4
Transport of Substances Through Cell Membranes
Figure 4-1 lists the approximate concentrations of important electrolytes and other substances in the extracellular fluid and intracellular fluid. The structure of the membrane covering the outside of every cell of the body is discussed in Chapter 2 and illustrated in Figures 2-3 and 4-2. This membrane not only consists almost entirely of a lipid bilayer but also contains large numbers of protein molecules in the lipid, many of which penetrate all the way through the membrane.

Figure 4-1 Chemical compositions of extracellular and intracellular fluids. The question mark indicates that precise values for intracellular fluid are unknown. The red line indicates the cell membrane.

Figure 4-2 Transport pathways through the cell membrane, and the basic mechanisms of transport.
The lipid bilayer is not miscible with either the extracellular fluid or the intracellular fluid. Therefore, it constitutes a barrier against movement of water molecules and water-soluble substances between the extracellular and intracellular fluid compartments. However, as demonstrated in Figure 4-2 by the leftmost arrow, lipid-soluble substances can penetrate this lipid bilayer, diffusing directly through the lipid substance itself.
The protein molecules in the membrane have entirely different properties for transporting substances. Their molecular structures interrupt the continuity of the lipid bilayer, constituting an alternative pathway through the cell membrane. Many of these penetrating proteins can function as transport proteins. Different proteins function differently. Some proteins have watery spaces all the way through the molecule and allow free movement of water as well as selected ions or molecules; these proteins are called channel proteins. Other proteins, called carrier proteins, bind with molecules or ions that are to be transported; conformational changes in the protein molecules then move the substances through the interstices of the protein to the other side of the membrane. Channel proteins and the carrier proteins are usually selective for the types of molecules or ions that are allowed to cross the membrane.
“Diffusion” Versus “Active Transport”
Transport through the cell membrane, either directly through the lipid bilayer or through the proteins, occurs via one of two basic processes: diffusion or active transport.
Although many variations of these basic mechanisms exist, diffusion means random molecular movement of substances molecule by molecule, either through intermolecular spaces in the membrane or in combination with a carrier protein. The energy that causes diffusion is the energy of the normal kinetic motion of matter.
In contrast, active transport means movement of ions or other substances across the membrane in combination with a carrier protein in such a way that the carrier protein causes the substance to move against an energy gradient, such as from a low-concentration state to a high-concentration state. This movement requires an additional source of energy besides kinetic energy. A more detailed explanation of the basic physics and physical chemistry of these two processes is provided in this chapter.
Diffusion
All molecules and ions in the body fluids, including water molecules and dissolved substances, are in constant motion, with each particle moving its separate way. The motion of these particles is what physicists call “heat”—the greater the motion, the higher is the temperature—and the motion never ceases except at absolute zero temperature. When a moving molecule, A, approaches a stationary molecule, B, the electrostatic and other nuclear forces of molecule A repel molecule B, transferring some of the energy of motion of molecule A to molecule B. Consequently, molecule B gains kinetic energy of motion, while molecule A slows down, losing some of its kinetic energy. As shown in Figure 4-3, a single molecule in a solution bounces among the other molecules first in one direction, then another, then another, and so forth, randomly bouncing thousands of times each second. This continual movement of molecules among one another in liquids or in gases is called diffusion.

