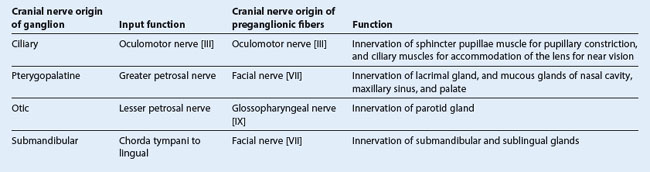
CRANIAL NERVES
The 12 pairs of cranial nerves are part of the peripheral nervous system (PNS) and pass through foramina or fissures in the cranial cavity. All nerves except one, the accessory nerve [XI], originate from the brain.
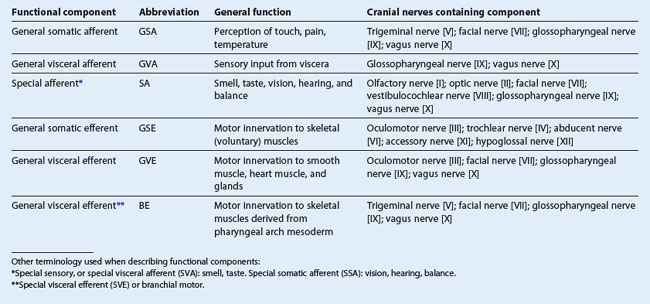
In addition to having similar somatic and visceral components as spinal nerves, some cranial nerves also contain special sensory and motor components (Tables 8.4 and 8.5).
The special sensory components are associated with hearing, seeing, smelling, balancing, and tasting.
Special motor components include those that innervate skeletal muscles derived embryologically from the pharyngeal arches and not from somites.
In human embryology, six pharyngeal arches are designated, but the fifth pharyngeal arch never develops. Each of the pharyngeal arches that does develop is associated with a developing cranial nerve or one of its branches. These cranial nerves carry efferent fibers that innervate the musculature derived from the pharyngeal arch.
Innervation of the musculature derived from the five pharyngeal arches that do develop is as follows:

first arch—trigeminal nerve [V
3];
second arch—facial nerve [VII];
third arch—glossopharyngeal nerve [IX];
fourth arch—superior laryngeal branch of the vagus nerve [X];
sixth arch—recurrent laryngeal branch of the vagus nerve [X].
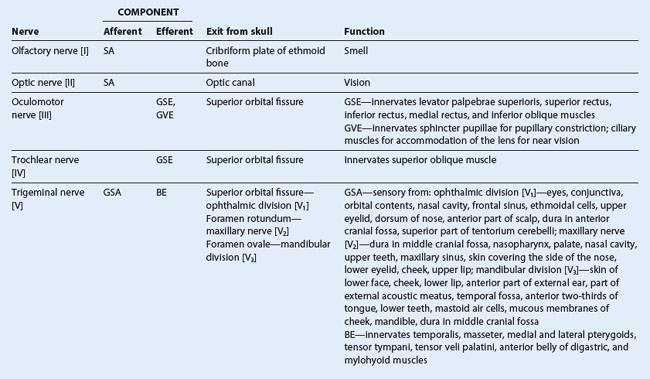
Olfactory nerve [I]
The olfactory nerve [I] carries special afferent (SA) fibers for the sense of smell. Its sensory neurons have:
peripheral processes that act as receptors in the nasal mucosa; and
central processes that return information to the brain.
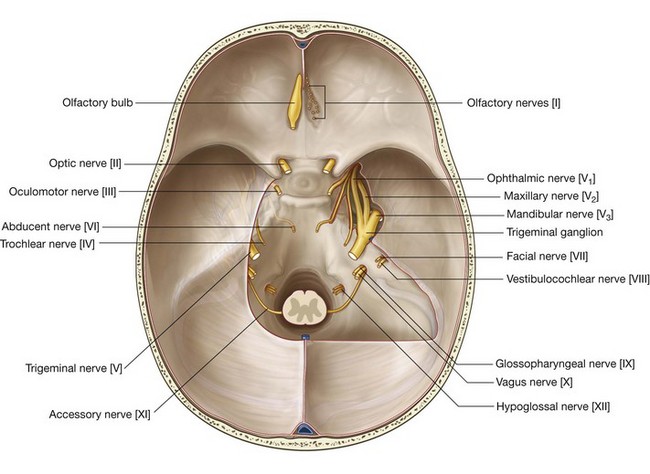
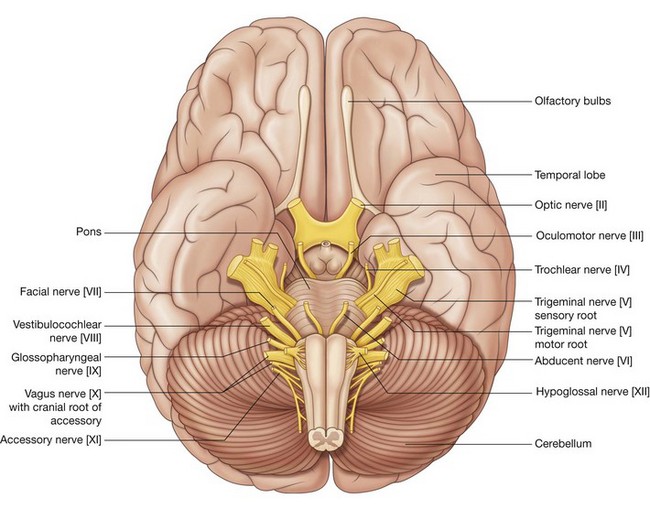
The receptors are in the roof and upper parts of the nasal cavity and the central processes, after joining into small bundles, enter the cranial cavity by passing through the cribriform plate of the ethmoid bone (Fig. 8.48). They terminate by synapsing with secondary neurons in the olfactory bulbs (Fig. 8.49).
Optic nerve [II]
The optic nerve [II] carries SA fibers for vision. These fibers return information to the brain from photoreceptors in the retina. Neuronal processes leave the retinal receptors, join into small bundles, and are carried by the optic nerves to other components of the visual system in the brain. The optic nerves enter the cranial cavity through the optic canals (Fig. 8.48).
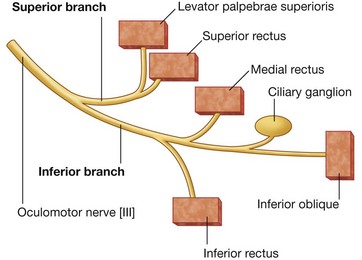
Oculomotor nerve [III]
The oculomotor nerve [III] carries two types of fibers:
general somatic efferent (GSE) fibers innervate most of the extra-ocular muscles;
general visceral efferent (GVE) fibers are part of the parasympathetic part of the autonomic division of the PNS.
The oculomotor nerve [III] leaves the anterior surface of the brainstem between the midbrain and the pons (Fig. 8.49). It enters the anterior edge of the tentorium cerebelli, continues in an anterior direction in the lateral wall of the cavernous sinus (Figs. 8.48 and 8.49), and leaves the cranial cavity through the superior orbital fissure.
In the orbit, the GSE fibers in the oculomotor nerve innervate levator palpebrae superioris, superior rectus, inferior rectus, medial rectus, and inferior oblique muscles.
The GVE fibers are preganglionic parasympathetic fibers that synapse in the ciliary ganglion and ultimately innervate the sphincter pupillae muscle, responsible for pupillary constriction, and the ciliary muscles, responsible for accommodation of the lens for near vision.
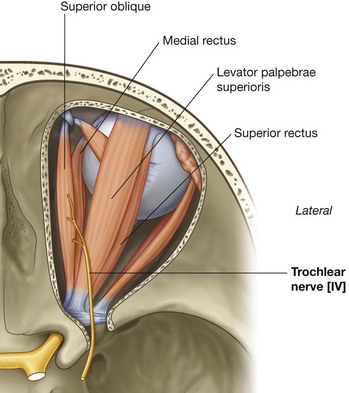
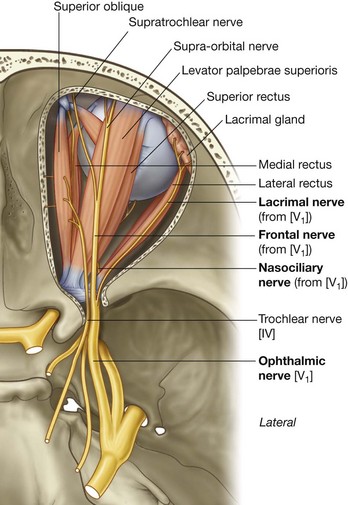
Trochlear nerve [IV]
The trochlear nerve [IV] is a cranial nerve that carries GSE fibers to innervate the superior oblique muscle, an extra-ocular muscle in the orbit. It arises in the midbrain and is the only cranial nerve to exit from the posterior surface of the brainstem (Fig. 8.49). After curving around the midbrain, it enters the inferior surface of the free edge of the tentorium cerebelli, continues in an anterior direction in the lateral wall of the cavernous sinus (Figs. 8.48 and 8.49), and enters the orbit, through the superior orbital fissure.
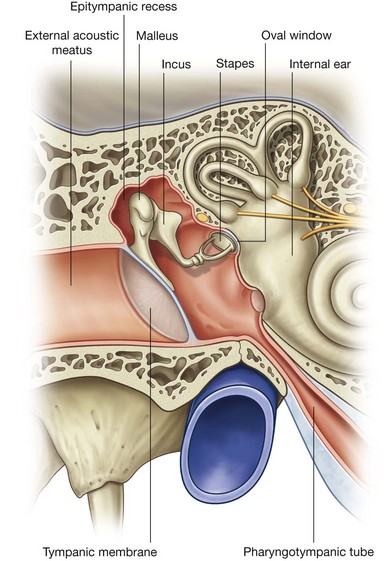
Trigeminal nerve [V]
The trigeminal nerve [V] is the major general sensory nerve of the head, and also innervates muscles that move the lower jaw. It carries general somatic afferent (GSA) and branchial efferent (BE) fibers:
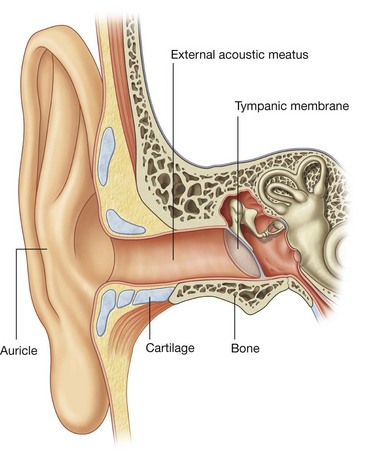
the GSA fibers provide sensory input from the face, anterior one-half of the scalp, mucous membranes of the oral and nasal cavities and the paranasal sinuses, the nasopharynx, part of the ear and external acoustic meatus, part of the tympanic membrane, orbital contents and conjunctiva, and the dura mater in the anterior and middle cranial fossae;
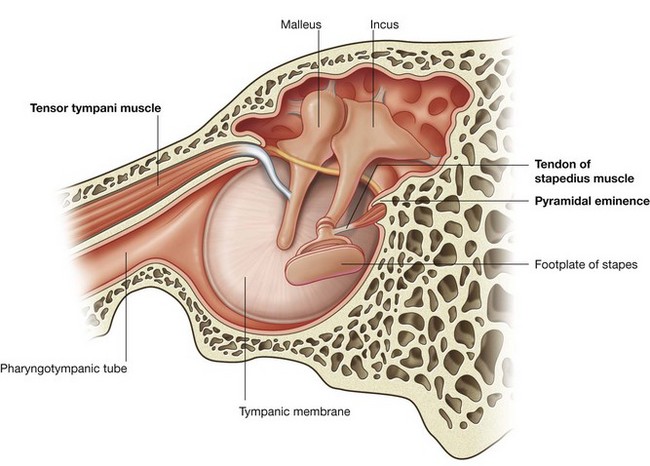
the BE fibers innervate the muscles of mastication, the tensor tympani, the tensor veli palatini, the mylohyoid, and the anterior belly of the digastric.
