Secretory Functions of the Alimentary Tract
Throughout the gastrointestinal tract, secretory glands subserve two primary functions: First, digestive enzymes are secreted in most areas of the alimentary tract, from the mouth to the distal end of the ileum. Second, mucous glands located from the mouth to the anus provide mucus for lubrication and protection of all parts of the alimentary tract.
Most digestive secretions are formed in response to the presence of food in the alimentary tract, and the quantity secreted in each segment of the tract is usually the amount needed for proper digestion. Furthermore, in some portions of the gastrointestinal tract, even the types of enzymes and other constituents of the secretions are varied in accordance with the types of food present. The purpose of this chapter is to describe the different alimentary secretions, their functions, and regulation of their production.
General Principles of Alimentary Tract Secretion
 Types of Alimentary Tract Glands
Types of Alimentary Tract Glands
Several types of glands provide the different types of alimentary tract secretions. First, on the surface of the epithelium in most parts of the gastrointestinal tract are billions of single-cell mucous glands called simply mucous cells or sometimes goblet cells because they look like goblets. They function mainly in response to local irritation of the epithelium: They extrude mucus directly onto the epithelial surface to act as a lubricant that also protects the surfaces from excoriation and digestion.
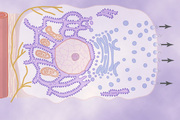
Second, many surface areas of the gastrointestinal tract are lined by pits that represent invaginations of the epithelium into the submucosa. In the small intestine, these pits, called crypts of Lieberkühn, are deep and contain specialized secretory cells. One of these cells is shown in Figure 65-1.
Third, in the stomach and upper duodenum are large numbers of deep tubular glands. A typical tubular gland can be seen in Figure 65-4, which shows an acid- and pepsinogen-secreting gland of the stomach (oxyntic gland).
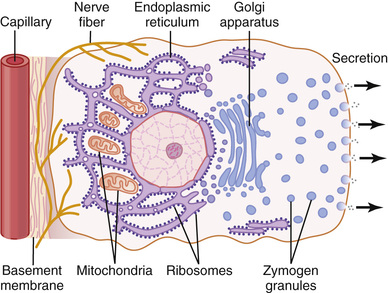
Fourth, also associated with the alimentary tract are several complex glands—the salivary glands, pancreas, and liver—that provide secretions for digestion or emulsification of food. The liver has a highly specialized structure that is discussed in Chapter 71. The salivary glands and the pancreas are compound acinous glands of the type shown in Figure 65-2. These glands lie outside the walls of the alimentary tract and, in this aspect, they differ from all other alimentary glands. They contain millions of acini lined with secreting glandular cells; these acini feed into a system of ducts that finally empty into the alimentary tract itself. 

Basic Mechanisms of Stimulation of the Alimentary Tract Glands
Contact of Food with the Epithelium Stimulates Secretion—Function of Enteric Nervous Stimuli
The presence of food in a particular segment of the gastrointestinal tract usually stimulates the glands of that region and adjacent regions to secrete moderate to large quantities of juices. Part of this local effect, especially the secretion of mucus by mucous cells, results from direct contact stimulation of the surface glandular cells by the food.
In addition, local epithelial stimulation also activates the enteric nervous system of the gut wall. The types of stimuli that activate this system are (1) tactile stimulation, (2) chemical irritation, and (3) distention of the gut wall. The resulting nervous reflexes stimulate both the mucous cells on the gut epithelial surface and the deep glands in the gut wall to increase their secretion.
Autonomic Stimulation of Secretion
Parasympathetic Stimulation Increases the Alimentary Tract Glandular Secretion Rate.
Stimulation of the parasympathetic nerves to the alimentary tract almost invariably increases the rates of alimentary glandular secretion. This increased secretion rate is especially true of the glands in the upper portion of the tract (innervated by the glossopharyngeal and vagus parasympathetic nerves) such as the salivary glands, esophageal glands, gastric glands, pancreas, and Brunner's glands in the duodenum. It is also true of some glands in the distal portion of the large intestine, which are innervated by pelvic parasympathetic nerves. Secretion in the remainder of the small intestine and in the first two thirds of the large intestine occurs mainly in response to local neural and hormonal stimuli in each segment of the gut.
Sympathetic Stimulation Has a Dual Effect on the Alimentary Tract Glandular Secretion Rate.
Stimulation of the sympathetic nerves going to the gastrointestinal tract causes a slight to moderate increase in secretion by some of the local glands. However, sympathetic stimulation also results in constriction of the blood vessels that supply the glands. Therefore, sympathetic stimulation can have a dual effect: (1) sympathetic stimulation alone usually slightly increases secretion and (2) if parasympathetic or hormonal stimulation is already causing copious secretion by the glands, superimposed sympathetic stimulation usually reduces the secretion, sometimes significantly so, mainly because of vasoconstrictive reduction of the blood supply.
Regulation of Glandular Secretion by Hormones.
In the stomach and intestine, several different gastrointestinal hormones help regulate the volume and character of the secretions. These hormones are liberated from the gastrointestinal mucosa in response to the presence of food in the lumen of the gut. The hormones are then absorbed into the blood and carried to the glands, where they stimulate secretion. This type of stimulation is particularly valuable to increase the output of gastric juice and pancreatic juice when food enters the stomach or duodenum.
Chemically, the gastrointestinal hormones are polypeptides or polypeptide derivatives and will be discussed in more detail later.
Basic Mechanism of Secretion by Glandular Cells
Secretion of Organic Substances.
Although all the basic mechanisms by which glandular cells function are not known, experimental evidence points to the following principles of secretion, as shown in Figure 65-1.
1. The nutrient material needed for formation of the secretion must first diffuse or be actively transported by the blood in the capillaries into the base of the glandular cell.
2. Many mitochondria located inside the glandular cell near its base use oxidative energy to form adenosine triphosphate (ATP).
3. Energy from the ATP, along with appropriate substrates provided by the nutrients, is then used to synthesize the organic secretory substances; this synthesis occurs almost entirely in the endoplasmic reticulum and Golgi complex of the glandular cell. Ribosomes adherent to the reticulum are specifically responsible for formation of the proteins that are secreted.
4. The secretory materials are transported through the tubules of the endoplasmic reticulum, passing in about 20 minutes all the way to the vesicles of the Golgi complex.
5. In the Golgi complex, the materials are modified, added to, concentrated, and discharged into the cytoplasm in the form of secretory vesicles, which are stored in the apical ends of the secretory cells.
6. These vesicles remain stored until nervous or hormonal control signals cause the cells to extrude the vesicular contents through the cells' surface. This action probably occurs in the following way: The hormone binds to its receptor and, through one of several possible cell signaling mechanisms, increases the cell membrane permeability to calcium ions. Calcium enters the cell and causes many of the vesicles to fuse with the apical cell membrane. The apical cell membrane then breaks open, thus emptying the vesicles to the exterior; this process is called exocytosis.
Water and Electrolyte Secretion.
A second necessity for glandular secretion is secretion of sufficient water and electrolytes to go along with the organic substances. Secretion by the salivary glands, discussed in more detail later, provides an example of how nervous stimulation causes water and salts to pass through the glandular cells in great profusion, washing the organic substances through the secretory border of the cells at the same time. It is believed that hormones acting on the cell membrane of some glandular cells also cause secretory effects similar to those caused by nervous stimulation.
