Principles of Gas Exchange; Diffusion of Oxygen and Carbon Dioxide Through the Respiratory Membrane
After the alveoli are ventilated with fresh air, the next step in respiration is diffusion of oxygen (O2) from the alveoli into the pulmonary blood and diffusion of carbon dioxide (CO2) in the opposite direction, out of the blood into the alveoli. The process of diffusion is simply the random motion of molecules in all directions through the respiratory membrane and adjacent fluids. However, in respiratory physiology, we are concerned not only with the basic mechanism by which diffusion occurs but also with the rate at which it occurs, which is a much more complex issue, requiring a deeper understanding of the physics of diffusion and gas exchange.
 Physics of Gas Diffusion and Gas Partial Pressures
Physics of Gas Diffusion and Gas Partial Pressures
 Molecular Basis of Gas Diffusion
Molecular Basis of Gas Diffusion
All the gases of concern in respiratory physiology are simple molecules that are free to move among one another by diffusion. This is also true of gases dissolved in the fluids and tissues of the body.
For diffusion to occur, there must be a source of energy. This source of energy is provided by the kinetic motion of the molecules. Except at absolute zero temperature, all molecules of all matter are continually undergoing motion. For free molecules that are not physically attached to others, this means linear movement at high velocity until they strike other molecules. They then bounce away in new directions and continue moving until they strike other molecules again. In this way, the molecules move rapidly and randomly among one another.
Net Diffusion of a Gas in One Direction—Effect of a Concentration Gradient.
If a gas chamber or a solution has a high concentration of a particular gas at one end of the chamber and a low concentration at the other end, as shown in Figure 40-1, net diffusion of the gas will occur from the high-concentration area toward the low-concentration area. The reason is obvious: There are far more molecules at end A of the chamber to diffuse toward end B than there are molecules to diffuse in the opposite direction. Therefore, the rates of diffusion in each of the two directions are proportionately different, as demonstrated by the lengths of the arrows in the figure.

Gas Pressures in a Mixture of Gases—“Partial Pressures” of Individual Gases
Pressure is caused by multiple impacts of moving molecules against a surface. Therefore, the pressure of a gas acting on the surfaces of the respiratory passages and alveoli is proportional to the summated force of impact of all the molecules of that gas striking the surface at any given instant. This means that the pressure is directly proportional to the concentration of the gas molecules.
In respiratory physiology, one deals with mixtures of gases, mainly oxygen, nitrogen, and carbon dioxide. The rate of diffusion of each of these gases is directly proportional to the pressure caused by that gas alone, which is called the partial pressure of that gas. The concept of partial pressure can be explained as follows.
Consider air, which has an approximate composition of 79 percent nitrogen and 21 percent oxygen. The total pressure of this mixture at sea level averages 760 mm Hg. It is clear from the preceding description of the molecular basis of pressure that each gas contributes to the total pressure in direct proportion to its concentration. Therefore, 79 percent of the 760 mm Hg is caused by nitrogen (600 mm Hg) and 21 percent by O2 (160 mm Hg). Thus, the “partial pressure” of nitrogen in the mixture is 600 mm Hg, and the “partial pressure” of O2 is 160 mm Hg; the total pressure is 760 mm Hg, the sum of the individual partial pressures. The partial pressures of individual gases in a mixture are designated by the symbols PO2, PCO2, PN2, PHe, and so forth.
Pressures of Gases Dissolved in Water and Tissues
Gases dissolved in water or in body tissues also exert pressure because the dissolved gas molecules are moving randomly and have kinetic energy. Further, when the gas dissolved in fluid encounters a surface, such as the membrane of a cell, it exerts its own partial pressure in the same way that a gas in the gas phase does. The partial pressures of the separate dissolved gases are designated the same as the partial pressures in the gas state—that is, PO2, PCO2, PN2, PHe, and so forth.
Factors That Determine the Partial Pressure of a Gas Dissolved in a Fluid.
The partial pressure of a gas in a solution is determined not only by its concentration but also by the solubility coefficient of the gas. That is, some types of molecules, especially CO2, are physically or chemically attracted to water molecules, whereas other types of molecules are repelled. When molecules are attracted, far more of them can be dissolved without building up excess partial pressure within the solution. Conversely, in the case of molecules that are repelled, high partial pressure will develop with fewer dissolved molecules. These relations are expressed by the following formula, which is Henry's law:

When partial pressure is expressed in atmospheres (1 atmosphere pressure equals 760 mm Hg) and concentration is expressed in volume of gas dissolved in each volume of water, the solubility coefficients for important respiratory gases at body temperature are the following:
| Oxygen | 0.024 |
| Carbon dioxide | 0.57 |
| Carbon monoxide | 0.018 |
| Nitrogen | 0.012 |
| Helium | 0.008 |
From this table, one can see that CO2 is more than 20 times as soluble as O2. Therefore, the partial pressure of CO2 (for a given concentration) is less than one twentieth that exerted by O2.
Diffusion of Gases Between the Gas Phase in the Alveoli and the Dissolved Phase in the Pulmonary Blood.
