Chapter 4
4.1 Introduction
4.2 History
Bentham and Hooker’s System
Table 4.1 Scheme of systematic classification of drugs |
||
| Division | Phanerogam | Phanerogam |
| Subdivision | Angiosperm | Angiosperm |
| Class | Dicotyledonae | Monocotyledonae |
| Subclass | Polypetalae | – |
| Series | Calyciflorae | Epigynae |
| Order | Resales | Scitamineae |
| Family | Leguminosae | Zingiberaceae |
| Subfamily | Caesalpinieae | – |
| Genus | Cassia | Zingiber |
| Species | angustifolia | officinalis |
Hutchinson’s System of Classification
4.3 Study of Different Tissue Systems
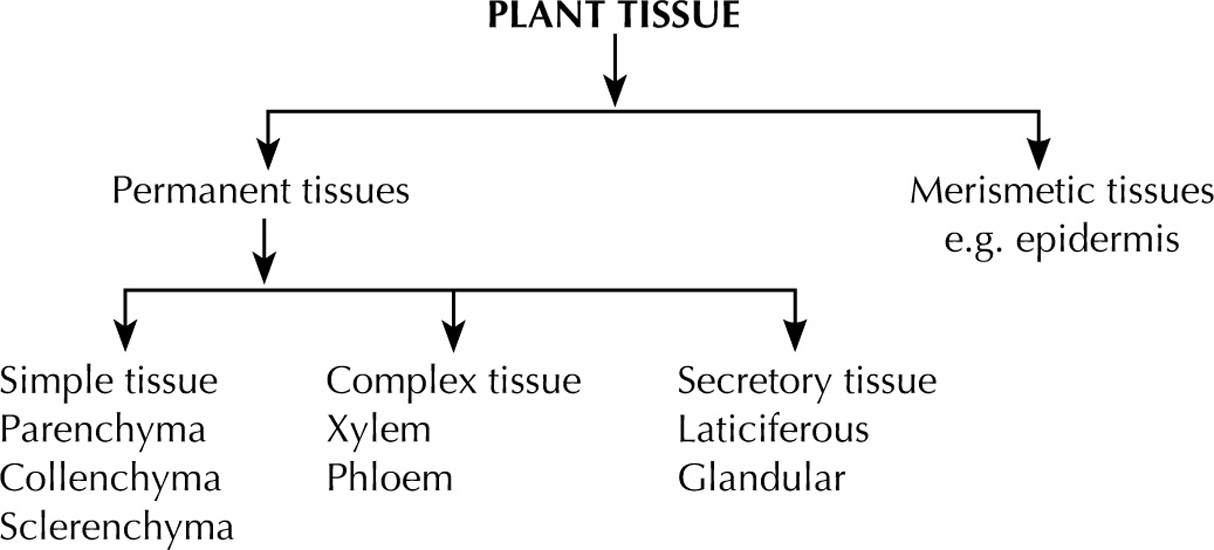
Difference between Merismetic and Permanent Tissues |
||
| Sr. No. | Merismetic Tissue | Permanent Tissue |
| 1. | Comprises of young cells which have the power to redivide and multiply. | These cells are living or dead having attained their definite form and size. |