The trigeminal nerve exits from the anterolateral surface of the pons as a large sensory root and a small motor root (Fig. 8.49). These roots continue forward out of the posterior cranial fossa and into the middle cranial fossa by passing over the medial tip of the petrous part of the temporal bone (Fig. 8.48).
In the middle cranial fossa the sensory root expands into the trigeminal ganglion (Fig. 8.48), which contains cell bodies for the sensory neurons in the trigeminal nerve and is comparable to a spinal ganglion. The ganglion is in a depression (the trigeminal depression) on the anterior surface of the petrous part of the temporal bone, in a dural cave (the trigeminal cave). The motor root is below and completely separate from the sensory root at this point.
Arising from the anterior border of the trigeminal ganglion are the three terminal divisions of the trigeminal nerve, which in descending order are:
the
ophthalmic nerve (ophthalmic division [V1]);
the
maxillary nerve (maxillary division [V2]); and
the
mandibular nerve (mandibular division [V3]).
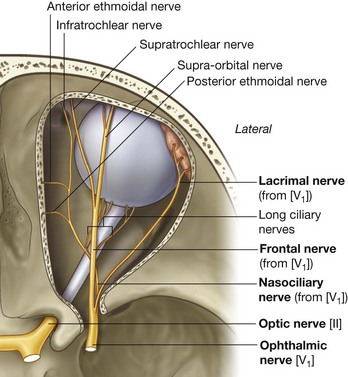
Ophthalmic nerve [V1]
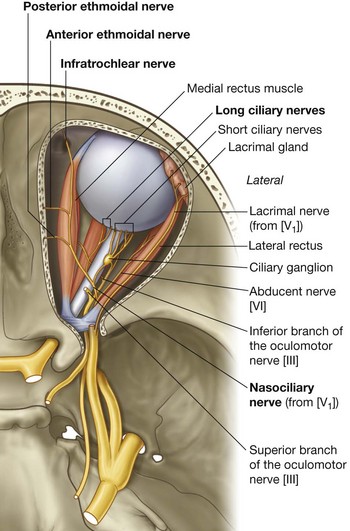
The ophthalmic nerve [V1] passes forward in the dura of the lateral wall of the cavernous sinus (see Fig. 8.44), leaves the cranial cavity, and enters the orbit through the superior orbital fissure.
The ophthalmic nerve [V1] carries sensory branches from the eyes, conjunctiva, and orbital contents, including the lacrimal gland. It also receives sensory branches from the nasal cavity, frontal sinus, ethmoidal cells, falx cerebri, dura in the anterior cranial fossa and superior parts of the tentorium cerebelli, upper eyelid, dorsum of the nose, and the anterior part of the scalp.
Maxillary nerve [V2]
The maxillary nerve [V2] passes forward in the dura mater of the lateral wall of the cavernous sinus just inferior to the ophthalmic nerve [V1] (see Fig. 8.44), leaves the cranial cavity through the foramen rotundum, and enters the pteryatine fossa.
The maxillary nerve [V2] receives sensory branches from the dura in the middle cranial fossa, the nasopharynx, the palate, the nasal cavity, teeth of the upper jaw, maxillary sinus, and skin covering the side of the nose, the lower eyelid, the cheek, and the upper lip.
Mandibular nerve [V3]
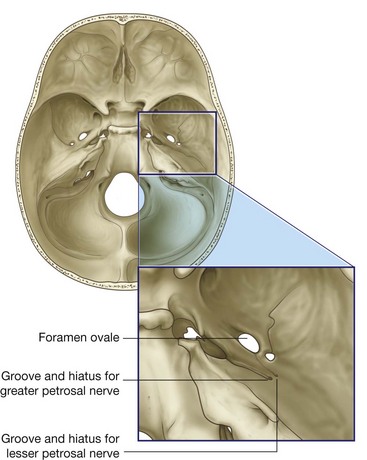
The mandibular nerve [V3] leaves the inferior margin of the trigeminal ganglion and leaves the skull through the foramen ovale.
The motor root of the trigeminal nerve also passes through the foramen ovale and unites with the sensory component of the mandibular nerve [V3] outside the skull. Thus the mandibular nerve [V3] is the only division of the trigeminal nerve that contains a motor component.
Outside the skull the motor fibers innervate the four muscles of mastication (temporalis, masseter, and medial and lateral pterygoids), as well as the tensor tympani, the tensor veli palatini, the anterior belly of the digastric, and the mylohyoid muscles.
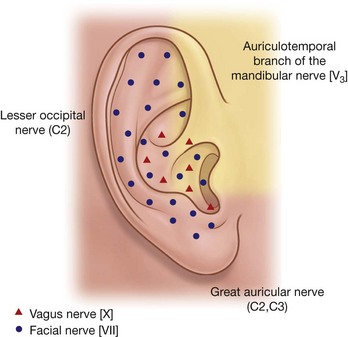
The mandibular nerve [V3] also receives sensory branches from the skin of the lower face, cheek, lower lip, anterior part of the external ear, part of the external acoustic meatus and the temporal region, the anterior two-thirds of the tongue, the teeth of the lower jaw, the mastoid air cells, the mucous membranes of the cheek, the mandible, and dura in the middle cranial fossa.
Abducent nerve [VI]
The abducent nerve [VI] carries GSE fibers to innervate the lateral rectus muscle in the orbit. It arises from the brainstem between the pons and medulla and passes forward, piercing the dura covering the clivus (Figs. 8.48 and 8.49). Continuing upward in a dural canal, it crosses the superior edge of the petrous temporal bone, enters and crosses the cavernous sinus (see Fig. 8.44) just inferolateral to the internal carotid artery, and enters the orbit through the superior orbital fissure.
Facial nerve [VII]
The facial nerve [VII] carries GSA, SA, GVE, and BE fibers:
the GSA fibers provide sensory input from part of the external acoustic meatus and deeper parts of the auricle;
the SA fibers are for taste from the anterior two-thirds of the tongue;
the GVE fibers are part of the parasympathetic part of the autonomic division of the PNS and stimulate secretomotor activity in the lacrimal gland, submandibular and sublingual salivary glands, and glands in the mucous membranes of the nasal cavity, and hard and soft palates;
the BE fibers innervate the muscles of the face (muscles of facial expression) and scalp derived from the second pharyngeal arch, and the stapedius, the posterior belly of the digastric, and the stylohyoid muscles.
The facial nerve [VII] attaches to the lateral surface of the brainstem, between the pons and medulla oblongata (Fig. 8.49). It consists of a large motor root and a smaller sensory root (the intermediate nerve):
the intermediate nerve contains the SA fibers for taste, the parasympathetic GVE fibers and the GSA fibers;
the larger motor root contains the BE fibers.
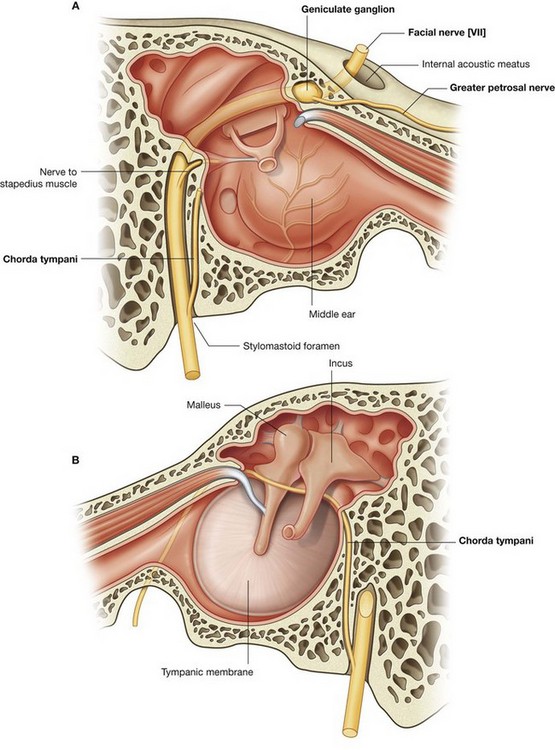
The motor and sensory roots cross the posterior cranial fossa and leave the cranial cavity through the internal acoustic meatus (Fig. 8.48). After entering the facial canal in the petrous part of the temporal bone, the two roots fuse and form the facial nerve [VII]. Near this point the nerve enlarges as the geniculate ganglion, which is similar to a spinal ganglion containing cell bodies for sensory neurons.
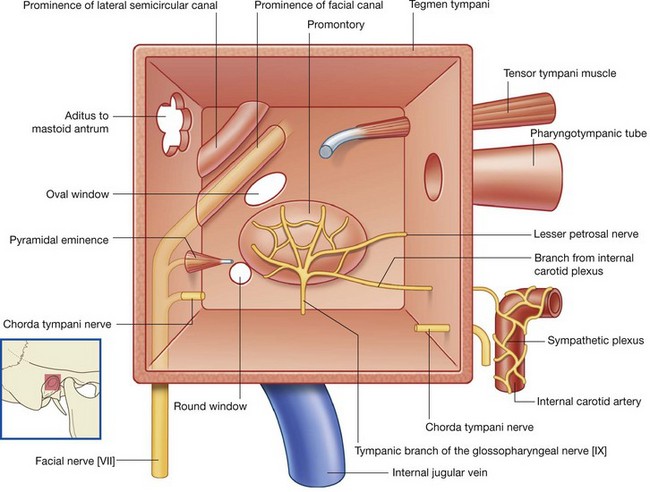
At the geniculate ganglion the facial nerve [VII] turns and gives off the greater petrosal nerve, which carries mainly preganglionic parasympathetic (GVE) fibers (Table 8.6).
The facial nerve [VII] continues along the bony canal, giving off the nerve to stapedius and the chorda tympani, before exiting the skull through the stylomastoid foramen.
The chorda tympani carries taste (SA) fibers from the anterior two-thirds of the tongue and preganglionic parasympathetic (GVE) fibers destined for the submandibular ganglion (Table 8.6).