Figure 4-3 Diffusion of a fluid molecule during one-thousandth of a second.
Ions diffuse in the same manner as whole molecules, and even suspended colloid particles diffuse in a similar manner, except that the colloids diffuse far less rapidly than do molecular substances because of their large size.
Diffusion Through the Cell Membrane
Diffusion through the cell membrane is divided into two subtypes called simple diffusion and facilitated diffusion. Simple diffusion means that kinetic movement of molecules or ions occurs through a membrane opening or through intermolecular spaces without any interaction with carrier proteins in the membrane. The rate of diffusion is determined by the amount of substance available, the velocity of kinetic motion, and the number and sizes of openings in the membrane through which the molecules or ions can move.
Facilitated diffusion requires interaction of a carrier protein. The carrier protein aids passage of the molecules or ions through the membrane by binding chemically with them and shuttling them through the membrane in this form.
Simple diffusion can occur through the cell membrane by two pathways: (1) through the interstices of the lipid bilayer if the diffusing substance is lipid soluble and (2) through watery channels that penetrate all the way through some of the large transport proteins, as shown to the left in Figure 4-2.
Diffusion of Lipid-Soluble Substances Through the Lipid Bilayer
An important factor that determines how rapidly a substance diffuses through the lipid bilayer is the lipid solubility of the substance. For instance, the lipid solubilities of oxygen, nitrogen, carbon dioxide, and alcohols are high, and all these substances can dissolve in the lipid bilayer and diffuse through the cell membrane in the same manner that diffusion of water solutes occurs in a watery solution. The rate of diffusion of each of these substances through the membrane is directly proportional to its lipid solubility. Especially large amounts of oxygen can be transported in this way; therefore, oxygen can be delivered to the interior of the cell almost as though the cell membrane did not exist.
Diffusion of Water and Other Lipid-Insoluble Molecules Through Protein Channels
Even though water is highly insoluble in the membrane lipids, it readily passes through channels in protein molecules that penetrate all the way through the membrane. Many of body’s cell membranes contain “pores” called aquaporins that selectively permit passage of water through the cell membrane. The aquaporins are highly specialized, and there are at least 13 different types in various cells of mammals.
The rapidity with which water molecules can diffuse through most cell membranes is astounding. For example, the total amount of water that diffuses in each direction through the red blood cell membrane during each second is about 100 times as great as the volume of the red blood cell itself.
Other lipid-insoluble molecules can pass through the protein pore channels in the same way as water molecules if they are water soluble and small enough. However, as they become larger, their penetration falls off rapidly. For instance, the diameter of the urea molecule is only 20% greater than that of water, yet its penetration through the cell membrane pores is about 1000 times less than that of water. Even so, given the astonishing rate of water penetration, this amount of urea penetration still allows rapid transport of urea through the membrane within minutes.
Diffusion Through Protein Pores and Channels—Selective Permeability and “Gating” of Channels
Computerized three-dimensional reconstructions of protein pores and channels have demonstrated tubular pathways all the way from the extracellular to the intracellular fluid. Therefore, substances can move by simple diffusion directly along these pores and channels from one side of the membrane to the other.
Pores are composed of integral cell membrane proteins that form open tubes through the membrane and are always open. However, the diameter of a pore and its electrical charges provide selectivity that permits only certain molecules to pass through. For example, protein pores called aquaporins or water channels permit rapid passage of water through cell membranes but exclude other molecules. At least 13 different types of aquaporins have been found in various cells of the human body. Aquaporins have a narrow pore that permits water molecules to diffuse through the membrane in single file. The pore is too narrow to permit passage of any hydrated ions. The density of some aquaporins (eg, aquaporin-2) in cell membranes is not static but is altered in different physiological conditions.
Selective Permeability of Protein Channels
Many of the protein channels are highly selective for transport of one or more specific ions or molecules. This selectivity results from the characteristics of the channel, such as its diameter, its shape, and the nature of the electrical charges and chemical bonds along its inside surfaces.
Potassium channels permit passage of potassium ions across the cell membrane about 1000 times more readily than they permit passage of sodium ions. This high degree of selectivity cannot be explained entirely by the molecular diameters of the ions because potassium ions are slightly larger than sodium ions. What is the mechanism for this remarkable ion selectivity? This question was partially answered when the structure of a bacterial potassium channel was determined by X-ray crystallography. Potassium channels were found to have a tetrameric structure consisting of four identical protein subunits surrounding a central pore (Figure 4-4). At the top of the channel pore are pore loops that form a narrow selectivity filter. Lining the selectivity filter are carbonyl oxygens. When hydrated potassium ions enter the selectivity filter, they interact with the carbonyl oxygens and shed most of their bound water molecules, permitting the dehydrated potassium ions to pass through the channel. The carbonyl oxygens are too far apart, however, to enable them to interact closely with the smaller sodium ions, which are therefore effectively excluded by the selectivity filter from passing through the pore.

Figure 4-4 The structure of a potassium channel. The channel is composed of four subunits (only two of which are shown), each with two transmembrane helices. A narrow selectivity filter is formed from the pore loops and carbonyl oxygens line the walls of the selectivity filter, forming sites for transiently binding dehydrated potassium ions. The interaction of the potassium ions with carbonyl oxygens causes the potassium ions to shed their bound water molecules, permitting the dehydrated potassium ions to pass through the pore.
Different selectivity filters for the various ion channels are believed to determine, in large part, the specificity of the various channels for cations or anions or for particular ions, such as sodium (Na+), potassium (K+), and calcium (Ca++) that gain access to the channels.
One of the most important of the protein channels, the sodium channel, is only 0.3 × 0.5 nm in diameter, but more important, the inner surfaces of this channel are lined with amino acids that are strongly negatively charged, as shown by the negative signs inside the channel proteins in the top panel of Figure 4-5. These strong negative charges can pull small dehydrated sodium ions into these channels, actually pulling the sodium ions away from their hydrating water molecules. Once in the channel, the sodium ions diffuse in either direction according to the usual laws of diffusion. Thus, the sodium channel is specifically highly selective for passage of sodium ions.