| 2. | These cells are present at growing points, i.e. tips of roots, shoots and epidermis. | Usually present in the ground tissue and make the fundamental tissue system. |
| 3. | These cells are closely packed without intracellular spaces. | Intracellular spaces are present. |
Dermal Tissue System
Epidermis
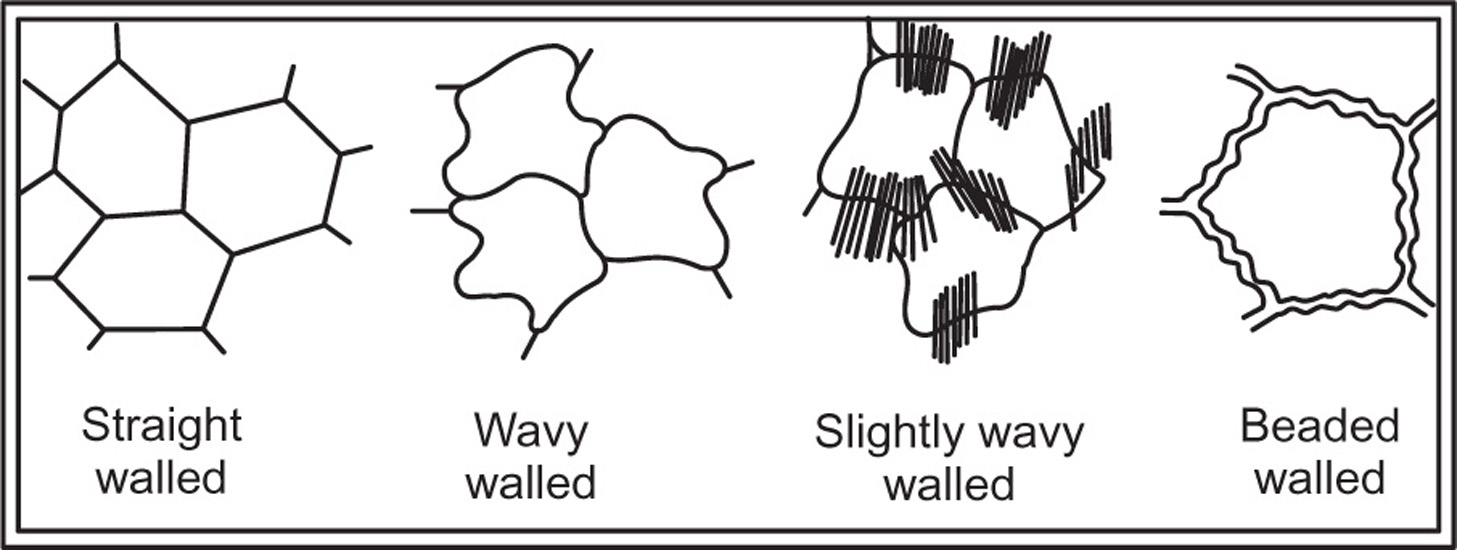
Fig. 4.1 Different type of cell walls of epidermis
Functions
Stomata
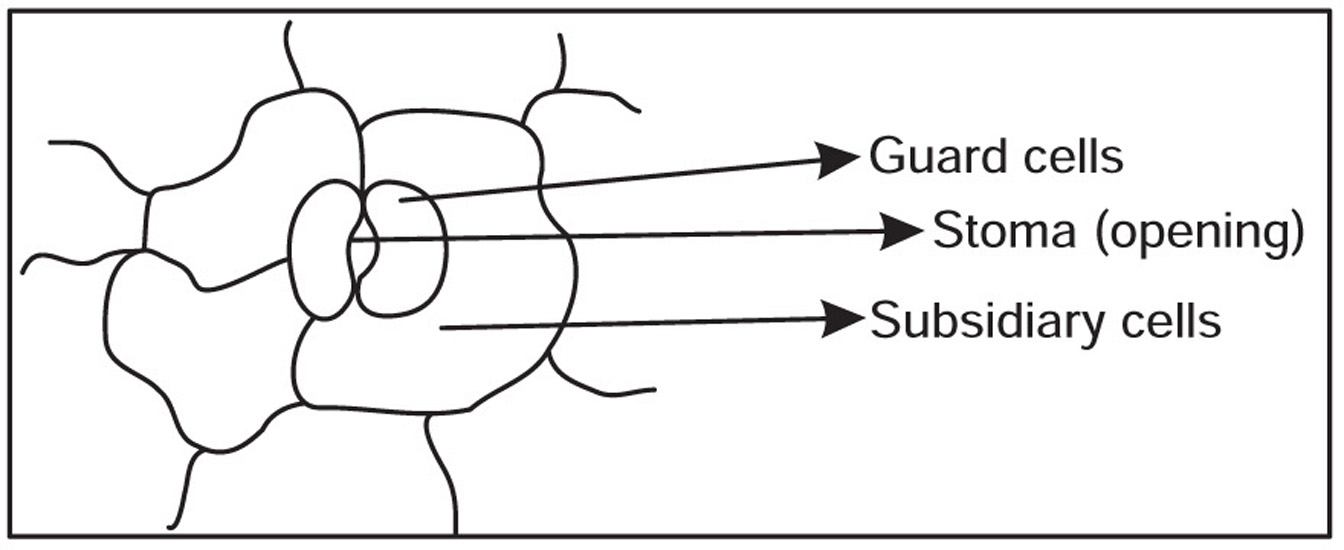
Fig. 4.2 Stomata
Trichomes
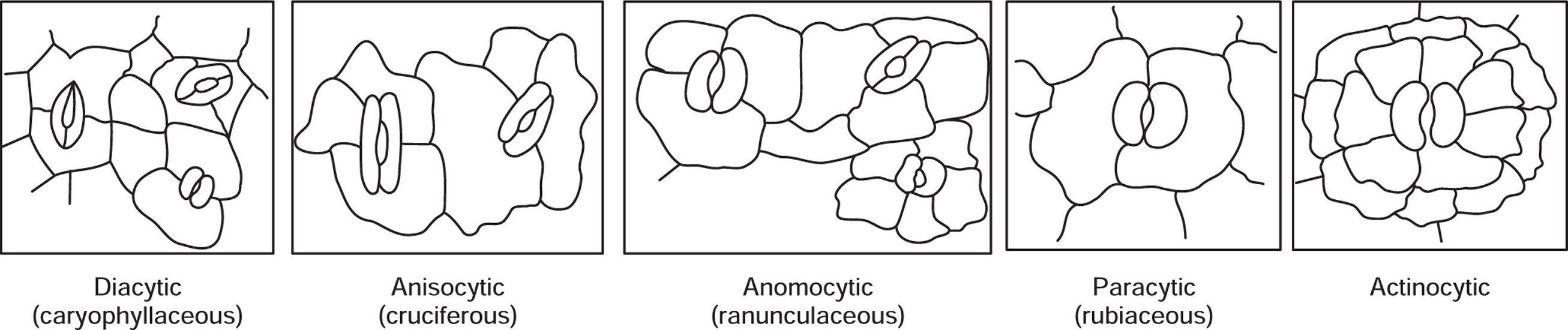
Fig. 4.3 Different types of stomata
[A] Covering Trichomes
[B] Glandular Trichomes
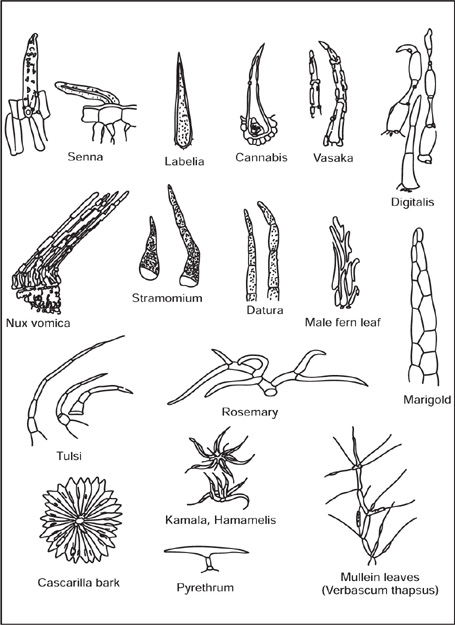
Fig. 4.4 Covering trichomes
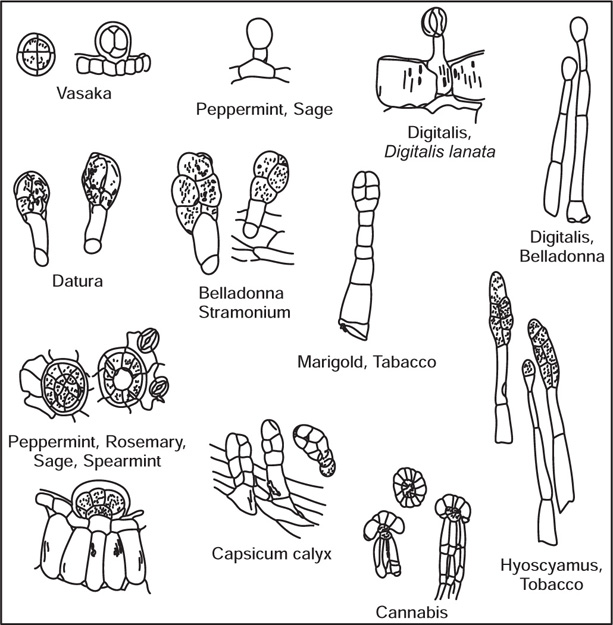
Fig. 4.5 Glandular trichomes
Periderm
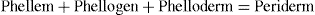
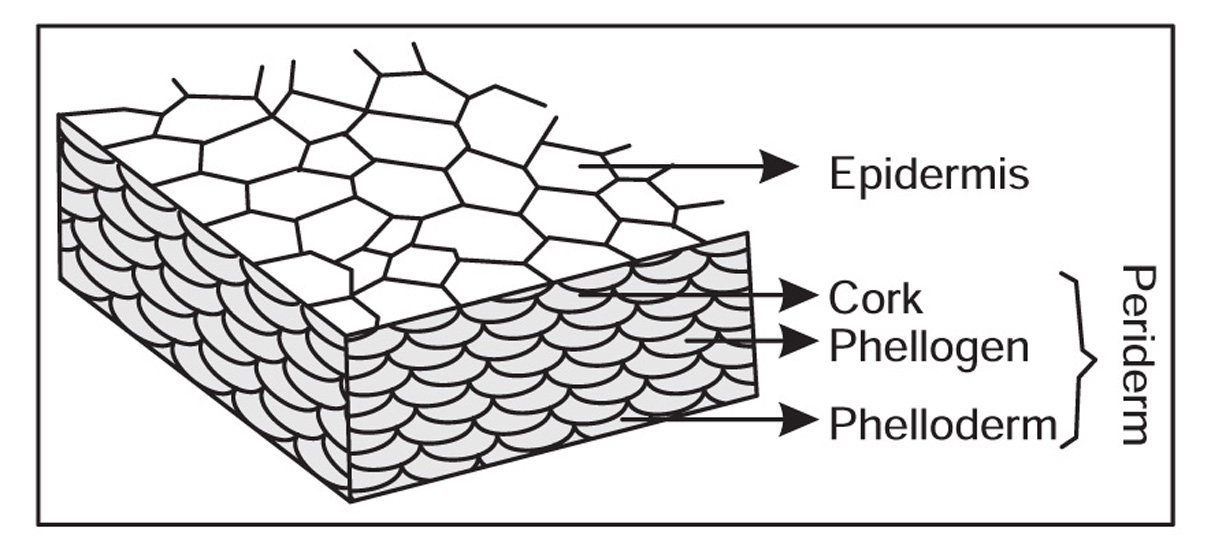