Lubricating and Protective Properties of Mucus, and the Importance of Mucus in the Gastrointestinal Tract
Mucus is a thick secretion composed mainly of water, electrolytes, and a mixture of several glycoproteins that are composed of large polysaccharides bound with much smaller quantities of protein. Mucus is slightly different in different parts of the gastrointestinal tract, but in all locations it has several important characteristics that make it both an excellent lubricant and a protectant for the wall of the gut. First, mucus has adherent qualities that make it adhere tightly to the food or other particles and to spread as a thin film over the surfaces. Second, it has sufficient body that it coats the wall of the gut and prevents actual contact of most food particles with the mucosa. Third, mucus has a low resistance for slippage, so the particles can slide along the epithelium with great ease. Fourth, mucus causes fecal particles to adhere to one another to form the feces that are expelled during a bowel movement. Fifth, mucus is strongly resistant to digestion by the gastrointestinal enzymes. And sixth, the glycoproteins of mucus have amphoteric properties, which means that they are capable of buffering small amounts of either acids or alkalies; also, mucus often contains moderate quantities of bicarbonate ions, which specifically neutralize acids.
Thus, mucus has the ability to allow easy slippage of food along the gastrointestinal tract and to prevent excoriative or chemical damage to the epithelium. A person becomes acutely aware of the lubricating qualities of mucus when the salivary glands fail to secrete saliva, because then it is difficult to swallow solid food even when it is eaten along with large amounts of water.
Secretion of Saliva
Saliva Contains a Serous Secretion and a Mucus Secretion.
The principal glands of salivation are the parotid, submandibular, and sublingual glands; in addition, there are many tiny buccal glands. Daily secretion of saliva normally ranges between 800 and 1500 milliliters, as shown by the average value of 1000 milliliters in Table 65-1.
Table 65-1
Daily Secretion of Intestinal Juices
| Type of Secretion | Daily Volume (ml) | pH |
| Saliva | 1000 | 6.0-7.0 |
| Gastric secretion | 1500 | 1.0-3.5 |
| Pancreatic secretion | 1000 | 8.0-8.3 |
| Bile | 1000 | 7.8 |
| Small intestine secretion | 1800 | 7.5-8.0 |
| Brunner's gland secretion | 200 | 8.0-8.9 |
| Large intestinal secretion | 200 | 7.5-8.0 |
| Total | 6700 |
Saliva contains two major types of protein secretion: (1) a serous secretion that contains ptyalin (an α-amylase), which is an enzyme for digesting starches, and (2) mucus secretion that contains mucin for lubricating and for surface protective purposes.
The parotid glands secrete almost entirely the serous type of secretion, whereas the submandibular and sublingual glands secrete both serous secretion and mucus. The buccal glands secrete only mucus. Saliva has a pH between 6.0 and 7.0, which is a favorable range for the digestive action of ptyalin.
Secretion of Ions in Saliva.
Saliva contains especially large quantities of potassium and bicarbonate ions. Conversely, the concentrations of both sodium and chloride ions are several times less in saliva than in plasma. One can understand these special concentrations of ions in the saliva from the following description of the mechanism for secretion of saliva.
Figure 65-2 shows secretion by the submandibular gland, a typical compound gland that contains acini and salivary ducts. Salivary secretion is a two-stage operation: The first stage involves the acini, and the second stage involves the salivary ducts. The acini secrete a primary secretion that contains ptyalin and/or mucin in a solution of ions with concentrations not greatly different from those of typical extracellular fluid. As the primary secretion flows through the ducts, two major active transport processes take place that markedly modify the ionic composition of the fluid in the saliva.
First, sodium ions are actively reabsorbed from all the salivary ducts and potassium ions are actively secreted in exchange for the sodium. Therefore, the sodium ion concentration of the saliva becomes greatly reduced, whereas the potassium ion concentration becomes increased. However, there is excess sodium reabsorption compared with potassium secretion, which creates electrical negativity of about −70 millivolts in the salivary ducts; this negativity in turn causes chloride ions to be reabsorbed passively. Therefore, the chloride ion concentration in the salivary fluid falls to a very low level, matching the ductal decrease in sodium ion concentration.
Second, bicarbonate ions are secreted by the ductal epithelium into the lumen of the duct. This secretion is at least partly caused by the passive exchange of bicarbonate for chloride ions, but it may also result partly from an active secretory process.
The net result of these transport processes is that under resting conditions, the concentrations of sodium and chloride ions in the saliva are only about 15 mEq/L each, about one seventh to one tenth their concentrations in plasma. Conversely, the concentration of potassium ions is about 30 mEq/L, seven times as great as in plasma, and the concentration of bicarbonate ions is 50 to 70 mEq/L, about two to three times that of plasma.
During maximal salivation, the salivary ionic concentrations change considerably because the rate of formation of primary secretion by the acini can increase as much as 20-fold. This acinar secretion then flows through the ducts so rapidly that the ductal reconditioning of the secretion is considerably reduced. Therefore, when copious quantities of saliva are being secreted, the sodium chloride concentration is about one half or two thirds that of plasma, and the potassium concentration rises to only four times that of plasma.
Function of Saliva for Oral Hygiene.
Under basal awake conditions, about 0.5 milliliter of saliva, almost entirely of the mucous type, is secreted each minute; however, during sleep, little secretion occurs. This secretion plays an exceedingly important role for maintaining healthy oral tissues. The mouth is loaded with pathogenic bacteria that can easily destroy tissues and cause dental caries. Saliva helps prevent the deteriorative processes in several ways:
1. The flow of saliva helps wash away pathogenic bacteria, as well as food particles that provide their metabolic support.
2. Saliva contains several factors that destroy bacteria. One of these is thiocyanate ions and another is several proteolytic enzymes—most important, lysozyme—that (a) attack the bacteria, (b) aid the thiocyanate ions in entering the bacteria where these ions in turn become bactericidal, and (c) digest food particles, thus helping further to remove the bacterial metabolic support.
3. Saliva often contains significant amounts of antibodies that can destroy oral bacteria, including some that cause dental caries. In the absence of salivation, oral tissues often become ulcerated and otherwise infected, and caries of the teeth can become rampant.
Nervous Regulation of Salivary Secretion
Figure 65-3 shows the parasympathetic nervous pathways for regulating salivation and demonstrates that the salivary glands are controlled mainly by parasympathetic nervous signals all the way from the superior and inferior salivatory nuclei in the brain stem.

The salivatory nuclei are located approximately at the juncture of the medulla and pons and are excited by both taste and tactile stimuli from the tongue and other areas of the mouth and pharynx. Many taste stimuli, especially the sour taste (caused by acids), elicit copious secretion of saliva—often 8 to 20 times the basal rate of secretion. Also, certain tactile stimuli, such as the presence of smooth objects in the mouth (e.g., a pebble), cause marked salivation, whereas rough objects cause less salivation and occasionally even inhibit salivation.
Salivation can also be stimulated or inhibited by nervous signals arriving in the salivatory nuclei from higher centers of the central nervous system. For instance, when a person smells or eats favorite foods, salivation is greater than when food that is disliked is smelled or eaten. The appetite area of the brain, which partially regulates these effects, is located in proximity to the parasympathetic centers of the anterior hypothalamus, and it functions to a great extent in response to signals from the taste and smell areas of the cerebral cortex or amygdala.
Salivation also occurs in response to reflexes originating in the stomach and upper small intestines—particularly when irritating foods are swallowed or when a person is nauseated because of some gastrointestinal abnormality. The saliva, when swallowed, helps to remove the irritating factor in the gastrointestinal tract by diluting or neutralizing the irritant substances.