Figure 4-5 Transport of sodium and potassium ions through protein channels. Also shown are conformational changes in the protein molecules to open or close “gates” guarding the channels.
Gating of Protein Channels
Gating of protein channels provides a means of controlling ion permeability of the channels. This mechanism is shown in both panels of Figure 4-5 for selective gating of sodium and potassium ions. It is believed that some of the gates are actual gate-like extensions of the transport protein molecule, which can close the opening of the channel or can be lifted away from the opening by a conformational change in the shape of the protein molecule itself.
The opening and closing of gates are controlled in two principal ways:
1. Voltage gating: In the case of voltage gating, the molecular conformation of the gate or of its chemical bonds responds to the electrical potential across the cell membrane. For instance, in the top panel of Figure 4-5, a strong negative charge on the inside of the cell membrane could cause the outside sodium gates to remain tightly closed; conversely, when the inside of the membrane loses its negative charge, these gates would open suddenly and allow sodium to pass inward through the sodium pores. This process is the basic mechanism for eliciting action potentials in nerves that are responsible for nerve signals. In the bottom panel of Figure 4-5, the potassium gates are on the intracellular ends of the potassium channels, and they open when the inside of the cell membrane becomes positively charged. The opening of these gates is partly responsible for terminating the action potential, a process discussed more fully in Chapter 9.
2. Chemical (ligand) gating: Some protein channel gates are opened by the binding of a chemical substance (a ligand) with the protein, which causes a conformational or chemical bonding change in the protein molecule that opens or closes the gate. One of the most important instances of chemical gating is the effect of acetylcholine on the so-called acetylcholine channel. Acetylcholine opens the gate of this channel, providing a negatively charged pore of about 0.65 nm in diameter that allows uncharged molecules or positive ions smaller than this diameter to pass through. This gate is exceedingly important for the transmission of nerve signals from one nerve cell to another (see Chapter 101) and from nerve cells to muscle cells to cause muscle contraction (see Chapter 12).
Open State Versus Closed State of Gated Channels
Figure 4-6A displays an interesting characteristic of most voltage-gated channels. This figure shows two recordings of electrical current flowing through a single sodium channel when there was an approximate 25-mV potential gradient across the membrane. Note that the channel conducts current “in an all-or-none” fashion. That is, the gate of the channel snaps open and then snaps close, with each open state lasting for only a fraction of a millisecond up to several milliseconds demonstrating the rapidity with which changes can occur during the opening and closing of the protein molecular gates. At one voltage potential, the channel may remain closed all the time or almost all the time, whereas at another voltage level, it may remain open either all or most of the time. At in-between voltages, as shown in the figure, the gates tend to snap open and close intermittently, resulting in an average current flow somewhere between the minimum and the maximum.