Fig. 4.6 Periderm
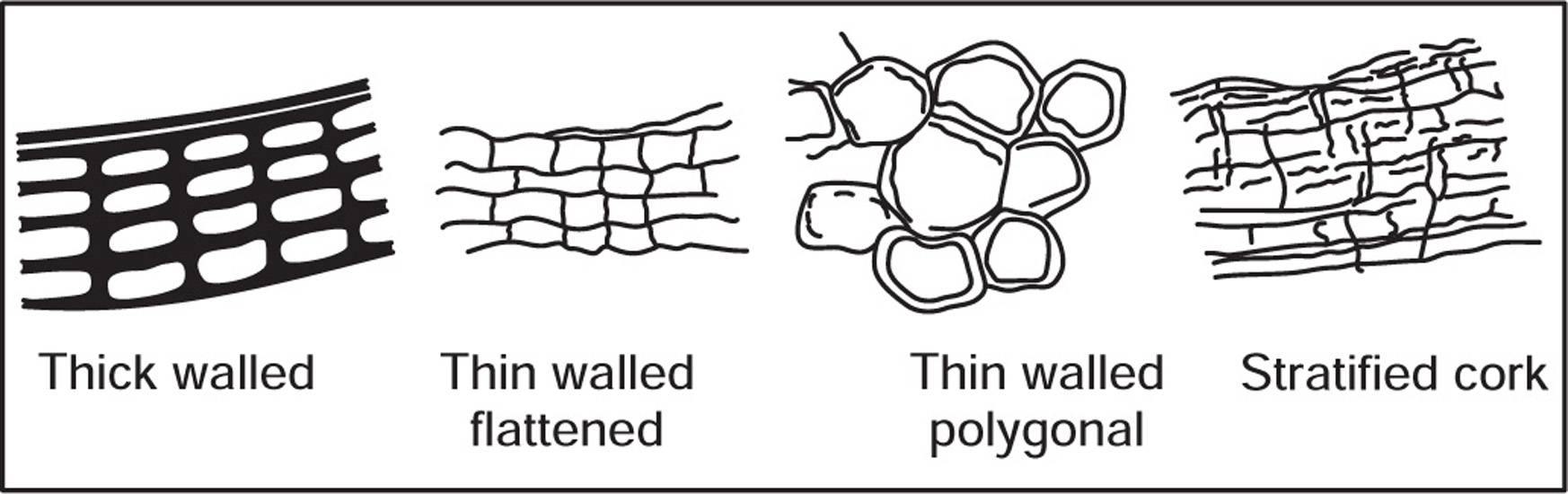
Fig. 4.7 Various types of cork cells
Vascular Tissue System
Function
[1] Xylem
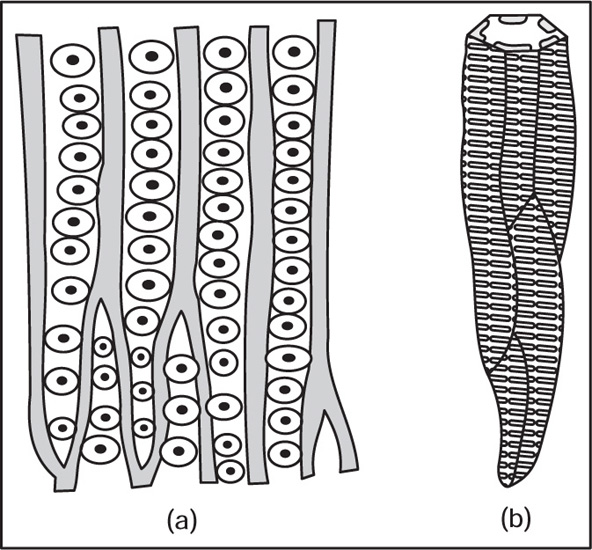
Fig. 4.8 (a) Tracheids with bordered pits (b) Scalariform tracheid
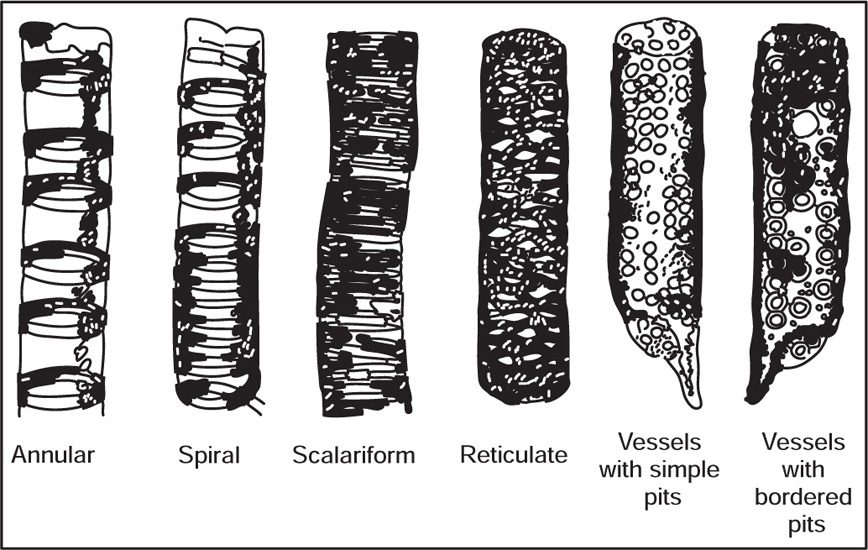
Fig. 4.9 Different kinds of vessels
[2] Phloem
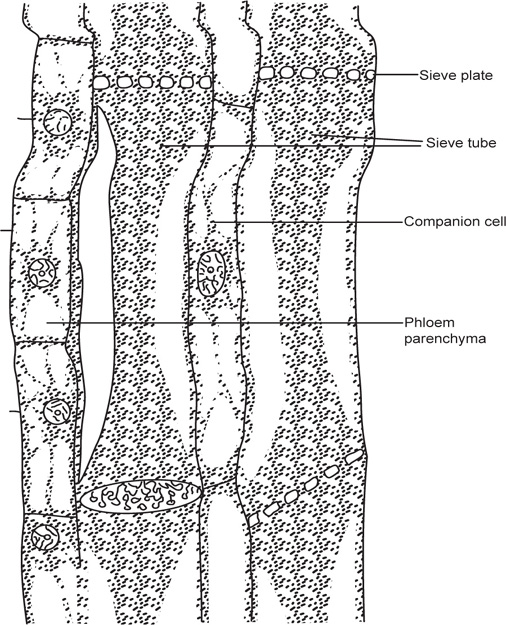
Fig. 4.10 A sieve tube in longitudinal section
[3] Cambium
Types of Vascular Bundles
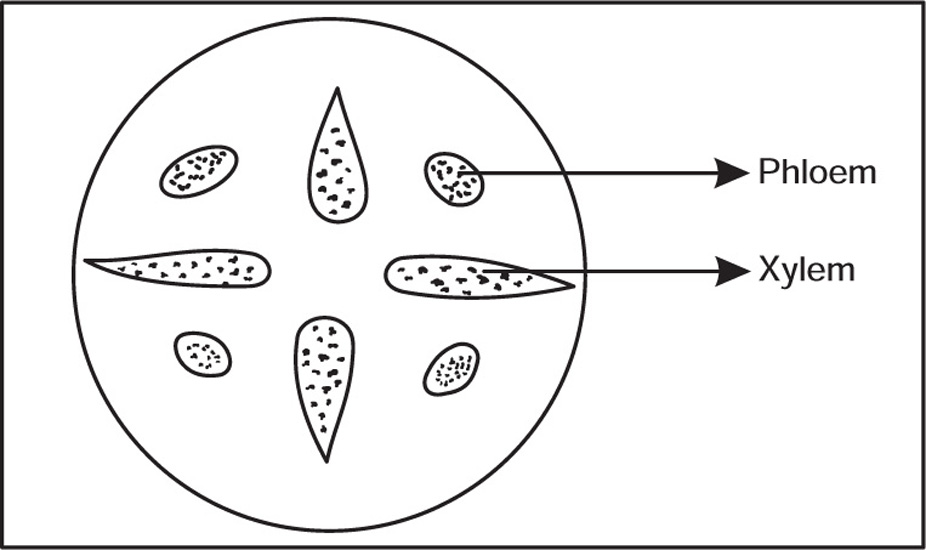
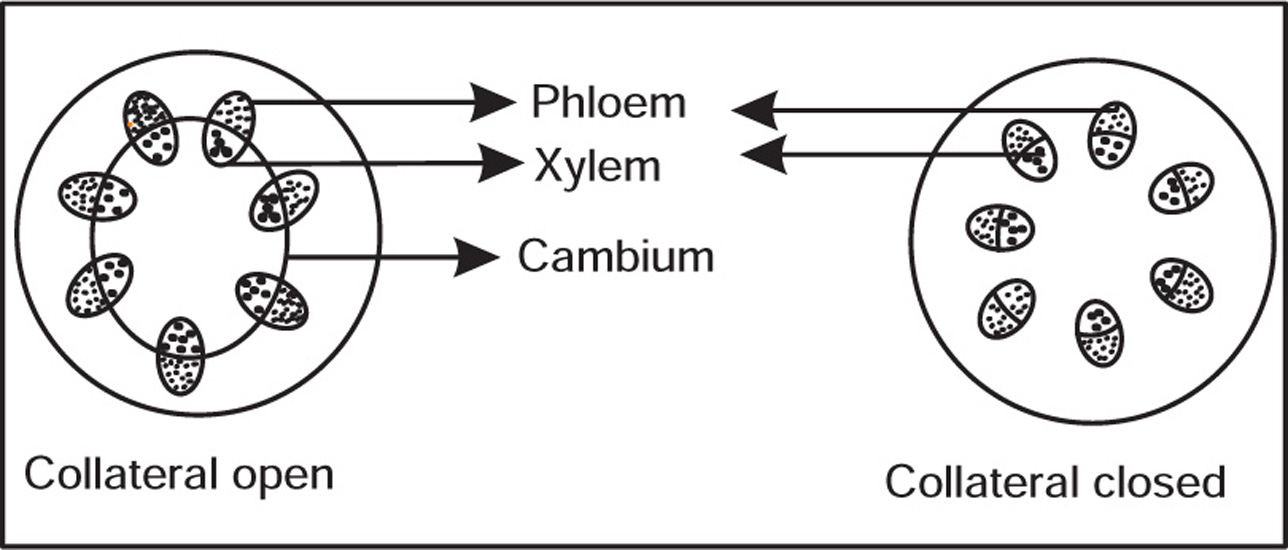
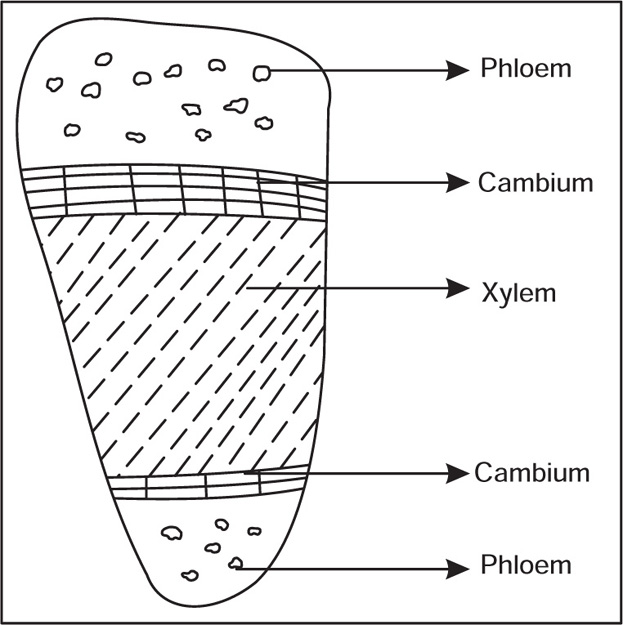
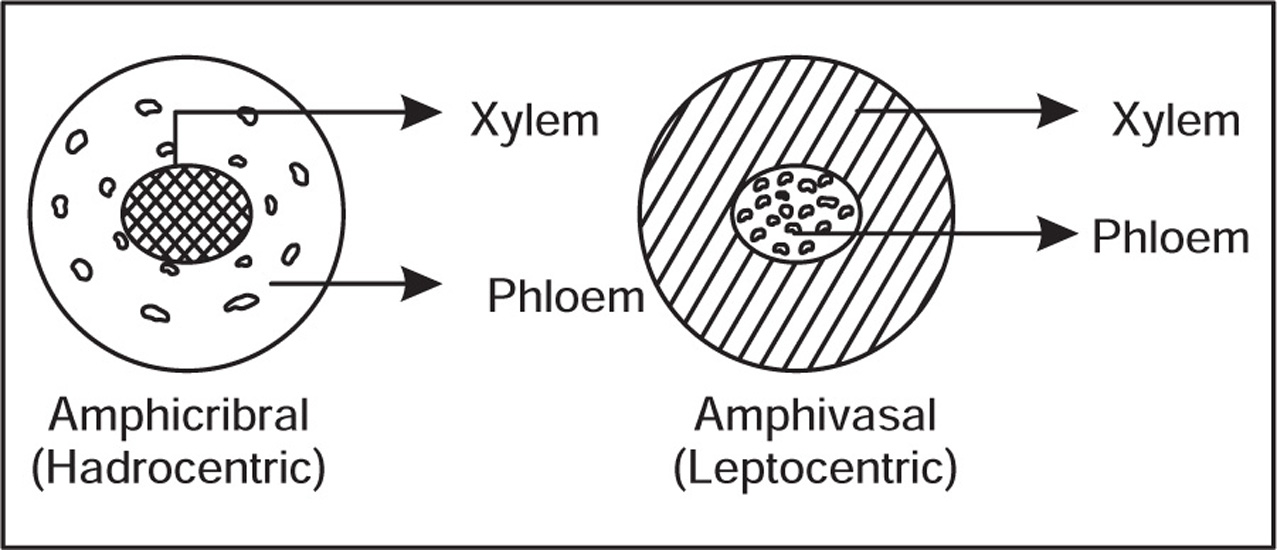
Ground Tissue System
(a) Parenchyma
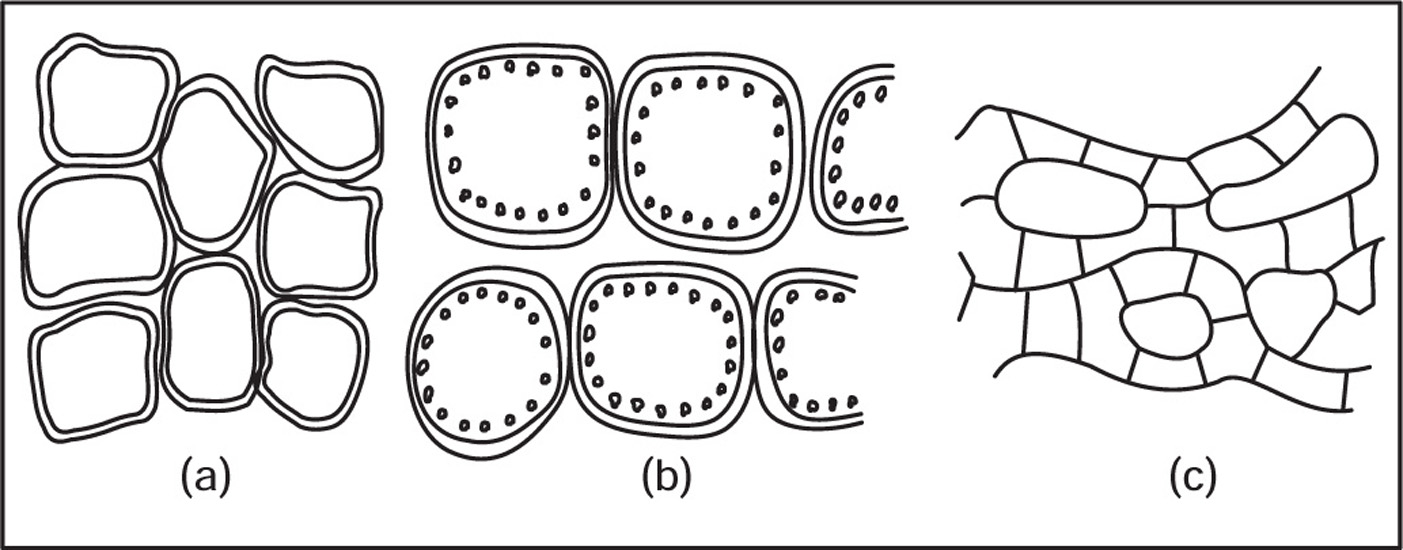
Fig. 4.11 (a) Parenchyma, (b) Chlorenchyma and (c) Aerenchyma
(b) Collenchyma
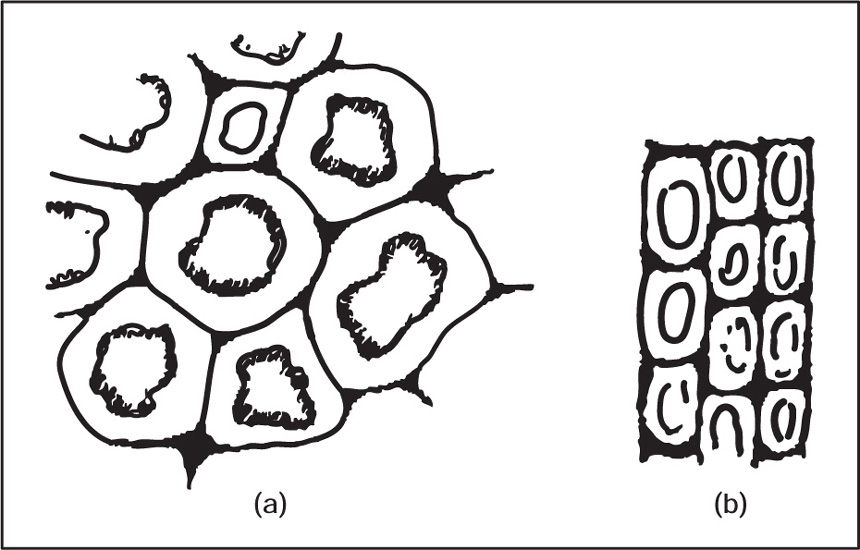
Fig. 4.12 (a) Collenchyma in transaction and (b) Collenchyma in longitudinal section
