacting with the female cervical mucus to make it more receptive to sperm movement and (2) by possibly causing backward, reverse peristaltic contractions in the uterus and fallopian tubes to move the ejaculated sperms toward the ovaries (a few sperms reach the upper ends of the fallopian tubes within 5 minutes).
Function of the Prostate Gland
The prostate gland secretes a thin, milky fluid that contains calcium, citrate ion, phosphate ion, a clotting enzyme, and a profibrinolysin. During emission, the capsule of the prostate gland contracts simultaneously with the contractions of the vas deferens so that the thin, milky fluid of the prostate gland adds further to the bulk of the semen. A slightly alkaline characteristic of the prostatic fluid may be quite important for successful fertilization of the ovum because the fluid of the vas deferens is relatively acidic owing to the presence of citric acid and metabolic end products of the sperm and, consequently, helps to inhibit sperm fertility. Also, the vaginal secretions of the female are acidic (pH of 3.5 to 4.0). Sperms do not become optimally motile until the pH of the surrounding fluids rises to about 6.0 to 6.5. Consequently, it is probable that the slightly alkaline prostatic fluid helps to neutralize the acidity of the other seminal fluids during ejaculation and thus enhances the motility and fertility of the sperms.
Semen
Semen, which is ejaculated during the male sexual act, is composed of the fluid and sperm from the vas deferens (about 10% of the total), fluid from the seminal vesicles (almost 60%), fluid from the prostate gland (about 30%), and small amounts from the mucous glands, especially the bulbourethral glands. Thus, the bulk of the semen is seminal vesicle fluid, which is the last to be ejaculated and serves to wash the sperm through the ejaculatory duct and urethra.
The average pH of the combined semen is about 7.5, the alkaline prostatic fluid having more than neutralized the mild acidity of the other portions of the semen. The prostatic fluid gives the semen a milky appearance, and fluid from the seminal vesicles and mucous glands gives the semen a mucoid consistency. Also, a clotting enzyme from the prostatic fluid causes the fibrinogen of the seminal vesicle fluid to form a weak fibrin coagulum that holds the semen in the deeper regions of the vagina where the uterine cervix lies. The coagulum then dissolves during the next 15 to 30 minutes because of lysis by fibrinolysin formed from the prostatic profibrinolysin. In the early minutes after ejaculation, the sperms remain relatively immobile, possibly because of the viscosity of the coagulum. As the coagulum dissolves, the sperms simultaneously become highly motile.
Abnormal Spermatogenesis and Male Fertility
The seminiferous tubular epithelium can be destroyed by a number of diseases. For instance, bilateral orchitis (inflammation) of the testes resulting from mumps causes sterility in some affected males.
Effect of Temperature on Spermatogenesis
Increasing the temperature of the testes can prevent spermatogenesis by causing degeneration of most cells of the seminiferous tubules besides the spermatogonia. It has often been stated that the reason the testes are located in the dangling scrotum is to maintain the temperature of these glands below the internal temperature of the body, although usually only about 2°C below the internal temperature. On cold days, scrotal reflexes cause the musculature of the scrotum to contract, pulling the testes close to the body to maintain this 2° differential. Thus, the scrotum acts as a cooling mechanism for the testes (but a controlled cooling), without which spermatogenesis might be deficient during hot weather.
Cryptorchidism
Cryptorchidism means failure of a testis to descend from the abdomen into the scrotum at or near the time of birth of a fetus. During development of the male fetus, the testes are derived from the genital ridges in the abdomen. However, at about 3 weeks to 1 month before birth of the baby, the testes normally descend through the inguinal canals into the scrotum. Occasionally, this descent does not occur or occurs incompletely, so one or both testes remain in the abdomen, in the inguinal canal, or elsewhere along the route of descent.
A testis that remains throughout life in the abdominal cavity is incapable of forming sperms. The tubular epithelium becomes degenerate, leaving only the interstitial structures of the testis. It has been claimed that even the few degrees’ higher temperature in the abdomen than in the scrotum is sufficient to cause this degeneration of the tubular epithelium and, consequently, to cause sterility, although this is not certain. Nevertheless, for this reason, operations to relocate the cryptorchid testes from the abdominal cavity into the scrotum before the beginning of adult sexual life can be performed on boys who have undescended testes.
Testosterone secretion by the fetal testes is the normal stimulus that causes the testes to descend into the scrotum from the abdomen. Therefore, many, if not most, instances of cryptorchidism are caused by abnormally formed testes that are unable to secrete enough testosterone. The surgical operation for cryptorchidism in these patients is unlikely to be successful.
Effect of Sperm Count on Fertility
The usual quantity of semen ejaculated during each coitus averages about 3.5 milliliters, and in each milliliter of semen is an average of about 120 million sperms, although even in “normal” males this can vary from 35 to 200 million. This means an average total of 400 million sperms are usually present in the several milliliters of each ejaculate. When the number of sperms in each milliliter falls below about 20 million, the person is likely to be infertile. A reduction in the sperm count below 20 million is referred to as oligospermia while the absence of sperm in the ejaculate is called azoospermia. Thus, even though only a single sperm is necessary to fertilize the ovum, for reasons not understood, the ejaculate usually must contain a tremendous number of sperm for only one sperm to fertilize the ovum. There are many causes of low counts and these include hormonal imbalances, infections or tumors involving the testes, exposure to environmental toxins, and lifestyle issues (chronic alcoholism, smoking, and substance abuse) among others.
Effect of Sperm Morphology and Motility on Fertility
Occasionally, a man has a normal number of sperms but is still infertile. When this occurs, sometimes as many as one-half the sperms are found to be abnormal physically, having two heads, abnormally shaped heads, or abnormal tails, as shown in Figure 94-5. At other times, the sperm appear to be structurally normal, but for reasons not understood, they are either entirely nonmotile or relatively nonmotile. Whenever the majority of the sperms are morphologically abnormal or are nonmotile, the person is likely to be infertile, even though the remainder of the sperms appear to be normal.

Table 94-1 summarizes the composition of the semen that has been discussed in the sections above.
Table 94-1
| Appearance | Translucent, thick, with a whitish tint |
| Volume | ~ 3.5 ml (1–6.5 milliliters) |
| pH | ~ 7.5 |
| Number of sperms | ~ 35–200 million/milliliters (> 15 million is normal) |
| Percent of motile sperms | > 50%, Measured within 60 minutes of collection of the sample |
| Percent contribution by: Vas deferens Seminal vesicle Prostate Bulbourethral glands |
~ 10% ~ 60% ~ 30% Minimal ~ 1% |