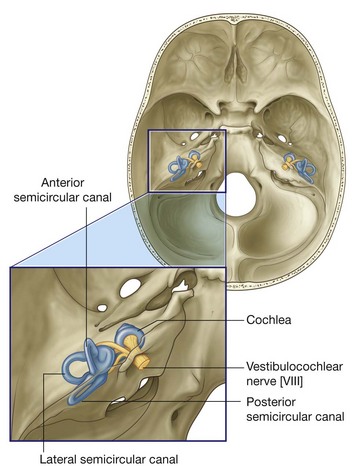
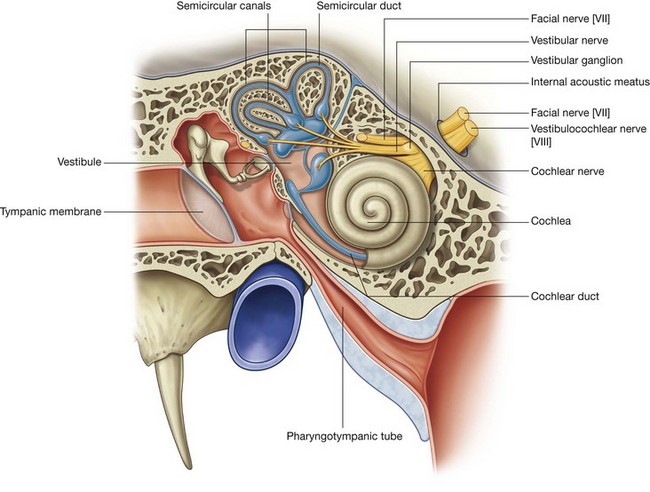
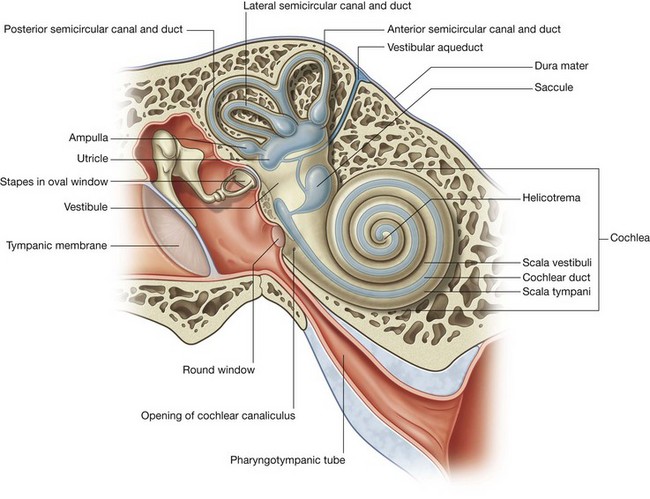
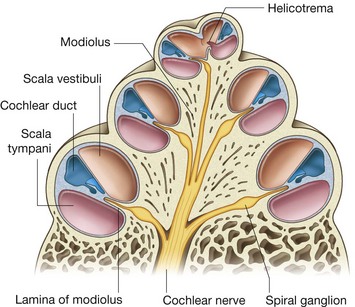
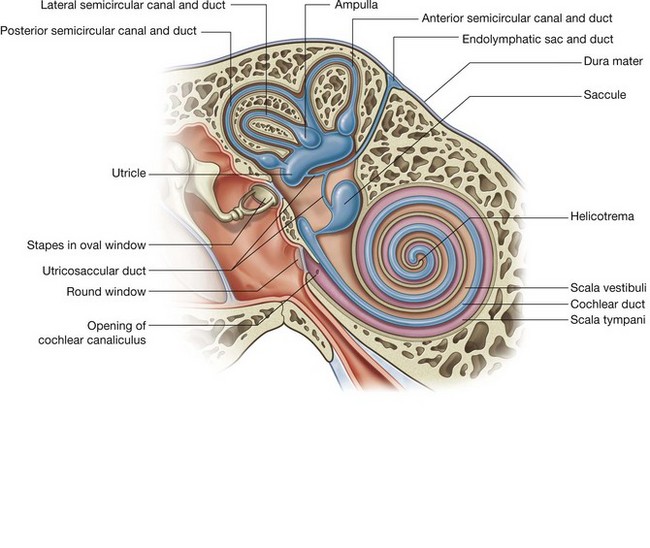
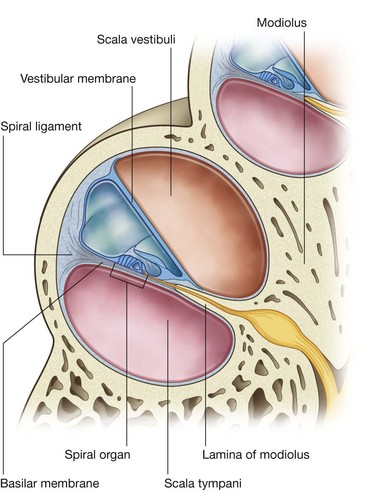
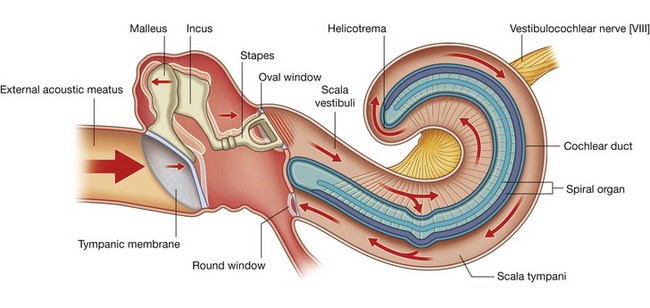
Vestibulocochlear nerve [VIII]
The vestibulocochlear nerve [VIII] carries SA fibers for hearing and balance, and consists of two divisions:
a vestibular component for balance;
a cochlear component for hearing.
The vestibulocochlear nerve [VIII] attaches to the lateral surface of the brainstem, between the pons and medulla, after emerging from the internal acoustic meatus and crossing the posterior cranial fossa (Figs. 8.48 and 8.49). The two divisions combine into the single nerve seen in the posterior cranial fossa within the substance of the petrous part of the temporal bone.
Glossopharyngeal nerve [IX]
The glossopharyngeal nerve [IX] carries GVA, SA, GVE, and BE fibers:
the GVA fibers provide sensory input from the carotid body and sinus;
the GSA fibers provide sensory input from posterior one-third of the tongue, palatine tonsils, oropharynx, and mucosa of the middle ear and pharyngotympanic tube;
the SA fibers are for taste from the posterior one-third of the tongue;
the GVE fibers are part of the parasympathetic part of the autonomic division of the PNS and stimulate secretomotor activity in the parotid salivary gland;
the BE fibers innervate the muscle derived from the third pharyngeal arch (the stylopharyngeus muscle).
The glossopharyngeal nerve [IX] arises as several rootlets on the anterolateral surface of the upper medulla oblongata (Fig. 8.49). The rootlets cross the posterior cranial fossa and enter the jugular foramen (Fig. 8.48). Within the jugular foramen, and before exiting from it, the rootlets merge to form the glossopharyngeal nerve.
Within or immediately outside the jugular foramen are two ganglia (the superior and inferior ganglia), which contain the cell bodies of the sensory neurons in the glossopharyngeal nerve [IX].
Tympanic nerve
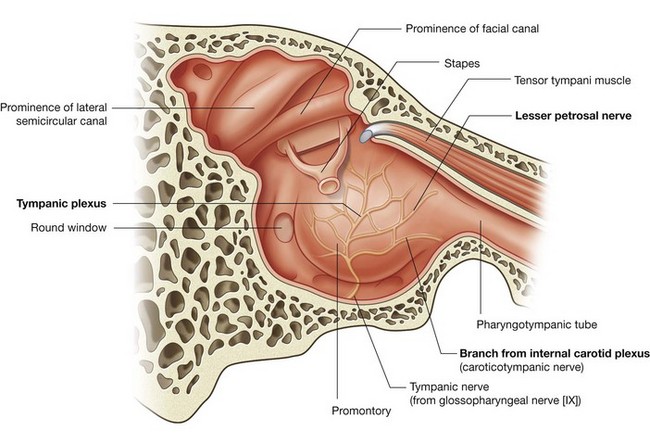
Branching from the glossopharyngeal nerve [IX] either within or immediately outside the jugular foramen is the tympanic nerve. This branch re-enters the temporal bone, enters the middle ear cavity, and participates in the formation of the tympanic plexus. Within the middle ear cavity it provides sensory innervation to the mucosa of the cavity, pharyngotympanic tube, and mastoid air cells.
The tympanic nerve also contributes GVE fibers, which leave the tympanic plexus in the lesser petrosal nerve—a small nerve that exits the temporal bone, enters the middle cranial fossa, and descends through the foramen ovale to exit the cranial cavity carrying preganglionic parasympathetic fibers to the otic ganglion (Table 8.6).
Vagus nerve [X]
The vagus nerve [X] carries GSA, GVA, SA, GVE, and BE fibers:
the GSA fibers provide sensory input from the larynx, laryngopharynx, deeper parts of the auricle, part of the external acoustic meatus, and the dura mater in the posterior cranial fossa;
the GVA fibers provide sensory input from the aortic body chemoreceptors and aortic arch baroreceptors, and the esophagus, bronchi, lungs, heart, and abdominal viscera in the foregut and midgut;
the SA fibers are for taste around the epiglottis and pharynx;
the GVE fibers are part of the parasympathetic part of the autonomic division of the PNS and stimulate smooth muscle and glands in the pharynx, larynx, thoracic viscera, and abdominal viscera of the foregut and midgut;
the BE fibers innervate one muscle of the tongue (palatoglossus), the muscles of the soft palate (except tensor veli palatini), pharynx (except stylopharyngeus), and larynx.
The vagus nerve arises as a group of rootlets on the anterolateral surface of the medulla oblongata just inferior to the rootlets arising to form the glossopharyngeal nerve [IX] (Fig. 8.49). The rootlets cross the posterior cranial fossa and enter the jugular foramen (Fig. 8.48). Within this foramen, and before exiting from it, the rootlets merge to form the vagus nerve [X]. Within or immediately outside the jugular foramen are two ganglia, the superior (jugular) and inferior (nodose) ganglia, which contain the cell bodies of the sensory neurons in the vagus nerve [X].
Accessory nerve [XI]
The accessory nerve [XI] is a cranial nerve that carries GSE fibers to innervate the sternocleidomastoid and trapezius muscles. It is a unique cranial nerve because its roots arise from motor neurons in the upper five segments of the cervical spinal cord. These fibers leave the lateral surface of the spinal cord and, joining together as they ascend, enter the cranial cavity through the foramen magnum (Fig. 8.49). The accessory nerve [XI] continues through the posterior cranial fossa and exits through the jugular foramen (Fig. 8.48). It then descends in the neck to innervate the sternocleidomastoid and trapezius muscles from their deep surfaces.
Cranial root of the accessory nerve
Some descriptions of the accessory nerve [XI] refer to a few rootlets arising from the caudal part of the medulla oblongata on the anterolateral surface just inferior to the rootlets arising to form the vagus nerve [X] as the “cranial” root of the accessory nerve (Fig. 8.49). Leaving the medulla, the cranial roots course with the “spinal” roots of the accessory nerve [XI] into the jugular foramen, at which point the cranial roots join the vagus nerve [X]. As part of the vagus nerve [X], they are distributed to the pharyngeal musculature innervated by the vagus nerve [X] and are therefore described as being part of the vagus nerve [X].
