General Principles of Endocrinal System
Hormones: definition and classification
Definition
The word hormone is derived from the Greek word hormaein, which means to execute or to arouse. In the classic definition, hormones are secretory products of the ductless glands, which are released in catalytic amounts into the blood stream and transported to specific target cells (or organs), where they elicit physiologic, morphologic and biochemical responses.
Classification of hormones
A Depending upon the chemical nature
B Depending upon the mechanism of action
1. Group I hormones. These act by binding to intracellular receptor and mediate their actions via formation of a hormone–receptor complex. These include steroid, retinoid and thyroid hormones.
2. Group II hormones. These involve second messenger to mediate their effect. Depending upon the chemical nature of the second messengers, group II hormones are further divided into four subgroups: A, B, C and D (Table 8.1-1).
Table 8.1-1
Types of group II hormones based on the chemical nature of second messenger involved in their mechanism of action
| Group | Second messenger | Hormones |
| Group II-A | Cyclic AMP (cAMP) | Adrenocorticotropic hormone (ACTH) |
| Antidiuretic hormone (ADH) | ||
| Angiotensin II | ||
| Calcitonin | ||
| Corticotropic hormones (CRH) | ||
| Catecholamine (α2 adrenergic) | ||
| Follicular-stimulating hormone (FSH) | ||
| Glucagon | ||
| Luteinizing hormone (LH) | ||
| Parathormone (PTH) | ||
| Somatostatin | ||
| Thyroid-stimulating hormone (TSH) | ||
| Group II-B | Cyclic GMP (cGMP) | Atrial natriuretic factor (ANF) and nitric oxide |
| Group II-C | Calcium/or phosphatidyl inositol/ or both | Acetylcholine (ACh), catecholamines (α1 adrenergic), gastrin, oxytocin, thyrotropin-releasing hormone (TRH), gonadotropin-releasing hormone (GnRH), platelet-derived growth factor (PDGF) and substances. |
| Group II-D | Kinase or phosphatase cascade | Human chorionic somatotropin (HCS), erythropoietin, growth hormone, insulin and insulin-like growth factors (IGF-I and IGF-II), nerve growth factor (NGF), prolactin and other growth factors |
Hormones: general considerations
Hormone transport, plasma concentration and half-life
Hormone transport
After secretion into the blood stream, the hormones may circulate in two forms:
Unbound form. Some hormones circulate as free molecule, e.g. catecholamines and most peptide and protein hormones circulate unbound.
Bound form. Some hormones, such as steroids, thyroid hormones and vitamin D circulate bound to specific globulins that are synthesized in the liver. The binding of hormones to proteins is advantageous as it:
Plasma concentration
Hormones are usually secreted into the circulation in extremely low concentrations:
Half-life
Most hormones are metabolized rapidly after secretion. In general:
• Peptide hormones have short half-life.
• Steroids and thyroid hormones have significantly longer half-life because they are bound to the plasma proteins. Table 8.1-2 depicts half-life of some of the hormones.
Table 8.1-2
Half-life of some of the important hormones
| Class of hormone | Hormone | Half-life |
| Protein and peptide hormones | ADH | < 1 min |
| Oxytocin | < 1 min | |
| Insulin | 5 min | |
| Prolactin | 12 s | |
| Growth hormone | < 30 min | |
| ACTH | 15−25 s | |
| LH | 15−45 min | |
| FSH | 180 min | |
| Amines | Epinephrine | 10 s |
| Norepinephrine | 15 s | |
| Thyroxine (T4) | 5−7 days | |
| Triiodothyronine (T3) | 1−3 days | |
| Steroid hormones | Aldosterone | 30 min |
| Cortisol | 90−100 min | |
| 1,25-Dihydroxycholecalciferol | 15 h | |
| 25 Hydroxycholecalciferol | 15 days |
Functions of hormones
Hormones regulate existing fundamental processes but do not initiate reactions de novo.
1. Regulation of biochemical reactions. Hormones regulate the metabolic functions in a variety of ways:
• They stimulate or inhibit the rate and magnitude of biochemical reactions by controlling enzymes and thereby cause morphologic, biochemical and functional changes in target tissues.
• They modulate energy-producing processes and regulate the circulating levels of energy-yielding substances (e.g. glucose, fatty acids). However, they are not used as energy sources in biochemical reactions.