Sympathetic stimulation can also increase salivation a slight amount—much less so than parasympathetic stimulation. The sympathetic nerves originate from the superior cervical ganglia and travel along the surfaces of the blood vessel walls to the salivary glands.
A secondary factor that also affects salivary secretion is the blood supply to the glands because secretion always requires adequate nutrients from the blood. The parasympathetic nerve signals that induce copious salivation also moderately dilate the blood vessels. In addition, salivation directly dilates the blood vessels, thus providing increased salivatory gland nutrition as needed by the secreting cells. Part of this additional vasodilator effect is caused by kallikrein secreted by the activated salivary cells, which in turn acts as an enzyme to split one of the blood proteins, an alpha2-globulin, to form bradykinin, a strong vasodilator.
Esophageal Secretion
Esophageal secretions are entirely mucous and mainly provide lubrication for swallowing. The main body of the esophagus is lined with many simple mucous glands. At the gastric end and to a lesser extent in the initial portion of the esophagus, many compound mucous glands can also be found. The mucus secreted by the compound glands in the upper esophagus prevents mucosal excoriation by newly entering food, whereas the compound glands located near the esophagogastric junction protect the esophageal wall from digestion by acidic gastric juices that often reflux from the stomach back into the lower esophagus. Despite this protection, a peptic ulcer at times can still occur at the gastric end of the esophagus.
Gastric Secretion
Characteristics of the Gastric Secretions
In addition to mucus-secreting cells that line the entire surface of the stomach, the stomach mucosa has two important types of tubular glands: oxyntic glands (also called gastric glands) and pyloric glands. The oxyntic (acid-forming) glands secrete hydrochloric acid, pepsinogen, intrinsic factor, and mucus. The pyloric glands secrete mainly mucus for protection of the pyloric mucosa from the stomach acid. They also secrete the hormone gastrin.
The oxyntic glands are located on the inside surfaces of the body and fundus of the stomach—the proximal 80 percent of the stomach. The pyloric glands are located in the antral portion of the stomach—the distal 20 percent of the stomach.
Secretions from the Oxyntic (Gastric) Glands
A typical stomach oxyntic gland is shown in Figure 65-4. It is composed of three types of cells: (1) mucous neck cells, which secrete mainly mucus; (2) peptic (or chief) cells, which secrete large quantities of pepsinogen; and (3) parietal (or oxyntic) cells, which secrete hydrochloric acid and intrinsic factor. Secretion of hydrochloric acid by the parietal cells involves special mechanisms, as follows.

Basic Mechanism of Hydrochloric Acid Secretion.
When stimulated, the parietal cells secrete an acid solution that contains about 160 mmol/L of hydrochloric acid, which is nearly isotonic with the body fluids. The pH of this acid is about 0.8, demonstrating its extreme acidity. At this pH, the hydrogen ion concentration is about 3 million times that of the arterial blood. To concentrate the hydrogen ions this tremendous amount requires more than 1500 calories of energy per liter of gastric juice. At the same time that hydrogen ions are secreted, bicarbonate ions diffuse into the blood so that gastric venous blood has a higher pH than arterial blood when the stomach is secreting acid.
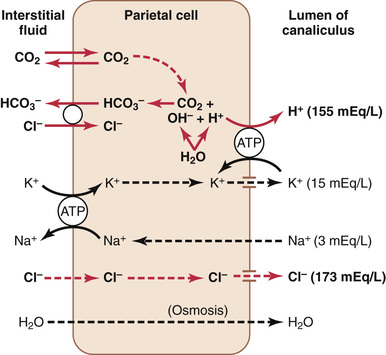
Figure 65-5 shows schematically the functional structure of a parietal cell (also called an oxyntic cell), demonstrating that it contains large branching intracellular canaliculi. The hydrochloric acid is formed at the villus-like projections inside these canaliculi and is then conducted through the canaliculi to the secretory end of the cell.

The main driving force for hydrochloric acid secretion by the parietal cells is a hydrogen-potassium pump (H+-K+ adenosine triphosphatase [ATPase]). The chemical mechanism of hydrochloric acid formation is shown in Figure 65-6 and consists of the following steps:
1. Water inside the parietal cell becomes dissociated into H+ and hydroxide (OH−) in the cell cytoplasm. The H+ is then actively secreted into the canaliculus in exchange for K+, an active exchange process that is catalyzed by H+-K+ ATPase. Potassium ions transported into the cell by the sodium Na+-K+ ATPase pump on the basolateral (extracellular) side of the membrane tend to leak into the lumen but are recycled back into the cell by the H+-K+ ATPase. The basolateral Na+-K+ ATPase creates low intracellular Na+, which contributes to Na+ reabsorption from the lumen of the canaliculus. Thus, most of the K+ and Na+ in the canaliculus is reabsorbed into the cell cytoplasm, and hydrogen ions take their place in the canaliculus.
2. The pumping of H+ out of the cell by the H+-K+ ATPase permits OH− to accumulate and form bicarbonate (HCO3−) from CO2, either formed during metabolism in the cell or while entering the cell from the blood. This reaction is catalyzed by carbonic anhydrase. The HCO3− is then transported across the basolateral membrane into the extracellular fluid in exchange for chloride ions, which enter the cell and are secreted through chloride channels into the canaliculus, giving a strong solution of hydrochloric acid in the canaliculus. The hydrochloric acid is then secreted outward through the open end of the canaliculus into the lumen of the gland.
3. Water passes into the canaliculus by osmosis because of extra ions secreted into the canaliculus. Thus, the final secretion from the canaliculus contains water, hydrochloric acid at a concentration of about 150 to 160 mEq/L, potassium chloride at a concentration of 15 mEq/L, and a small amount of sodium chloride.
To produce a concentration of hydrogen ions as great as that found in gastric juice requires minimal backleak into the mucosa of the secreted acid. A major part of the stomach's ability to prevent backleak of acid can be attributed to the gastric barrier due to the formation of alkaline mucus and to tight junctions between epithelia cells, as described later. If this barrier is damaged by toxic substances, such as occurs with excessive use of aspirin or alcohol, the secreted acid does leak down an electrochemical gradient into the mucosa, causing stomach mucosal damage.
The Basic Factors That Stimulate Gastric Secretion Are Acetylcholine, Gastrin, and Histamine.
Acetylcholine released by parasympathetic stimulation excites secretion of pepsinogen by peptic cells, hydrochloric acid by parietal cells, and mucus by mucous cells. In comparison, both gastrin and histamine strongly stimulate secretion of acid by parietal cells but have little effect on the other cells.
Secretion and Activation of Pepsinogen.
Several slightly different types of pepsinogen are secreted by the peptic and mucous cells of the gastric glands, but all the pepsinogens perform the same basic functions.
When pepsinogen is first secreted, it has no digestive activity. However, as soon as it comes in contact with hydrochloric acid, it is activated to form active pepsin. In this process, the pepsinogen molecule, having a molecular weight of about 42,500, is split to form a pepsin molecule, having a molecular weight of about 35,000.
Pepsin functions as an active proteolytic enzyme in a highly acidic medium (optimum pH 1.8 to 3.5), but above a pH of about 5 it has almost no proteolytic activity and becomes completely inactivated in a short time. Hydrochloric acid is as necessary as pepsin for protein digestion in the stomach, as discussed in Chapter 66.
Secretion of Intrinsic Factor by Parietal Cells.