Hypoglossal nerve [XII]
The hypoglossal nerve [XII] carries GSE fibers to innervate all intrinsic and most of the extrinsic muscles of the tongue. It arises as several rootlets from the anterior surface of the medulla, passes laterally across the posterior cranial fossa and exits through the hypoglossal canal (Figs. 8.48 and 8.49). This nerve innervates the hyoglossus, styloglossus, and genioglossus muscles and all intrinsic muscles of the tongue.
In the clinic
Cranial nerve lesions
| Cranial nerve |
Clinical findings |
Example of lesion |
| Olfactory nerve [I] |
Loss of smell (anosmia) |
Injury to the cribriform plate; congenital absence |
| Optic nerve [II] |
Blindness/visual field abnormalities, loss of pupillary constriction |
Direct trauma to the orbit; disruption of the optic pathway |
| Oculomotor nerve [III] |
Dilated pupil, ptosis, loss of normal pupillary reflex, eye moves down inferiorly and laterally (down and out) |
Pressure from an aneurysm arising from the posterior communicating, posterior cerebral, or superior cerebellar artery; pressure from a herniating cerebral uncus (false localizing sign); cavernous sinus mass or thrombosis |
| Trochlear nerve [IV] |
Inability to look inferiorly when the eye is adducted (down and in) |
Along the course of the nerve around the brainstem; orbital fracture |
| Trigeminal nerve [V] |
Loss of sensation and pain in the region supplied by the three divisions of the nerve over the face; loss of motor function of the muscles of mastication on the side of the lesion |
Typically, in the region of the trigeminal ganglion, though local masses around the foramina through which the divisions pass can produce symptoms |
| Abducent nerve [VI] |
Inability of lateral eye movement |
Brain lesion or cavernous sinus lesion extending onto the orbit |
| Facial nerve [VII] |
Paralysis of facial muscles
Abnormal taste sensation from the anterior two-thirds of the tongue and dry conjunctivae
Paralysis of contralateral facial muscles below the eye |
Damage to the branches within the parotid gland
Injury to temporal bone; viral inflammation of nerve
Brainstem injury |
| Vestibulocochlear nerve [VIII] |
Progressive unilateral hearing loss and tinnitus (ringing in the ear) |
Tumor at the cerebellopontine angle |
| Glossopharyngeal nerve [IX] |
Loss of taste to the posterior one-third of the tongue and sensation of the soft palate |
Brainstem lesion; penetrating neck injury |
| Vagus nerve [X] |
Soft palate deviation with deviation of the uvula to the normal side; vocal cord paralysis |
Brainstem lesion; penetrating neck injury |
| Accessory nerve [XI] |
Paralysis of sternocleidomastoid and trapezius muscles |
Penetrating injury to the posterior triangle of the neck |
| Hypoglossal nerve [XII] |
Atrophy of ipsilateral muscles of the tongue and deviation toward the affected side; speech disturbance |
Penetrating injury to the neck and skull base pathology |
FACE
A face-to-face meeting is an important initial contact between individuals. Part of this exchange is the use of facial expressions to convey emotions. In fact, a physician can gain important information about an individual’s general health by observing a patient’s face.
Thus an understanding of the unique organization of the various structures between the superciliary arches superiorly, the lower edge of the mandible inferiorly, and as far back as the ears on either side, the area defined as the face, is particularly useful in the practice of medicine.
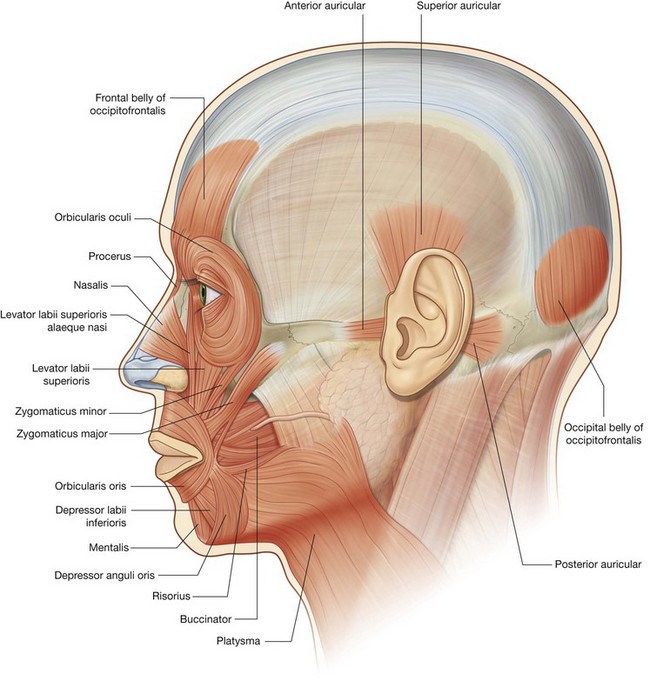
Muscles
The muscles of the face (Fig. 8.50) develop from the second pharyngeal arch and are innervated by branches of the facial nerve [VII]. They are in the superficial fascia, with origins from either bone or fascia, and insertions into the skin.
Because these muscles control expressions of the face, they are sometimes referred to as muscles of “facial expression.” They also act as sphincters and dilators of the orifices of the face (i.e., the orbits, nose, and mouth). This organizational arrangement into functional groups provides a logical approach to understanding these muscles (Table 8.7).
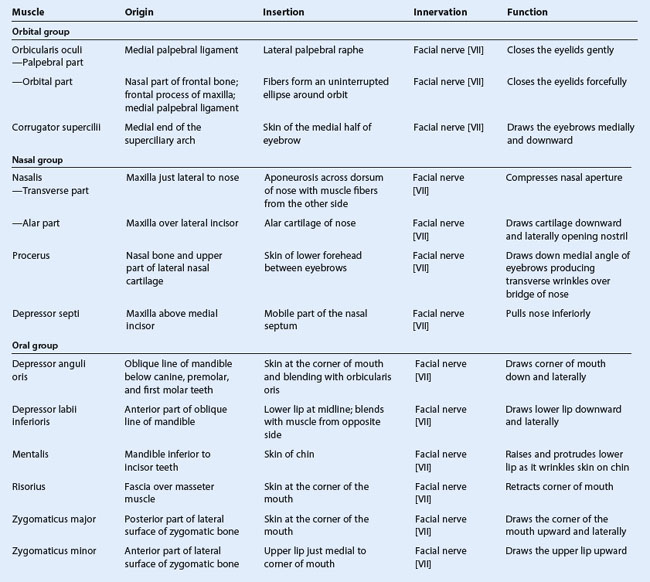
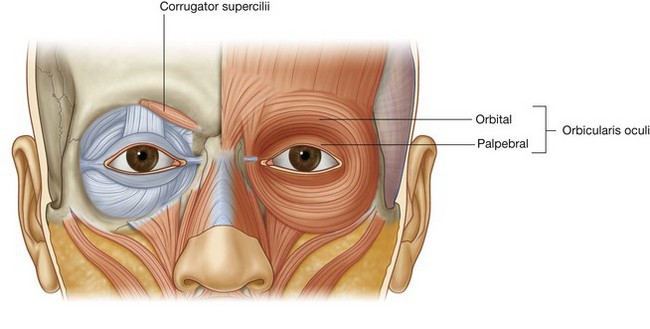
Orbital group
Two muscles are associated with the orbital group—the orbicularis oculi and the corrugator supercilii.
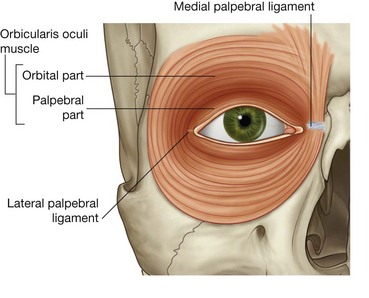
Orbicularis oculi
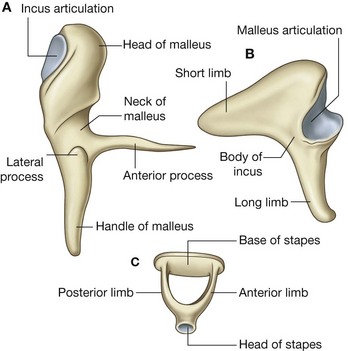
The orbicularis oculi is a large muscle that completely surrounds each orbital orifice and extends into each eyelid (Fig. 8.51). It closes the eyelids. It has two major parts:
the outer
orbital part is a broad ring that encircles the orbital orifice and extends outward beyond the orbital rim;
the inner
palpebral part is in the eyelids and consists of muscle fibers originating in the medial corner of the eye that arch across each lid to attach laterally.
The orbital and palpebral parts have specific roles to play during eyelid closure. The palpebral part closes the eye gently, whereas the orbital part closes the eye more forcefully and produces some wrinkling on the forehead.
An additional small lacrimal part of the orbicularis oculi muscle is deep, medial in position, and attaches to bone posterior to the lacrimal sac of the lacrimal apparatus in the orbit.
Corrugator supercilii
The second muscle in the orbital group is the much smaller corrugator supercilii (Fig. 8.51), which is deep to the eyebrows and the orbicularis oculi muscle and is active when frowning. It arises from the medial end of the superciliary arch, passing upward and laterally to insert into the skin of the medial half of the eyebrow. It draws the eyebrows toward the midline, causing vertical wrinkles above the nose.
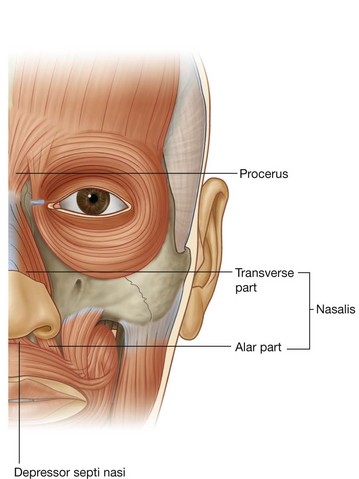
Nasal group
Three muscles are associated with the nasal group—the nasalis, the procerus, and the depressor septi nasi (Fig. 8.52).
Nasalis
The largest and best developed of the muscles of the nasal group is the nasalis, which is active when the nares are flared (Fig. 8.52). It consists of a transverse part (the compressor naris) and an alar part (the dilator naris):
the
transverse part of the nasalis compresses the nares—it originates from the maxilla and its fibers pass upward and medially to insert, along with fibers from the same muscle on the opposite side, into an aponeurosis across the dorsum of the nose;
the
alar part of the nasalis draws the alar cartilages downward and laterally, so opening the nares—it originates from the maxilla, below and medial to the transverse part, and inserts into the alar cartilage.
Procerus
The procerus is a small muscle superficial to the nasal bone and is active when an individual frowns (Fig. 8.52). It arises from the nasal bone and the upper part of the lateral nasal cartilage and inserts into the skin over the lower part of the forehead between the eyebrows. It may be continuous with the frontal belly of the occipitofrontalis muscle of the scalp.
Procerus draws the medial border of the eyebrows downward to produce transverse wrinkles over the bridge of the nose.
Depressor septi nasi
The final muscle in the nasal group is the depressor septi nasi, another muscle that assists in widening the nares (Fig. 8.52). Its fibers arise from the maxilla above the central incisor tooth and ascend to insert into the lower part of the nasal septum.
Depressor septi nasi pulls the nose inferiorly, so assisting the alar part of the nasalis in opening the nares.