Figure 4-6 (A) A recording of current flow through a single voltage-gated sodium channel, demonstrating the “all-or-none” principle for opening and closing of the channel. (B) The “patch-clamp” method for recording current flow through a single protein channel. To the left, recording is performed from a “patch” of a living cell membrane. To the right, recording is from a membrane patch that has been torn away from the cell.
Patch-Clamp Method for Recording Ion Current Flow Through Single Channels
The patch-clamp method for recording ion current flow through single protein channels is illustrated in Figure 4-6A and B. A micropipette, with a tip diameter of only 1 or 2 μm, is abutted against the outside of a cell membrane. Suction is then applied inside the pipette to pull the membrane against the tip of the pipette, which creates a seal where the edges of the pipette touch the cell membrane. The result is a minute membrane “patch” at the tip of the pipette through which electrical current flow can be recorded.
Alternatively, as shown at the bottom right in Figure 4-6B, the small cell membrane patch at the end of the pipette can be torn away from the cell. The pipette with its sealed patch is then inserted into a free solution, which allows the concentrations of ions both inside the micropipette and in the outside solution to be altered as desired. Also, the voltage between the two sides of the membrane can be set or “clamped” to a given voltage.
Facilitated Diffusion Requires Membrane Carrier Proteins
Facilitated diffusion is also called carrier-mediated diffusion because a substance transported in this manner diffuses through the membrane with the help of a specific carrier protein.
Facilitated diffusion differs from simple diffusion in the following important way: Although the rate of simple diffusion through an open channel increases proportionately with the concentration of the diffusing substance, in facilitated diffusion the rate of diffusion approaches a maximum, called Vmax, as the concentration of the diffusing substance increases. This difference between simple diffusion and facilitated diffusion is demonstrated in Figure 4-7. The figure shows that as the concentration of the diffusing substance increases the rate of simple diffusion continues to increase proportionately, but in the case of facilitated diffusion the rate of diffusion cannot rise greater than the Vmax level.
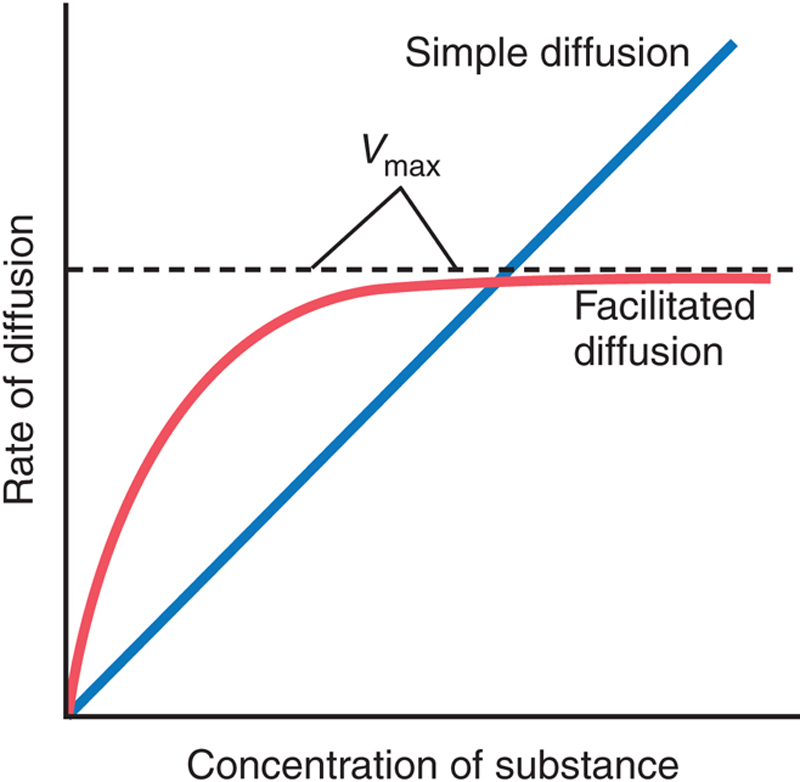
Figure 4-7 The effect of concentration of a substance on the rate of diffusion through a membrane by simple diffusion and facilitated diffusion. This graph shows that facilitated diffusion approaches a maximum rate called the Vmax.
What is it that limits the rate of facilitated diffusion? A probable answer is the mechanism illustrated in Figure 4-8. This figure shows a carrier protein with a pore large enough to transport a specific molecule partway through. It also shows a binding “receptor” on the inside of the protein carrier. The molecule to be transported enters the pore and becomes bound. Then, in a fraction of a second, a conformational or chemical change occurs in the carrier protein, so the pore now opens to the opposite side of the membrane. Because the binding force of the receptor is weak, the thermal motion of the attached molecule causes it to break away and be released on the opposite side of the membrane. The rate at which molecules can be transported by this mechanism can never be greater than the rate at which the carrier protein molecule can undergo change back and forth between its two states. Note specifically, though, that this mechanism allows the transported molecule to move—that is, to “diffuse”—in either direction through the membrane.

Figure 4-8 The postulated mechanism for facilitated diffusion.
Among the many important substances that cross cell membranes by facilitated diffusion are glucose and most of the amino acids. In the case of glucose at least 14 members of a family of membrane proteins (called GLUT) that transport glucose transporter molecules have been discovered in various tissues. Some of these GLUT can also transport other monosaccharides that have structures similar to that of glucose, including galactose and fructose. One of these, glucose transporter 4 (GLUT4), is activated by insulin, which can increase the rate of facilitated diffusion of glucose as much as 10- to 20-fold in insulin-sensitive tissues. This is the principal mechanism by which insulin controls glucose use in the body, as discussed in Chapter 93.
A comparison of simple and facilitated diffusion is summarized in Table 4-1.
Table 4-1
Comparison of Simple and Facilitated Diffusion
| Simple Diffusion | Facilitated Diffusion | |
| Movement | Along concentration gradient | Along concentration gradient |
| Energy required | No | No |
| Carrier protein required | No | Yes |
| Saturation kinetics | No | Yes |
| Type of molecules transported | Small, lipid soluble | Large, lipid insoluble |
Factors that Affect Net Rate of Diffusion
By now it is evident that many substances can diffuse through the cell membrane. What is usually important is the net rate of diffusion of a substance in the desired direction. This net rate is determined by several factors.
Net Diffusion Rate Is Proportional to the Concentration Difference Across a Membrane
Figure 4-9A shows a cell membrane with a high concentration of a substance on the outside and a low concentration on the inside. The rate at which the substance diffuses inward is proportional to the concentration of molecules on the outside because this concentration determines how many molecules strike the outside of the membrane each second. Conversely, the rate at which molecules diffuse outward is proportional to their concentration inside the membrane. Therefore, the rate of net diffusion into the cell is proportional to the concentration on the outside minus the concentration on the inside, or:

Figure 4-9 The effect of concentration difference (A), electrical potential difference affecting negative ions (B), and pressure difference (C) to cause diffusion of molecules and ions through a cell membrane.