The substance intrinsic factor, which is essential for absorption of vitamin B12 in the ileum, is secreted by the parietal cells along with the secretion of hydrochloric acid. When the acid-producing parietal cells of the stomach are destroyed, which frequently occurs in persons with chronic gastritis, not only does achlorhydria (lack of stomach acid secretion) develop, but pernicious anemia also often develops because of failure of maturation of the red blood cells in the absence of vitamin B12 stimulation of the bone marrow. This condition is discussed in detail in Chapter 33.
Pyloric Glands—Secretion of Mucus and Gastrin
The pyloric glands are structurally similar to the oxyntic glands but contain few peptic cells and almost no parietal cells. Instead, they contain mostly mucous cells that are identical with the mucous neck cells of the oxyntic glands. These cells secrete a small amount of pepsinogen, as discussed earlier, and an especially large amount of thin mucus that helps to lubricate food movement, as well as to protect the stomach wall from digestion by the gastric enzymes. The pyloric glands also secrete the hormone gastrin, which plays a key role in controlling gastric secretion, as we discuss shortly.
Surface Mucous Cells
The entire surface of the stomach mucosa between glands has a continuous layer of a special type of mucous cells called simply “surface mucous cells.” They secrete large quantities of viscid mucus that coats the stomach mucosa with a gel layer of mucus often more than 1 millimeter thick, thus providing a major shell of protection for the stomach wall, as well as contributing to lubrication of food transport.
Another characteristic of this mucus is that it is alkaline. Therefore, the normal underlying stomach wall is not directly exposed to the highly acidic, proteolytic stomach secretion. Even the slightest contact with food or any irritation of the mucosa directly stimulates the surface mucous cells to secrete additional quantities of this thick, alkaline, viscid mucus.
Stimulation of Gastric Acid Secretion
Parietal Cells of the Oxyntic Glands Are the Only Cells That Secrete Hydrochloric Acid.
As noted earlier in the chapter, the acidity of the fluid secreted by the parietal cells of the oxyntic glands can be great, with pH as low as 0.8. However, secretion of this acid is under continuous control by both endocrine and nervous signals. Furthermore, the parietal cells operate in close association with another type of cell called enterochromaffin-like cells (ECL cells), the primary function of which is to secrete histamine.
The ECL cells lie in the deep recesses of the oxyntic glands and therefore release histamine in direct contact with the parietal cells of the glands. The rate of formation and secretion of hydrochloric acid by the parietal cells is directly related to the amount of histamine secreted by the ECL cells. In turn, the ECL cells are stimulated to secrete histamine by the hormone gastrin, which is formed almost entirely in the antral portion of the stomach mucosa in response to proteins in the foods being digested. The ECL cells may also be stimulated by hormones secreted by the enteric nervous system of the stomach wall. We will first discuss the gastrin mechanism for control of the ECL cells and their subsequent control of parietal cell secretion of hydrochloric acid.
Stimulation of Acid Secretion by Gastrin.
Gastrin is a hormone secreted by gastrin cells, also called G cells. These cells are located in the pyloric glands in the distal end of the stomach. Gastrin is a large polypeptide secreted in two forms: a large form called G-34, which contains 34 amino acids, and a smaller form, G-17, which contains 17 amino acids. Although both of these forms are important, the smaller form is more abundant.
When meats or other foods containing protein reach the antral end of the stomach, some of the proteins from these foods have a special stimulatory effect on the gastrin cells in the pyloric glands to cause release of gastrin into the blood to be transported to the ECL cells of the stomach. The vigorous mixing of the gastric juices transports the gastrin rapidly to the ECL cells in the body of the stomach, causing release of histamine directly into the deep oxyntic glands. The histamine then acts quickly to stimulate gastric hydrochloric acid secretion.
Regulation of Pepsinogen Secretion
Stimulation of pepsinogen secretion by the peptic cells in the oxyntic glands occurs in response to two main types of signals: (1) acetylcholine released from the vagus nerves or from the gastric enteric nervous plexus, and (2) acid in the stomach. The acid probably does not stimulate the peptic cells directly but instead elicits additional enteric nervous reflexes that support the original nervous signals to the peptic cells. Therefore, the rate of secretion of pepsinogen, the precursor of the enzyme pepsin that causes protein digestion, is strongly influenced by the amount of acid in the stomach. In people who have lost the ability to secrete normal amounts of acid, secretion of pepsinogen is also decreased, even though the peptic cells may otherwise appear to be normal.
Phases of Gastric Secretion
Gastric secretion is said to occur in three “phases” (as shown in Figure 65-7): a cephalic phase, a gastric phase, and an intestinal phase.

 Cephalic Phase.
Cephalic Phase.
The cephalic phase of gastric secretion occurs even before food enters the stomach, especially while it is being eaten. It results from the sight, smell, thought, or taste of food, and the greater the appetite, the more intense is the stimulation. Neurogenic signals that cause the cephalic phase of gastric secretion originate in the cerebral cortex and in the appetite centers of the amygdala and hypothalamus. They are transmitted through the dorsal motor nuclei of the vagi and thence through the vagus nerves to the stomach. This phase of secretion normally accounts for about 30 percent of the gastric secretion associated with eating a meal.
Gastric Phase.
Once food enters the stomach, it excites (1) long vagovagal reflexes from the stomach to the brain and back to the stomach, (2) local enteric reflexes, and (3) the gastrin mechanism, all of which cause secretion of gastric juice during several hours while food remains in the stomach. The gastric phase of secretion accounts for about 60 percent of the total gastric secretion associated with eating a meal and therefore accounts for most of the total daily gastric secretion of about 1500 milliliters.
Intestinal Phase.
The presence of food in the upper portion of the small intestine, particularly in the duodenum, will continue to cause stomach secretion of small amounts of gastric juice, probably partly because of small amounts of gastrin released by the duodenal mucosa. This secretion accounts for about 10 percent of the acid response to a meal.
Inhibition of Gastric Secretion by Other Intestinal Factors
Although intestinal chyme slightly stimulates gastric secretion during the early intestinal phase of stomach secretion, it paradoxically inhibits gastric secretion at other times. This inhibition results from at least two influences.
1. The presence of food in the small intestine initiates a reverse enterogastric reflex, transmitted through the myenteric nervous system and extrinsic sympathetic and vagus nerves, that inhibits stomach secretion. This reflex can be initiated by (a) distending the small bowel, (b) the presence of acid in the upper intestine, (c) the presence of protein breakdown products, or (d) irritation of the mucosa. This reflex is part of the complex mechanism discussed in Chapter 64 for slowing stomach emptying when the intestines are already filled.
2. The presence of acid, fat, protein breakdown products, hyperosmotic or hypo-osmotic fluids, or any irritating factor in the upper small intestine causes release of several intestinal hormones. One of these hormones is secretin, which is especially important for control of pancreatic secretion. However, secretin opposes stomach secretion. Three other hormones—glucose-dependent insulinotropic peptide (gastric inhibitory peptide), vasoactive intestinal polypeptide, and somatostatin—also have slight to moderate effects in inhibiting gastric secretion.
The purpose of intestinal factors that inhibit gastric secretion is presumably to slow passage of chyme from the stomach when the small intestine is already filled or already overactive. In fact, the enterogastric inhibitory reflexes plus inhibitory hormones usually also reduce stomach motility at the same time that they reduce gastric secretion, as was discussed in Chapter 64.
Gastric Secretion During the Interdigestive Period.