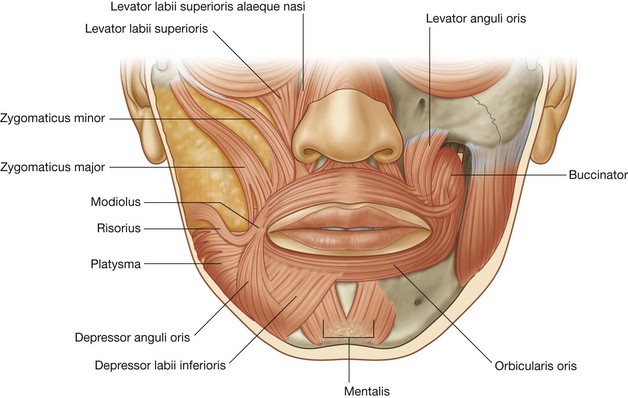
Oral group
The muscles in the oral group move the lips and cheek. They include the orbicularis oris and buccinator muscles, and a lower and upper group of muscles (Fig. 8.50). Many of these muscles intersect just lateral to the corner of the mouth on each side at a structure termed the modiolus.
Orbicularis oris
The orbicularis oris is a complex muscle consisting of fibers that completely encircle the mouth (Fig. 8.53). Its function is apparent when pursing the lips, as occurs during whistling. Some of its fibers originate near the midline from the maxilla superiorly and the mandible inferiorly, whereas other fibers are derived from both the buccinator, in the cheek, and the numerous other muscles acting on the lips. It inserts into the skin and mucous membrane of the lips, and into itself.
Contraction of the orbicularis oris narrows the mouth and closes the lips.
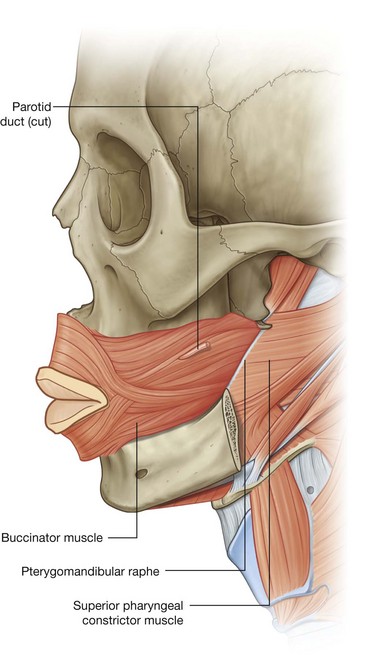
Buccinator
The buccinator forms the muscular component of the cheek and is used every time air expanding the cheeks is forcefully expelled (Figs. 8.53 and 8.54). It is in the space between the mandible and the maxilla, deep to the other facial muscles in the area.
The buccinator arises from the posterior part of the maxilla and mandible opposite the molar teeth and the pterygomandibular raphe, which is a tendinous band between the pterygoid hamulus superiorly and the mandible inferiorly and is a point of attachment for the buccinator and superior pharyngeal constrictor muscles.
The fibers of the buccinator pass toward the corner of the mouth to insert into the lips, blending with fibers from the orbicularis oris in a unique fashion. Central fibers of the buccinator cross so that lower fibers enter the upper lip and upper fibers enter the lower lip (Fig. 8.54). The highest and lowest fibers of the buccinator do not cross and enter the upper and lower lips, respectively.
Contraction of the buccinator presses the cheek against the teeth. This keeps the cheek taut and aids in mastication by preventing food from accumulating between the teeth and the cheek. It also assists in the forceful expulsion of air from the cheeks.
Lower group of oral muscles
The muscles in the lower group consist of the depressor anguli oris, depressor labii inferioris. and mentalis (Fig. 8.53).
Depressor anguli oris is active during frowning. It arises along the side of the mandible below the canine, premolar, and first molar teeth and inserts into skin and the upper part of the orbicularis oris near the corner of the mouth. It depresses the corner of the mouth.
Depressor labii inferioris arises from the front of the mandible, deep to depressor anguli oris. Its fibers move superiorly and medially, some merging with fibers from the same muscle on the opposite side and fibers from the orbicularis oris before inserting into the lower lip. It depresses the lower lip and moves it laterally.
Mentalis helps position the lip when drinking from a cup or when pouting. It is the deepest muscle of the lower group arising from the mandible just inferior to the incisor teeth, with its fibers passing downward and medially to insert into the skin of the chin. It raises and protrudes the lower lip as it wrinkles the skin of the chin.
Upper group of oral muscles
The muscles of the upper group of oral muscles consist of risorius, zygomaticus major, zygomaticus minor, levator labii superioris, levator labii superioris alaeque nasi, and levator anguli oris (Fig. 8.53).
Risorius helps produce a grin (
Fig. 8.53). It is a thin, superficial muscle that extends laterally from the corner of the mouth in a slightly upward direction. Contraction of its fibers pulls the corner of the mouth laterally and upward.
Zygomaticus major and
zygomaticus minor help produce a smile (
Fig. 8.53). Zygomaticus major is a superficial muscle that arises deep to the orbicularis oculi along the posterior part of the lateral surface of the zygomatic bone, and passes downward and forward, blending with the orbicularis oris and inserting into skin at the corner of the mouth. Zygomaticus minor arises from the zygomatic bone anterior to the origin of zygomaticus major, parallels the path of zygomaticus major, and inserts into the upper lip medial to the corner of the mouth. Both zygomaticus muscles raise the corner of the mouth and move it laterally.
Levator labii superioris deepens the furrow between the nose and the corner of the mouth during sadness (
Fig. 8.53). It arises from the maxilla just superior to the infra-orbital foramen, and its fibers pass downward and medially to blend with the orbicularis oris and insert into the skin of the upper lip.
Levator labii superioris alaeque nasi is medial to the levator labii superioris, arises from the maxilla next to the nose, and inserts into both the alar cartilage of the nose and skin of the upper lip (
Fig. 8.53). It may assist in flaring the nares.
Levator anguli oris is more deeply placed and covered by the other two levators and the zygomaticus muscles (
Fig. 8.53). It arises from the maxilla, just inferior to the infra-orbital foramen and inserts into the skin at the corner of the mouth. It elevates the corner of the mouth and may help deepen the furrow between the nose and the corner of the mouth during sadness.
Other muscles or muscle groups
Several additional muscles or groups of muscles not in the area defined as the face, but derived from the second pharyngeal arch and innervated by the facial nerve [VII], are considered muscles of facial expression. They include the platysma, auricular, and occipitofrontalis muscles (Fig. 8.50).
Platysma
Platysma is a large, thin sheet of muscle in the superficial fascia of the neck. It arises below the clavicle in the upper part of the thorax and ascends through the neck to the mandible. At this point, the more medial fibers insert on the mandible, whereas the lateral fibers join with muscles around the mouth.
Platysma tenses the skin of the neck and can move the lower lip and corners of the mouth down.
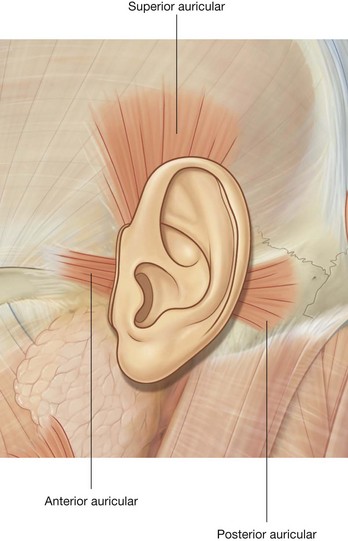
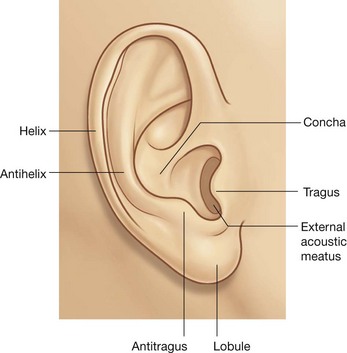
Auricular muscles
Three of these muscles, “other muscles of facial expression,” are associated with the ear—the anterior, superior, and posterior auricular muscles (Fig. 8.55):
the anterior muscle is anterolateral and pulls the ear upward and forward;
the superior muscle is superior and elevates the ear;
the posterior muscle is posterior and retracts and elevates the ear.
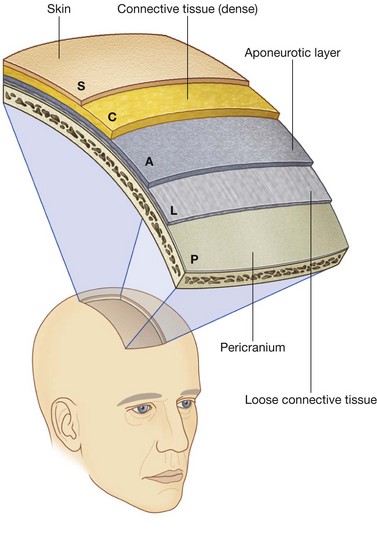
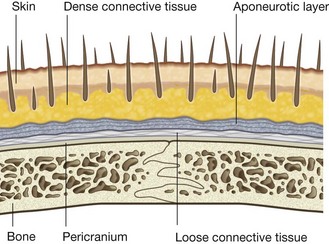
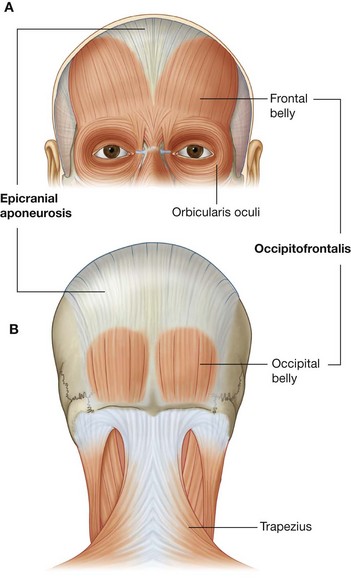
Occipitofrontalis
Occipitofrontalis is the final muscle in this category of “other muscles of facial expression” and is associated with the scalp (Fig. 8.50). It consists of a frontal belly anteriorly and an occipital belly posteriorly. An aponeurotic tendon connects the two:
the frontal belly covers the forehead and is attached to the skin of the eyebrows;
the occipital belly arises from the posterior aspect of the skull and is smaller than the frontal belly.
The occipitofrontalis muscles move the scalp and wrinkle the forehead.