2. Regulation of bodily processes. Hormones regulate different bodily processes, such as growth, maturation, differentiation, regeneration, reproduction and behaviour. Thus, main function of the endocrine glands is to maintain homeostasis in an internal environment.
Regulation of hormone secretion
The quantity of hormones secreted is regulated in accordance with their requirement. General mechanisms that govern the secretion of hormone include:
Feedback control is of two types:
Negative feedback control. Generally, the influence of blood concentration of the hormone concerned or its effect is to inhibit further secretion of the hormone and is called negative feedback control (Fig. 8.1-1A).
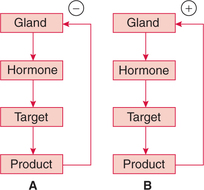
Fig. 8.1-1 Hormonal regulation by feedback control mechanism: A, negative feedback and B, positive feedback.
Positive feedback control. It is less common and acts to amplify the initial biological effects of the hormone (Fig. 8.1-1B).
Depending upon the product involved the feedback mechanism may be:
(i) Hormone–hormone feedback control
The best example of hormone–hormone negative feedback control is the regulation of hormone secretions by the hypothalamus and pituitary, which involves three loops (Fig. 8.1-2):
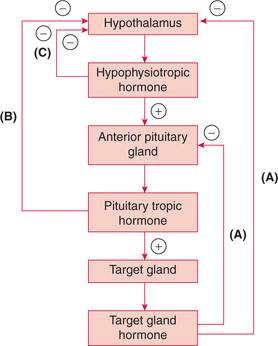
Fig. 8.1-2 Hormone–hormone negative feedback control by the hypothalamus and pituitary: A, long-loop feedback; B, short-loop feedback and C, ultra-short-loop feedback.
• Long-loop feedback (Fig. 8.1-2A). The peripheral gland hormone (e.g. thyroid, adrenocortical and gonads) can exert long-loop negative feedback control on both the hypothalamus and the anterior lobe of pituitary.
• Short-loop feedback (Fig. 8.1-2B). The pituitary tropic hormones decrease the secretion of hypophysiotropic hormone (e.g. GHRH, GHIH, TRH, GnRH, etc.) by shortloop feedback.
• Ultra-short-loop feedback (Fig. 8.1-2C). The hypophysiotropic hormones may inhibit their own synthesis and secretion via an ultra-short-loop feedback mechanism.
(ii) Substrate–hormone feedback control
The best example of substrate–hormone feedback control is regulation of insulin secretion from the pancreatic beta cells of the islets of Langerhans and glucagon secretion from the α cells by blood glucose levels. A rise in blood glucose level promotes the secretion of insulin, whereas a fall in blood glucose promotes secretion of glucagon. These responses keep the blood glucose level within narrow limits in spite of variation in carbohydrate intake in diet.
2 Neural control
Neural control acts to evoke or suppress hormone secretion in response to both external and internal stimuli.
External stimuli, which can modulate hormone release through neural mechanisms, may be visual, auditory, olfactory, gustatory and tactile.
Internal stimuli, which influence hormonal release through neural mechanism include pain, emotion, sexual excitement, fright, stress and changes in blood volume.
3 Chronotropic control
Chronotropic control of hormone secretion accounts for:
The source of regular oscillatory cycles is a pulse generator(s) located in the suprachiasmatic nucleus (SCN) of the hypothalamus (Fig. 8.1-3).
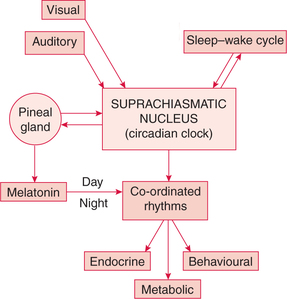
Fig. 8.1-3 The origin of circadian rhythms in endocrine gland secretion, metabolic process and behavioural activity.
The intrinsic circadian clock is also located in the SCN, which is responsible for endocrinal, metabolic and behavioural co-ordinated rhythms.
Hormone: receptors and mechanism of action
Hormone receptors
All hormones act through specific receptors. Almost all hormone receptors are large proteins present in hormonesensitive target cells.
Characteristics of hormone receptors
Receptor specificity. There are specific receptors for each hormone. This is the reason that all hormones circulate to all parts of the body, yet each hormone has a specific target tissue for its action (Fig. 8.1-4).