The stomach secretes a few milliliters of gastric juice each hour during the “interdigestive period,” when little or no digestion is occurring anywhere in the gut. The secretion that does occur is usually almost entirely of the nonoxyntic type, composed mainly of mucus but little pepsin and almost no acid.
Emotional stimuli may increase interdigestive gastric secretion (which is highly peptic and acidic) to 50 milliliters or more per hour, in much the same way that the cephalic phase of gastric secretion excites secretion at the onset of a meal. This increase of secretion in response to emotional stimuli is believed to contribute to the development of peptic ulcers, as discussed in Chapter 67.
Chemical Composition of Gastrin and Other Gastrointestinal Hormones
Gastrin, cholecystokinin (CCK), and secretin are all large polypeptides with approximate molecular weights of 2000, 4200, and 3400, respectively. The terminal five amino acids in the gastrin and CCK molecular chains are the same. The functional activity of gastrin resides in the terminal four amino acids, and the activity for CCK resides in the terminal eight amino acids. All the amino acids in the secretin molecule are essential.
A synthetic gastrin, composed of the terminal four amino acids of natural gastrin plus the amino acid alanine, has all the same physiological properties as the natural gastrin. This synthetic product is called pentagastrin.
Pancreatic Secretion
The pancreas, which lies parallel to and beneath the stomach (illustrated in Figure 65-10), is a large compound gland, and most of its internal structure is similar to that of the salivary glands shown in Figure 65-2. The pancreatic digestive enzymes are secreted by pancreatic acini, and large volumes of sodium bicarbonate solution are secreted by the small ductules and larger ducts leading from the acini. The combined product of enzymes and sodium bicarbonate then flows through a long pancreatic duct that normally joins the hepatic duct immediately before it empties into the duodenum through the papilla of Vater, surrounded by the sphincter of Oddi.
Pancreatic juice is secreted most abundantly in response to the presence of chyme in the upper portions of the small intestine, and the characteristics of the pancreatic juice are determined to some extent by the types of food in the chyme. (The pancreas also secretes insulin, but it is not secreted by the same pancreatic tissue that secretes intestinal pancreatic juice. Instead, insulin is secreted directly into the blood—not into the intestine—by the islets of Langerhans that occur in islet patches throughout the pancreas. These structures are discussed in Chapter 79.)
Pancreatic Digestive Enzymes
Pancreatic secretion contains multiple enzymes for digesting all of the three major types of food: proteins, carbohydrates, and fats. It also contains large quantities of bicarbonate ions, which play an important role in neutralizing the acidity of the chyme emptied from the stomach into the duodenum.
The most important of the pancreatic enzymes for digesting proteins are trypsin, chymotrypsin, and carboxypolypeptidase. By far the most abundant of these is trypsin.
Trypsin and chymotrypsin split whole and partially digested proteins into peptides of various sizes but do not cause release of individual amino acids. However, carboxypolypeptidase splits some peptides into individual amino acids, thus completing digestion of some proteins all the way to the amino acid state.
The pancreatic enzyme for digesting carbohydrates is pancreatic amylase, which hydrolyzes starches, glycogen, and most other carbohydrates (except cellulose) to form mostly disaccharides and a few trisaccharides.
The main enzymes for fat digestion are (1) pancreatic lipase, which is capable of hydrolyzing neutral fat into fatty acids and monoglycerides; (2) cholesterol esterase, which causes hydrolysis of cholesterol esters; and (3) phospholipase, which splits fatty acids from phospholipids.
When first synthesized in the pancreatic cells, the proteolytic digestive enzymes are in their enzymatically inactive forms trypsinogen, chymotrypsinogen, and procarboxypolypeptidase. They become activated only after they are secreted into the intestinal tract. Trypsinogen is activated by an enzyme called enterokinase, which is secreted by the intestinal mucosa when chyme comes in contact with the mucosa. Trypsinogen also can be autocatalytically activated by trypsin that has already been formed from previously secreted trypsinogen. Chymotrypsinogen is activated by trypsin to form chymotrypsin, and procarboxypolypeptidase is activated in a similar manner.
Secretion of Trypsin Inhibitor Prevents Digestion of the Pancreas.
It is important that the proteolytic enzymes of the pancreatic juice not become activated until after they have been secreted into the intestine because the trypsin and the other enzymes would digest the pancreas. Fortunately, the same cells that secrete proteolytic enzymes into the acini of the pancreas simultaneously secrete another substance called trypsin inhibitor. This substance, which is formed in the cytoplasm of the glandular cells, prevents activation of trypsin both inside the secretory cells and in the acini and ducts of the pancreas. In addition, because it is trypsin that activates the other pancreatic proteolytic enzymes, trypsin inhibitor prevents activation of the other enzymes as well.
When the pancreas becomes severely damaged or when a duct becomes blocked, large quantities of pancreatic secretion sometimes become pooled in the damaged areas of the pancreas. Under these conditions, the effect of trypsin inhibitor is often overwhelmed, in which case the pancreatic secretions rapidly become activated and can literally digest the entire pancreas within a few hours, giving rise to the condition called acute pancreatitis. This condition is sometimes lethal because of accompanying circulatory shock; even if it is not lethal, it usually leads to a subsequent lifetime of pancreatic insufficiency.
Secretion of Bicarbonate Ions
Although the enzymes of the pancreatic juice are secreted entirely by the acini of the pancreatic glands, the other two important components of pancreatic juice, bicarbonate ions and water, are secreted mainly by the epithelial cells of the ductules and ducts that lead from the acini. When the pancreas is stimulated to secrete copious quantities of pancreatic juice, the bicarbonate ion concentration can rise to as high as 145 mEq/L, a value about five times that of bicarbonate ions in the plasma. This high concentration provides a large quantity of alkali in the pancreatic juice that serves to neutralize the hydrochloric acid emptied into the duodenum from the stomach.
The basic steps in the cellular mechanism for secreting sodium bicarbonate solution into the pancreatic ductules and ducts, shown in Figure 65-8, are as follows:
1. Carbon dioxide diffuses to the interior of the cell from the blood and, under the influence of carbonic anhydrase, combines with water to form carbonic acid (H2CO3). The carbonic acid dissociates into bicarbonate ions and hydrogen ions (HCO3− and H+). Additional bicarbonate ions enter the cell through the basolateral membrane by co-transport with sodium ions (Na+). The bicarbonate ions are then exchanged for chloride ions (Cl−) by secondary active transport through the luminal border of the cell into the lumen of the duct. The chloride that enters the cell is recycled back into the lumen by special chloride channels.
2. The hydrogen ions formed by dissociation of carbonic acid inside the cell are exchanged for sodium ions through the basolateral membrane of the cell by secondary active transport. Sodium ions also enter the cell by co-transport with bicarbonate across the basolateral membrane. Sodium ions are then transported across the luminal border into the pancreatic duct lumen. The negative voltage of the lumen also pulls the positively charged sodium ions across the tight junctions between the cells.
3. The overall movement of sodium and bicarbonate ions from the blood into the duct lumen creates an osmotic pressure gradient that causes osmosis of water also into the pancreatic duct, thus forming an almost completely isosmotic bicarbonate solution.