The partial pressure of each gas in the alveolar respiratory gas mixture tends to force molecules of that gas into solution in the blood of the alveolar capillaries. Conversely, the molecules of the same gas that are already dissolved in the blood are bouncing randomly in the fluid of the blood, and some of these bouncing molecules escape back into the alveoli. The rate at which they escape is directly proportional to their partial pressure in the blood.
But in which direction will net diffusion of the gas occur? The answer is that net diffusion is determined by the difference between the two partial pressures. If the partial pressure is greater in the gas phase in the alveoli, as is normally true for oxygen, then more molecules will diffuse into the blood than in the other direction. Alternatively, if the partial pressure of the gas is greater in the dissolved state in the blood, which is normally true for CO2, then net diffusion will occur toward the gas phase in the alveoli.
Vapor Pressure of Water
When non-humidified air is breathed into the respiratory passageways, water immediately evaporates from the surfaces of these passages and humidifies the air. This results from the fact that water molecules, like the different dissolved gas molecules, are continually escaping from the water surface into the gas phase. The partial pressure that the water molecules exert to escape through the surface is called the vapor pressure of the water. At normal body temperature, 37°C, this vapor pressure is 47 mm Hg. Therefore, once the gas mixture has become fully humidified—that is, once it is in “equilibrium” with the water—the partial pressure of the water vapor in the gas mixture is 47 mm Hg. This partial pressure, like the other partial pressures, is designated PH2O.
The vapor pressure of water depends entirely on the temperature of the water. The greater the temperature, the greater the kinetic activity of the molecules and, therefore, the greater the likelihood that the water molecules will escape from the surface of the water into the gas phase. For instance, the water vapor pressure at 0°C is 5 mm Hg, and at 100°C it is 760 mm Hg. The most important value to remember is the vapor pressure at body temperature, 47 mm Hg. This value appears in many of our subsequent discussions. 
Pressure Difference Causes Net Diffusion of Gases Through Fluids
From the preceding discussion, it is clear that when the partial pressure of a gas is greater in one area than in another area, there will be net diffusion from the high-pressure area toward the low-pressure area. For instance, returning to Figure 40-1, one can readily see that the molecules in the area of high pressure, because of their greater number, have a greater chance of moving randomly into the area of low pressure than do molecules attempting to go in the other direction. However, some molecules do bounce randomly from the area of low pressure toward the area of high pressure. Therefore, the net diffusion of gas from the area of high pressure to the area of low pressure is equal to the number of molecules bouncing in this forward direction minus the number bouncing in the opposite direction, which is proportional to the gas partial pressure difference between the two areas, called simply the pressure difference for causing diffusion.
Quantifying the Net Rate of Diffusion in Fluids.
In addition to the pressure difference, several other factors affect the rate of gas diffusion in a fluid. They are (1) the solubility of the gas in the fluid, (2) the cross-sectional area of the fluid, (3) the distance through which the gas must diffuse, (4) the molecular weight of the gas, and (5) the temperature of the fluid. In the body, the temperature remains reasonably constant and usually need not be considered.
The greater the solubility of the gas, the greater the number of molecules available to diffuse for any given partial pressure difference. The greater the cross-sectional area of the diffusion pathway, the greater the total number of molecules that diffuse. Conversely, the greater the distance the molecules must diffuse, the longer it will take the molecules to diffuse the entire distance. Finally, the greater the velocity of kinetic movement of the molecules, which is inversely proportional to the square root of the molecular weight, the greater the rate of diffusion of the gas. All these factors can be expressed in a single formula, as follows:
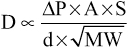
in which D is the diffusion rate, ΔP is the partial pressure difference between the two ends of the diffusion pathway, A is the cross-sectional area of the pathway, S is the solubility of the gas, d is the distance of diffusion, and MW is the molecular weight of the gas.
It is obvious from this formula that the characteristics of the gas determine two factors of the formula: solubility and molecular weight. Together, these two factors determine the diffusion coefficient of the gas, which is proportional to  ; that is, the relative rates at which different gases at the same partial pressure levels will diffuse are proportional to their diffusion coefficients. Assuming that the diffusion coefficient for O2 is 1, the relative diffusion coefficients for different gases of respiratory importance in the body fluids are as follows:
; that is, the relative rates at which different gases at the same partial pressure levels will diffuse are proportional to their diffusion coefficients. Assuming that the diffusion coefficient for O2 is 1, the relative diffusion coefficients for different gases of respiratory importance in the body fluids are as follows:
| Oxygen | 1.0 |
| Carbon dioxide | 20.3 |
| Carbon monoxide | 0.81 |
| Nitrogen | 0.53 |
| Helium | 0.95 |
Diffusion of Gases Through Tissues
The gases that are of respiratory importance are all highly soluble in lipids and, consequently, are highly soluble in cell membranes. Because of this, the major limitation to the movement of gases in tissues is the rate at which the gases can diffuse through the tissue water instead of through the cell membranes. Therefore, diffusion of gases through the tissues, including through the respiratory membrane, is almost equal to the diffusion of gases in water, as given in the preceding list.
