Chapter 12
Antigen
Introduction
Antigen is a molecule, which when introduced parenterally into the body, initiates the production of an antibody with which it reacts specifically in an observable manner.
Molecules that can be recognised by the immunoglobulin receptor of B cells or by the T-cell receptor when complexed with major histocompatibility complex (MHC) are called antigens. The word antigen is a shortened form of the words ‘antibody generator.’ Antigens are substances that react with antibodies, whereas immunogens are molecules that induce an immune response. In most cases, antigens are immunogens, and the terms are used interchangeably. The antigens that are not immunogenic but can take part in immune reactions are termed as haptens. The term immunogenicity means the ability of an antigen to elicit an immune reaction in the form of a B-cell or T-cell response, whereas the term antigenicity means just the ability to combine specifically with the products of the above responses. All molecules that are immunogenic are antigenic too, but all antigenic molecules cannot be considered immunogenic. Thus, haptens can be said to lack immunogenicity.
Attributes of Antigenicity
Immunogenicity (the ability of an antigen to elicit an immune reaction) and immunological reactivity (specific reaction with antibodies or sensitised cells) are two important attributes of the antigen. On the basis of these properties antigens can be classified as (1) antigen and (2) hapten.
An antigen can induce formation of antibody and also produce a specific and observable reaction with the antibody so produced.
Haptens
Haptens are small organic molecules that are antigenic but not immunogenic. They are not immunogenic because they cannot activate helper T cells. Failure of hapten to activate helper T cells is due to their inability to bind to MHC proteins; they cannot bind because they are not proteins and only proteins can be presented by MHC proteins. Moreover, haptens are univalent hence cannot activate B cells by themselves.
The haptens, however, can activate B cells when covalently bound to a ‘carrier’ protein. When bound with a carrier molecule, they form an immunogenic hapten-carrier conjugate (Fig. 12-1). In this process, the haptens combine with an IgM receptor on the B cells, and the hapten-carrier protein complex is internalised. A peptide of the carrier protein is presented in association with Class-II MHC protein to the helper T cells. The activated helper T cells then produce interleukins, which stimulate the B cells to produce antibodies to hapten.

Fig. 12-1 Hapten-carrier conjugate.
Animals immunised with such a conjugate produce antibodies specific for (1) the hapten determinant, (2) un-altered epitopes on the carrier protein, and (3) new epitopes formed by combined parts of both the hapten and carrier. In fact, the hapten-carrier molecule is bound to surface immunoglobulins on B cells via the hapten epitopes. The hapten-carrier molecule is then taken in, processed and pieces of the carrier are presented by these B cells and TH cells. In the body, the formation of hapten-carrier conjugates is the basis for development of allergic responses to drugs, such as penicillin.
Determinants of Antigenicity
A number of factors have been identified that make a substance immunogenic. Some of the important determinants of antigenicity include:
1. molecular size
2. foreignness
3. chemical-structural complexity
4. stability
5. other factors
Molecular Size
In general, protein molecules with large molecular weight are highly antigenic. Substances with molecular weights of about 100,000 Da and more are highly immunogenic, whereas substances with molecular weights of less than 5000 Da are generally not immunogenic. This property has been exploited in experimental studies by using high molecular weight proteins such as bovine gamma globulin (MW 150,000 Da) to induce an immune reaction. Substances with low molecular weight may be made antigenic by adsorbing these on carrier particles, such as bentonite, kaolin and other inert particles.
Foreignness
To be immunogenic, a molecule must be recognised as non-self, that is foreign. The molecule is considered self or non-self by the immune system depending on whether or not the molecule was exposed to the immune system during foetal development.
Foreignness implies ability of the host to tolerate self-antigens. Tolerance to self-antigens develops by contact with them in the initial phases of the development of immune system, particularly during the development of lymphocytes.
In general, the more distantly related two species are, the greater the immunogenicity of a molecule from one species will be when exposed to the other. For example, the bovine serum albumin is more immunogenic in a chicken than in a goat. A graft from an unrelated human will be rejected within about 2 weeks unless immunosuppressive drugs are used, but a graft from a chimpanzee will be rejected within hours even if drugs are used. In contrast, a kidney graft from an identical twin will be accepted readily.