Regulation of Pancreatic Secretion
Basic Stimuli That Cause Pancreatic Secretion
Three basic stimuli are important in causing pancreatic secretion:
1. Acetylcholine, which is released from the parasympathetic vagus nerve endings and from other cholinergic nerves in the enteric nervous system
2. Cholecystokinin, which is secreted by the duodenal and upper jejunal mucosa when food enters the small intestine
3. Secretin, which is also secreted by the duodenal and jejunal mucosa when highly acidic food enters the small intestine
The first two of these stimuli, acetylcholine and cholecystokinin, stimulate the acinar cells of the pancreas, causing production of large quantities of pancreatic digestive enzymes but relatively small quantities of water and electrolytes to go with the enzymes. Without the water, most of the enzymes remain temporarily stored in the acini and ducts until more fluid secretion comes along to wash them into the duodenum. Secretin, in contrast to the first two basic stimuli, stimulates secretion of large quantities of water solution of sodium bicarbonate by the pancreatic ductal epithelium.
Multiplicative Effects of Different Stimuli.
When all the different stimuli of pancreatic secretion occur at once, the total secretion is far greater than the sum of the secretions caused by each one separately. Therefore, the various stimuli are said to “multiply,” or “potentiate,” one another. Thus, pancreatic secretion normally results from the combined effects of the multiple basic stimuli, not from one alone.
Phases of Pancreatic Secretion
Pancreatic secretion, as with gastric secretion, occurs in three phases: the cephalic phase, the gastric phase, and the intestinal phase. Their characteristics are described in the following sections.
Cephalic and Gastric Phases.
During the cephalic phase of pancreatic secretion, the same nervous signals from the brain that cause secretion in the stomach also cause acetylcholine release by the vagal nerve endings in the pancreas. This signaling causes moderate amounts of enzymes to be secreted into the pancreatic acini, accounting for about 20 percent of the total secretion of pancreatic enzymes after a meal. However, little of the secretion flows immediately through the pancreatic ducts into the intestine because only small amounts of water and electrolytes are secreted along with the enzymes.
During the gastric phase, the nervous stimulation of enzyme secretion continues, accounting for another 5 to 10 percent of pancreatic enzymes secreted after a meal. However, again, only small amounts reach the duodenum because of continued lack of significant fluid secretion.
Intestinal Phase.
After chyme leaves the stomach and enters the small intestine, pancreatic secretion becomes copious, mainly in response to the hormone secretin.
Secretin Stimulates Copious Secretion of Bicarbonate Ions, Which Neutralizes Acidic Stomach Chyme.
Secretin is a polypeptide containing 27 amino acids (with a molecular weight of about 3400). It is present in an inactive form, prosecretin, in so-called S cells in the mucosa of the duodenum and jejunum. When acid chyme with a pH less than 4.5 to 5.0 enters the duodenum from the stomach, it causes duodenal mucosal release and activation of secretin, which is then absorbed into the blood. The one truly potent constituent of chyme that causes this secretin release is the hydrochloric acid from the stomach.
Secretin in turn causes the pancreas to secrete large quantities of fluid containing a high concentration of bicarbonate ion (up to 145 mEq/L) but a low concentration of chloride ion. The secretin mechanism is especially important for two reasons: First, secretin begins to be released from the mucosa of the small intestine when the pH of the duodenal contents falls below 4.5 to 5.0, and its release increases greatly as the pH falls to 3.0. This mechanism immediately causes copious secretion of pancreatic juice that contains abundant amounts of sodium bicarbonate. The net result is then the following reaction in the duodenum:
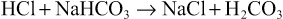
The carbonic acid then immediately dissociates into carbon dioxide and water. The carbon dioxide is absorbed into the blood and expired through the lungs, thus leaving a neutral solution of sodium chloride in the duodenum. In this way, the acid contents that are emptied into the duodenum from the stomach become neutralized, and thus further peptic digestive activity by the gastric juices in the duodenum is immediately blocked. Because the mucosa of the small intestine cannot withstand the digestive action of acid gastric juice, this protective mechanism is essential to prevent the development of duodenal ulcers, as is discussed in further detail in Chapter 67.
Bicarbonate ion secretion by the pancreas provides an appropriate pH for action of the pancreatic digestive enzymes, which function optimally in a slightly alkaline or neutral medium, at a pH of 7.0 to 8.0. Fortunately, the pH of the sodium bicarbonate secretion averages 8.0.
Cholecystokinin Contributes to Control of Digestive Enzyme Secretion by the Pancreas.
The presence of food in the upper small intestine also causes a second hormone, cholecystokinin (CCK), a polypeptide containing 33 amino acids, to be released from yet another group of cells, the I cells, in the mucosa of the duodenum and upper jejunum. This release of CCK results especially from the presence of proteoses and peptones (products of partial protein digestion) and long-chain fatty acids in the chyme coming from the stomach.
CCK, like secretin, passes by way of the blood to the pancreas, but instead of causing sodium bicarbonate secretion, it mainly causes secretion of much more pancreatic digestive enzymes by the acinar cells. This effect is similar to that caused by vagal stimulation but is even more pronounced, accounting for 70 to 80 percent of the total secretion of the pancreatic digestive enzymes after a meal.
The differences between the pancreatic stimulatory effects of secretin and CCK are shown in Figure 65-9, which demonstrates (1) intense sodium bicarbonate secretion in response to acid in the duodenum, stimulated by secretin; (2) a dual effect in response to soap (a fat); and (3) intense digestive enzyme secretion (when peptones enter the duodenum) stimulated by CCK.
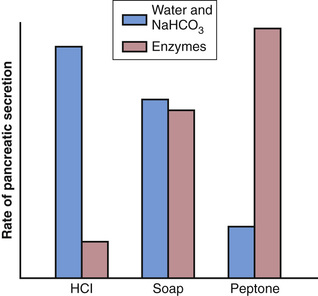
Figure 65-10 summarizes the more important factors in the regulation of pancreatic secretion. The total amount secreted each day is about 1 liter.

Bile Secretion by the Liver
One of the many functions of the liver is to secrete bile, normally between 600 and 1000 ml/day. Bile serves two important functions.
First, bile plays an important role in fat digestion and absorption, not because of any enzymes in the bile that cause fat digestion, but because bile acids in the bile perform two functions: (1) They help emulsify the large fat particles of the food into many minute particles, the surface of which can then be attacked by lipase enzymes secreted in pancreatic juice, and (2) they aid in absorption of the digested fat end products through the intestinal mucosal membrane.
Second, bile serves as a means for excretion of several important waste products from the blood. These waste products include in particular bilirubin, an end product of hemoglobin destruction, and excesses of cholesterol.
Physiologic Anatomy of Biliary Secretion
Bile is secreted in two stages by the liver:
1. The initial portion is secreted by the principal functional cells of the liver, the hepatocytes; this initial secretion contains large amounts of bile acids, cholesterol, and other organic constituents. It is secreted into minute bile canaliculi that originate between the hepatic cells.
2. Next, the bile flows in the canaliculi toward the interlobular septa, where the canaliculi empty into terminal bile ducts and then into progressively larger ducts, finally reaching the hepatic duct and common bile duct. From these ducts the bile either empties directly into the duodenum or is diverted for minutes up to several hours through the cystic duct into the gallbladder, shown in Figure 65-11.

In its course through the bile ducts, a second portion of liver secretion is added to the initial bile. This additional secretion is a watery solution of sodium and bicarbonate ions secreted by epithelial cells that line the ductules and ducts. This second secretion sometimes increases the total quantity of bile by as much as 100 percent. The second secretion is stimulated especially by secretin, which causes release of additional quantities of bicarbonate ions to supplement the bicarbonate ions in pancreatic secretion (for neutralizing acid that empties into the duodenum from the stomach).
