The Cell Physiology
 Cell membrane
Cell membraneCell structure
The cell is the smallest structural and functional unit of the body. The human body contains about 100 trillion cells. Different types of cells of the body possess features which distinguish one type from the other, and are specially adapted to perform particular functions, e.g. the red blood cells transport oxygen from lungs to the tissues, muscle cell is specialized for the function of contraction.
A typical cell, as seen by the light microscope, consists of three basic components:
Cell membrane
Cell membrane or the plasma membrane is the protective sheath enveloping the cell body. It separates the contents of cell from the external environment and controls exchange of materials between the fluid outside the cell (extracellular fluid) and the fluid inside the cell (intracellular fluid). A detailed knowledge of its structure (Fig. 1.2-1) is essential for the understanding of cell functions. Therefore, it will be discussed separately.
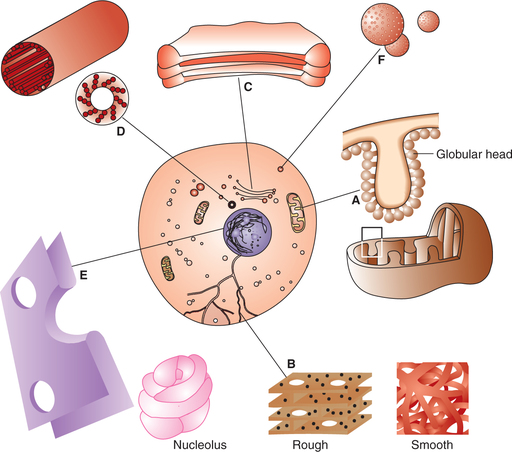
Cytoplasm
Cytoplasm is an aqueous substance (cytosol) containing a variety of cell organelles and other structures. The structures dispersed in the cytoplasm can be broadly divided into three groups: organelles, inclusion bodies and cytoskeleton.
A. Organelles
The organelles are the permanent components of the cells which are bounded by limiting membrane and contain enzymes; hence participate in the cellular metabolic activity. These include the following.
1. Mitochondria
Mitochondria are the major sites for aerobic respiration. These are oval structures and more numerous in metabolically active cells.
Structure. The mitochondria consist of:
• Membrane. There are two layers of the membrane. The outer smooth and inner folded into incomplete septa called cristae (Fig. 1.2-1A).
• Matrix of the mitochondria contains enzymes required in Krebs’ cycle by which products of carbohydrate, fat and protein metabolism are oxidised to produce energy that is stored in the form of ATP in the lollipop-like globular structures.
Functions. In addition to their role as power generating units, the mitochondria may have a role in synthesizing membrane-bound proteins since they also possess deoxyribonucleic acid (DNA) and ribosomes.
2. Endoplasmic reticulum
Endoplasmic reticulum (ER) is a system of flattened membrane-bound vesicles and tubules called cisternae (Fig. 1.2-1B). It is continuous with the outer membrane of the nuclear envelop, Golgi apparatus and possibly with the cell membrane. Morphologically, two types of ER can be identified: rough or granular and smooth or agranular.
(i) Rough endoplasmic reticulum. The rough ER is characterized by the presence of a number of ribosomes on its surface and transports proteins made by the ribosomes through the cisternae. Thus, the rough ER is especially well developed in cells active in protein synthesis, e.g. Russell's bodies of plasma cells, Nissl granules of nerve cells and acinar cells of pancreas.
(ii) Smooth endoplasmic reticulum. Smooth ER is devoid of ribosomes on its surface. It is a site of lipid and steroid synthesis. Therefore, it is found in abundance with the Leydig cells and cells of the adrenal cortex. In the skeletal and cardiac muscles, smooth ER is modified to form sarcoplasmic reticulum which is involved in the release and sequestration of calcium ions during muscular contraction.
3. Golgi apparatus
The Golgi apparatus or complex is a collection of membranous vesicles, sacs or tubules which is generally located close to the nucleus. It is continuous with the ER. Golgi apparatus is particularly well developed in exocrine glandular cells (Fig. 1.2-1C).
4. Ribosomes
Ribosomes are spherical particles which contain 80%-85% of the cell's ribonucleic acid (RNA). They may be present in the cytosol as free (unattached) or in bound form (attached to the membrane of ER). Slightly smaller form of ribosomes is also found in mitochondria.