(c) Sclerenchyma
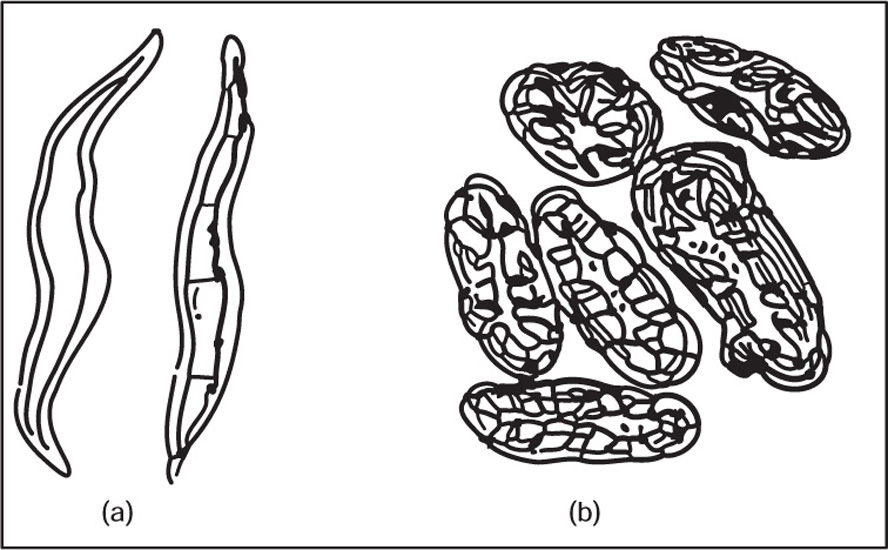
Fig. 4.13 (a) Sclerenchymatous fibres and (b) Sclereids (Stone cells)
4.4 Cell Contents
Starch
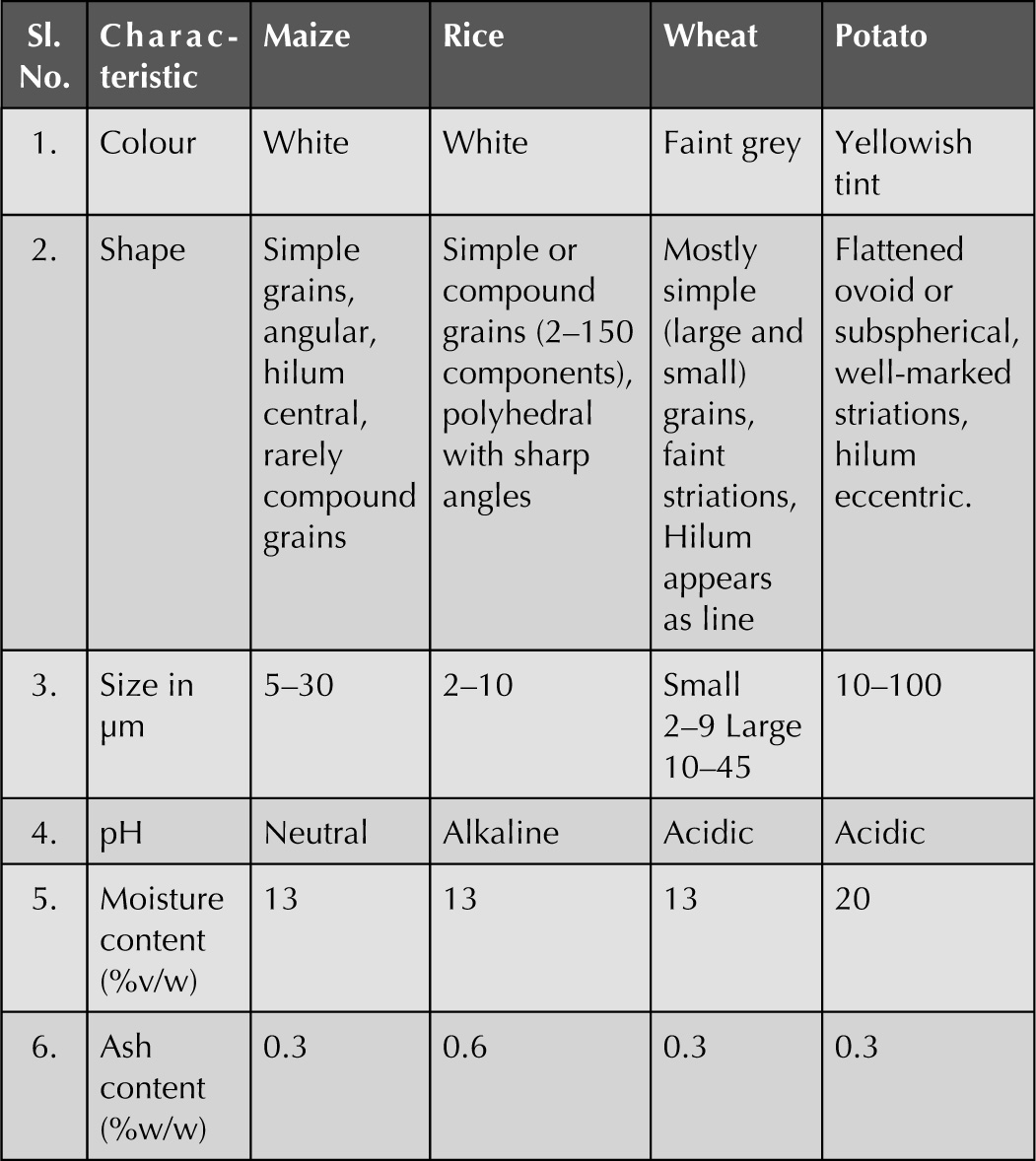
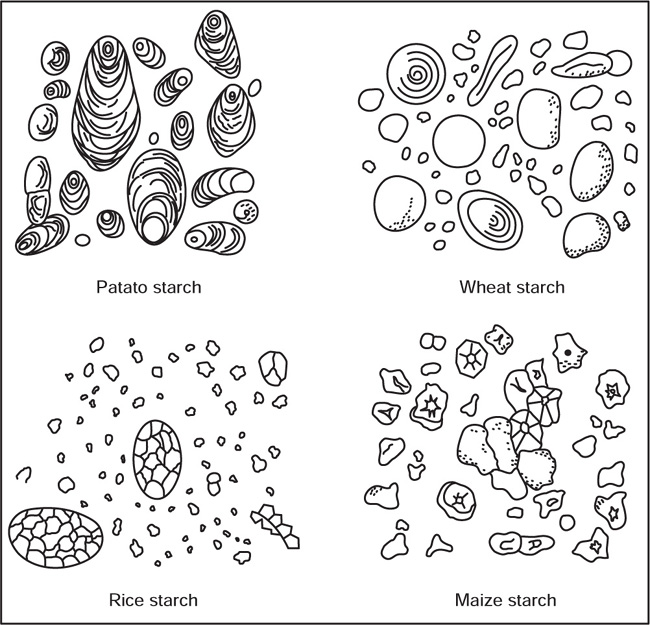
Fig. 4.14 Starch grains obtained from the different sources
Aleurone Grain
Calcium Oxalate Crystals
Calcium Carbonate
Fixed Oils and Fats
4.5 Cell Division
Mitosis
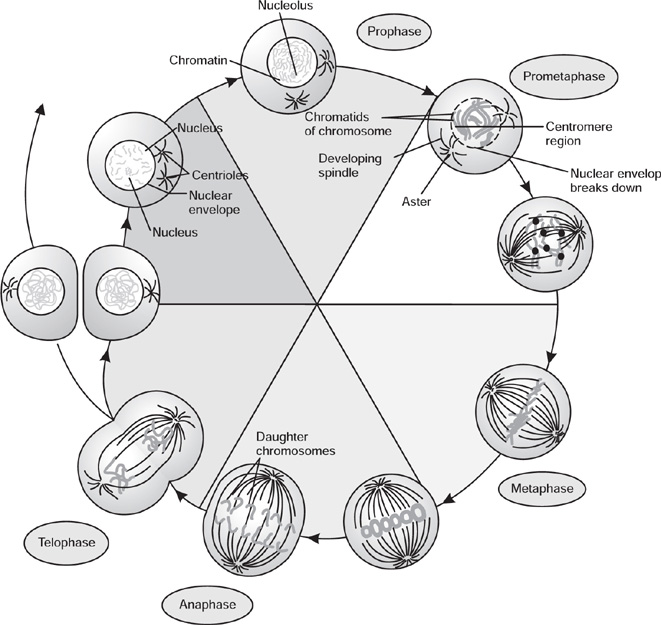
Fig. 4.15 Phases of mitotic cell division
Prophase
Metaphase
Protometaphase
Anaphase
Telophase
Cytokinesis
Meiosis
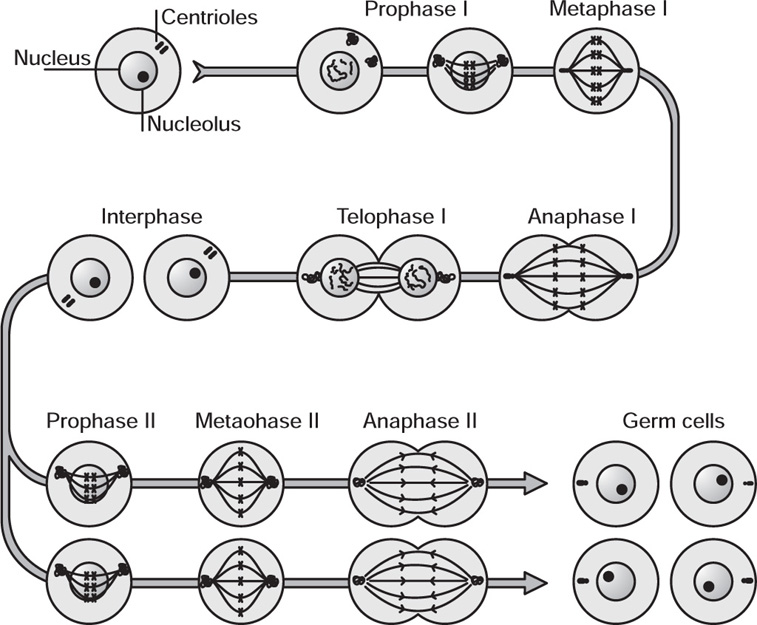
Fig. 4.16 Phases of meiosis
Division I
Division II
4.6 Morphological Study
Morphology of Bark
Methods of Collection of Barks
Morphology of Bark
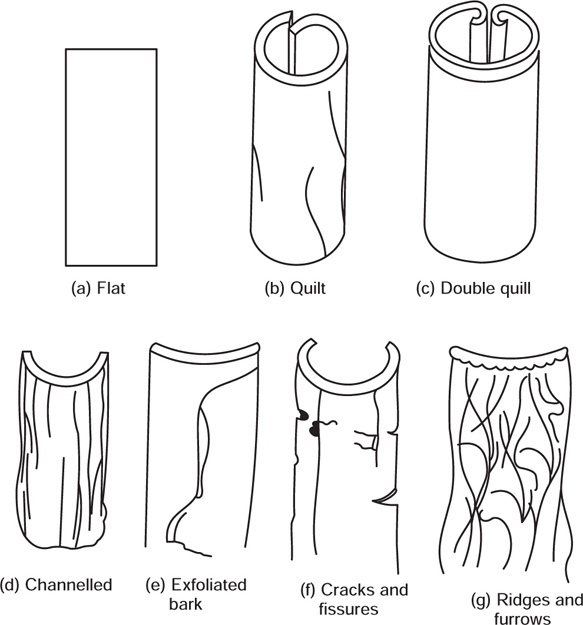
Fig. 4.17 Morphological characters of bark