Compositions of Alveolar Air and Atmospheric Air are Different
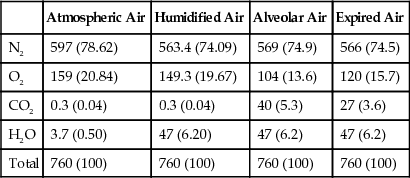
Alveolar air does not have the same concentrations of gases as atmospheric air (Table 40-1). There are several reasons for the differences. First, alveolar air is only partially replaced by atmospheric air with each breath. Second, O2 is constantly being absorbed into the pulmonary blood from the alveolar air. Third, CO2 is constantly diffusing from the pulmonary blood into the alveoli. And fourth, dry atmospheric air that enters the respiratory passages is humidified even before it reaches the alveoli.
Table 40-1
Partial Pressures of Respiratory Gases (in mm Hg) as They Enter and Leave the Lungs (at Sea Level)
| Atmospheric Air | Humidified Air | Alveolar Air | Expired Air | |
| N2 | 597 (78.62) | 563.4 (74.09) | 569 (74.9) | 566 (74.5) |
| O2 | 159 (20.84) | 149.3 (19.67) | 104 (13.6) | 120 (15.7) |
| CO2 | 0.3 (0.04) | 0.3 (0.04) | 40 (5.3) | 27 (3.6) |
| H2O | 3.7 (0.50) | 47 (6.20) | 47 (6.2) | 47 (6.2) |
| Total | 760 (100) | 760 (100) | 760 (100) | 760 (100) |
Humidification of the Air in the Respiratory Passages
Table 40-1 shows that atmospheric air is composed almost entirely of nitrogen and O2; it normally contains almost no CO2 and little water vapor. However, as soon as the atmospheric air enters the respiratory passages, it is exposed to the fluids that cover the respiratory surfaces. Even before the air enters the alveoli, it becomes almost totally humidified.
The partial pressure of water vapor at a normal body temperature of 37°C is 47 mm Hg, which is therefore the partial pressure of water vapor in the alveolar air. Because the total pressure in the alveoli cannot rise to more than the atmospheric pressure (760 mm Hg at sea level), this water vapor simply dilutes all the other gases in the inspired air. Table 40-1 also shows that humidification of the air dilutes the oxygen partial pressure at sea level from an average of 159 mm Hg in atmospheric air to 149 mm Hg in the humidified air, and it dilutes the nitrogen partial pressure from 597 to 563 mm Hg.
Alveolar Air is Slowly Renewed by Atmospheric Air
In Chapter 38, we pointed out that the average male functional residual capacity of the lungs (the volume of air remaining in the lungs at the end of normal expiration) measures about 2300 milliliters. Yet only 350 milliliters of new air is brought into the alveoli with each normal inspiration, and this same amount of old alveolar air is expired. Therefore, the volume of alveolar air replaced by new atmospheric air with each breath is only one seventh of the total, so multiple breaths are required to exchange most of the alveolar air. Figure 40-2 shows this slow rate of renewal of the alveolar air. In the first alveolus of the figure, excess gas is present in the alveoli but note that even at the end of 16 breaths the excess gas still has not been completely removed from the alveoli.

Figure 40-3 demonstrates graphically the rate at which excess gas in the alveoli is normally removed, showing that with normal alveolar ventilation, about one half the gas is removed in 17 seconds. When a person's rate of alveolar ventilation is only one-half normal, one half the gas is removed in 34 seconds, and when the rate of ventilation is twice normal, one half is removed in about 8 seconds.

Importance of the Slow Replacement of Alveolar Air.
The slow replacement of alveolar air is of particular importance in preventing sudden changes in gas concentrations in the blood. This makes the respiratory control mechanism much more stable than it would be otherwise, and it helps prevent excessive increases and decreases in tissue oxygenation, tissue CO2 concentration, and tissue pH when respiration is temporarily interrupted.
Oxygen Concentration and Partial Pressure in the Alveoli
Oxygen is continually being absorbed from the alveoli into the blood of the lungs, and new O2 is continually being breathed into the alveoli from the atmosphere. The more rapidly O2 is absorbed, the lower its concentration in the alveoli becomes; conversely, the more rapidly new O2 is breathed into the alveoli from the atmosphere, the higher its concentration becomes. Therefore, O2 concentration in the alveoli, as well as its partial pressure, is controlled by (1) the rate of absorption of O2 into the blood and (2) the rate of entry of new O2 into the lungs by the ventilatory process.
Figure 40-4 shows the effect of alveolar ventilation and rate of O2 absorption into the blood on the alveolar partial pressure of O2 (PO2). One curve represents O2 absorption at a rate of 250 ml/min, and the other curve represents a rate of 1000 ml/min. At a normal ventilatory rate of 4.2 L/min and an O2 consumption of 250 ml/min, the normal operating point in Figure 40-4 is point A. The figure also shows that when 1000 milliliters of O2 are being absorbed each minute, as occurs during moderate exercise, the rate of alveolar ventilation must increase fourfold to maintain the alveolar Po2 at the normal value of 104 mm Hg.
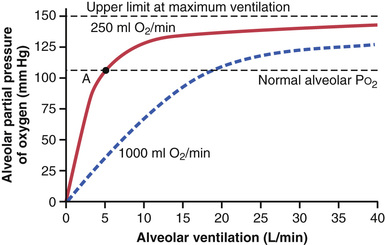
Another effect shown in Figure 40-4 is that even an extreme increase in alveolar ventilation can never increase the alveolar PO2 above 149 mm Hg as long as the person is breathing normal atmospheric air at sea level pressure, because 149 mm Hg is the maximum PO2 in humidified air at this pressure. If the person breathes gases that contain partial pressures of O2 higher than 149 mm Hg, the alveolar PO2 can approach these higher pressures at high rates of ventilation.