where Co is concentration outside and Ci is concentration inside.
Effect of Membrane Electrical Potential on Diffusion of Ions—The “Nernst Potential”
If an electrical potential is applied across the membrane, as shown in Figure 4-9B, the electrical charges of the ions cause them to move through the membrane even though no concentration difference exists to cause movement. Thus, in the left panel of Figure 4-9B, the concentration of negative ions is the same on both sides of the membrane, but a positive charge has been applied to the right side of the membrane and a negative charge has been applied to the left, creating an electrical gradient across the membrane. The positive charge attracts the negative ions, whereas the negative charge repels them. Therefore, net diffusion occurs from left to right. After some time large quantities of negative ions have moved to the right, creating the condition shown in the right panel of Figure 4-9B, in which a concentration difference of the ions has developed in the direction opposite to the electrical potential difference. The concentration difference now tends to move the ions to the left, while the electrical difference tends to move them to the right. When the concentration difference rises high enough, the two effects balance each other. At normal body temperature (37°C), the electrical difference that will balance a given concentration difference of univalent ions—such as Na+ ions—can be determined from the following formula, called the Nernst equation:

where EMF is the electromotive force (voltage) between side 1 and side 2 of the membrane, C1 is the concentration on side 1, and C2 is the concentration on side 2. This equation is extremely important in understanding the transmission of nerve impulses and is discussed in the context of the generation of the resting membrane potential in Chapter 7 and in the propagation of the nerve impulse in Chapter 10.
Effect of a Pressure Difference Across the Membrane
At times, considerable pressure difference develops between the two sides of a diffusible membrane. This pressure difference occurs, for instance, at the blood capillary membrane in all tissues of the body. The pressure is about 20 mmHg greater inside the capillary than outside.
Pressure actually means the sum of all the forces of the different molecules striking a unit surface area at a given instant. Therefore, having a higher pressure on one side of a membrane than on the other side means that the sum of all the forces of the molecules striking the channels on that side of the membrane is greater than on the other side. In most instances, this situation is caused by greater numbers of molecules striking the membrane per second on one side than on the other side. The result is that increased amounts of energy are available to cause net movement of molecules from the high-pressure side toward the low-pressure side. This effect is demonstrated in Figure 4-9C, which shows a piston developing high pressure on one side of a “pore,” thereby causing more molecules to strike the pore on this side and, therefore, more molecules to “diffuse” to the other side.
Osmosis Across Selectively Permeable Membranes—“Net Diffusion” of Water
By far the most abundant substance that diffuses through the cell membrane is water. Enough water ordinarily diffuses in each direction through the red blood cell membrane per second to equal about 100 times the volume of the cell itself. Yet normally the amount that diffuses in the two directions is balanced so precisely that zero net movement of water occurs. Therefore, the volume of the cell remains constant. However, under certain conditions, a concentration difference for water can develop across a membrane. When this concentration gradient for water develops, net movement of water does occur across the cell membrane, causing the cell to either swell or shrink, depending on the direction of the water movement. This process of net movement of water caused by a concentration difference of water is called osmosis.
To illustrate osmosis, let us assume the conditions shown in Figure 4-10, with pure water on one side of the cell membrane and a solution of sodium chloride on the other side. Water molecules pass through the cell membrane with ease, whereas sodium and chloride ions pass through only with difficulty. Therefore, sodium chloride solution is actually a mixture of permeant water molecules and nonpermeant sodium and chloride ions, and the membrane is said to be selectively permeable to water but much less so to sodium and chloride ions. Yet the presence of the sodium and chloride has displaced some of the water molecules on the side of the membrane where these ions are present and, therefore, has reduced the concentration of water molecules to less than that of pure water. As a result, in the example of Figure 4-10, more water molecules strike the channels on the left side, where there is pure water, than on the right side, where the water concentration has been reduced. Thus, net movement of water occurs from left to right—that is, osmosis occurs from the pure water into the sodium chloride solution.

Figure 4-10 Osmosis at a cell membrane when a sodium chloride solution is placed on one side of the membrane and water is placed on the other side.
Osmotic Pressure
If in Figure 4-10 pressure was applied to the sodium chloride solution, osmosis of water into this solution would be slowed, stopped, or even reversed. The amount of pressure required to stop osmosis is called the osmotic pressure of the sodium chloride solution.
The principle of a pressure difference opposing osmosis is demonstrated in Figure 4-11, which shows a selectively permeable membrane separating two columns of fluid, one containing pure water and the other containing a solution of water and any solute that will not penetrate the membrane. Osmosis of water from chamber B into chamber A causes the levels of the fluid columns to become farther and farther apart, until eventually a pressure difference develops between the two sides of the membrane great enough to oppose the osmotic effect. The pressure difference across the membrane at this point is equal to the osmotic pressure of the solution that contains the nondiffusible solute.