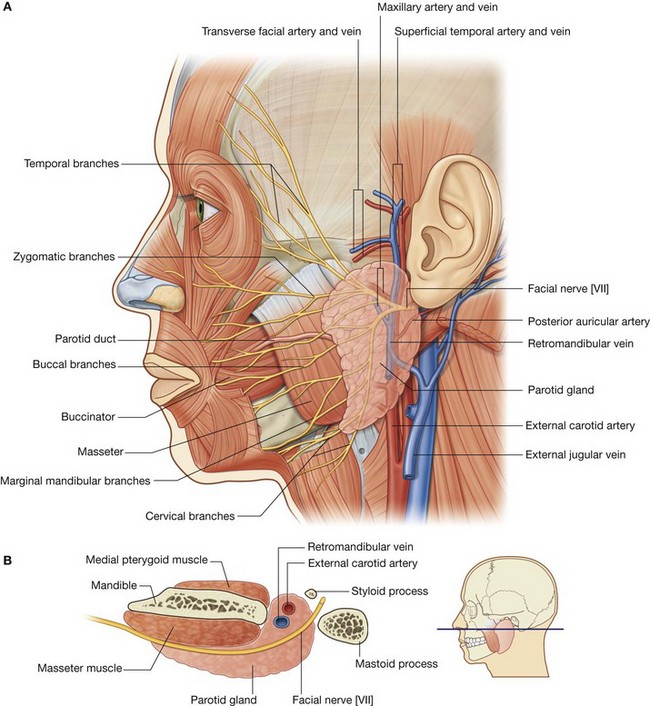
Parotid gland
The parotid glands are the largest of the three pairs of main salivary glands in the head and numerous structures pass through them. They are anterior to and below the lower half of the ear, superficial, posterior, and deep to the ramus of mandible (Fig. 8.56). They extend down to the lower border of the mandible and up to the zygomatic arch. Posteriorly they covers the anterior part of the sternocleidomastoid muscle and continues anteriorly to halfway across the masseter muscle.
The parotid duct leaves the anterior edge of the parotid gland midway between the zygomatic arch and the corner of the mouth (Fig. 8.56). It crosses the face in a transverse direction and, after crossing the medial border of the masseter muscle, turns deeply into the buccal fat pad and pierces the buccinator muscle. It opens into the oral cavity near the second upper molar tooth.
Important relationships
Several major structures enter and pass through or pass just deep to the parotid gland. These include the facial nerve [VII], the external carotid artery and its branches, and the retromandibular vein and its tributaries (Fig. 8.56).
Facial nerve
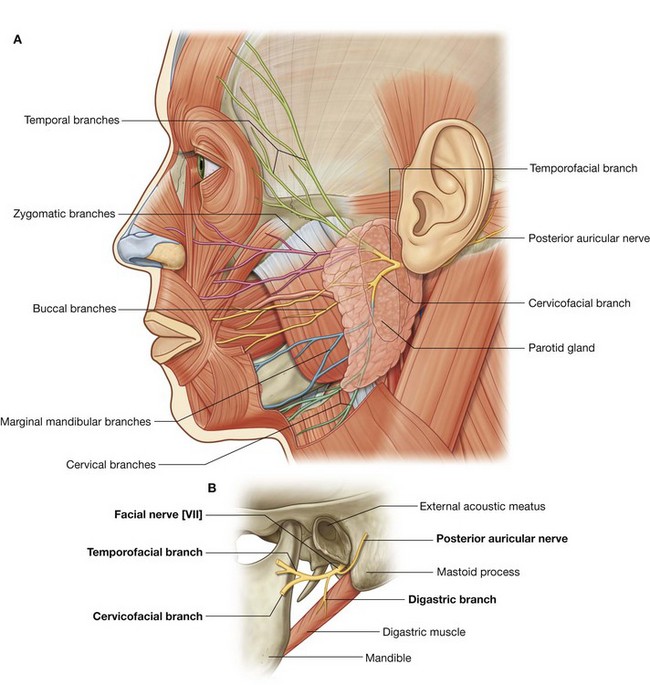
The facial nerve [VII] exits the skull through the stylomastoid foramen and then passes into the parotid gland, where it usually divides into upper and lower trunks. These pass through the substance of the parotid gland, where there may be further branching and anastomosing of the nerves.
Five terminal groups of branches of the facial nerve [VII]—the temporal, zygomatic, buccal, marginal mandibular, and cervical branches—emerge from the upper, anterior, and lower borders of the parotid gland (Fig. 8.56).
The intimate relationships between the facial nerve [VII] and the parotid gland mean that surgical removal of the parotid gland is a difficult dissection if all branches of the facial nerve [VII] are to be spared.
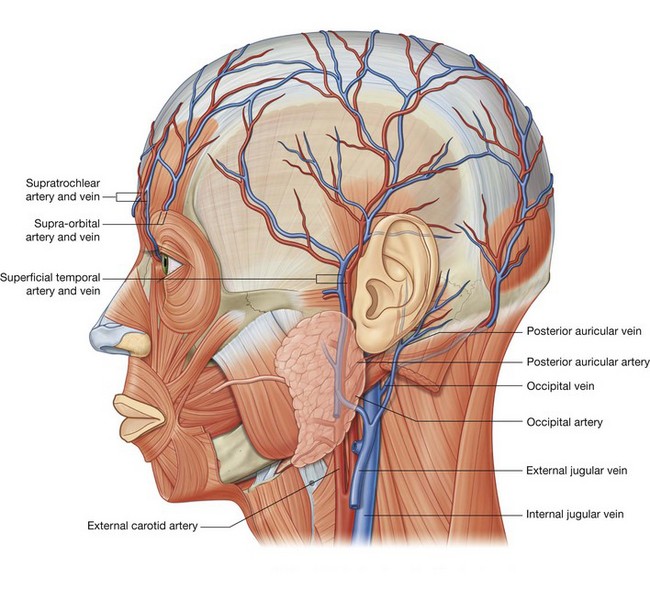
External carotid artery and its branches
The external carotid artery enters into or passes deep to the inferior border of the parotid gland (Fig. 8.56). As it continues in a superior direction, it gives off the posterior auricular artery before dividing into its two terminal branches (the maxillary and superficial temporal arteries) near the lower border of the ear:
the maxillary artery passes horizontally, deep to the mandible;
the superficial temporal artery continues in a superior direction and emerges from the upper border of the gland after giving off the
transverse facial artery.
Retromandibular vein and its tributaries
The retromandibular vein is formed in the substance of the parotid gland when the superficial temporal and maxillary veins join together (Fig. 8.56), and passes inferiorly in the substance of the parotid gland. It usually divides into anterior and posterior branches just below the inferior border of the gland.
Arterial supply
The parotid gland receives its arterial supply from the numerous arteries that pass through its substance.
Innervation
Sensory innervation of the parotid gland is provided by the auriculotemporal nerve, which is a branch of the mandibular nerve [V3]. This division of the trigeminal nerve exits the skull through the foramen ovale.
The auriculotemporal nerve also carries secretomotor fibers to the parotid gland. These postganglionic parasympathetic fibers have their origin in the otic ganglion associated with the mandibular nerve [V3] and are just inferior to the foramen ovale.
Preganglionic parasympathetic fibers to the otic ganglion come from the glossopharyngeal nerve [IX].
In the clinic
Parotid gland
The parotid gland is the largest of the paired salivary glands and is enclosed within the split investing layer of deep cervical fascia.
The parotid gland produces a watery saliva and salivary amylase, which are necessary for food bolus formation, oral digestion, and smooth passage of the bolus into the upper gastrointestinal tract.
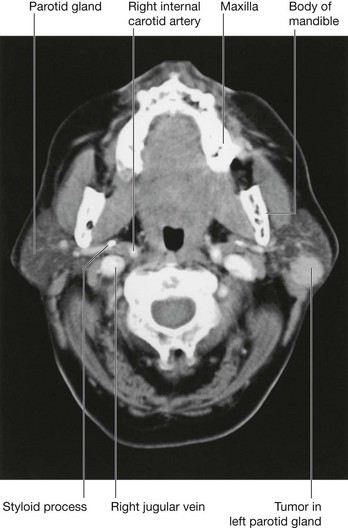
Tumors of the parotid gland
The commonest tumors of the parotid gland (Fig. 8.57) are benign and typically involve the superficial gland. These include pleomorphic adenoma and adenolymphoma. Their importance is in relation to their anatomical position. Critically, the relationship of the tumor to the branches of the facial nerve [VII] must be defined because resection may damage the facial nerve [VII]. In addition, if the tumor extends to the deep part of the gland the patient will need to provide consent for potential facial nerve [VII] damage.
Parotid gland stones
It is not uncommon for stones to develop within the parotid gland. They typically occur within the main confluence of the ducts and within the main parotid duct. The patient usually complains of intense pain when salivating and tends to avoid foods that produce this symptom. The pain can be easily reproduced in clinic by squirting lemon juice into the patient’s mouth.
Surgery depends upon where the stone is. If it is within the anterior aspect of the duct, a simple incision in the buccal mucosa with a sphincterotomy may allow removal. If the stone is farther back within the main duct, complete gland excision may be necessary.
Innervation
During development a cranial nerve becomes associated with each of the pharyngeal arches. Because the face is primarily derived from the first and second pharyngeal arches, innervation of neighboring facial structures varies as follows:
the trigeminal nerve [V] innervates facial structures derived from the first arch;
the facial nerve [VII] innervates facial structures derived from the second arch.
Sensory innervation
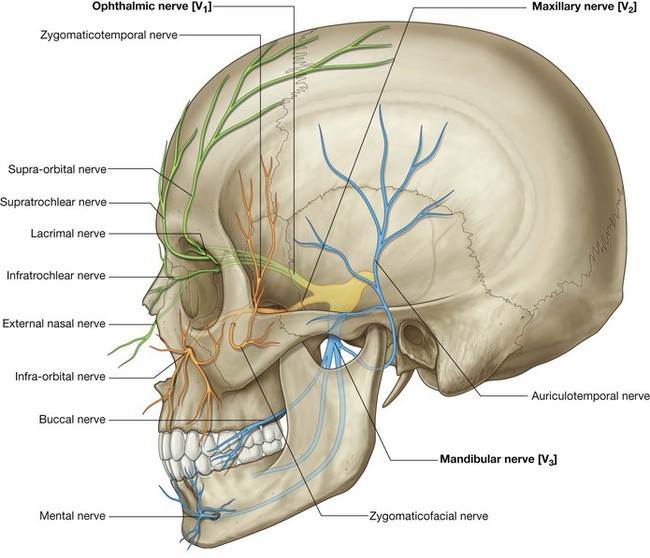
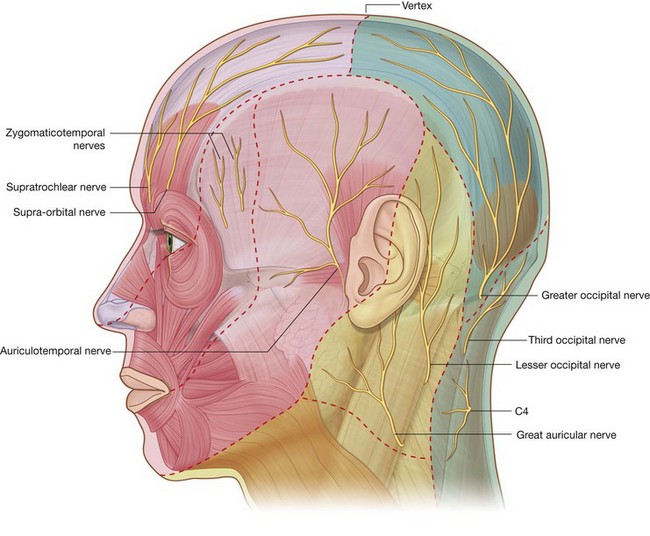
Because the face is derived developmentally from a number of structures originating from the first pharyngeal arch, cutaneous innervation of the face is by branches of the trigeminal nerve [V].