Functions. They are the site of protein synthesis. They synthesize all transmembrane proteins, secreted proteins and most proteins that are stored in the Golgi apparatus, lysosomes and endosomes.
5. Lysosomes
Lysosomes are rounded to oval membrane-bound organelles containing powerful lysosomal digestive (hydrolytic) enzymes. They are formed by the Golgi apparatus. As many as 40 different lysosomal enzymes have been synthesized. Lysosomes are particularly abundant in cells involved in phagocytic activity, e.g. neutrophils and macrophages. There are three forms of lysosomes:
• Primary lysosomes or storage vacuoles are formed from the various hydrolytic enzymes synthesized by rough ER and packaged in the Golgi apparatus.
• Secondary lysosomes or autophagic vacuoles are formed by fusion of primary lysosomes with parts of damaged or worn-out cell components.
• Residual bodies are undigestible materials in the lysosomes.
6. Peroxisomes
Peroxisomes, also known as microbodies, are spherical structures enclosed by a single layer of unit membrane. These are predominantly present in hepatocytes and tubular epithelial cells.
7. Centrosome
The centrosome consists of two short cylindrical structures called centrioles (Fig. 1.2-1D). It is situated near the centre of the cell close to the nucleus. The centrioles are responsible for movement of chromosomes during cell division.
B. Cytoplasmic inclusions
The cytoplasmic inclusions are the temporary components of certain cells. These may or may not be enclosed in the membrane. A few examples of cytoplasmic inclusions are given below.
• Lipid droplets. These are seen in the cells of adipose tissue, liver and adrenal cortex.
• Glycogen. It is seen in the cells of liver and skeletal muscles.
• Proteins as secretory granules are seen in the secretory glandular cells (Fig. 1.2-1F).
• Melanin pigment is seen in the cells of epidermis, retina and basal ganglia.
• Lipofuscin. It is a yellow-brown pigment believed to be derived from secondary lysosomes and is seen in the cardiac muscle and brain cells of elderly people.
C. Cytoskeleton
The cytoskeleton is a complex network of fibres that maintains the structure of the cell and allows it to change shape and move. It primarily consists of the following (Fig. 1.2-2).
1. Microtubules
Microtubules are long, hollow, tubular structures without limiting membrane about 25 nm in diameter. These are made up of two globular protein subunits α- and β-tubulin. The bundles of tubulin give structural strength to the cells. Microtubules form the transport system of the cells. Some of the other organelles and protein molecules move to a different part of the cell through the microtubules. Kinesin and dynein known as molecular motors help in the movement of molecules through the microtubules.
The cilia and flagella which project from surface of certain cells (spermatozoa, respiratory mucosa and fallopian tubes) are also composed of microtubules enclosed in the plasma membrane and are active in the locomotion of the cells.
2. Intermediate filaments
Intermediate filaments are filamentous structures about 10 nm in diameter. Some of these filaments connect the nuclear membrane to the cell membrane. Their main function is to mechanically integrate the cell organelles within the cytoplasm. In their absence, cells rupture more easily; and when they are abnormal in human, blistering of the skin is common.
3. Microfilaments
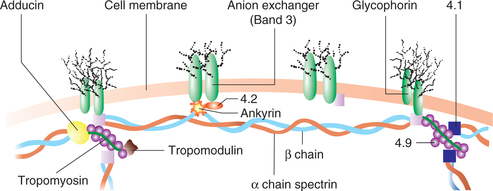
Microfilaments are long, solid, filamentous structures having a diameter of 6-8 nm. These are made up of contractile proteins, actin and myosin. Actin is the most abundant protein in the mammalian cell. It attaches to various parts of cytoskeleton by other proteins (anchor proteins). These are identified by numbers as 4.1, 4.2 and 4.9. Extension of microfilaments along with the plasma membrane on the surface of the cells form microvilli, which increase the absorptive surface of the cells (e.g. intestinal epithelium). In the skeletal muscle, presence of actin and myosin filaments is responsible for their contractile property.
Molecular motors
Molecular motors help in the movement of different proteins, organelles and other cell parts (their cargo) to all parts of the cell. These can be divided into two types:
1. Microtubule-based molecular motors. This is a super-family of molecular motors that produce motion along microtubules. Two important molecular motors are:
2. Actin-based molecular motors. This is a superfamily of molecular motors that produce motion along the actin. The important example of this group is myosin.