Figure 4-11 Demonstration of osmotic pressure caused by osmosis at a semipermeable membrane.
Importance of Number of Osmotic Particles (Molar Concentration) in Determining Osmotic Pressure
The osmotic pressure exerted by particles in a solution, whether they are molecules or ions, is determined by the number of particles per unit volume of fluid, not by the mass of the particles. The reason for this is that each particle in a solution, regardless of its mass, exerts, on average, the same amount of pressure against the membrane. That is, large particles, which have greater mass (m) than do small particles, move at slower velocities (v). The small particles move at higher velocities in such a way that their average kinetic energies (k), determined by the equation:

are the same for each small particle as for each large particle. Consequently, the factor that determines the osmotic pressure of a solution is the concentration of the solution in terms of number of particles (which is the same as its molar concentration if it is a nondissociated molecule), not in terms of mass of the solute.
“Osmolality”—The Osmole
To express the concentration of a solution in terms of numbers of particles the unit called the osmole is used in place of grams.
One osmole is 1 g molecular weight of osmotically active solute. Thus, 180 g of glucose, which is 1 g molecular weight of glucose, is equal to 1 osm of glucose because glucose does not dissociate into ions. If a solute dissociates into two ions, 1 g molecular weight of the solute will become 2 osm because the number of osmotically active particles is now twice as great as is the case for the nondissociated solute. Therefore, when fully dissociated, 1 g molecular weight of sodium chloride, 58.5 g, is equal to 2 osm.
Thus, a solution that has 1 osm of solute dissolved in each kilogram of water is said to have an osmolality of 1 osm/kg, and a solution that has 1/1000 osm dissolved per kilogram has an osmolality of 1 mOsm/kg. The normal osmolality of the extracellular and intracellular fluids is about 300 mOsm/kg of water.
Relation of Osmolality to Osmotic Pressure
At normal body temperature, 37°C, a concentration of 1 osm/L will cause 19,300 mmHg osmotic pressure in the solution. Likewise, 1 mOsm/L concentration is equivalent to 19.3 mmHg osmotic pressure. Multiplying this value by the 300-mOsm concentration of the body fluids gives a total calculated osmotic pressure of the body fluids of 5790 mmHg. The measured value for this, however, averages only about 5500 mmHg. The reason for this difference is that many of the ions in the body fluids, such as sodium and chloride ions, are highly attracted to one another; consequently, they cannot move entirely unrestrained in the fluids and create their full osmotic pressure potential. Therefore, on average, the actual osmotic pressure of the body fluids is about 0.93 times the calculated value.
The Term “Osmolarity”
Osmolarity is the osmolar concentration expressed as osmoles per liter of solution rather than osmoles per kilogram of water. Although, strictly speaking, it is osmoles per kilogram of water (osmolality) that determines osmotic pressure for dilute solutions such as those in the body, the quantitative differences between osmolarity and osmolality are less than 1%. Because it is far more practical to measure osmolarity than osmolality, measuring osmolarity is the usual practice in almost all physiological studies.
“Active Transport” of Substances Through Membranes
At times, a large concentration of a substance is required in the intracellular fluid even though the extracellular fluid contains only a small concentration. This situation is true, for instance, for potassium ions. Conversely, it is important to keep the concentrations of other ions very low inside the cell even though their concentrations in the extracellular fluid are great. This situation is especially true for sodium ions. Neither of these two effects could occur by simple diffusion because simple diffusion eventually equilibrates concentrations on the two sides of the membrane. Instead, some energy source must cause excess movement of potassium ions to the inside of cells and excess movement of sodium ions to the outside of cells. When a cell membrane moves molecules or ions “uphill” against a concentration gradient (or “uphill” against an electrical or pressure gradient), the process is called active transport.
Different substances that are actively transported through at least some cell membranes include sodium, potassium, calcium, iron, hydrogen, chloride, iodide, and urate ions, several different sugars, and most of the amino acids.
Primary Active Transport and Secondary Active Transport
Active transport is divided into two types according to the source of the energy used to facilitate the transport: primary active transport and secondary active transport. In primary active transport, the energy is derived directly from breakdown of adenosine triphosphate (ATP) or some other high-energy phosphate compound. In secondary active transport, the energy is derived secondarily from energy that has been stored in the form of ionic concentration differences of secondary molecular or ionic substances between the two sides of a cell membrane, created originally by primary active transport. In both instances, transport depends on carrier proteins that penetrate through the cell membrane, as is true for facilitated diffusion. However, in active transport, the carrier protein functions differently from the carrier in facilitated diffusion because it is capable of imparting energy to the transported substance to move it against the electrochemical gradient. The following sections provide some examples of primary active transport and secondary active transport, with more detailed explanations of their principles of function.
Primary Active Transport
Sodium–Potassium Pump Transports Sodium Ions Out of Cells and Potassium Ions into Cells
Among the substances that are transported by primary active transport are sodium, potassium, calcium, hydrogen, chloride, and a few other ions.
The active transport mechanism that has been studied in greatest detail is the sodium–potassium (Na+–K+) pump, a transport process that pumps sodium ions outward through the cell membrane of all cells and at the same time pumps potassium ions from the outside to the inside. This pump is responsible for maintaining the sodium and potassium concentration differences across the cell membrane, as well as for establishing a negative electrical voltage inside the cells as discussed in Chapter 7.
Figure 4-12 shows the basic physical components of the Na+–K+ pump. The carrier protein is a complex of two separate globular proteins: a larger one called the α subunit, with a molecular weight of about 100,000, and a smaller one called the β subunit, with a molecular weight of about 55,000. Although the function of the smaller protein is not known (except that it might anchor the protein complex in the lipid membrane), the larger protein has three specific features that are important for the functioning of the pump:
1. It has three binding sites for sodium ions on the portion of the protein that protrudes to the inside of the cell.
2. It has two binding sites for potassium ions on the outside.
3. The inside portion of this protein near the sodium binding sites has adenosine triphosphatse (ATPase) activity.