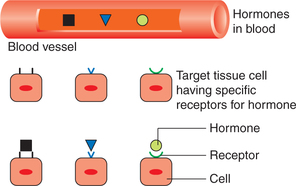
Change in receptor number. Number of receptors of a cell vary depending upon the situation. It is regulated by two mechanisms: down regulation and up regulation.
(i) Down regulation refers to a decrease in the number of active receptors. It occurs to regulate the hormone sensitivity when it is present in excess.
(ii) Up regulation refers to an increase in the number of active receptors on a cell. It occurs to regulate the hormone action when its concentration is less. This phenomenon tends to reduce the effect of hormone deficiency.
Mechanism of action of hormones
The main mechanisms of hormone actions are:
• Action through change in the membrane permeability,
• Action through effect on gene expression by binding of hormones with intracellular receptors,
• Action through secondary messengers which activate intracellular enzymes when hormones combine with membrane receptors and
Action through change in membrane permeability
Certain hormones bind with the receptors present in the cell membrane (external receptors) and cause conformational change in the protein of the receptors, this results into either opening or closing of the ions channels (such as Na+ channels, K+ channels and Ca2+ channels). The movement of ions through Ca2+ channels causes the subsequent effect, e.g. adrenaline, noradrenaline act by this mechanism.
Action through effect on gene expression by binding of hormones with intracellular receptors
Group I hormones act by their effect on the gene expression include steroid hormones, retinoids and thyroid hormones. These hormones are lipophilic in nature and can easily pass across the cell membrane. They act through intracellular receptors located either in the cytosol or in the nucleus. The sequence of events involved is (Fig. 8.1-5):
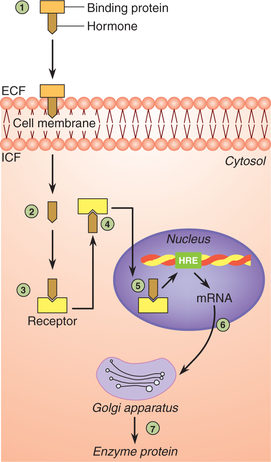
Fig. 8.1-5 Action of hormones through their effect of gene expression. Note 1–7 represent the steps involved in the process (for details see text).
1. Transport. After secretion, the hormone is carried to the target tissue on serum-binding protein.
2. Internalization. Being lipophilic, the hormone easily diffuses through the plasma membrane.
3. Receptor–hormone complex is formed by binding of hormone to the specific receptor inside the cell.
4. Conformational change occurs in the receptor proteins leading to activation of receptors.
5. The activated receptor–hormone complex then diffuses into the nucleus and binds on the specific region of the DNA known as hormone responsive element (HRE), which initiates gene transcription.
6. Binding of the receptor–hormone complex to DNA alters the rate of transcription of messenger RNA (mRNA).
7. The mRNA diffuses in the cytoplasm, where it promotes the translation process at the ribosomes. In this way new proteins are formed which result in specific responses. Some of the new proteins synthesized are enzymes.
Note. The hormonal action mediated through intracellular receptors is comparatively slower. Therefore glucocorticoids may take hours to few days to achieve the therapeutic effect.
Action through second messengers
Group II hormones (Table 8.1-1) release of second messenger is mediated by GTP binding proteins also called G proteins.
Coupling by G proteins
Events involved in coupling by G protein which lead onto changes in the cellular concentration of the second messengers are summarized (Fig. 8.1-6):
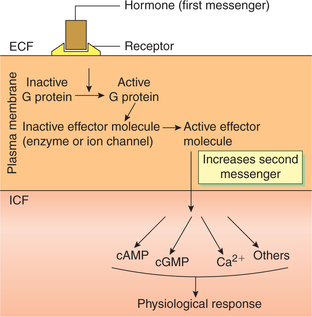
Fig. 8.1-6 Schematic mechanism of coupling by G protein leading to increase in second messenger, which mediates hormone's physiological response.
• Group II hormones are water soluble and bind to the plasma membrane of the target cell via cell surface receptors.
• The hormone-bearing receptor then interacts with a G protein and activates it by binding GTP. There are two classes of G proteins: stimulatory G protein (Gs) and inhibitory G protein (Gi).
• In its activated (“on”) state, the G protein interacts with one or more of the effector protein (most of which are enzymes or ion channels such as adenylyl cyclase; Ca2+ or K+ channels or phospholipase C, A2 or D) to activate or inhibit them.