Storing and Concentrating Bile in the Gallbladder.
Bile is secreted continually by the liver cells, but most of it is normally stored in the gallbladder until it is needed in the duodenum. The maximum volume that the gallbladder can hold is only 30 to 60 milliliters. Nevertheless, as much as 12 hours of bile secretion (usually about 450 milliliters) can be stored in the gallbladder because water, sodium, chloride, and most other small electrolytes are continually absorbed through the gallbladder mucosa, concentrating the remaining bile constituents that contain the bile salts, cholesterol, lecithin, and bilirubin.
Most of this gallbladder absorption is caused by active transport of sodium through the gallbladder epithelium, and this transport is followed by secondary absorption of chloride ions, water, and most other diffusible constituents. Bile is normally concentrated in this way about 5-fold, but it can be concentrated up to a maximum of 20-fold.
Composition of Bile.
Table 65-2 lists the composition of bile when it is first secreted by the liver and then after it has been concentrated in the gallbladder. By far the most abundant substances secreted in the bile are bile salts, which account for about one half of the total solutes also in the bile. Also secreted or excreted in large concentrations are bilirubin, cholesterol, lecithin, and the usual electrolytes of plasma.
Table 65-2
Composition of Bile
| Substance | Liver Bile | Gallbladder Bile |
| Water | 97.5 g/dl | 92 g/dl |
| Bile salts | 1.1 g/dl | 6 g/dl |
| Bilirubin | 0.04 g/dl | 0.3 g/dl |
| Cholesterol | 0.1 g/dl | 0.3 to 0.9 g/dl |
| Fatty acids | 0.12 g/dl | 0.3 to 1.2 g/dl |
| Lecithin | 0.04 g/dl | 0.3 g/dl |
| Na+ | 145 mEq/L | 130 mEq/L |
| K+ | 5 mEq/L | 12 mEq/L |
| Ca++ | 5 mEq/L | 23 mEq/L |
| Cl− | 100 mEq/L | 25 mEq/L |
| HCO3− | 28 mEq/L | 10 mEq/L |
In the concentrating process in the gallbladder, water and large portions of the electrolytes (except calcium ions) are reabsorbed by the gallbladder mucosa; essentially all other constituents, especially the bile salts and the lipid substances cholesterol and lecithin, are not reabsorbed and, therefore, become highly concentrated in the gallbladder bile.
Cholecystokinin Stimulates Gallbladder Emptying.
When food begins to be digested in the upper gastrointestinal tract, the gallbladder begins to empty, especially when fatty foods reach the duodenum about 30 minutes after a meal. The mechanism of gallbladder emptying is rhythmical contractions of the gallbladder wall, but effective emptying also requires simultaneous relaxation of the sphincter of Oddi, which guards the exit of the common bile duct into the duodenum.
By far the most potent stimulus for causing the gallbladder contractions is the hormone CCK. This is the same CCK discussed earlier that causes increased secretion of digestive enzymes by the acinar cells of the pancreas. The stimulus for CCK entry into the blood from the duodenal mucosa is mainly the presence of fatty foods in the duodenum.
The gallbladder is also stimulated less strongly by acetylcholine-secreting nerve fibers from both the vagi and the intestinal enteric nervous system. They are the same nerves that promote motility and secretion in other parts of the upper gastrointestinal tract.
In summary, the gallbladder empties its store of concentrated bile into the duodenum mainly in response to the CCK stimulus that is initiated mainly by fatty foods. When fat is not in the food, the gallbladder empties poorly, but when significant quantities of fat are present, the gallbladder normally empties completely in about 1 hour. Figure 65-11 summarizes the secretion of bile, its storage in the gallbladder, and its ultimate release from the bladder to the duodenum.
Function of Bile Salts in Fat Digestion and Absorption
The liver cells synthesize about 6 grams of bile salts daily. The precursor of the bile salts is cholesterol, which is either present in the diet or synthesized in the liver cells during the course of fat metabolism. The cholesterol is first converted to cholic acid or chenodeoxycholic acid in about equal quantities. These acids in turn combine principally with glycine and to a lesser extent with taurine to form glyco- and tauro-conjugated bile acids. The salts of these acids, mainly sodium salts, are then secreted in the bile.
The bile salts have two important actions in the intestinal tract:
First, they have a detergent action on the fat particles in the food. This action, which decreases the surface tension of the particles and allows agitation in the intestinal tract to break the fat globules into minute sizes, is called the emulsifying or detergent function of bile salts.
Second, and even more important than the emulsifying function, bile salts help in the absorption of (1) fatty acids, (2) monoglycerides, (3) cholesterol, and (4) other lipids from the intestinal tract. They help in this absorption by forming small physical complexes with these lipids; the complexes are called micelles, and they are semisoluble in the chyme because of the electrical charges of the bile salts. The intestinal lipids are “ferried” in this form to the intestinal mucosa, where they are then absorbed into the blood, as will be described in detail in Chapter 66. Without the presence of bile salts in the intestinal tract, up to 40 percent of the ingested fats are lost into the feces and a metabolic deficit often develops because of this nutrient loss.
Enterohepatic Circulation of Bile Salts.
About 94 percent of the bile salts are reabsorbed into the blood from the small intestine, about one half of this by diffusion through the mucosa in the early portions of the small intestine and the remainder by an active transport process through the intestinal mucosa in the distal ileum. They then enter the portal blood and pass back to the liver. Upon reaching the liver and during first passage through the venous sinusoids, these salts are absorbed almost entirely back into the hepatic cells and are then resecreted into the bile.
In this way, about 94 percent of all the bile salts are recirculated into the bile, so on average these salts make the entire circuit some 17 times before being carried out in the feces. The small quantities of bile salts lost into the feces are replaced by new amounts formed continually by the liver cells. This recirculation of the bile salts is called the enterohepatic circulation of bile salts.
The quantity of bile secreted by the liver each day is highly dependent on the availability of bile salts—the greater the quantity of bile salts in the enterohepatic circulation (usually a total of only about 2.5 grams), the greater the rate of bile secretion. Indeed, ingestion of supplemental bile salts can increase bile secretion by several hundred milliliters per day.
If a bile fistula empties the bile salts to the exterior for several days to several weeks so they cannot be reabsorbed from the ileum, the liver increases its production of bile salts 6- to 10-fold, which increases the rate of bile secretion most of the way back to normal. This demonstrates that the daily rate of liver bile salt secretion is actively controlled by the availability (or lack of availability) of bile salts in the enterohepatic circulation.
Role of Secretin in Controlling Bile Secretion.
In addition to the strong stimulating effect of bile acids to cause bile secretion, the hormone secretin—which also stimulates pancreatic secretion—increases bile secretion, sometimes more than doubling its secretion for several hours after a meal. This increase in secretion consists almost entirely of secretion of a sodium bicarbonate–rich watery solution by the epithelial cells of the bile ductules and ducts and does not represent increased secretion by the liver parenchymal cells themselves. The bicarbonate in turn passes into the small intestine and joins the bicarbonate from the pancreas in neutralizing the hydrochloric acid from the stomach. Thus, the secretin feedback mechanism for neutralizing duodenal acid operates not only through its effects on pancreatic secretion but also to a lesser extent through its effect on secretion by the liver ductules and ducts.
Liver Secretion of Cholesterol and Gallstone Formation
Bile salts are formed in the hepatic cells from cholesterol in the blood plasma. In the process of secreting the bile salts, about 1 to 2 grams of cholesterol are removed from the blood plasma and secreted into the bile each day.