Nucleus
Nucleus is present in all the eukaryotic cells. It controls all the cellular activities including reproduction of the cell. Most of the cells are uninucleated except few types of cells like skeletal muscle cells which are multinucleated. The nucleus consists of the following (Fig. 1.2-1E).
1. Nuclear membrane
The nuclear membrane is double-layered porous structure having a 40–70 nm wide space called perinuclear cistern, which is continuous with the lumen of ER. The outer layer of the nuclear membrane is continuous with ER. The exchange of materials between the nucleoplasm and cytoplasm occurs through the nuclear membrane.
2. Nucleoplasm
The nucleoplasm or the nuclear matrix is a gel-like ground substance containing a large quantity of genetic material in the form of DNA. When a cell is not dividing the nucleoplasm appears as dark staining thread-like material called nuclear chromatin. During cell division the chromatin material is converted into rod-shaped structures, the chromosomes. There are 46 chromosomes (23 pairs) in all the dividing cells of the body except the gamete (sex cells) which contains only 23 chromosomes (haploid number). Each chromosome is composed of two chromatids connected at the centromere to form ’X’ configuration having variation of the location of centromere. The chromosomes are composed of three components: DNA, RNA and other nuclear proteins. The nuclear DNA carries the genetic information which is passed via RNA into the cytoplasm for synthesis of proteins of similar composition.
The cell membrane
An understanding of the structure and properties of the cell membrane is most essential to understand the various physiological activities of the cell. Electron microscopy has shown that cell membrane/plasma membrane has a trilayer structure having a total thickness of 7–10 nm (70–100 Å) and is known as unit membrane. The three layers consist of two electron-dense layers separated by an electron-lucent layer (clear zone). Biochemically the cell membrane is composed of a complex mixture of lipids (40%), proteins (55%) and carbohydrates (5%).
Hypothesis for cell membrane structure
A few hypotheses have been proposed to explain the distribution of various biochemical components in the cell membrane. The most important hypothesis is the fluid mosaic model of Singer and Nicholson.
Fluid mosaic model of membrane structure
In 1972, Singer and Nicholson put forward the fluid mosaic model of membrane structure (Fig. 1.2-3), which is presently most accepted. According to this model:
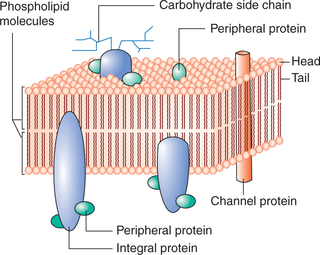
• Phospholipid bilayer is the basic continuous structure forming the cell membrane. The phospholipids are present in fluid form. This fluidity makes the membrane quite flexible and thus allows the cells to undergo considerable changes in the shape without disruption of structural integrity.
• The protein molecules are present as a discontinuous mosaic of globular proteins which float about in the fluid phospholipid bilayer forming a fluid mosaic pattern.
Arrangement of different molecules in cell membrane
Arrangement of lipid bilayer of the cell membrane
Each lipid molecule in the lipid bilayer of the cell membrane primarily consists of phospholipid, cholesterol and glycolipids. The lipid molecule is clothespin shape and consists of a head end and a tail end.
The head end or the globular end of the molecule contains phosphate moiety of phospholipid or hydroxyl radical of cholesterol. It is positively charged and quite soluble in water (i.e. polar or hydrophilic). The tail end consists of two chains of fatty acids or steroid radicle of cholesterol. It is quite insoluble in water (non-polar or hydrophobic). These lipid molecules are arranged as bilayer in such a way that their non-polar hydrophobic tail ends are directed towards the centre of the membrane whereas their polar hydrophilic head ends are directed outwards on either side of the membrane (Fig. 1.2-3). In this way head ends of molecules face the aqueous phase, i.e. extracellular fluid on outside and the intracellular fluid (cytoplasm) on inner side. The major types of phospholipids are phosphatidyl choline, phospha-tidyl ethanolamine, glycosphingolipids, sphingomyelin and cholesterol.
Functional significance of the lipid bilayer. The lipid bilayer of the cell membrane makes it a semipermeable membrane which constitutes the major barrier for the water-soluble molecules like electrolytes, urea and glucose. On the other hand, fat-soluble substances like oxygen, fatty acids and alcohol can pass through the membrane with ease.
Arrangement of proteins in the cell membrane
Most protein molecules float about in the phospholipid bilayer forming a fluid mosaic pattern. The two types of proteins recognized in the cell membrane are:
• Lipoproteins, i.e. the proteins containing lipids which function as enzymes and ion channels, and
• Glycoproteins, i.e. the proteins containing carbohydrates which function as receptors for hormones and neurotransmitters.