• The changed effector molecules, in turn, generate second messenger that mediates the hormone's intracellular action.
Mechanism of action of hormone via tyrosine kinase activation
Group-II D hormones act by activating tyrosine kinase system (Table 8.1-1).
The activation of tyrosine kinase occurs by two mechanisms:
1. Hormone receptors possessing intrinsic tyrosine activity, e.g. those for insulin and epidermal growth factor involve following steps (Fig. 8.1-7A):
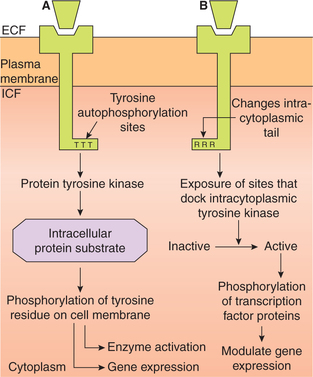
Fig. 8.1-7 Mechanism of action of hormone via tyrosin kinase activity: A, by receptors that possess intrinsic tyrosine activity and B, by receptors that do not possess intrinsic tyrosine activity.
• Binding of hormone to the receptor changes its conformation and exposes sites on its intracellular portion that are capable of receptor autophosphorylation at specific tyrosine sites.
• As a result, the receptor itself becomes a tyrosine kinase that phosphorylates tyrosine residue on the intracellular protein substrates.
• This latter activity sets into motion a cascade of events leading to enzyme activation and gene transcription.
2. Hormone receptors that do not possess intrinsic tyrosine activity, e.g. those for growth hormone, prolactin-releasing hormones, cytokines, etc. act as follows (Fig. 8.1.7B):
• Hormone binding to extracellular portion of the receptor changes its intracytoplasmic tail.
• The changes produced in the intracytoplasmic tail of receptor exposes sites which attract and dock the intracytoplasmic tyrosine kinases [such as janus tyrosine kinases (JAK) and signal transducer and activator of transcription (STAT) kinases] and then activates them.
• The activated intracytoplasmic tyrosine kinases phosphorylate cytoplasmic substrates, such as transcription factor proteins and ultimately modulate gene expression.
Measurement of hormones
Measurement of blood level of hormones is essential to confirm the endocrinal disorders associated with either deficiency or excess of a hormone. Since the hormones exist in the blood in very low concentration, the conventional methods of estimation such as colorimetry are not of much use. Therefore, they are measured by hormone assays and some special techniques which include:
Bioassay
In this method, hormone levels were assessed by injecting the unknown sample of plasma in the experimental animals and observing quantitatively the specific biological effect. The effect chosen was a characteristic action of the hormone for which a clear dose–response relationship existed.
Immunoassay
The immunoassay methods, frequently employed for estimation of hormone levels, include:
1 Radioimmunoassay
The radioimmunoassay is performed as:
• An unknown sample of plasma in which the concentration of a particular hormone (H) to be estimated is mixed with commercially available purified specific antibody (anti-H), and an appropriate amount of the purified hormone tagged with radioactive isotope (H+). The mixture is incubated in the cold.
• The antibodies have high affinity for the hormone. There occurs a competition between the free hormone (H) present in the unknown sample of plasma and the tagged hormone (H+) for binding to the specific antibody (anti-H).
2 Enzyme-linked immunosorbent assay method
Enzyme-linked immunosorbent assay (ELISA) method is principally similar to RIA, i.e. it is also based on the principle of antigen–antibody reaction. Any antigen that is protein can be measured by this technique. In this method, radioactivity is not measured, instead specific antibody– hormone (antigen) complex is stained with a suitable dye, and the intensity of colour is measured by the spectrophotometer. This technique is useful in estimating peptide and steroid hormones.
Cytochemicalassay
This test is much more sensitive than the immunoassay. In this technique, genesis of hormone can be detected in slices cut out of the endocrine gland by incubating them in a culture medium. This test is very useful in measuring the minute basal levels of hormone secretion.
Dynamic tests
Dynamic tests are needed in certain situations when simple blood hormone level estimation is not enough. Two types of dynamic tests are:
Suppression type of dynamic tests are useful in certain conditions, e.g. to know whether a lung cancer is secreting ACTH.
Stimulation type of dynamic tests are useful in certain other conditions, e.g. metyrapone test is performed to know whether the corticotrophs of the pituitary (which secrete ACTH) are normally functioning or not.