CO2 Concentration and Partial Pressure in the Alveoli
Carbon dioxide is continually formed in the body and then carried in the blood to the alveoli, and it is continually removed from the alveoli by ventilation. Figure 40-5 shows the effects on the alveolar partial pressure of CO2 (PCO2) of both alveolar ventilation and two rates of CO2 excretion, 200 and 800 ml/min. One curve represents a normal rate of CO2 excretion of 200 ml/min. At the normal rate of alveolar ventilation of 4.2 L/min, the operating point for alveolar PCO2 is at point A in Figure 40-5 (i.e., 40 mm Hg).
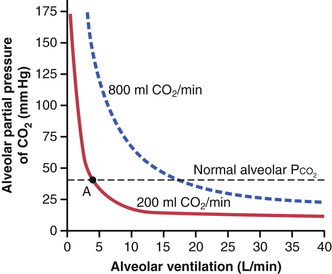
Two other facts are also evident from Figure 40-5: First, the alveolar PCO2 increases directly in proportion to the rate of CO2 excretion, as represented by the fourfold elevation of the curve (when 800 milliliters of CO2 are excreted per minute). Second, the alveolar PCO2 decreases in inverse proportion to alveolar ventilation. Therefore, the concentrations and partial pressures of both O2 and CO2 in the alveoli are determined by the rates of absorption or excretion of the two gases and by the amount of alveolar ventilation.
Expired Air Is a Combination of Dead Space Air and Alveolar Air
The overall composition of expired air is determined by (1) the amount of the expired air that is dead space air and (2) the amount that is alveolar air. Figure 40-6 shows the progressive changes in O2 and CO2 partial pressures in the expired air during the course of expiration. The first portion of this air, the dead space air from the respiratory passageways, is typical humidified air, as shown in Table 40-1. Then, progressively more and more alveolar air becomes mixed with the dead space air until all the dead space air has finally been washed out and nothing but alveolar air is expired at the end of expiration. Therefore, the method of collecting alveolar air for study is simply to collect a sample of the last portion of the expired air after forceful expiration has removed all the dead space air.
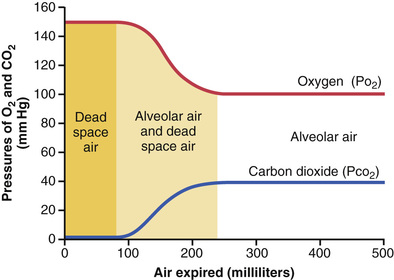
Normal expired air, containing both dead space air and alveolar air, has gas concentrations and partial pressures approximately as shown in Table 40-1 (i.e., concentrations between those of alveolar air and humidified atmospheric air).
Diffusion of Gases Through the Respiratory Membrane
Respiratory Unit.
Figure 40-7 shows the respiratory unit (also called “respiratory lobule”), which is composed of a respiratory bronchiole, alveolar ducts, atria, and alveoli. There are about 300 million alveoli in the two lungs, and each alveolus has an average diameter of about 0.2 millimeter. The alveolar walls are extremely thin, and between the alveoli is an almost solid network of interconnecting capillaries, shown in Figure 40-8. Indeed, because of the extensiveness of the capillary plexus, the flow of blood in the alveolar wall has been described as a “sheet” of flowing blood. Thus, it is obvious that the alveolar gases are in very close proximity to the blood of the pulmonary capillaries. Further, gas exchange between the alveolar air and the pulmonary blood occurs through the membranes of all the terminal portions of the lungs, not merely in the alveoli. All these membranes are collectively known as the respiratory membrane, also called the pulmonary membrane.

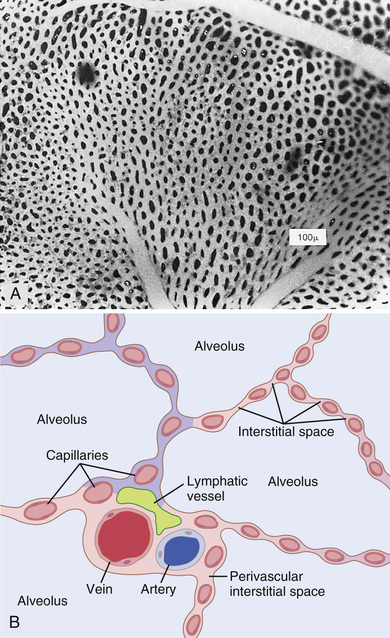
Respiratory Membrane.