Figure 4-12 The postulated mechanism of the sodium–potassium pump. ADP, adenosine diphosphate; ATP, adenosine triphosphate; Pi, phosphate ion.
When two potassium ions bind on the outside of the carrier protein and three sodium ions bind on the inside, the ATPase function of the protein becomes activated. Activation of the ATPase function leads to cleavage of one molecule of ATP, splitting it to adenosine diphosphate (ADP) and liberating a high-energy phosphate bond of energy. This liberated energy is then believed to cause a chemical and conformational change in the protein carrier molecule, extruding the three sodium ions to the outside and the two potassium ions to the inside.
As with other enzymes, the Na+–K+ ATPase pump can run in reverse. If the electrochemical gradients for Na+ and K+ are experimentally increased to the degree that the energy stored in their gradients is greater than the chemical energy of ATP hydrolysis, these ions will move down their concentration gradients and the Na+–K+ pump will synthesize ATP from ADP and phosphate. The phosphorylated form of the Na+–K+ pump, therefore, can either donate its phosphate to ADP to produce ATP or use the energy to change its conformation and pump Na+ out of the cell and K+ into the cell. The relative concentrations of ATP, ADP, and phosphate, as well as the electrochemical gradients for Na+ and K+, determine the direction of the enzyme reaction. For some cells, such as electrically active nerve cells, 60–70% of the cells’ energy requirement may be devoted to pumping Na+ out of the cell and K+ into the cell.
The Na+–K+ Pump Is Important for Controlling Cell Volume
One of the most important functions of the Na+–K+ pump is to control the volume of each cell. Without function of this pump, most cells of the body would swell until they burst. The mechanism for controlling the volume is as follows: Inside the cell are large numbers of proteins and other organic molecules that cannot escape from the cell. Most of these proteins and other organic molecules are negatively charged and therefore attract large numbers of potassium, sodium, and other positive ions as well. All these molecules and ions then cause osmosis of water to the interior of the cell. Unless this process is checked, the cell will swell indefinitely until it bursts. The normal mechanism for preventing this outcome is the Na+–K+ pump. Note again that this device pumps three Na+ ions to the outside of the cell for every two K+ ions pumped to the interior. Also, the membrane is far less permeable to sodium ions than it is to potassium ions, and thus once the sodium ions are on the outside, they have a strong tendency to stay there. This process thus represents a net loss of ions out of the cell, which initiates osmosis of water out of the cell as well.
If a cell begins to swell for any reason, the Na+–K+ pump is automatically activated, moving still more ions to the exterior and carrying water with them. Therefore, the Na+–K+ pump performs a continual surveillance role in maintaining normal cell volume.
Electrogenic Nature of the Na+–K+ Pump
The fact that the Na+–K+ pump moves three Na+ ions to the exterior for every two K+ ions that are moved to the interior means that a net of one positive charge is moved from the interior of the cell to the exterior for each cycle of the pump. This action creates positivity outside the cell but results in a deficit of positive ions inside the cell; that is, it causes negativity on the inside. Therefore, the Na+–K+ pump is said to be electrogenic because it creates an electrical potential across the cell membrane. As discussed in Chapter 7 this electrical potential is a basic requirement in nerve and muscle fibers for transmitting nerve and muscle signals.
Secondary Active Transport—Cotransport and Countertransport
When sodium ions are transported out of cells by primary active transport, a large concentration gradient of sodium ions across the cell membrane usually develops with high concentration outside the cell and low concentration inside. This gradient represents a storehouse of energy because the excess sodium outside the cell membrane is always attempting to diffuse to the interior. Under appropriate conditions, this diffusion energy of sodium can pull other substances along with the sodium through the cell membrane. This phenomenon, called cotransport, is one form of secondary active transport.
For sodium to pull another substance along with it, a coupling mechanism is required, which is achieved by means of still another carrier protein in the cell membrane. The carrier in this instance serves as an attachment point for both the sodium ion and the substance to be cotransported. Once both of them are attached, the energy gradient of the sodium ion causes both the sodium ion and the other substance to be transported together to the interior of the cell.
In countertransport, sodium ions again attempt to diffuse to the interior of the cell because of their large concentration gradient. However, this time, the substance to be transported is on the inside of the cell and must be transported to the outside. Therefore, the sodium ion binds to the carrier protein where it projects to the exterior surface of the membrane, while the substance to be countertransported binds to the interior projection of the carrier protein. Once both have become bound, a conformational change occurs, and energy released by the action of the sodium ion moving to the interior causes the other substance to move to the exterior.
Cotransport of Glucose and Amino Acids Along with Sodium Ions
Glucose and many amino acids are transported into most cells against large concentration gradients; the mechanism of this action is entirely by cotransport, as shown in Figure 4-13. Note that the transport carrier protein has two binding sites on its exterior side, one for sodium and one for glucose. Also, the concentration of sodium ions is high on the outside and low inside, which provides energy for the transport. A special property of the transport protein is that a conformational change to allow sodium movement to the interior will not occur until a glucose molecule also attaches. When both of them become attached, the conformational change takes place, and the sodium and glucose are transported to the inside of the cell at the same time. Hence, this is a sodium–glucose cotransport mechanism. Sodium–glucose cotransporters are especially important mechanisms in transporting glucose across renal and intestinal epithelial cells, as discussed in Chapters 70 and 78.