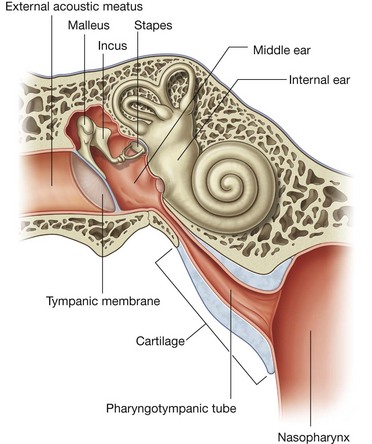
The trigeminal nerve [V] divides into three major divisions—the ophthalmic [V1], maxillary [V2], and mandibular [V3] nerves—before leaving the middle cranial fossa (Fig. 8.58). Each of these divisions passes out of the cranial cavity to innervate a part of the face, so most of the skin covering the face is innervated by branches of the trigeminal nerve [V]. The exception is a small area covering the angle and lower border of the ramus of mandible and parts of the ear, which are innervated by the trigeminal [V], facial [VII], vagus [X], and cervical nerves.
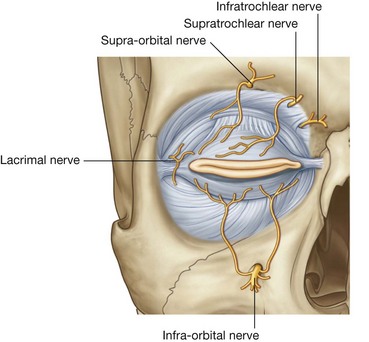
Ophthalmic nerve [V1]
The ophthalmic nerve [V1] exits the skull through the superior orbital fissure and enters the orbit. Its branches (Fig. 8.58) that innervate the face include:
the
supra-orbital and
supratrochlear nerves, which leave the orbit superiorly and innervate the upper eyelid, forehead, and scalp;
the
infratrochlear nerve, which exits the orbit in the medial angle to innervate the medial half of the upper eyelid, the skin in the area of the medial angle, and the side of the nose;
the
lacrimal nerve, which exits the orbit in the lateral angle to innervate the lateral half of the upper eyelid and the skin in the area of the lateral angle; and
the
external nasal nerve, which supplies the anterior part of the nose (
Fig. 8.59).
Maxillary nerve [V2]
The maxillary nerve [V2] exits the skull through the foramen rotundum. Branches (Fig. 8.58) that innervate the face include:
a small
zygomaticotemporal branch, which exits the zygomatic bone and supplies a small area of the anterior temple above the zygomatic arch;
a small
zygomaticofacial branch, which exits the zygomatic bone and supplies a small area of skin over the zygomatic bone; and
the large
infra-orbital nerve, which exits the maxilla through the infra-orbital foramen and immediately divides into multiple branches to supply the lower eyelid, cheek, side of the nose, and upper lip (
Fig. 8.59).
Mandibular nerve [V3]
The mandibular nerve [V3] exits the skull through the foramen ovale. Branches (Fig. 8.58) innervating the face include:
the auriculotemporal nerve, which enters the face just posterior to the temporomandibular joint, passes through the parotid gland, and ascends just anterior to the ear to supply the external acoustic meatus, the surface of the tympanic membrane (eardrum), and a large area of the temple;
the
buccal nerve, which is on the surface of the buccinator muscle supplying the cheek; and
the
mental nerve, which exits the mandible through the mental foramen and immediately divides into multiple branches to supply the skin and mucous membrane of the lower lip and skin of the chin (
Fig. 8.59).
Motor innervation
The muscles of the face, as well as those associated with the ear and the scalp, are derived from the second pharyngeal arch. The cranial nerve associated with this arch is the facial nerve [VII] and therefore branches of the facial nerve [VII] innervate all these muscles.
The facial nerve [VII] exits the posterior cranial fossa through the internal acoustic meatus. It passes through the temporal bone, giving off several branches, and emerges from the base of the skull through the stylomastoid foramen (Fig. 8.60). At this point it gives off the posterior auricular nerve. This branch passes upward, behind the ear, to supply the occipital belly of the occipitofrontalis muscle of the scalp and the posterior auricular muscle of the ear.
The main stem of the facial nerve [VII] then gives off another branch, which innervates the posterior belly of the digastric muscle and the stylohyoid muscle. At this point, the facial nerve [VII] enters the deep surface of the parotid gland (Fig. 8.60B).
Once in the parotid gland, the main stem of the facial nerve [VII] usually divides into upper (temporofacial) and lower (cervicofacial) branches. As these branches pass through the substance of the parotid gland they may branch further or take part in an anastomotic network (the parotid plexus).
Whatever types of interconnections occur, five terminal groups of branches of the facial nerve [VII]—the temporal, zygomatic, buccal, marginal mandibular, and cervical branches—emerge from the parotid gland (Fig. 8.60A).
Although there are variations in the pattern of distribution of the five terminal groups of branches, the basic pattern is as follows:
temporal branches exit from the superior border of the parotid gland to supply muscles in the area of the temple, forehead, and supra-orbital area;
zygomatic branches emerge from the anterosuperior border of the parotid gland to supply muscles in the infra-orbital area, the lateral nasal area, and the upper lip;
buccal branches emerge from the anterior border of the parotid gland to supply muscles in the cheek, the upper lip, and the corner of the mouth;
marginal mandibular branches emerge from the anteroinferior border of the parotid gland to supply muscles of the lower lip and chin;
cervical branches emerge from the inferior border of the parotid gland to supply the platysma.
Vessels
The arterial supply to the face is primarily from branches of the external carotid artery, though there is some limited supply from a branch of the internal carotid artery.
Similarly, most of the venous return is back to the internal jugular vein, though some important connections from the face result in venous return through a clinically relevant intracranial pathway involving the cavernous sinus.
Arteries
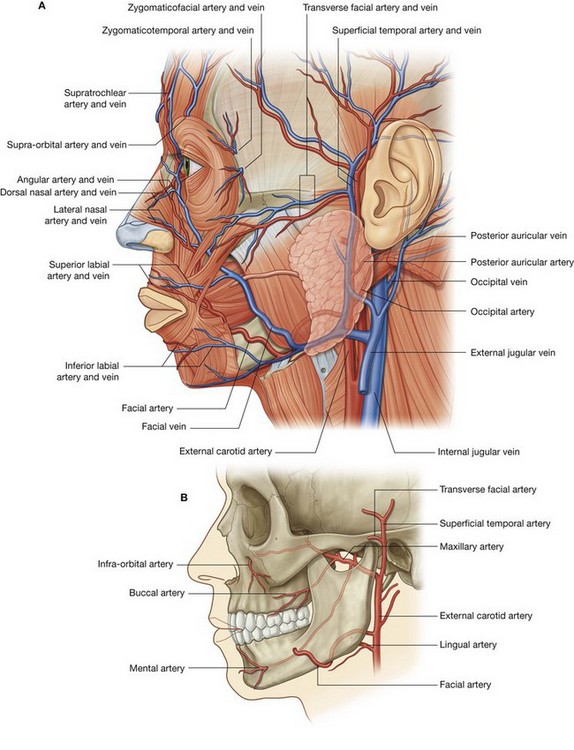
Facial artery
The facial artery is the major vessel supplying the face (Fig. 8.61). It branches from the anterior surface of the external carotid artery, passes up through the deep structures of the neck and appears at the lower border of the mandible after passing posterior to the submandibular gland.
Curving around the inferior border of the mandible just anterior to the masseter, where its pulse can be felt, the facial artery then enters the face.
From this point the facial artery runs upward and medially in a tortuous course. It passes along the side of the nose and terminates as the angular artery at the medial corner of the eye.
Along its path the facial artery is deep to the platysma, risorius, and zygomaticus major and minor, superficial to the buccinator and levator anguli oris, and may pass superficially to or through the levator labii superioris.
Branches of the facial artery include the superior and inferior labial branches and the lateral nasal branch (Fig. 8.61).
The labial branches arise near the corner of the mouth:
the
inferior labial branch supplies the lower lip;
the
superior labial branch supplies the upper lip, and also provides a branch to the nasal septum.
Near the midline, the superior and inferior labial branches anastomose with their companion arteries from the opposite side of the face. This provides an important connection between the facial arteries and the external carotid arteries of opposite sides.
The lateral nasal branch is a small branch arising from the facial artery as it passes along the side of the nose. It supplies the lateral surface and dorsum of the nose.
Transverse facial artery
Another contributor to the vascular supply of the face is the transverse facial artery (Fig. 8.61), which is a branch of the superficial temporal artery (the smaller of the two terminal branches of the external carotid artery).
The transverse facial artery arises from the superficial temporal artery within the substance of the parotid gland, passes through the gland, and crosses the face in a transverse direction. Lying on the superficial surface of the masseter muscle, it is between the zygomatic arch and the parotid duct.
Branches of the maxillary artery
The maxillary artery, the larger of the two terminal branches of the external carotid artery, gives off several small branches which contribute to the arterial supply to the face:
the
infra-orbital artery enters the face through the infra-orbital foramen and supplies the lower eyelid, upper lip, and the area between these structures.
the
buccal artery enters the face on the superficial surface of the buccinator muscle and supplies structures in this area;
the
mental artery enters the face through the mental foramen and supplies the chin.
Branches of the ophthalmic artery
Three small arteries from the internal carotid artery also contribute to the arterial supply of the face. These vessels arise from the ophthalmic artery, a branch of the internal carotid artery, after the ophthalmic artery enters the orbit:
the
zygomaticofacial and
zygomaticotemporal arteries come from the lacrimal branch of the ophthalmic artery (
Fig. 8.61), enter the face through the zygomaticofacial and zygomaticotemporal foramina, and supply the area of the face over the zygomatic bone;
the
dorsal nasal artery, a terminal branch of the ophthalmic artery, exits the orbit in the medial corner, and supplies the dorsum of the nose.
Other branches of the ophthalmic artery (the supra-orbital and supratrochlear arteries) supply the anterior scalp.
Veins
Facial vein
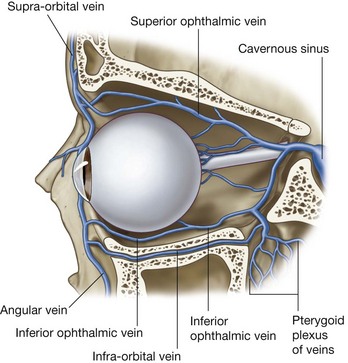
The facial vein is the major vein draining the face (Fig. 8.61). Its point of origin is near the medial corner of the orbit as the supratrochlear and supra-orbital veins come together to form the angular vein. This vein becomes the facial vein as it proceeds inferiorly and assumes a position just posterior to the facial artery. The facial vein descends across the face with the facial artery until it reaches the inferior border of the mandible. Here the artery and vein part company and the facial vein passes superficial to the submandibular gland to enter the internal jugular vein.
Throughout its course the facial vein receives tributaries from veins draining the eyelids, external nose, lips, cheek, and chin that accompany the various branches of the facial artery.
Transverse facial vein
The transverse facial vein is a small vein that accompanies the transverse facial artery in its journey across the face (Fig. 8.61). It empties into the superficial temporal vein within the substance of the parotid gland.