Figure 40-9 shows the ultrastructure of the respiratory membrane drawn in cross section on the left and a red blood cell on the right. It also shows the diffusion of O2 from the alveolus into the red blood cell and diffusion of CO2 in the opposite direction. Note the following different layers of the respiratory membrane:
1. A layer of fluid containing surfactant that lines the alveolus and reduces the surface tension of the alveolar fluid
2. The alveolar epithelium, which is composed of thin epithelial cells
3. An epithelial basement membrane
4. A thin interstitial space between the alveolar epithelium and the capillary membrane
5. A capillary basement membrane that in many places fuses with the alveolar epithelial basement membrane

Despite the large number of layers, the overall thickness of the respiratory membrane in some areas is as little as 0.2 micrometer and averages about 0.6 micrometer, except where there are cell nuclei. From histological studies, it has been estimated that the total surface area of the respiratory membrane is about 70 square meters in the healthy adult human male, which is equivalent to the floor area of a 25 × 30 foot room. The total quantity of blood in the capillaries of the lungs at any given instant is 60 to 140 milliliters. Now imagine this small amount of blood spread over the entire surface of a 25 × 30 foot floor, and it is easy to understand the rapidity of the respiratory exchange of O2 and CO2.
The average diameter of the pulmonary capillaries is only about 5 micrometers, which means that red blood cells must squeeze through them. The red blood cell membrane usually touches the capillary wall, so O2 and CO2 need not pass through significant amounts of plasma as they diffuse between the alveolus and the red blood cell. This, too, increases the rapidity of diffusion.
Factors That Affect the Rate of Gas Diffusion Through the Respiratory Membrane
Referring to the earlier discussion of diffusion of gases in water, one can apply the same principles to diffusion of gases through the respiratory membrane. Thus, the factors that determine how rapidly a gas will pass through the membrane are (1) the thickness of the membrane, (2) the surface area of the membrane, (3) the diffusion coefficient of the gas in the substance of the membrane, and (4) the partial pressure difference of the gas between the two sides of the membrane.
The thickness of the respiratory membrane occasionally increases—for instance, as a result of edema fluid in the interstitial space of the membrane and in the alveoli—so the respiratory gases must then diffuse not only through the membrane but also through this fluid. Also, some pulmonary diseases cause fibrosis of the lungs, which can increase the thickness of some portions of the respiratory membrane. Because the rate of diffusion through the membrane is inversely proportional to the thickness of the membrane, any factor that increases the thickness to more than two to three times normal can interfere significantly with normal respiratory exchange of gases.
The surface area of the respiratory membrane can be greatly decreased by many conditions. For instance, removal of an entire lung decreases the total surface area to one-half normal. Also, in emphysema, many of the alveoli coalesce, with dissolution of many alveolar walls. Therefore, the new alveolar chambers are much larger than the original alveoli, but the total surface area of the respiratory membrane is often decreased as much as fivefold because of loss of the alveolar walls. When the total surface area is decreased to about one-third to one-fourth normal, exchange of gases through the membrane is substantially impeded, even under resting conditions, and during competitive sports and other strenuous exercise even the slightest decrease in surface area of the lungs can be a serious detriment to respiratory exchange of gases.
The diffusion coefficient for transfer of each gas through the respiratory membrane depends on the gas's solubility in the membrane and, inversely, on the square root of the gas's molecular weight. The rate of diffusion in the respiratory membrane is almost exactly the same as that in water, for reasons explained earlier. Therefore, for a given pressure difference, CO2 diffuses about 20 times as rapidly as O2. Oxygen diffuses about twice as rapidly as nitrogen.
The pressure difference across the respiratory membrane is the difference between the partial pressure of the gas in the alveoli and the partial pressure of the gas in the pulmonary capillary blood. The partial pressure represents a measure of the total number of molecules of a particular gas striking a unit area of the alveolar surface of the membrane in unit time, and the pressure of the gas in the blood represents the number of molecules that attempt to escape from the blood in the opposite direction. Therefore, the difference between these two pressures is a measure of the net tendency for the gas molecules to move through the membrane.
When the partial pressure of a gas in the alveoli is greater than the pressure of the gas in the blood, as is true for O2, net diffusion from the alveoli into the blood occurs; when the pressure of the gas in the blood is greater than the partial pressure in the alveoli, as is true for CO2, net diffusion from the blood into the alveoli occurs.
Diffusing Capacity of the Respiratory Membrane
The ability of the respiratory membrane to exchange a gas between the alveoli and the pulmonary blood is expressed in quantitative terms by the respiratory membrane's diffusing capacity, which is defined as the volume of a gas that will diffuse through the membrane each minute for a partial pressure difference of 1 mm Hg. All the factors discussed earlier that affect diffusion through the respiratory membrane can affect this diffusing capacity.
Diffusing Capacity for Oxygen.
In the average young man, the diffusing capacity for O2 under resting conditions averages 21 ml/min/mm Hg. In functional terms, what does this mean? The mean O2 pressure difference across the respiratory membrane during normal, quiet breathing is about 11 mm Hg. Multiplication of this pressure by the diffusing capacity (11 × 21) gives a total of about 230 milliliters of oxygen diffusing through the respiratory membrane each minute, which is equal to the rate at which the resting body uses O2.
Increased Oxygen Diffusing Capacity during Exercise.
During strenuous exercise or other conditions that greatly increase pulmonary blood flow and alveolar ventilation, the diffusing capacity for O2 increases in young men to a maximum of about 65 ml/min/mm Hg, which is three times the diffusing capacity under resting conditions. This increase is caused by several factors, among which are (1) opening up of many previously dormant pulmonary capillaries or extra dilation of already open capillaries, thereby increasing the surface area of the blood into which the O2 can diffuse, and (2) a better match between the ventilation of the alveoli and the perfusion of the alveolar capillaries with blood, called the ventilation-perfusion ratio, which is explained later in this chapter. Therefore, during exercise, oxygenation of the blood is increased not only by increased alveolar ventilation but also by greater diffusing capacity of the respiratory membrane for transporting O2 into the blood.