Chemical-Structural Complexity
Proteins are the most potent immunogens followed by polysaccharides. Nucleic acids and lipids are not efficient in eliciting a good immune reaction, although they may act as haptens. Structural complexity of a protein contributes to its immunogenicity. Chains of single amino acids or single sugars are poorly immunogenic, but if different amino acids or sugars are combined in the same molecule, the immunogenicity is greatly enhanced.
In cell-mediated immunity, the response of T cells to the peptide component of the proteins depends on how the peptide is recognised and presented by the MHC cells. Therefore, the structure of protein plays an important role in its immunogenicity, especially in inducing cellular immunity.
The lipid-specific antibodies are not easily produced; hence, they do not play a major role in immunity. However, these antibodies have a role in the measurement of certain lipid-based molecules and drugs. These antibodies are produced first by treating lipids with haptens and then conjugating with suitable carrier molecules, such as the proteins (e.g., haemocyanin or bovine serum albumin).
Stability
Highly stable and non-degradable substances (e.g., some plastics, metals, or chains of d-amino acids) are not immunogenic. This is because internalisation, processing and presentation by antigen-presenting cells (APCs) are always essential to mount an immune response. Therefore, very stable substances (such as silicon) have been successful as non-immunogenic materials for reconstructive surgeries, such as breast implants.
On the other hand, if a substance is very unstable, it may break up before an APC can be internalised, and hence become immunogenic. In addition, large, insoluble complexes are more immunogenic than smaller, soluble ones. This is because macrophages find it easier to phagocytose, degrade and present the insoluble complexes than the soluble complexes.
Other Factors
Biological system: Biological system also plays an important role in determining the immunological efficiency of an antigen. Some substances are immunogenic in one individual but not in others (i.e., responders and non-responders). This is due to the fact that individuals may lack or have altered genes that code for the receptors for antigen on B cells and T cells, or they may not have the appropriate genes needed for the APC to present antigen to the helper T (TH) cells.
Dosage and route of the antigen: The dose of antigen and the route by which it comes into contact with the immune system also influence immunogenicity of the antigen. Very low doses of antigen do not stimulate immune response, either because too few lymphocytes are contacted or because a non-responsive state is elicited. Conversely, an extremely high dose also fails to elicit tolerance.
Repeated administration of antigens (booster doses) may be required to enhance immune response of the host to certain antigens. This is particularly important in case of vaccines where a pre-requisite immune level needs to be attained. Hence the booster doses of vaccines, such as DPT (diphtheria, pertussis, tetanus), DT (diphtheria, tetanus), etc., are given to ensure good protective levels of antibodies. Generally, antigens are administered by the parenteral route to produce good level of antibodies. The antigens can be given by (1) intravenous, (2) subcutaneous, (3) intradermal, (4) intramuscular, (5) intra-peritoneal and (6) mucosal routes. Usually, the subcutaneous route of administration proves to be better than intravenous routes at eliciting an immune response.
Adjuvants
Adjuvants are the substances that when mixed with an antigen and injected with it boost the immunogenicity of the antigen. Adjuvants increase both the strength and the duration of immune response. Adjuvants boost immunogenicity of antigens in several ways:
• Adjuvants such as aluminium potassium sulphate (alum) and Freund’s water-in-oil adjuvant prolong the persistence of antigen by forming a depot at the injection site. Alum precipitates the antigen and releases it a little at a time. The water-in-oil emulsion forms small droplets with the antigen and also releases these slowly over time.
• Freund’s complete adjuvant contains, in addition to the emulsifying factors, heat-killed mycobacteria. The bacterial components activate macrophages and increase both the production of IL-1 and the level of B7 membrane molecules, which enhances the immune response. The increased expression of Class-II MHC increases the ability of APC to present antigen to TH cells. B7 molecules on the APC bind to CD28, a cell-surface protein on TH cells, triggering costimulation, an enhancement of the T-cell immune response.