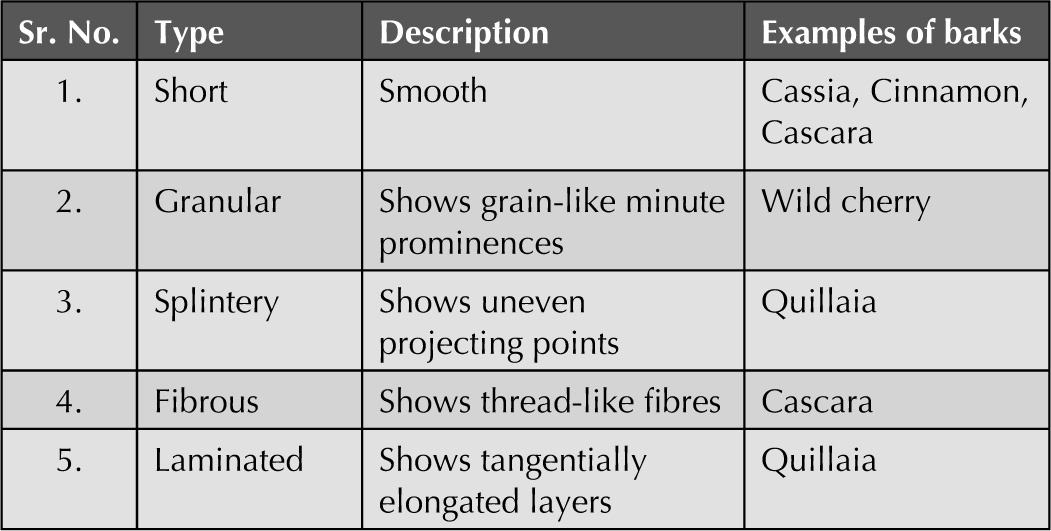
Histology of Barks
Morphology of Roots
[A] Functions of Roots
[B] Various Parts of a Root
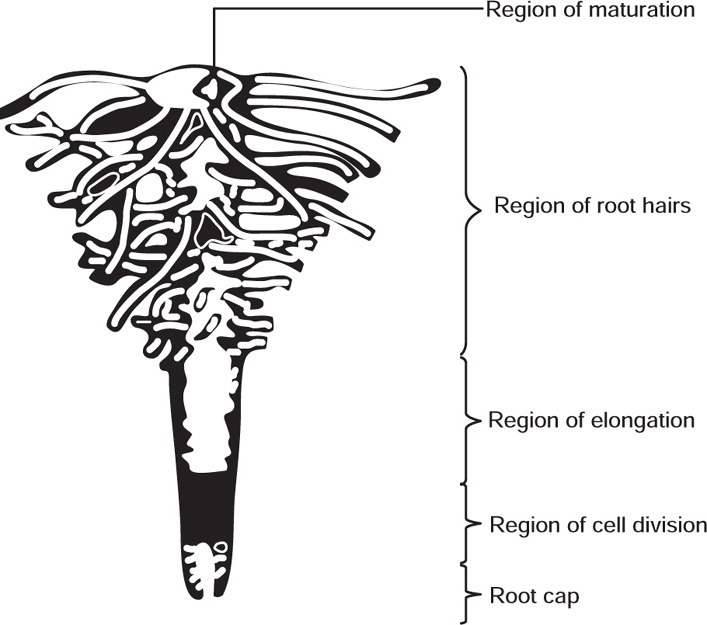
Fig. 4.18 Apex of root showing different regions
[C] Types of Roots
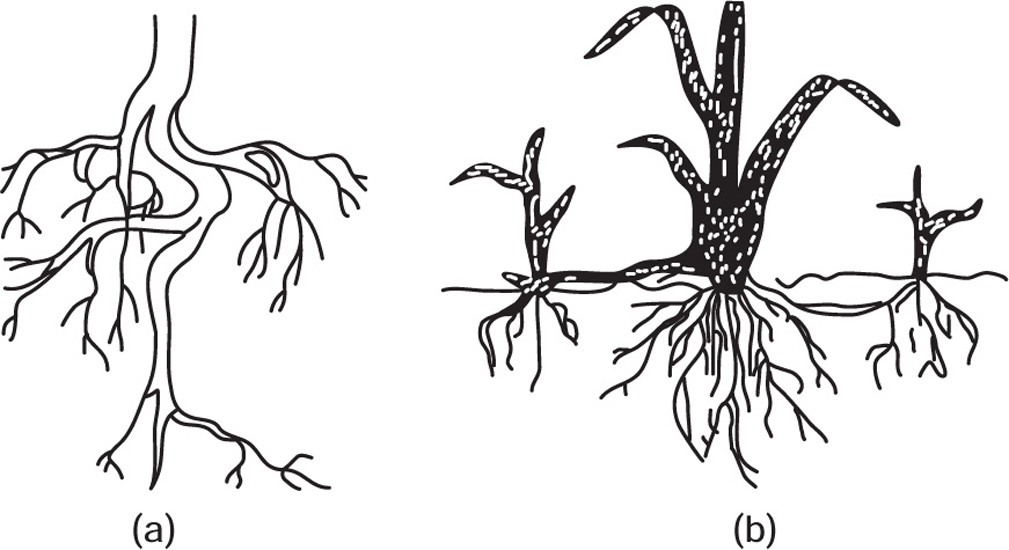
Fig. 4.19 (a) Tap root system and (b) Adventitious root system
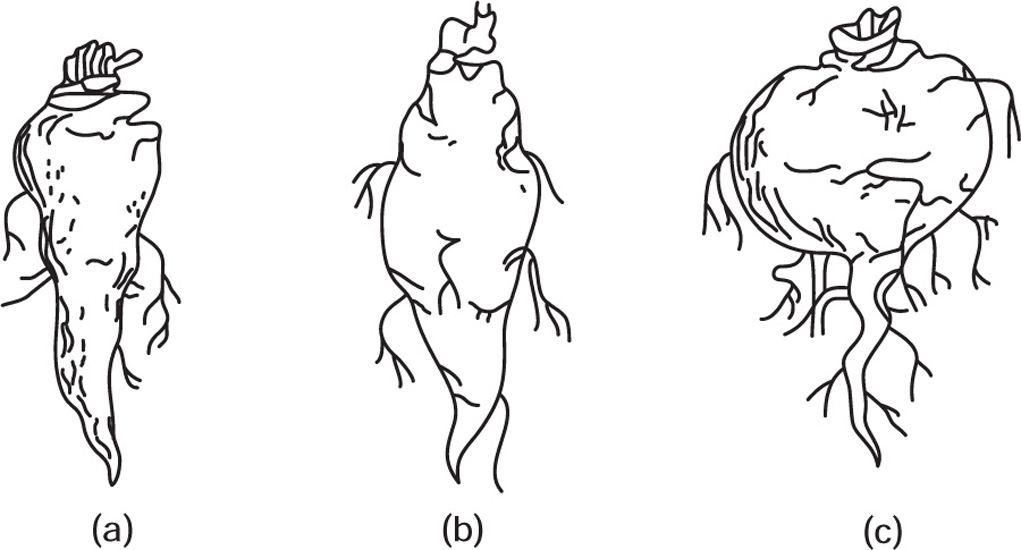
Fig. 4.20 (a) Conical root, (b) Fusiform root and (c) Napiform root
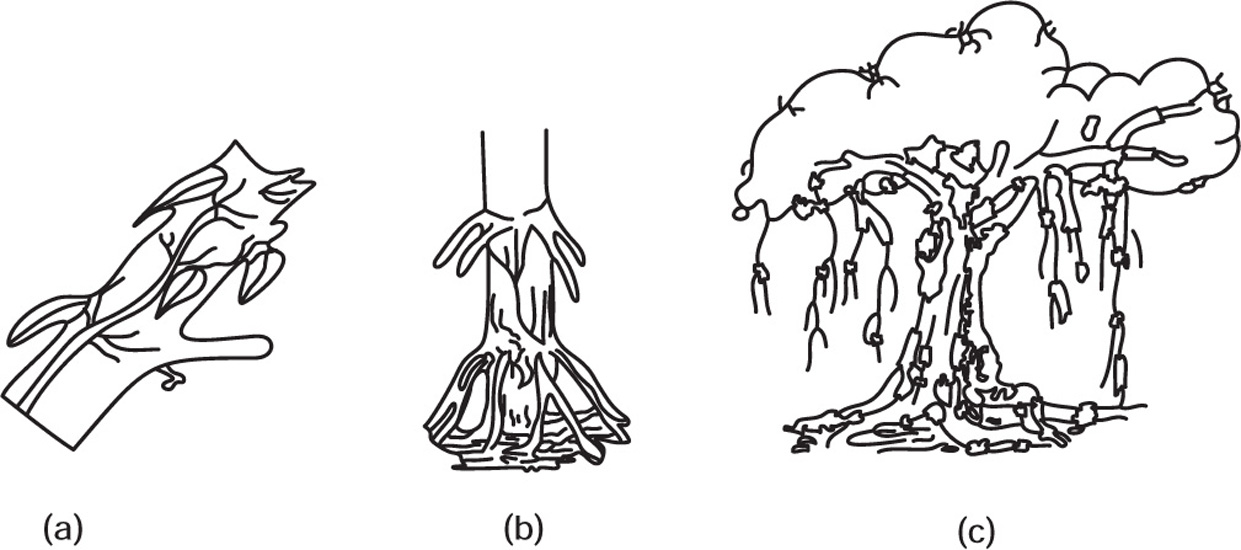
Fig. 4.21 (a) Climbing root (b) Stilt root (c) Columnar root
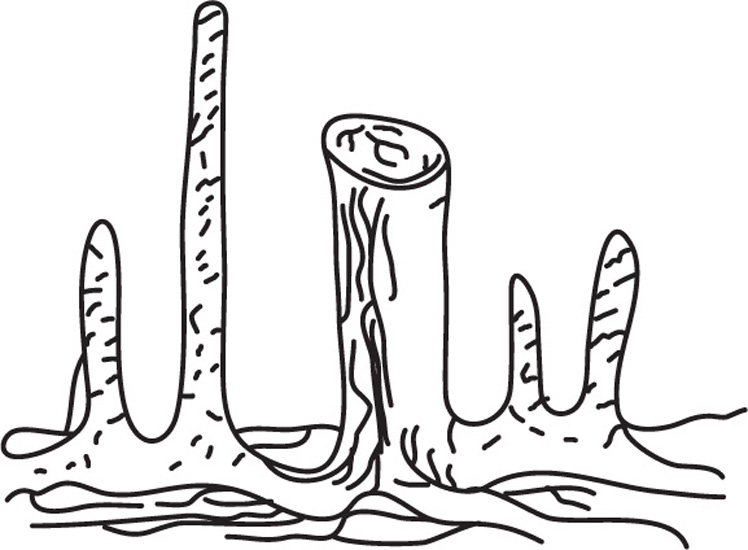
Fig. 4.22 Respiratory roots
Uses of Roots
Morphology of Stems
I Underground Modifications of Stems
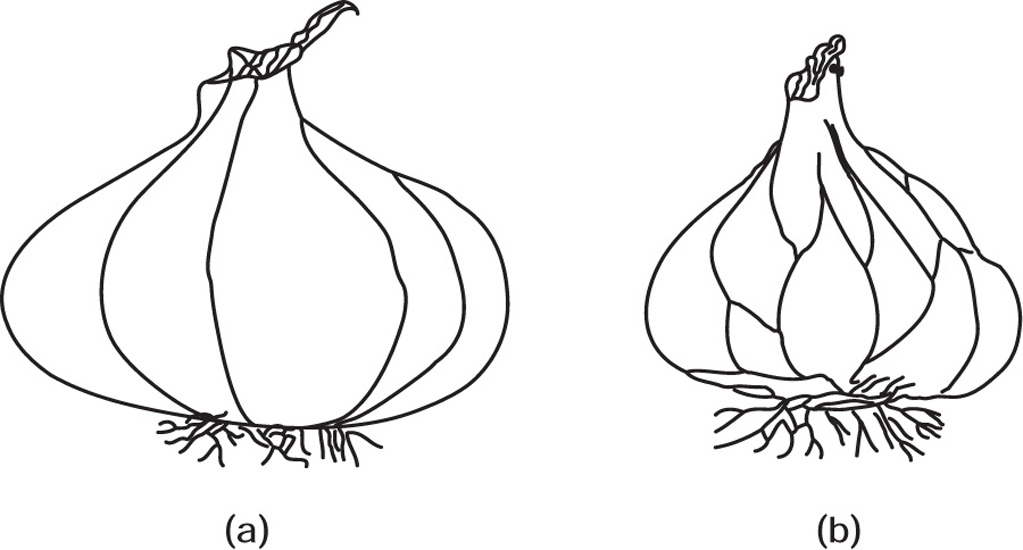
Fig. 4.23 Bulbs (a) Onion (b) Garlic
II Sub-Aerial Modifications of Stems
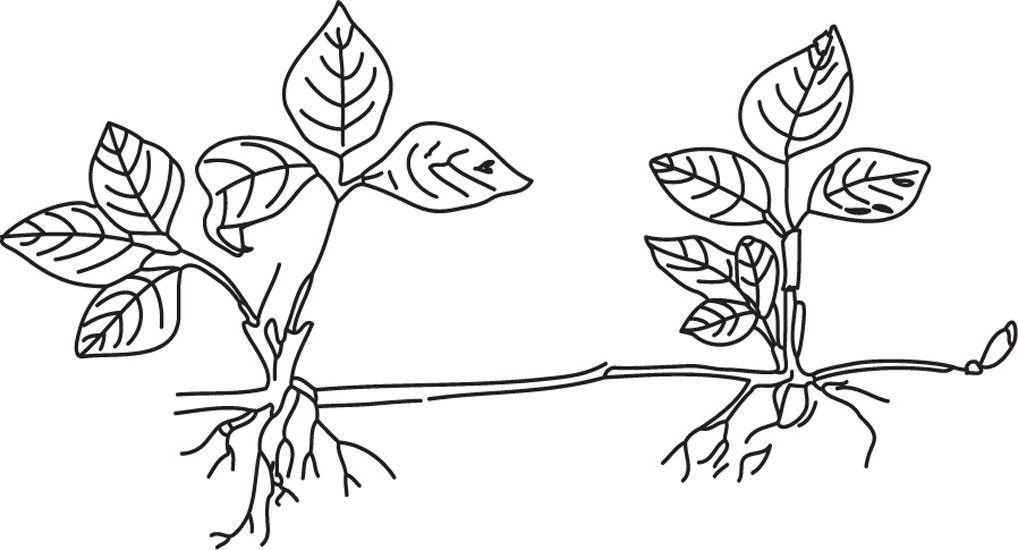