The proteins in the cell membrane are described below.
1. Peripheral proteins. These are present peripheral to the lipid bilayer, both inside and outside to it.
(i) Intrinsic proteins. These are located on the inner surface of the lipid bilayer and serve mainly as enzymes. Some of these are anchored to the cytoskeleton of the cell (Fig. 1.2-2).
(ii) Extrinsic or surface proteins. These are the proteins located on the outer surface of the lipid bilayer. These protein molecules are not associated tightly with the cell membrane and thus can dissociate readily from the cell membrane. Some of these proteins serve as cell adhesion molecules (CAMs) that anchor cells to the neighbouring cells and basal lamina.
2. Integral proteins or transmembrane proteins. These are the proteins which extend into the lipid bilayer (Fig. 1.2-3). Some roteins penetrate only part of the way into the membrane while others penetrate all the way through. The integral proteins on the basis of functions they serve are discussed below.
• Channel proteins serve as channels for water-soluble substances like glucose and electrolytes.
• Carrier proteins help in transport of substances across the cell membrane by means of active and passive (facilitated diffusion) transport.
• Receptor proteins. Some of the proteins function as receptors that bind neurotransmitters and hormones, initiating physiologic changes inside the cell.
• Antigens. Some proteins in the cell membrane also act as antigens.
• Pumps. There are certain proteins in the cell membrane which act as pumps and form active transport system of the cell, e.g. Na+–K+ ATPase pump, K+–H+ ATPase pump and Ca2+ pump.
Intercellular junctions
The cell membranes of the neighbouring cells are connected with one another through the intercellular junctions or the junctional complexes, which are of three types.
Types of intercellular junctions
1. Tight junction. This is also called zona occludens or the occluding zone (Fig. 1.2-4A). In this type of intercellular junction, the outer layer of the cell membrane of the neighbouring cells fuse with each other, thus obliterating the space between the cells. Such junctions form a barrier to the movement of ions and other solutes from one cell to another. A few examples of the cells where tight junctions are present:
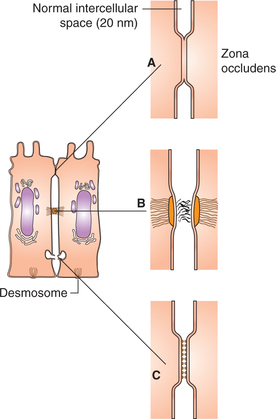
Fig. 1.2-4 Schematic diagram of a cell to show various intercellular junctions: A, tight junction; B, adherens junction and C, gap junction.
• Apical margins of epithelial cells, such as the intestinal mucosa.
• The renal tubular epithelial cells.
• Capillary endothelium in the brain forming blood-brain barrier.
2. Adherens junction. This is also called zonula adherens. In this type of junction, cell membranes of the adjacent cells are separated by a 15-20 nm wide space which is at focal places obliterated by the dense accumulation of the proteins at the cell surface. Bundles of intermediate filaments project from the intercellular junctional areas and radiate into the cytoplasm. This holds the adjacent cells at these focal places. These are of two types:
• Desmosomes are the adherens junctions where thickened focal areas are formed on both the apposing cell membranes (Fig. 1.2-4B).
• Hemidesmosomes are the adherens junctions where focal thickening is seen only on the membrane of one of the two adjacent cells. So, this is also known as half desmosome. Adherens junctions are seen in the cells of epidermis. Focal adhesions attach the cell to the basal lamina and also to actin filaments present inside the cell and thus help in cell movement.
3. Gap junction. Gap junctions or the nexus are the channels on the lateral surfaces of the two adjacent cells through which the molecules are exchanged between the cells (Fig. 1.2-4C). Each half of the channel is surrounded by six sub-units of proteins (the channel proteins). The intercellular space is reduced from the usual size of 15-20 nm to 2-3 nm at such junction. The gap junctions are seen in the heart and basal part of epithelial cells of intestinal mucous membrane. Gap junctions serve the following functions:
• These permit the intercellular passage of glucose, amino acids, ions and other substances which have a molecular weight of about 1000.
• These permit rapid propagation of electrical potential changes from one cell to another as seen in cardiac muscle and other smooth muscle cells.
• These help in the exchange of chemical messengers between the cells.