Cholesterol is almost completely insoluble in pure water, but the bile salts and lecithin in bile combine physically with the cholesterol to form ultramicroscopic micelles in the form of a colloidal solution, as explained in more detail in Chapter 66. When the bile becomes concentrated in the gallbladder, the bile salts and lecithin become concentrated along with the cholesterol, which keeps the cholesterol in solution.
Under abnormal conditions, the cholesterol may precipitate in the gallbladder, resulting in the formation of cholesterol gallstones, as shown in Figure 65-12. The amount of cholesterol in the bile is determined partly by the quantity of fat that the person eats, because liver cells synthesize cholesterol as one of the products of fat metabolism in the body. For this reason, people who consume a high-fat diet over a period of years are prone to development of gallstones.

Inflammation of the gallbladder epithelium, often resulting from a low-grade chronic infection, may also change the absorptive characteristics of the gallbladder mucosa, sometimes allowing excessive absorption of water and bile salts but leaving behind the cholesterol in the gallbladder in progressively greater concentrations. The cholesterol then begins to precipitate, first forming many small crystals of cholesterol on the surface of the inflamed mucosa, but then progressing to large gallstones.
Secretions of the Small Intestine
Secretion of Mucus by Brunner's Glands in the Duodenum
An extensive array of compound mucous glands, called Brunner's glands, is located in the wall of the first few centimeters of the duodenum, mainly between the pylorus of the stomach and the papilla of Vater, where pancreatic secretion and bile empty into the duodenum. These glands secrete large amounts of alkaline mucus in response to (1) tactile or irritating stimuli on the duodenal mucosa; (2) vagal stimulation, which causes increased Brunner gland secretion concurrently with increase in stomach secretion; and (3) gastrointestinal hormones, especially secretin.
The function of the mucus secreted by Brunner's glands is to protect the duodenal wall from digestion by the highly acidic gastric juice emptying from the stomach. In addition, the mucus contains a large excess of bicarbonate ions, which add to the bicarbonate ions from pancreatic secretion and liver bile in neutralizing the hydrochloric acid entering the duodenum from the stomach.
Brunner's glands are inhibited by sympathetic stimulation; therefore, such stimulation in very excitable persons is likely to leave the duodenal bulb unprotected and is perhaps one of the factors that cause this area of the gastrointestinal tract to be the site of peptic ulcers in about 50 percent of persons with ulcers.
Secretion of Intestinal Digestive Juices by the Crypts of Lieberkühn
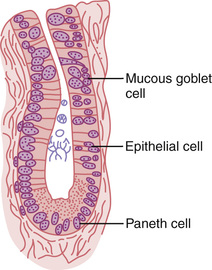
Located over the entire surface of the small intestine are small pits called crypts of Lieberkühn, one of which is illustrated in Figure 65-13. These crypts lie between the intestinal villi. The surfaces of both the crypts and the villi are covered by an epithelium composed of two types of cells: (1) a moderate number of goblet cells, which secrete mucus that lubricates and protects the intestinal surfaces, and (2) a large number of enterocytes, which, in the crypts, secrete large quantities of water and electrolytes and, over the surfaces of adjacent villi, reabsorb the water and electrolytes along with the end products of digestion.
The intestinal secretions are formed by the enterocytes of the crypts at a rate of about 1800 ml/day. These secretions are almost pure extracellular fluid and have a slightly alkaline pH in the range of 7.5 to 8.0. The secretions are also rapidly reabsorbed by the villi. This flow of fluid from the crypts into the villi supplies a watery vehicle for absorption of substances from chyme when it comes in contact with the villi. Thus, the primary function of the small intestine is to absorb nutrients and their digestive products into the blood.
Mechanism of Secretion of the Watery Fluid.
The exact mechanism that controls the marked secretion of watery fluid by the crypts of Lieberkühn is still unclear, but it is believed to involve at least two active secretory processes: (1) active secretion of chloride ions into the crypts and (2) active secretion of bicarbonate ions. The secretion of both ions causes electrical drag of positively charged sodium ions through the membrane and into the secreted fluid as well. Finally, all these ions together cause osmotic movement of water.
Digestive Enzymes in the Small Intestinal Secretion.
When secretions of the small intestine are collected without cellular debris, they have almost no enzymes. The enterocytes of the mucosa, especially those that cover the villi, contain digestive enzymes that digest specific food substances while they are being absorbed through the epithelium. These enzymes are the following: (1) several peptidases for splitting small peptides into amino acids; (2) four enzymes—sucrase, maltase, isomaltase, and lactase—for splitting disaccharides into monosaccharides; and (3) small amounts of intestinal lipase for splitting neutral fats into glycerol and fatty acids.
The epithelial cells deep in the crypts of Lieberkühn continually undergo mitosis, and new cells migrate along the basement membrane upward out of the crypts toward the tips of the villi, thus continually replacing the villus epithelium and also forming new digestive enzymes. As the villus cells age, they are finally shed into the intestinal secretions. The life cycle of an intestinal epithelial cell is about 5 days. This rapid growth of new cells also allows rapid repair of excoriations that occur in the mucosa.
Regulation of Small Intestine Secretion—Local Stimuli
By far the most important means for regulating small intestine secretion are local enteric nervous reflexes, especially reflexes initiated by tactile or irritative stimuli from the chyme in the intestines.
Secretion of Mucus by the Large Intestine
Mucus Secretion.
The mucosa of the large intestine, like that of the small intestine, has many crypts of Lieberkühn; however, unlike the small intestine, it contains no villi. The epithelial cells secrete almost no digestive enzymes. Instead, they contain mucous cells that secrete only mucus. This mucus contains moderate amounts of bicarbonate ions secreted by a few non–mucus-secreting epithelial cells. The rate of secretion of mucus is regulated principally by direct, tactile stimulation of the epithelial cells lining the large intestine and by local nervous reflexes to the mucous cells in the crypts of Lieberkühn.
Stimulation of the pelvic nerves from the spinal cord, which carry parasympathetic innervation to the distal one half to two thirds of the large intestine, also can cause a marked increase in mucus secretion. This increase occurs along with an increase in peristaltic motility of the colon, which was discussed in Chapter 64.
During extreme parasympathetic stimulation, often caused by emotional disturbances, so much mucus can occasionally be secreted into the large intestine that the person has a bowel movement of ropy mucus as often as every 30 minutes; this mucus often contains little or no fecal material.
Mucus in the large intestine protects the intestinal wall against excoriation, but in addition, it provides an adherent medium for holding fecal matter together. Furthermore, it protects the intestinal wall from the great amount of bacterial activity that takes place inside the feces, and, finally, the mucus plus the alkalinity of the secretion (a pH of 8.0 caused by large amounts of sodium bicarbonate) provides a barrier to keep acids formed in the feces from attacking the intestinal wall.
Diarrhea Caused by Excess Secretion of Water and Electrolytes in Response to Irritation.
Whenever a segment of the large intestine becomes intensely irritated, as occurs when bacterial infection becomes rampant during enteritis, the mucosa secretes extra large quantities of water and electrolytes in addition to the normal viscid alkaline mucus. This secretion acts to dilute the irritating factors and to cause rapid movement of the feces toward the anus. The result is diarrhea, with loss of large quantities of water and electrolytes. However, the diarrhea also washes away irritant factors, which promotes earlier recovery from the disease than might otherwise occur.