Fig. 4.24 Strawberry runner
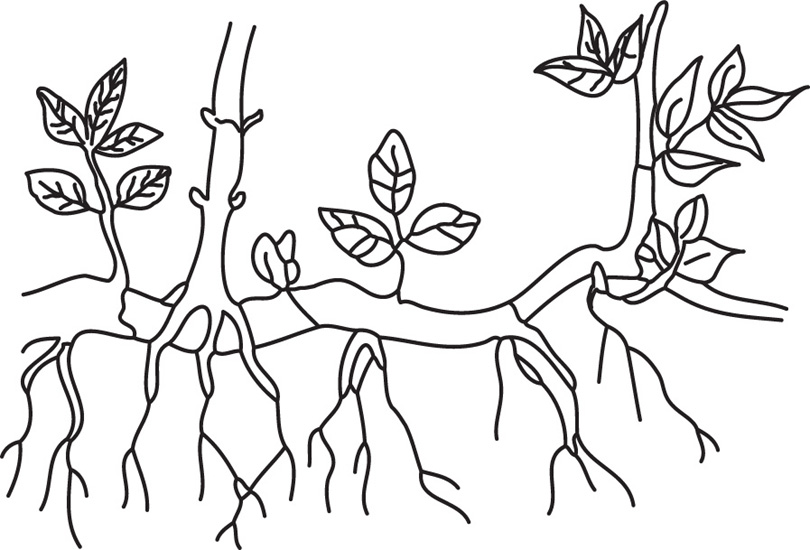
Fig. 4.25 Sucker of mentha
III Aerial Modification of Stems
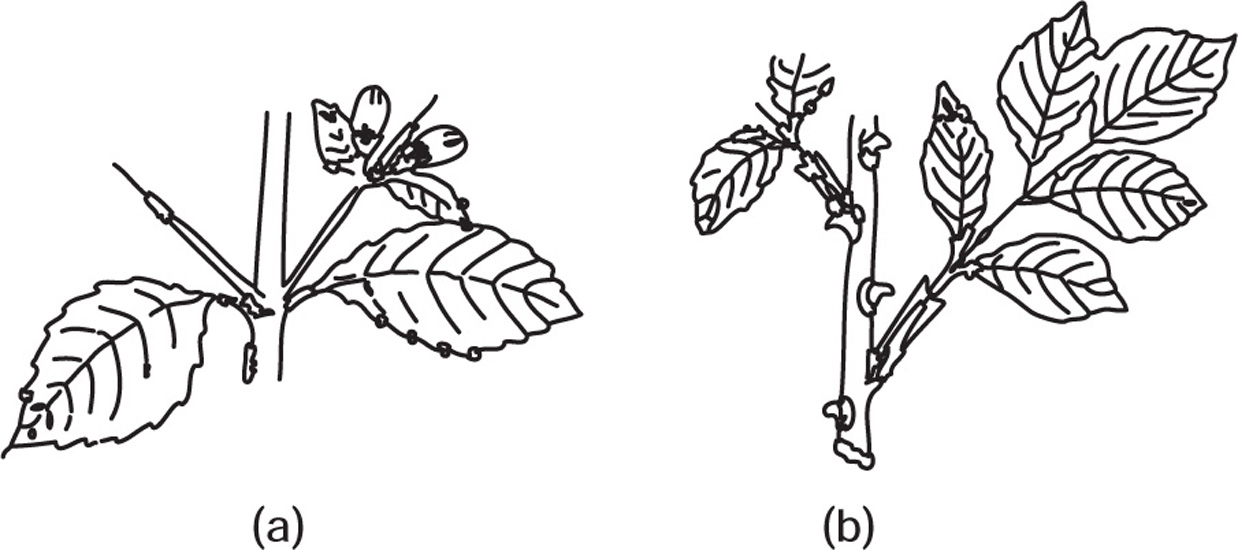
Fig. 4.26 (a) Thorns of duranta (b) Rose prickles
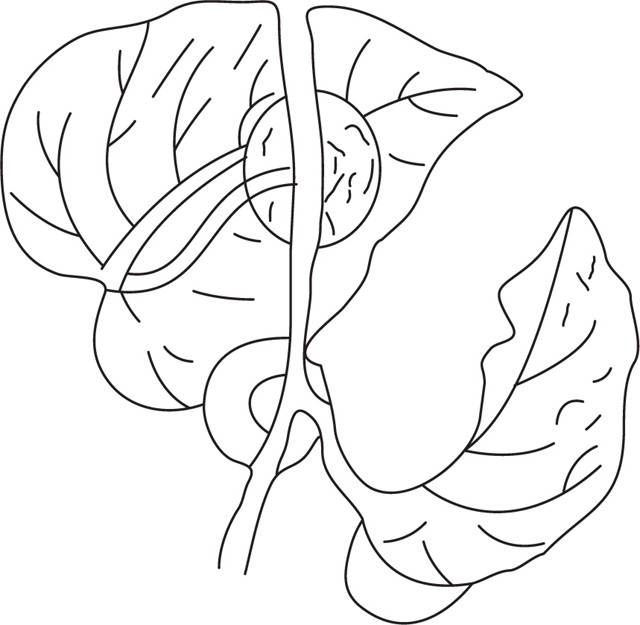
Uses of Stems
Morphology of Leaves
[A] A Typical Angiospermic Leaf Consists of the Following Parts
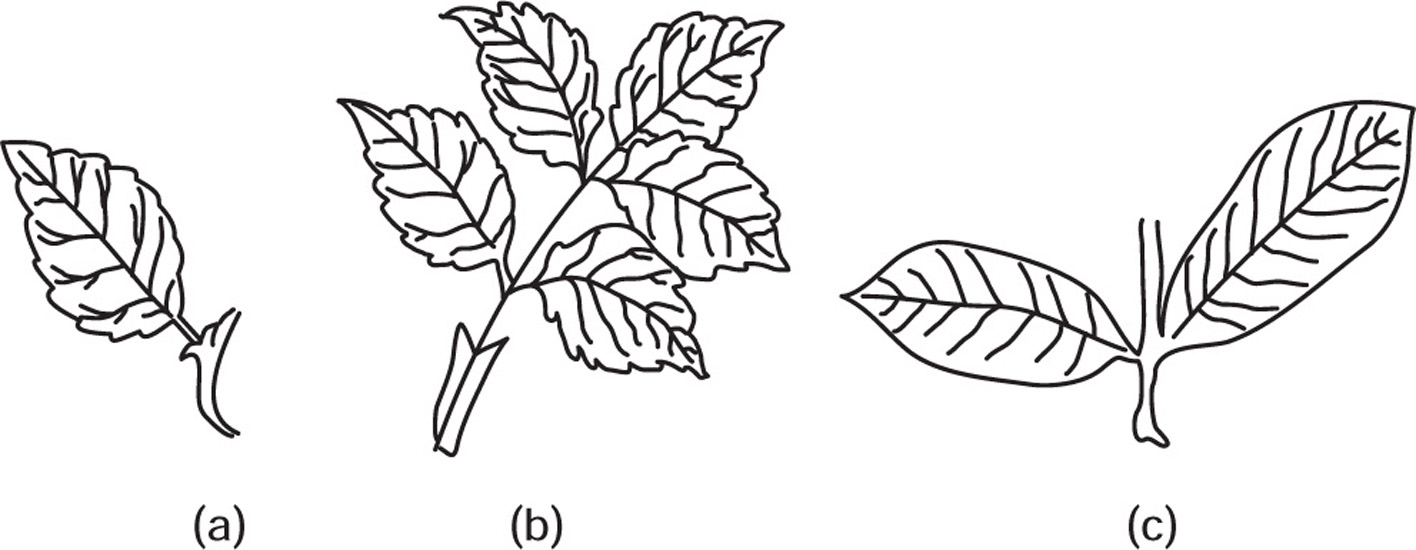
| Sr. No. | Leaf | Leaflet |
| 1. | Bud or branch is present in the axil. | Bud is absent. |
| 2. | Leaves are solitary and are arranged spirally | These are arranged in pairs |
| 3. | These lie in different planes | Leaflets lie in the same plane |
| 4. | Symmetrical at the bases, i.e. Belladonna, vasaka, eucalyptus, etc. | Asymmetrical at bases, i.e. Rose, senna, acacia, etc. |
[B] Shape of the Lamina of Leaves
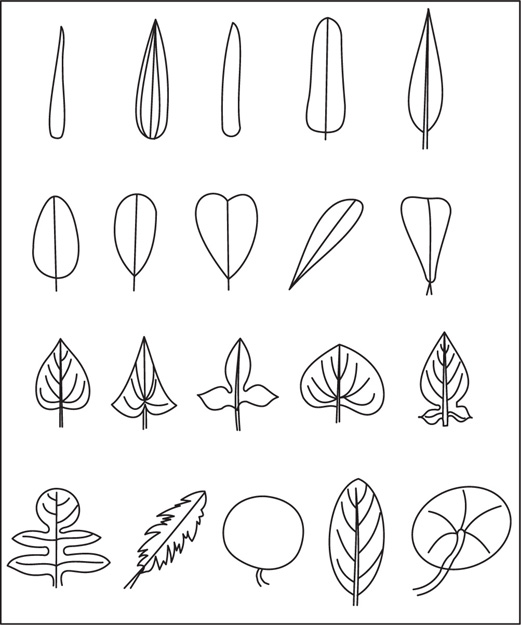