• Some adjuvants, such as synthetic polyribonucleotides and bacterial lipopolysaccharides, stimulate non-specific lymphocyte proliferation and bring about their action.
Antigenic Specificity
Antigenic specificity of the antigen depends on antigenic determinants or epitopes.
Epitopes
An epitope is defined as the immunologically active region of an immunogen that binds to antigen-specific membrane receptors on lymphocytes or secreted antibodies. The interaction between cells of the immune system and antigens takes place at many levels and the complexity of any antigen is mirrored by its epitope (Fig. 12-2). There are two types of epitopes: B-cell epitopes and T-cell epitopes.
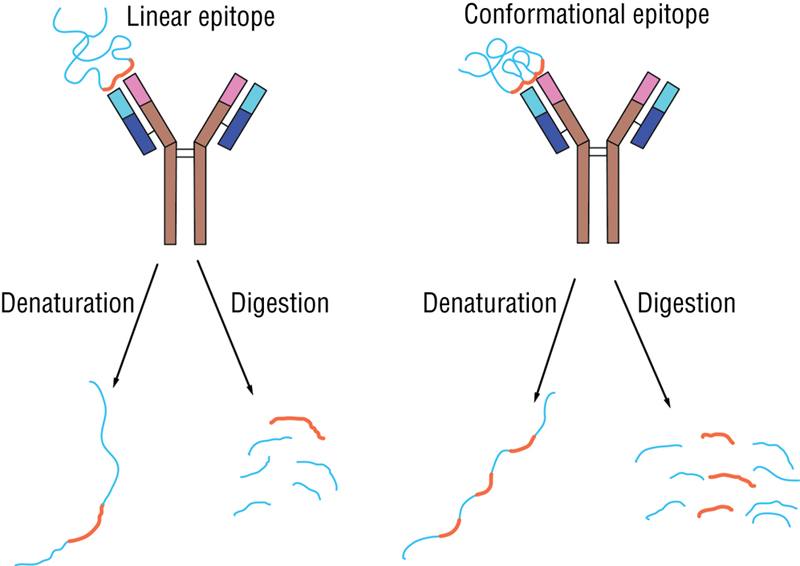
Fig. 12-2 Epitopes (linear and conformational).
B-Cell Epitopes
B-cell epitopes are antigenic determinants recognised by B cells. B-cell epitope can combine with its receptor only if the antigen molecule is in its native state. The complementary surfaces of the antibody and the antigen molecules appear to be relatively flat. Smaller molecules often fit nicely within a particular depression or groove in the antigen-binding site of the antibody molecule. The B-cell epitope is about six or seven sugar residues or amino acids long. They are also often found in regions of proteins, which have a higher mobility; this may make it possible for an epitope to shift just a bit to fit into an almost-right site.
T-Cell Epitopes
T cells recognise amino acids in proteins but do not recognise polysaccharide or nucleic acid antigens. This is the reason why polysaccharides are considered as T cell -independent antigens and proteins as T-cell-dependent antigens. The primary sequence of amino acids in proteins determines the antigenic determinants recognised by T cells. Free peptides are not recognised by T cells, whereas the complex of MHC molecules and peptide are recognised by T cells. Thus for a T-cell response, it should recognise both the antigenic determinant and also the MHC, and therefore it is said to be MHC restricted. In general, T-cell epitopes or antigenic determinants are small and are only 8–15 amino acids long. The antigenic determinants are limited to those parts of the antigen that can bind to MHC molecules.
Processing of an antigen by APCs is absolutely essential for a T cell to recognise it. Two different types of processing can prepare a protein antigen for antigen presentation. These include:
• Externally derived antigens’ processing: In this process, phagocytosed bacteria are killed and lysed by phagocytic cells, such as macrophages. Pieces of the bacteria are then processed and presented in the context of Class-II MHC molecules.
• Endogenously derived antigens’ processing: In this process, virus proteins synthesised in a cell are processed and then presented in the context of Class-I MHC molecules.