Diffusing Capacity for Carbon Dioxide.
The diffusing capacity for CO2 has never been measured because CO2 diffuses through the respiratory membrane so rapidly that the average PCO2 in the pulmonary blood is not far different from the PCO2 in the alveoli—the average difference is less than 1 mm Hg. With currently available techniques, this difference is too small to be measured.
Nevertheless, measurements of diffusion of other gases have shown that the diffusing capacity varies directly with the diffusion coefficient of the particular gas. Because the diffusion coefficient of CO2 is slightly more than 20 times that of O2, one would expect a diffusing capacity for CO2 under resting conditions of about 400 to 450 ml/min/mm Hg and during exercise of about 1200 to 1300 ml/min/mm Hg. Figure 40-10 compares the measured or calculated diffusing capacities of carbon monoxide, O2, and CO2 at rest and during exercise, showing the extreme diffusing capacity of CO2 and the effect of exercise on the diffusing capacity of each of these gases.

Measurement of Diffusing Capacity—The Carbon Monoxide Method.
The O2 diffusing capacity can be calculated from measurements of (1) alveolar PO2, (2) PO2 in the pulmonary capillary blood, and (3) the rate of O2 uptake by the blood. However, measuring the PO2 in the pulmonary capillary blood is so difficult and imprecise that it is not practical to measure oxygen diffusing capacity by such a direct procedure, except on an experimental basis.
To obviate the difficulties encountered in measuring oxygen diffusing capacity directly, physiologists usually measure carbon monoxide (CO) diffusing capacity instead and then calculate the O2 diffusing capacity from this. The principle of the CO method is the following: A small amount of CO is breathed into the alveoli, and the partial pressure of the CO in the alveoli is measured from appropriate alveolar air samples. The CO pressure in the blood is essentially zero because hemoglobin combines with this gas so rapidly that its pressure never has time to build up. Therefore, the pressure difference of CO across the respiratory membrane is equal to its partial pressure in the alveolar air sample. Then, by measuring the volume of CO absorbed in a short period and dividing this by the alveolar CO partial pressure, one can determine accurately the CO diffusing capacity.
To convert CO diffusing capacity to O2 diffusing capacity, the value is multiplied by a factor of 1.23 because the diffusion coefficient for O2 is 1.23 times that for CO. Thus, the average diffusing capacity for CO in healthy young men at rest is 17 ml/min/mm Hg, and the diffusing capacity for O2 is 1.23 times this, or 21 ml/min/mm Hg.
Effect of the Ventilation-Perfusion Ratio on Alveolar Gas Concentration
Earlier in this chapter, we learned that two factors determine the PO2 and the PCO2 in the alveoli: (1) the rate of alveolar ventilation and (2) the rate of transfer of O2 and CO2 through the respiratory membrane. These discussions made the assumption that all the alveoli are ventilated equally and that blood flow through the alveolar capillaries is the same for each alveolus. However, even normally to some extent, and especially in many lung diseases, some areas of the lungs are well ventilated but have almost no blood flow, whereas other areas may have excellent blood flow but little or no ventilation. In either of these conditions, gas exchange through the respiratory membrane is seriously impaired, and the person may suffer severe respiratory distress despite both normal total ventilation and normal total pulmonary blood flow, but with the ventilation and blood flow going to different parts of the lungs. Therefore, a highly quantitative concept has been developed to help us understand respiratory exchange when there is imbalance between alveolar ventilation and alveolar blood flow. This concept is called the ventilation-perfusion ratio.
In quantitative terms, the ventilation-perfusion ratio is expressed as  . When
. When  (alveolar ventilation) is normal for a given alveolus and
(alveolar ventilation) is normal for a given alveolus and  (blood flow) is also normal for the same alveolus, the ventilation-perfusion ratio (
(blood flow) is also normal for the same alveolus, the ventilation-perfusion ratio ( ) is also said to be normal. When the ventilation (
) is also said to be normal. When the ventilation ( ) is zero, yet there is still perfusion (
) is zero, yet there is still perfusion ( ) of the alveolus, the
) of the alveolus, the  is zero. Or, at the other extreme, when there is adequate ventilation (
is zero. Or, at the other extreme, when there is adequate ventilation ( ) but zero perfusion (
) but zero perfusion ( ), the ratio
), the ratio  is infinity. At a ratio of either zero or infinity, there is no exchange of gases through the respiratory membrane of the affected alveoli, which explains the importance of this concept. Therefore, let us explain the respiratory consequences of these two extremes.
is infinity. At a ratio of either zero or infinity, there is no exchange of gases through the respiratory membrane of the affected alveoli, which explains the importance of this concept. Therefore, let us explain the respiratory consequences of these two extremes.
Alveolar Oxygen and Carbon Dioxide Partial Pressures When  Equals Zero.
Equals Zero.