Fig. 4.29 Shape of the lamina of leaves
[C] Leaf Margins

[D] Leaf Apices
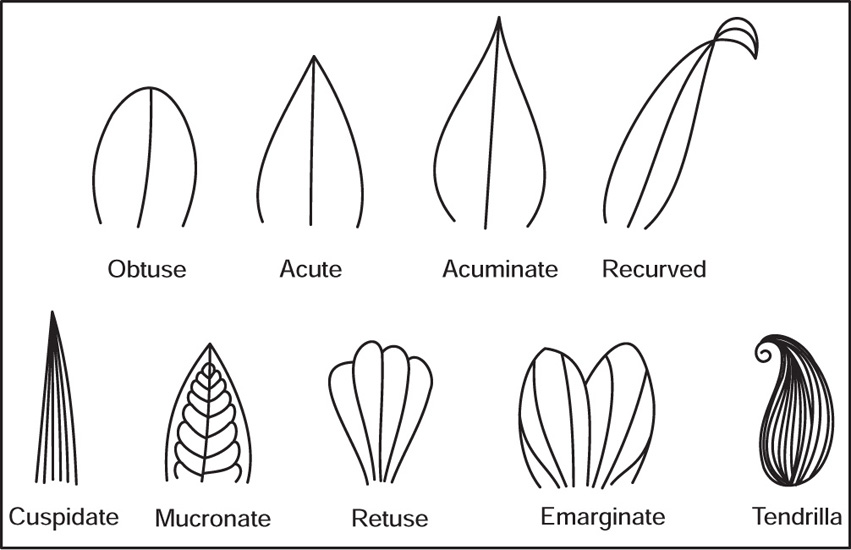
Fig. 4.31 Leaf apices
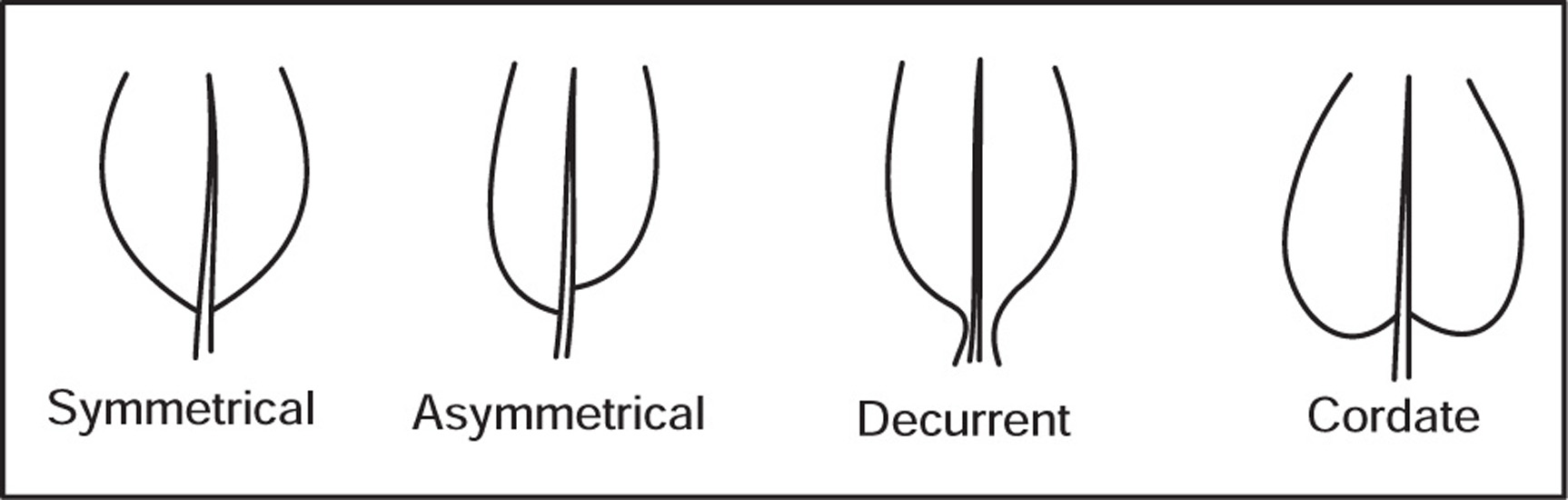
[F] Leaf Surface
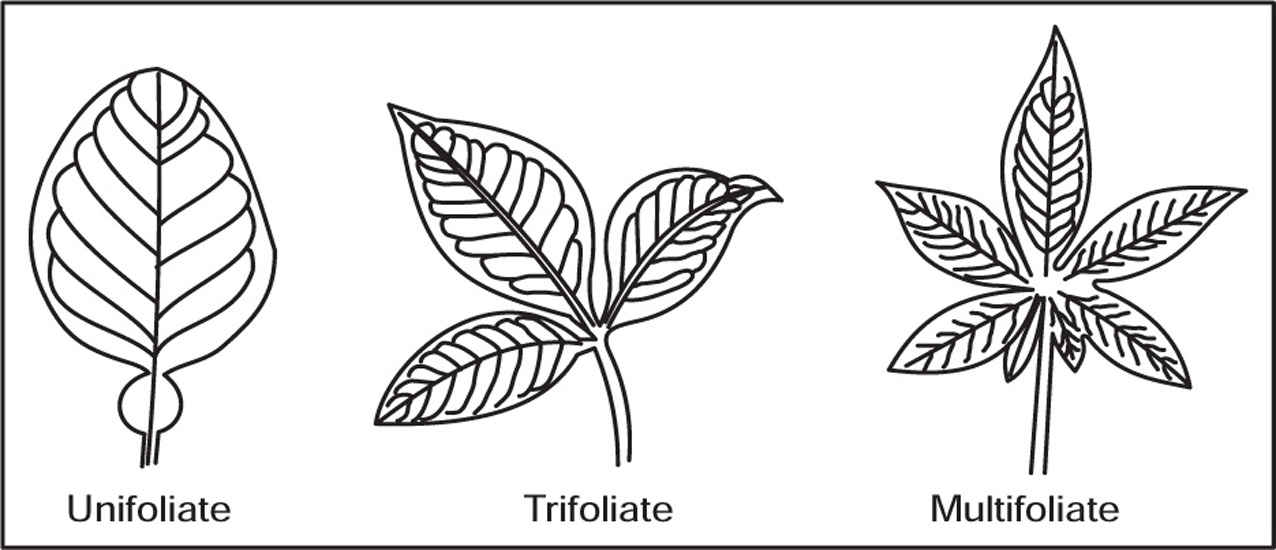
[G] Types of Leaves
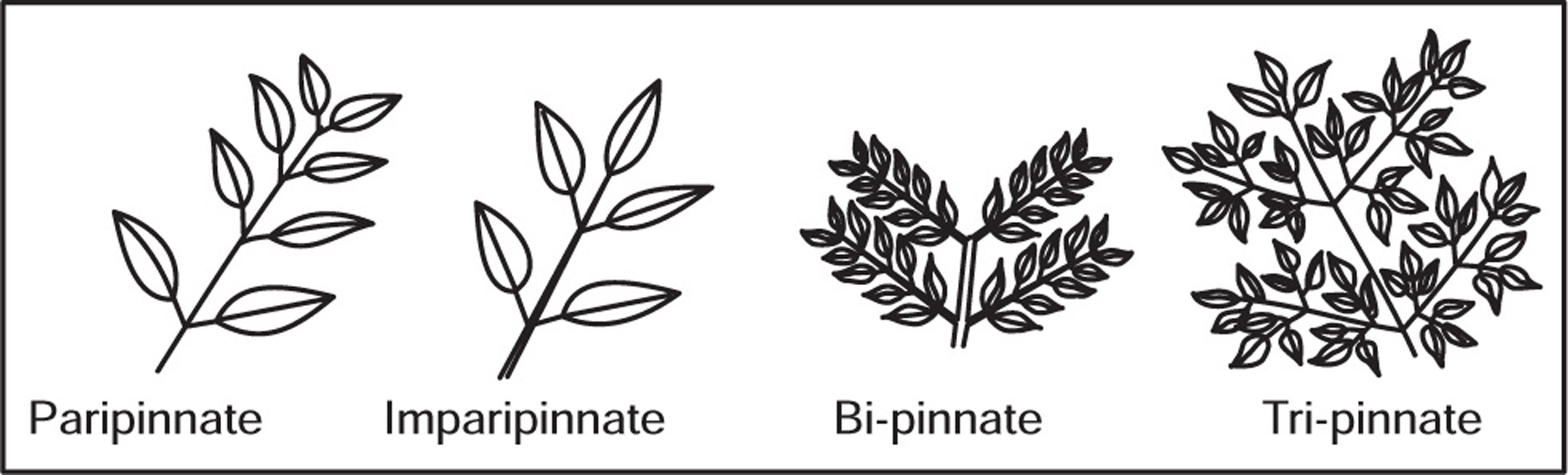
Fig. 4.33 Pinnate compound leaves
[H] Venation
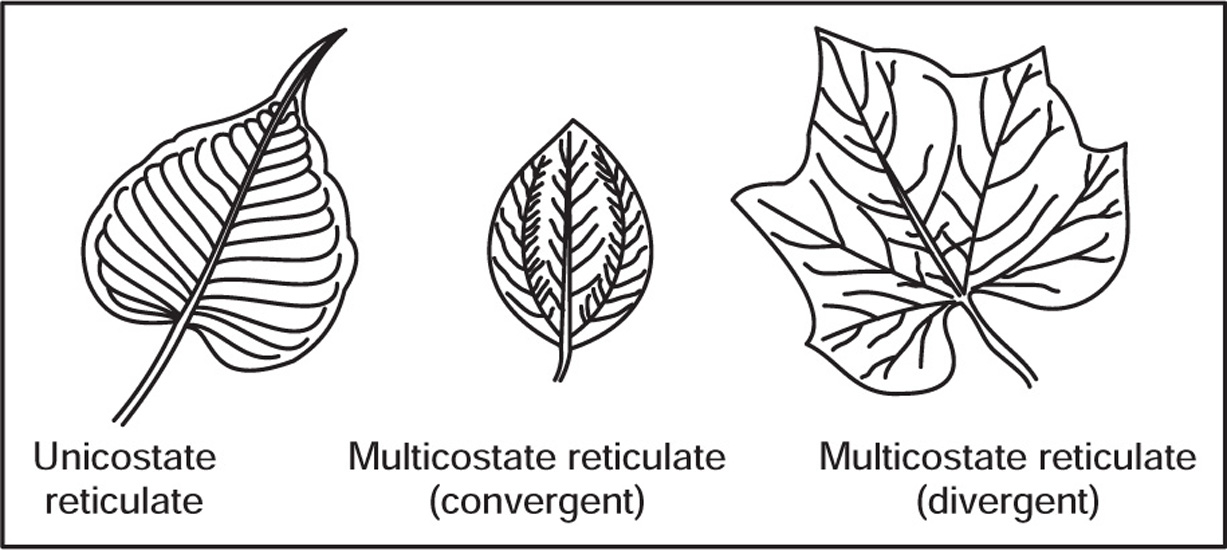
Fig. 4.35 Reticulate venation
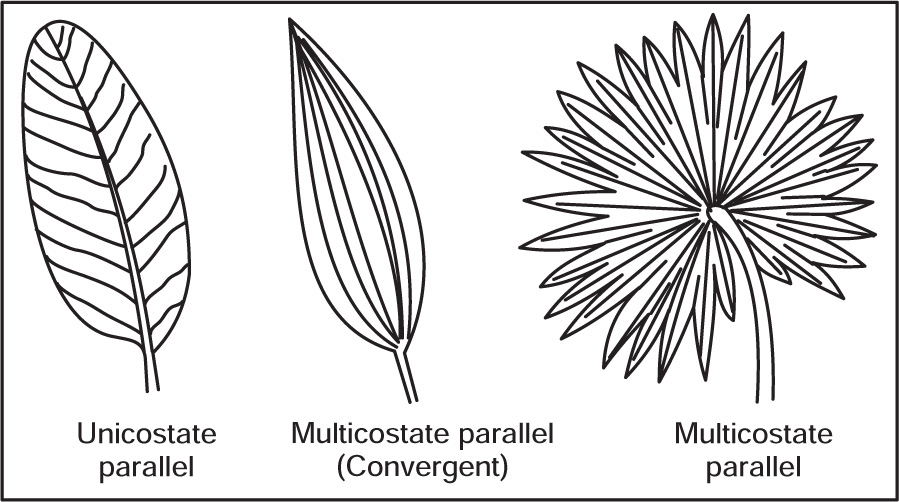
Fig. 4.36 Parallel venation
[I] Phyllotaxy
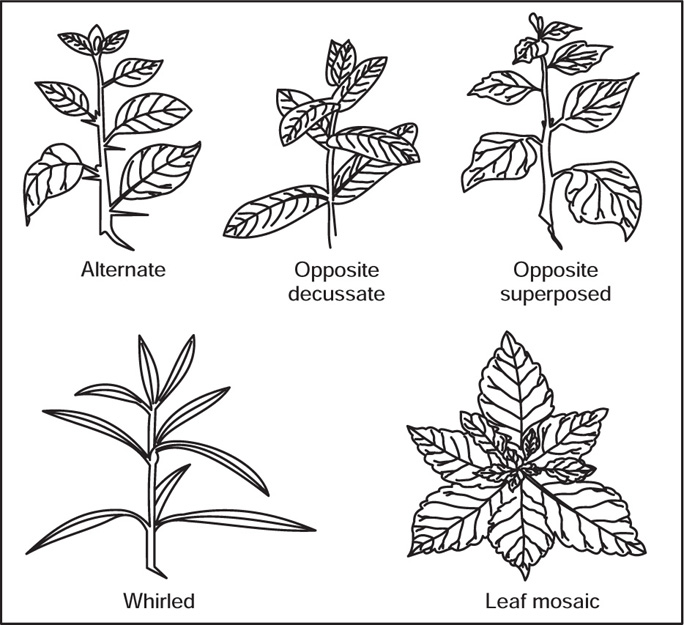
Fig. 4.37 Types of phyllotaxy
[J] Modifications of Leaves
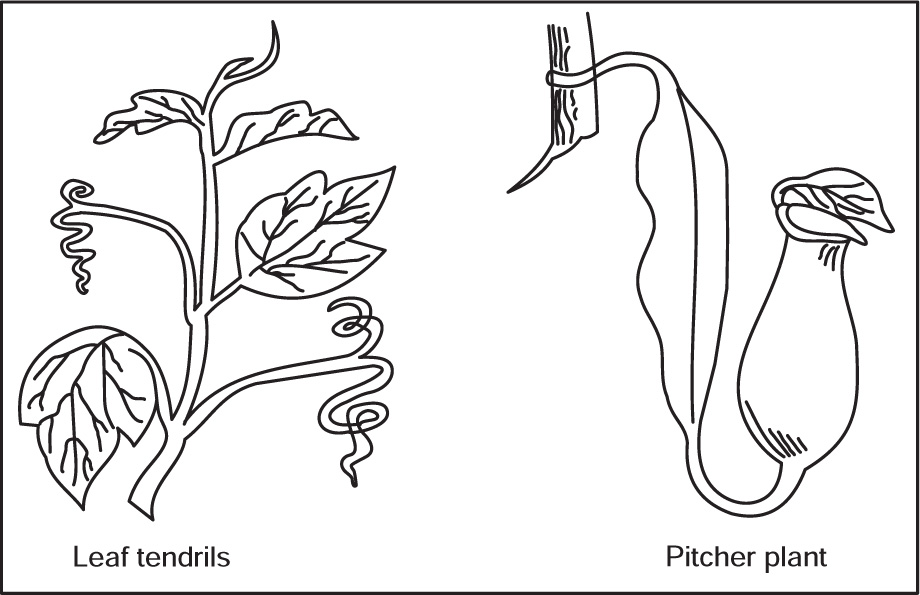
Fig. 4.38 Leaf modifications
Morphology of Flowers
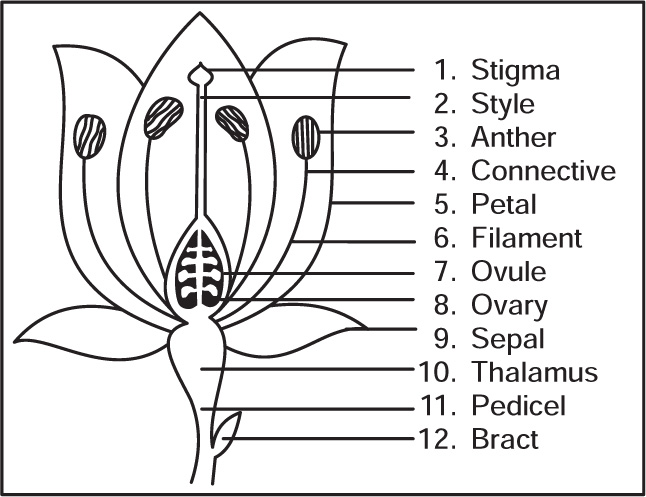
Fig. 4.39 Typical parts of flower
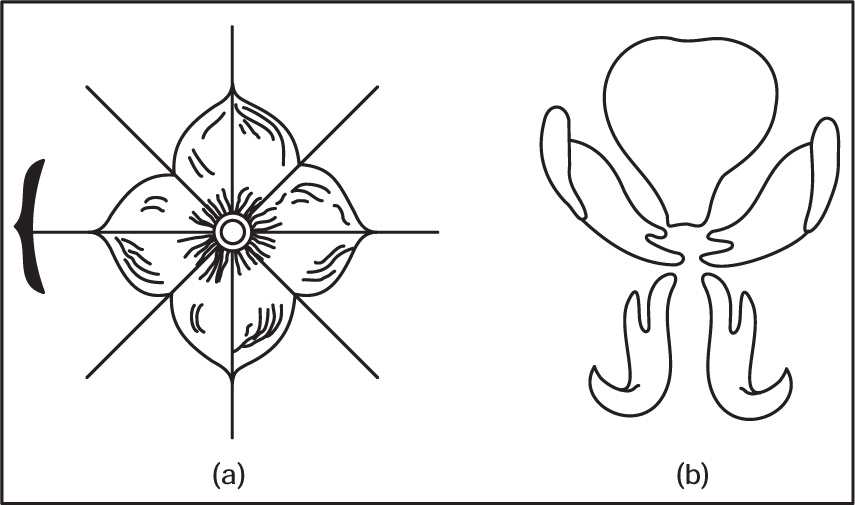
Fig. 4.40 (a) Actinomorphic flower (b) Zygomorphic flower