Species Specificity
Tissues of all individuals in a species possess certain species-specific antigens. However, some degree of cross-reaction occurs between antigens from related species. The species specificity shows phylogenetic relationship. The phylogenetic relationship is useful in:
• tracing evolutionary relationship between species;
• the species identification from blood and seminal stains in forensic medicine.
Isospecificity
Isospecificity is determined by the presence of isoantigens or histocompatibility antigens.
Isoantigens: Isoantigens are antigens found in some, but not all, members of a species. A species may be grouped depending on the presence of different isoantigens in its members. These are genetically determined. Human erythrocyte antigens, on the basis of which individuals are classified into different blood groups, are the best examples of isoantigens in humans. The blood groups are of primary importance in:
• transfusion of blood and blood products;
• isoimmunisation during pregnancy and
• providing valuable evidence in paternity disputes, the results of which are supplemented by more recent DNA fingerprinting tests.
Histocompatibility Antigens
Histocompatibility antigens are the cellular determinants specific for each individual of a species. These antigens are associated with the plasma membrane of tissue cells. Human leukocyte antigen (HLA) is the major histocompatibility antigen that determines the homograft rejection. Therefore, HLA typing is absolutely essential before carrying out transplantation of tissue or organ from one individual to another.
Autospecificity
Self-antigens are generally non-antigenic. Sequestrated antigens (such as eye lens protein and sperm) are, however, exceptions, because these are not recognised as self-antigens. This is because corneal tissue and sperm are never encountered by the immune system during the development of tolerance to self-antigens. Therefore, these tissues become immunogenic if accidentally or experimentally released into the blood or tissues.
Organ Specificity
Antigens characteristic of an organ or tissues are called organ-specific antigens. These antigens found in the brain, kidney and lens tissues, even of different animal species, share the same antigen specificity. Organ-specific antigens, such as brain-specific antigens, shared by human and sheep brain are one such example. The antirabies vaccines prepared from sheep brain, when given, may induce immune response in some humans, causing damage to neural tissues of the recipient. This may result in neuroparalytic complications in some individuals.
Heterophile Specificity
Heterophile specificity is determined by the presence of heterophile antigens. The same or closely related antigens, sometimes present in tissues of different biological species, classes, or kingdoms are known as heterophile antigens. Antibodies against the heterophile antigens produced by one of the species cross-react with the antigens of other species. This property is exploited for diagnosis of many infectious diseases. Weil–Felix reaction, Paul–Bunnell test, and cold agglutination tests are the examples of serological tests that use such heterophile antigens.
• Weil–Felix reaction is a test used for diagnosis of rickettsial infections, in which the strains of Proteus species (such as OX 19, OX 2, and OX K) are used to detect heterophile antibodies produced against rickettsial pathogens.
• Paul–Bunnel test is used for diagnosis of infectious mononucleosis caused by Epstein–Barr virus infection by demonstration of heterophile antibodies that agglutinate sheep erythrocytes.
• Cold agglutinin test is performed for diagnosis of primary atypical pneumonia caused by Mycoplasma pneumoniae by demonstration of heterophilic antibodies.
Biological Classification of Antigens
These are classified based on their ability to induce antibody production
• T-cell-dependent antigens
• T-cell-independent antigens
T-cell-dependent antigens: Majority of naturally occurring proteins are T-cell-dependent antigens and these antigens cannot induce antibody production upon direct contact with B cells. T-dependent antigens are made up of few copies of several antigenic determinants. These antigens bind to Ig receptor present on the B cells, which are then internalised and processed into smaller peptides and then expressed on B cells surface. Further, B cells form a complex with MHC II and present themselves to T helper cells. Upon recognition of processed antigenic determinants TH cell secretes cytokines and express CD 40 ligands on it. Thereafter, TH cell interacts with CD 40 markers present on the B cells in presence of cytokines to produce antibodies against the specific antigens.