When  is equal to zero—that is, without any alveolar ventilation—the air in the alveolus comes to equilibrium with the blood O2 and CO2 because these gases diffuse between the blood and the alveolar air. Because the blood that perfuses the capillaries is venous blood returning to the lungs from the systemic circulation, it is the gases in this blood with which the alveolar gases equilibrate. In Chapter 41, we describe how the normal venous blood (
is equal to zero—that is, without any alveolar ventilation—the air in the alveolus comes to equilibrium with the blood O2 and CO2 because these gases diffuse between the blood and the alveolar air. Because the blood that perfuses the capillaries is venous blood returning to the lungs from the systemic circulation, it is the gases in this blood with which the alveolar gases equilibrate. In Chapter 41, we describe how the normal venous blood ( ) has a PO2 of 40 mm Hg and a PCO2 of 45 mm Hg. Therefore, these are also the normal partial pressures of these two gases in alveoli that have blood flow but no ventilation.
) has a PO2 of 40 mm Hg and a PCO2 of 45 mm Hg. Therefore, these are also the normal partial pressures of these two gases in alveoli that have blood flow but no ventilation.
Alveolar Oxygen and Carbon Dioxide Partial Pressures When  Equals Infinity.
Equals Infinity.
The effect on the alveolar gas partial pressures when  equals infinity is entirely different from the effect when
equals infinity is entirely different from the effect when  equals zero because now there is no capillary blood flow to carry O2 away or to bring CO2 to the alveoli. Therefore, instead of the alveolar gases coming to equilibrium with the venous blood, the alveolar air becomes equal to the humidified inspired air. That is, the air that is inspired loses no O2 to the blood and gains no CO2 from the blood. Furthermore, because normal inspired and humidified air has a PO2 of 149 mm Hg and a PCO2 of 0 mm Hg, these will be the partial pressures of these two gases in the alveoli.
equals zero because now there is no capillary blood flow to carry O2 away or to bring CO2 to the alveoli. Therefore, instead of the alveolar gases coming to equilibrium with the venous blood, the alveolar air becomes equal to the humidified inspired air. That is, the air that is inspired loses no O2 to the blood and gains no CO2 from the blood. Furthermore, because normal inspired and humidified air has a PO2 of 149 mm Hg and a PCO2 of 0 mm Hg, these will be the partial pressures of these two gases in the alveoli.
Gas Exchange and Alveolar Partial Pressures When  Is Normal.
Is Normal.
When there is both normal alveolar ventilation and normal alveolar capillary blood flow (normal alveolar perfusion), exchange of O2 and CO2 through the respiratory membrane is nearly optimal, and alveolar PO2 is normally at a level of 104 mm Hg, which lies between that of the inspired air (149 mm Hg) and that of venous blood (40 mm Hg). Likewise, alveolar PCO2 lies between two extremes; it is normally 40 mm Hg, in contrast to 45 mm Hg in venous blood and 0 mm Hg in inspired air. Thus, under normal conditions, the alveolar air PO2 averages 104 mm Hg and the PCO2 averages 40 mm Hg.
PO2-PCO2,  Diagram
Diagram
The concepts presented in the preceding sections can be shown in graphical form, as demonstrated in Figure 40-11, called the Po2-Pco2,  diagram. The curve in the diagram represents all possible PO2 and PCO2 combinations between the limits of
diagram. The curve in the diagram represents all possible PO2 and PCO2 combinations between the limits of  equals zero and
equals zero and  equals infinity when the gas pressures in the venous blood are normal and the person is breathing air at sea-level pressure. Thus, point
equals infinity when the gas pressures in the venous blood are normal and the person is breathing air at sea-level pressure. Thus, point  is the plot of PO2 and PCO2 when
is the plot of PO2 and PCO2 when  equals zero. At this point, the PO2 is 40 mm Hg and the PCO2 is 45 mm Hg, which are the values in normal venous blood.
equals zero. At this point, the PO2 is 40 mm Hg and the PCO2 is 45 mm Hg, which are the values in normal venous blood.
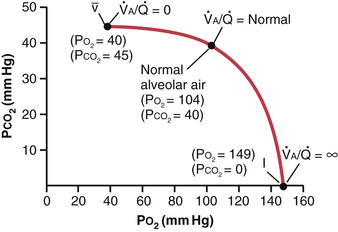

 ) diagram.
) diagram.At the other end of the curve, when  equals infinity, point I represents inspired air, showing PO2 to be 149 mm Hg while PCO2 is zero. Also plotted on the curve is the point that represents normal alveolar air when
equals infinity, point I represents inspired air, showing PO2 to be 149 mm Hg while PCO2 is zero. Also plotted on the curve is the point that represents normal alveolar air when  is normal. At this point, PO2 is 104 mm Hg and PCO2 is 40 mm Hg.
is normal. At this point, PO2 is 104 mm Hg and PCO2 is 40 mm Hg.
Concept of “Physiological Shunt” (When  Is Below Normal)
Is Below Normal)
Whenever  is below normal, there is inadequate ventilation to provide the O2 needed to fully oxygenate the blood flowing through the alveolar capillaries. Therefore, a certain fraction of the venous blood passing through the pulmonary capillaries does not become oxygenated. This fraction is called shunted blood. Also, some additional blood flows through bronchial vessels rather than through alveolar capillaries, normally about 2 percent of the cardiac output; this, too, is unoxygenated, shunted blood.
is below normal, there is inadequate ventilation to provide the O2 needed to fully oxygenate the blood flowing through the alveolar capillaries. Therefore, a certain fraction of the venous blood passing through the pulmonary capillaries does not become oxygenated. This fraction is called shunted blood. Also, some additional blood flows through bronchial vessels rather than through alveolar capillaries, normally about 2 percent of the cardiac output; this, too, is unoxygenated, shunted blood.