Arrangement of Floral Parts on Thalamus
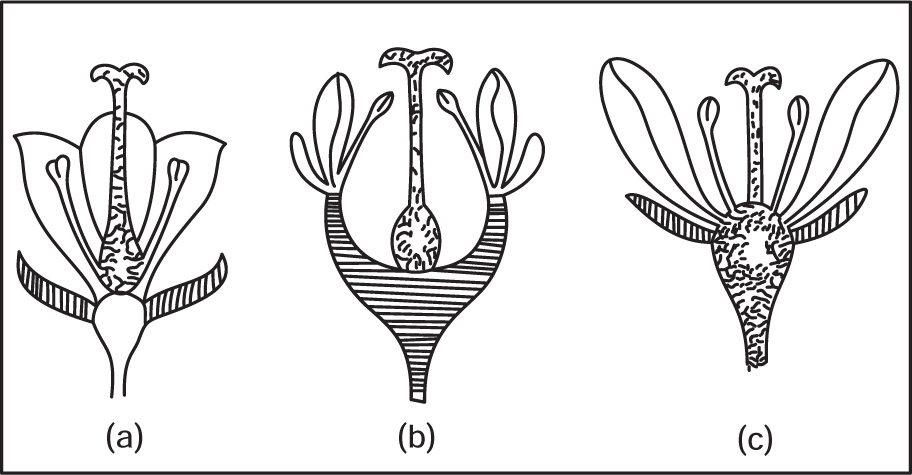
Fig. 4.41 (a) Superior ovary (b) Half-superior ovary (c) Inferior ovary
Placentation
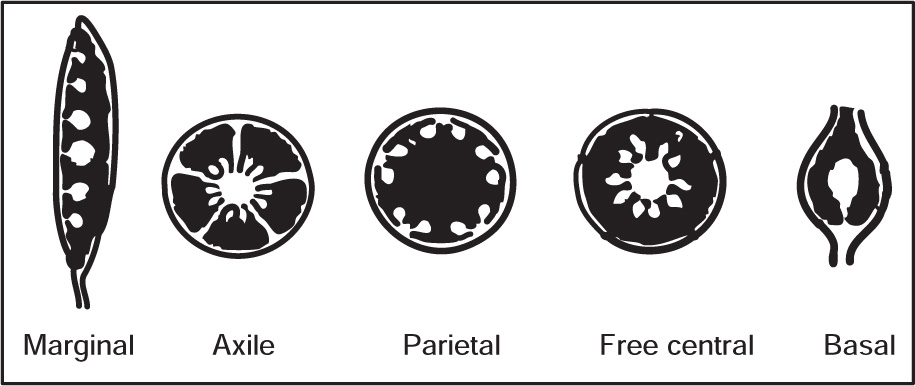
Fig. 4.42 Types of placentation
Pollination
Morphology of Inflorescence
Types of Inflorescences
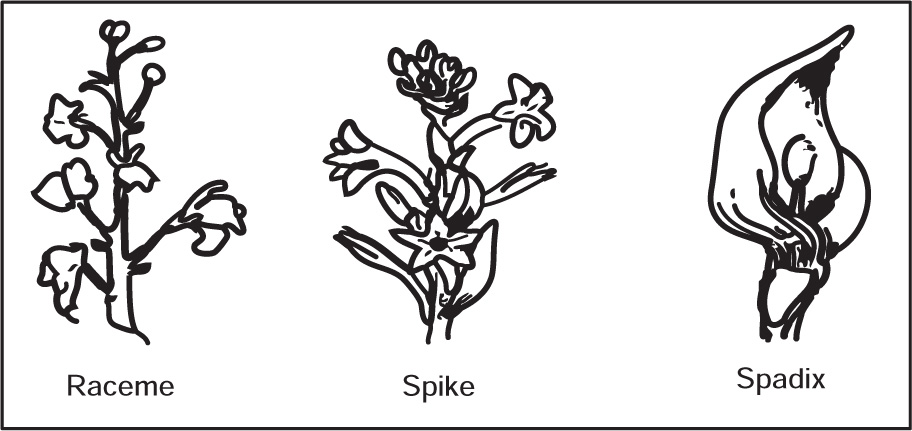
Fig. 4.43 Types of inflorescence
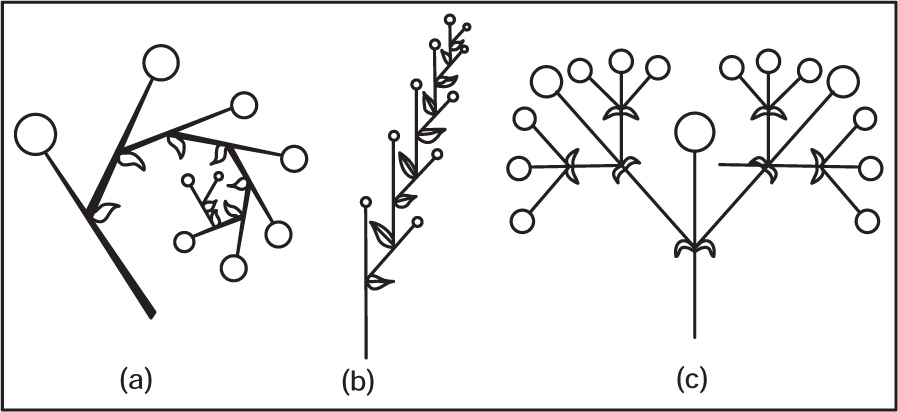
Fig. 4.44 (a) Uniparous helicoids cyme, (b) Uniparous scorpoid cyme (c) Biparous cyme
Morphology of Seeds
Seed Coat
Embryo
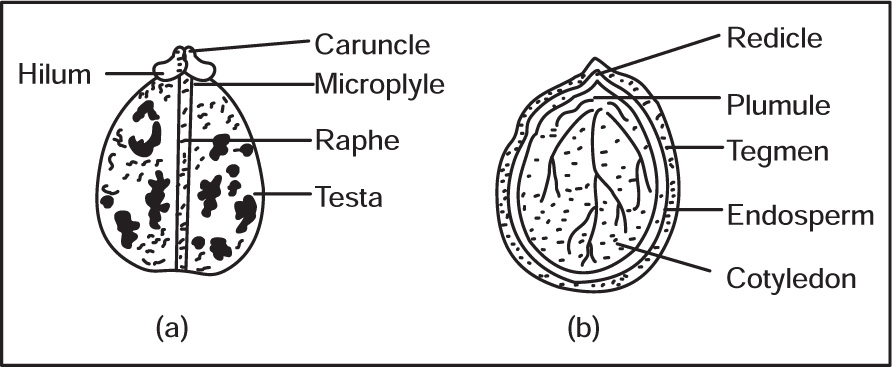
Fig. 4.45 (a) Castor seed (b) L.S. of Castor seed
Endosperm
Special Features of Seeds
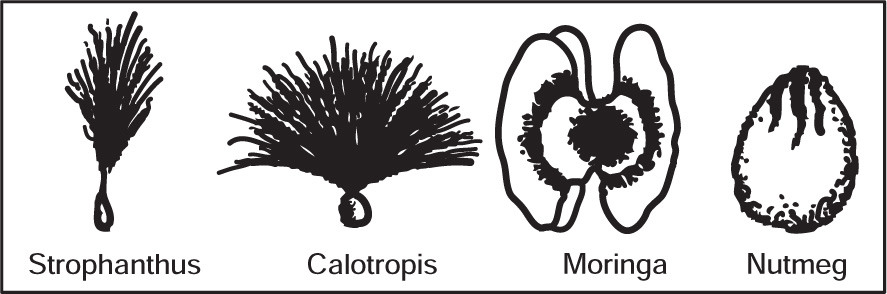
Fig. 4.46 Special features of seeds
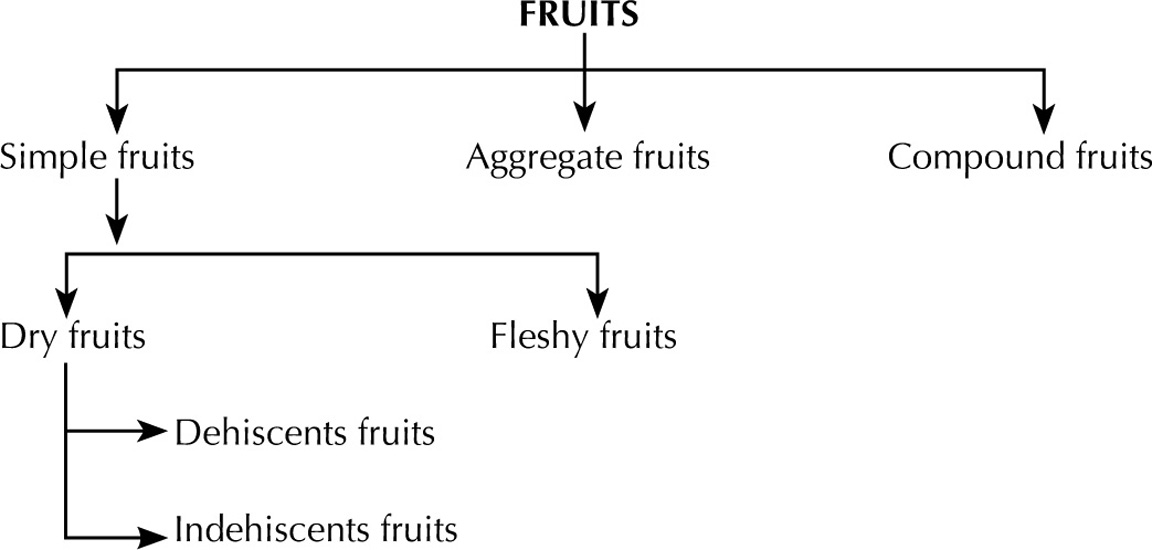
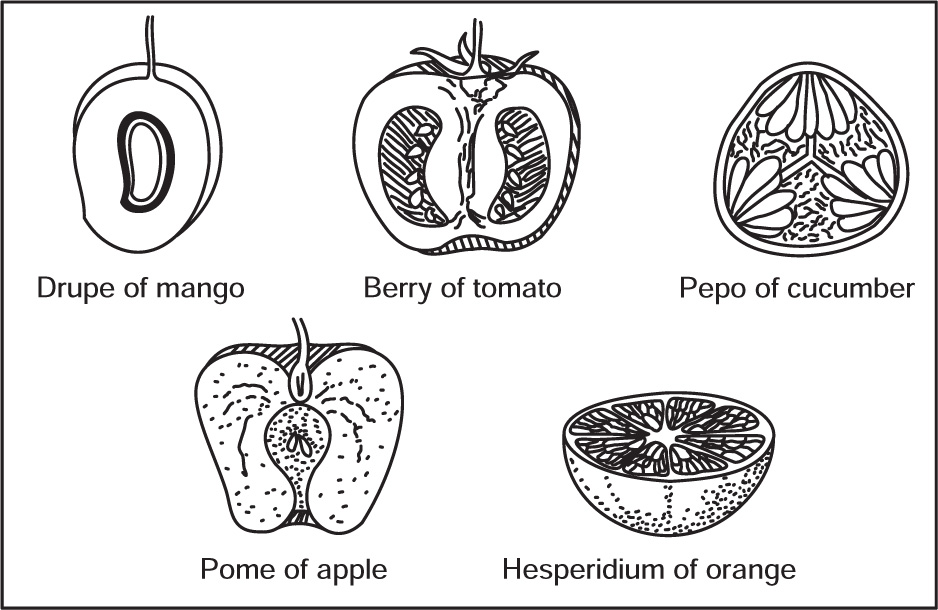
Morphology of Fruits
Simple Fruits
Compound Fruits