T cell-independent antigens: These antigens stimulate B cells to produce antibody without any direct involvement of T cells. Most of the microbial antigens such as carbohydrates, lipids and certain nucleic acids are T-independent antigens. Further, these antigens are of two types:
• Type-1 antigens include endotoxins and lipopolysaccharides (LPS) of microbes they are mitogenic in nature, thus help in B cell multiplication and polyclonal B cell activation.
• Type-2 antigens include polymeric compounds such as polysaccharides (pneumococcal capsular polysaccharides) or proteins (flagellar proteins). These antigens, along with cytokines activate B cells to produce specific antibodies and also augment functions of other immune cell such as dendritic cells, mast cells, macrophages and NK cells.
Superantigens
Superantigens are a class of molecules that can interact with APCs and T lymphocytes in a non-specific way. The superantigens act differently by interacting with MHC class II molecules of the APC and the Vb domain of the T-lymphocyte receptor (Fig. 12-3). This interaction results in the activation of a larger number of T cells (10%) than conventional antigens (1%), leading to massive cytokine expression and immunomodulation. Examples of superantigens are staphylococcal enterotoxins, toxic shock syndrome toxin, exfoliative toxins, and also some viral proteins.

Pathogen Determinants Recognition by Innate Immune System
Usually, adaptive immune system reacts specifically to a particular pathogen by recognising its discrete antigenic determinants with the help of specific receptors present on the immune cells. Whereas the components of the innate immune system have the ability to recognise the unique molecular patterns which are shared by many pathogens. This unique molecular patterns shared by multiple pathogens are termed as pathogen associated molecular patterns (PAMPs) and the innate immune receptors that recognise PAMPs are called as pattern recognition receptors (PRRs). A single PRR can recognise a particular PAMP present on different pathogens. Mainly, PRRs are categorised into three classes.
• Toll-like receptors (TLRs): TLRs are transmembrane receptors present on the host immune cells such as dendritic cells and macrophages with unique pattern recognition capability. There are 13 different types of TLRs which can recognise different pathogens and enhance innate immune mechanism by phagocytosis and inflammation at the site of infection.
• Scavenger receptors (SR): SRs are expressed by macrophages, dendritic cells and certain endothelial cells. They have the ability to bind to the bacterial cell wall components such as LPS, peptidoglycans as well as damaged or apoptotic cells. SRs are involved in lipid metabolism and play an important role in clearing damaged host cells and other molecules. Examples of scavenger receptors include CD-36, CD-68, SRB-1 etc.
• Mannose receptors: These are found on the surface of phagocytes and are capable of binding to the mannose rich glycans, the components of the microbial glycoproteins and glycolipids. Mannose rich glycans are short carbohydrate chains with mannose or fucose as the terminal sugar and they are usually present in microbes but not in humans.
Study questions
1. What are the determinants of antigenicity of the antigen?
Write about biological classification of antigens.
2. Short notes:
a. Eepitopes
b. Haptens
c. Superantigen
d. Heterospecificity of an antigen
Online study material
Multiple Choice Questions
1. The antigen that are not immunogenic but can take part in immune reactions are termed
A. Superantigens
B. Isoantigens
C. Heteroantigens
D. Haptens
2. The substances that are least ummunogenic are
A. Proteins
B. Polysaccharides
C. Nucleic acids
D. None of the above.
3. All the following statements are true for epitopes except for
A. Epitope called the antigenic determinant
B. Antigenic specificity does not depend on the epitope
C. Epitope is an immunologically active region of an immunogen.
D. An epitope binds to antigen-specific membrane receptors on lymphocytes.
4. All the following statements are true for histocompatibility antigens except
A. They are the cellular determinants specific for each individual of a species.
B. These are associated with the nuclear membrane.
C. HLA is the major histocompatibility antigen that determines the homograft rejection.
D. HLA typing is absolutely essential before carrying out transplantation of tissue.
5. The test that does not use heterophilic antigen is
A. TPHA test
B. Weil–Felix reaction
C. Paul–Bunnell test
D. Cold agglutination test
Keys: 1. D; 2. C; 3. B; 4. B; 5. A