The total quantitative amount of shunted blood per minute is called the physiological shunt. This physiological shunt is measured in clinical pulmonary function laboratories by analyzing the concentration of O2 in both mixed venous blood and arterial blood, along with simultaneous measurement of cardiac output. From these values, the physiological shunt can be calculated by the following equation:
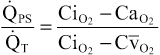
in which  is the physiological shunt blood flow per minute,
is the physiological shunt blood flow per minute,  is cardiac output per minute, CiO2 is the concentration of oxygen in the arterial blood if there is an “ideal” ventilation-perfusion ratio, CaO2 is the measured concentration of oxygen in the arterial blood, and
is cardiac output per minute, CiO2 is the concentration of oxygen in the arterial blood if there is an “ideal” ventilation-perfusion ratio, CaO2 is the measured concentration of oxygen in the arterial blood, and  is the measured concentration of oxygen in the mixed venous blood.
is the measured concentration of oxygen in the mixed venous blood.
The greater the physiological shunt, the greater the amount of blood that fails to be oxygenated as it passes through the lungs.
Concept of the “Physiological Dead Space” (When  Is Greater Than Normal)
Is Greater Than Normal)
When ventilation of some of the alveoli is great but alveolar blood flow is low, there is far more available oxygen in the alveoli than can be transported away from the alveoli by the flowing blood. Thus, the ventilation of these alveoli is said to be wasted. The ventilation of the anatomical dead space areas of the respiratory passageways is also wasted. The sum of these two types of wasted ventilation is called the physiological dead space. This space is measured in the clinical pulmonary function laboratory by making appropriate blood and expiratory gas measurements and using the following equation, called the Bohr equation:
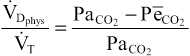
in which  is the physiological dead space,
is the physiological dead space,  is the tidal volume, Paco2 is the partial pressure of CO2 in the arterial blood, and PeCO2 is the average partial pressure of CO2 in the entire expired air.
is the tidal volume, Paco2 is the partial pressure of CO2 in the arterial blood, and PeCO2 is the average partial pressure of CO2 in the entire expired air.
When the physiological dead space is great, much of the work of ventilation is wasted effort because so much of the ventilating air never reaches the blood.
Abnormalities of Ventilation-Perfusion Ratio
Abnormal  in the Upper and Lower Normal Lung.
in the Upper and Lower Normal Lung.
In a normal person in the upright position, both pulmonary capillary blood flow and alveolar ventilation are considerably less in the upper part of the lung than in the lower part; however, the decrease of blood flow is considerably greater than the decrease in ventilation. Therefore, at the top of the lung,  is as much as 2.5 times as great as the ideal value, which causes a moderate degree of physiological dead space in this area of the lung.
is as much as 2.5 times as great as the ideal value, which causes a moderate degree of physiological dead space in this area of the lung.
At the other extreme, in the bottom of the lung, there is slightly too little ventilation in relation to blood flow, with  as low as 0.6 times the ideal value. In this area, a small fraction of the blood fails to become normally oxygenated, and this represents a physiological shunt.
as low as 0.6 times the ideal value. In this area, a small fraction of the blood fails to become normally oxygenated, and this represents a physiological shunt.
In both extremes, inequalities of ventilation and perfusion decrease slightly the lung's effectiveness for exchanging O2 and CO2. However, during exercise, blood flow to the upper part of the lung increases markedly, so far less physiological dead space occurs, and the effectiveness of gas exchange now approaches optimum.
Abnormal  in Chronic Obstructive Lung Disease.
in Chronic Obstructive Lung Disease.
Most people who smoke for many years develop various degrees of bronchial obstruction; in a large share of these persons, this condition eventually becomes so severe that serious alveolar air trapping develops, with resultant emphysema. The emphysema in turn causes many of the alveolar walls to be destroyed. Thus, two abnormalities occur in smokers to cause abnormal  . First, because many of the small bronchioles are obstructed, the alveoli beyond the obstructions are unventilated, causing a
. First, because many of the small bronchioles are obstructed, the alveoli beyond the obstructions are unventilated, causing a  that approaches zero. Second, in the areas of the lung where the alveolar walls have been mainly destroyed but there is still alveolar ventilation, most of the ventilation is wasted because of inadequate blood flow to transport the blood gases.
that approaches zero. Second, in the areas of the lung where the alveolar walls have been mainly destroyed but there is still alveolar ventilation, most of the ventilation is wasted because of inadequate blood flow to transport the blood gases.
Thus, in chronic obstructive lung disease, some areas of the lung exhibit serious physiological shunt, and other areas exhibit serious physiological dead space. Both conditions tremendously decrease the effectiveness of the lungs as gas exchange organs, sometimes reducing their effectiveness to as little as one-tenth normal. In fact, this condition is the most prevalent cause of pulmonary disability today.