Figure 4-13 The postulated mechanism for sodium cotransport of glucose.
Sodium cotransport of the amino acids occurs in the same manner as for glucose, except that it uses a different set of transport proteins. At least five amino acid transport proteins have been identified, each of which is responsible for transporting one subset of amino acids with specific molecular characteristics.
Sodium cotransport of glucose and amino acids occurs especially through the epithelial cells of the intestinal tract and the renal tubules of the kidneys to promote absorption of these substances into the blood. This is discussed in later chapters.
Other important cotransport mechanisms in at least some cells include cotransport of chloride, iodine, iron, and urate ions.
Active Transport Through Cellular Sheets
At many places in the body, substances must be transported all the way through a cellular sheet instead of simply through the cell membrane. Transport of this type occurs through the (1) intestinal epithelium, (2) epithelium of the renal tubules, (3) epithelium of all exocrine glands, (4) epithelium of the gallbladder, and (5) membrane of the choroid plexus of the brain, along with other membranes.
The basic mechanism for transport of a substance through a cellular sheet is (1) active transport through the cell membrane on one side of the transporting cells in the sheet, and then (2) either simple diffusion or facilitated diffusion through the membrane on the opposite side of the cell.
Figure 4-14 shows a mechanism for transport of sodium ions through the epithelial sheet of the intestines, gallbladder, and renal tubules. This figure shows that the epithelial cells are connected together tightly at the luminal pole by means of junctions. The brush border on the luminal surfaces of the cells is permeable to both sodium ions and water. Therefore, sodium and water diffuse readily from the lumen into the interior of the cell. Then, at the basal and lateral membranes of the cells, sodium ions are actively transported into the extracellular fluid of the surrounding connective tissue and blood vessels. This action creates a high sodium ion concentration gradient across these membranes, which in turn causes osmosis of water as well. Thus, active transport of sodium ions at the basolateral sides of the epithelial cells results in transport not only of sodium ions but also of water.

Figure 4-14 The basic mechanism of active transport across a layer of cells.
It is through these mechanisms that almost all nutrients, ions, and other substances are absorbed into the blood from the intestine. These mechanisms are also the way the same substances are reabsorbed from the glomerular filtrate by the renal tubules.
Vesicular Transport
In addition to the examples of transport described above, there is also a need to transport large molecules (macromolecules). This is achieved by two processes: endocytosis and exocytosis.
1. Endocytosis is the process by which large molecules are transported from the extracellular space into the cell by the invagination of the cell membrane around the molecule. The molecule can then be released inside the cell. Pinocytosis and phagocytosis are examples of endocytosis.
2. Exocytosis is the process by which large molecules are released to the exterior of the cell following fusion of a vesicle with the cell membrane. Hormone and neurotransmitter release are examples of exocytosis.
Numerous examples of the different types of transport discussed in this chapter are provided throughout this text.