Intracranial venous connections
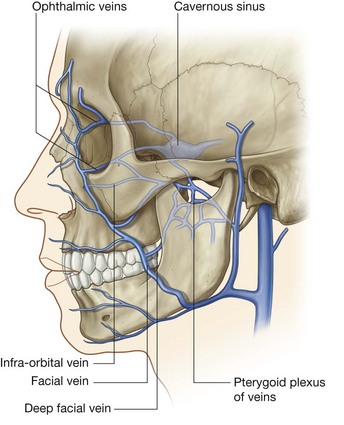
As it crosses the face, the facial vein has numerous connections with venous channels passing into deeper regions of the head (Fig. 8.62):
near the medial corner of the orbit, it communicates with ophthalmic veins;
in the area of the cheek it communicates with veins passing into the infra-orbital foramen;
it communicates with veins passing into deeper regions of the face (i.e., the deep facial vein connecting with the pterygoid plexus of veins).
All these venous channels have interconnections with the intracranial cavernous sinus through emissary veins that connect intracranial with extracranial veins. There are no valves in the facial vein or any other venous channels in the head, so blood can move in any direction. Because of the interconnections between the veins, infections of the face, primarily above the mouth (i.e., the “danger area”) should be handled with great care to prevent the dissemination of infectious material in an intracranial direction.
Lymphatic drainage
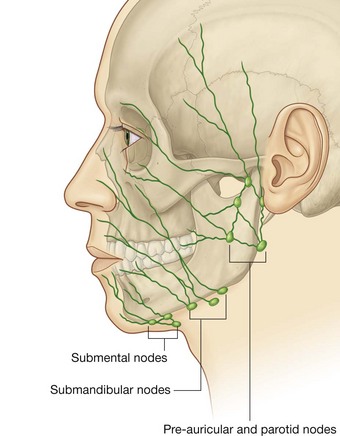
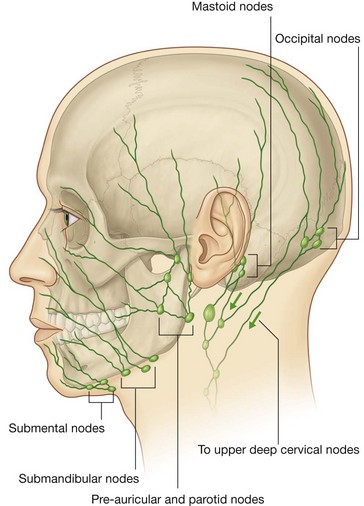
Lymphatic drainage from the face primarily moves toward three groups of lymph nodes (Fig. 8.63):
submental nodes inferior and posterior to the chin, which drain lymphatics from the medial part of the lower lip and chin bilaterally;
submandibular nodes superficial to the submandibular gland and inferior to the body of the mandible, which drain the lymphatics from the medial corner of the orbit, most of the external nose, medial part of the cheek, the upper lip and the lateral part of the lower lip that follow the course of the facial artery;
pre-auricular and parotid nodes anterior to the ear, which drain lymphatics from most of the eyelids, a part of the external nose, and the lateral part of the cheek.
In the clinic
Facial nerve [VII] palsy (Bell’s palsy)
The complexity of the facial nerve [VII] is demonstrated by the different pathological processes and sites at which these processes occur.
The facial nerve [VII] is formed from the nuclei within the brainstem emerging at the junction of the pons and the medulla. It enters the internal acoustic meatus, passes to the geniculate ganglion (which gives rise to further branches), and emerges from the skull base after a complex course within the temporal bone, leaving through the stylomastoid foramen. It enters the parotid gland and gives rise to five terminal groups of branches that supply muscles in the face and a number of additional branches that supply deeper or more posterior muscles. A series of lesions may affect the nerve along its course and it is possible, with good clinical expertise, to determine the exact site of the lesion in relation to the course of the nerve.
Central lesions
A primary brainstem lesion affecting the motor nucleus of the facial nerve (VII) would lead to ipsilateral (same side) weakness of the whole face. However, because the upper part of the nucleus receives motor input from the left and right cerebral hemispheres a lesion occurring above the nucleus leads to contralateral lower facial weakness. In this example, motor innervation to the upper face is spared because the upper part of the nucleus receives input from both hemispheres. Preservation and loss of the special functions are determined by the extent of the lesion.
Lesions at and around the geniculate ganglion
Typically lesions at and around the geniculate ganglion are accompanied by loss of motor function on the whole of the ipsilateral (same) side of the face. Taste to the anterior two-thirds of the tongue, lacrimation, and some salivation also are likely to be affected because the lesion is proximal to the greater petrosal and chorda tympani branches of the nerve.
Lesions at and around the stylomastoid foramen
Lesions at and around the stylomastoid foramen are the commonest abnormality of the facial nerve [VII] and usually result from a viral inflammation of the nerve within the bony canal before exiting through the stylomastoid foramen. Typically the patient has an ipsilateral loss of motor function of the whole side of the face. Not only does this produce an unusual appearance, but it also complicates chewing of food. Lacrimation and taste may not be affected if the lesion remains distal to the greater petrosal and chorda tympani branches that originate deep in the temporal bone.
In the clinic
Trigeminal neuralgia
Trigeminal neuralgia (tic douloureux) is a complex sensory disorder of the sensory root of the trigeminal nerve. Typically the pain is in the region of the mandibular [V3] and maxillary [V2] nerves, and is typically of sudden onset, excruciating in nature, and may be triggered by touching a sensitive region of skin.
The etiology of trigeminal neuralgia is unknown, although anomalous blood vessels lying adjacent to the sensory route of the maxillary [V2] and mandibular [V3] nerves may be involved.
If symptoms persist and are unresponsive to medical care, surgical exploration of the trigeminal nerve (which is not without risk) may be necessary to remove any aberrant vessels.
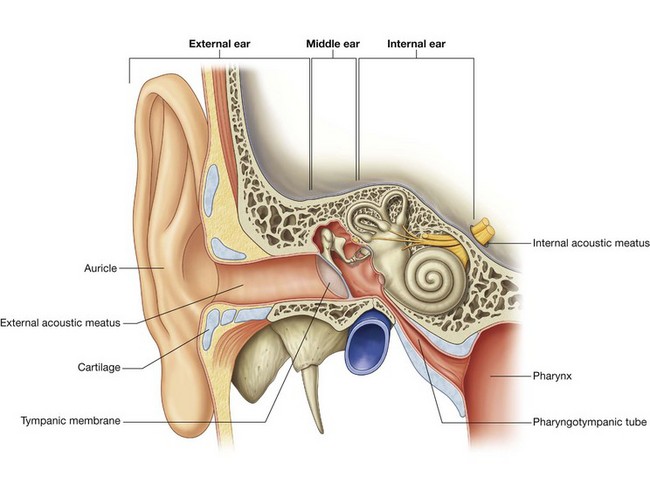
ORBIT
The orbits are bilateral structures in the upper half of the face below the anterior cranial fossa and anterior to the middle cranial fossa that contain the eyeball, the optic nerve, the extra-ocular muscles, the lacrimal apparatus, adipose tissue, fascia, and the nerves and vessels that supply these structures.
Bony orbit
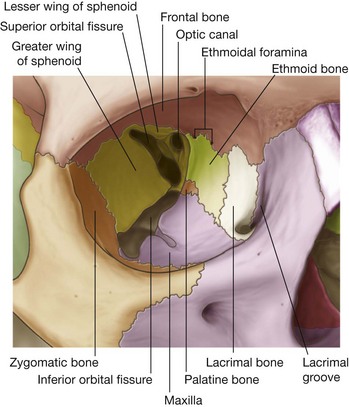
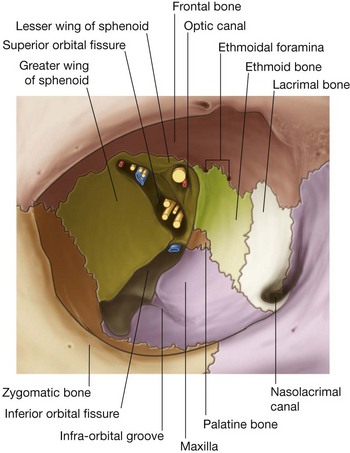
Seven bones contribute to the framework of each orbit (Fig. 8.70). They are the maxilla, zygomatic, frontal, ethmoid, lacrimal, sphenoid, and palatine bones. Together they give the bony orbit the shape of a pyramid, with its wide base opening anteriorly onto the face, and its apex extending in a posteromedial direction. Completing the pyramid configuration are medial, lateral, superior, and inferior walls.
The apex of the pyramid-shaped bony orbit is the optic foramen, whereas the base (the orbital rim) is formed:
superiorly by the frontal bone;
medially by the frontal process of the maxilla;
inferiorly by the zygomatic process of the maxilla and the zygomatic bone; and
laterally by the zygomatic bone, the frontal process of the zygomatic bone, and the zygomatic process of the frontal bone.
Roof
The roof (superior wall) of the bony orbit is made up of the orbital part of the frontal bone with a small contribution from the sphenoid bone (Fig. 8.70). This thin plate of bone separates the contents of the orbit from the brain in the anterior cranial fossa.
Unique features of the superior wall include:
anteromedially, the possible intrusion of part of the frontal sinus and the trochlear fovea, for the attachment of a pulley through which the superior oblique muscle passes;
anterolaterally, a depression (the lacrimal fossa) for the orbital part of the lacrimal gland.
Posteriorly, the lesser wing of the sphenoid bone completes the roof.
Medial wall
The medial walls of the paired bony orbits are parallel to each other and each consists of four bones—the maxilla, lacrimal, ethmoid, and sphenoid bones (Fig. 8.70).
The largest contributor to the medial wall is the orbital plate of the ethmoid bone. This part of the ethmoid bone contains collections of ethmoidal cells, which are clearly visible in a dried skull.
Also visible, at the junction between the roof and the medial wall, usually associated with the frontoethmoidal suture, are the anterior and posterior ethmoidal foramina. The anterior and posterior ethmoidal nerves and vessels leave the orbit through these openings.
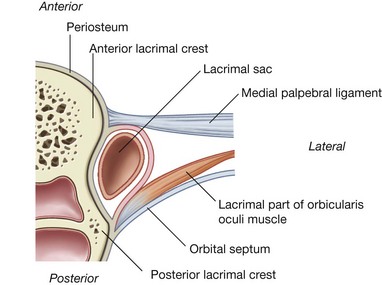
Anterior to the ethmoid bone is the small lacrimal bone, and completing the anterior part of the medial wall is the frontal process of the maxilla. These two bones participate in the formation of the lacrimal groove, which contains the lacrimal sac and is bound by the posterior lacrimal crest (part of the lacrimal bone) and the anterior lacrimal crest (part of the maxilla).
Posterior to the ethmoid bone the medial wall is completed by a small part of the sphenoid bone, which forms a part of the medial wall of the optic canal.
Floor
The floor (inferior wall) of the bony orbit, which is also the roof of the maxillary sinus, consists primarily of the orbital surface of the maxilla (Fig. 8.70), with small contributions from the zygomatic and palatine bones.
Beginning posteriorly and continuing along the lateral boundary of the floor of the bony orbit is the inferior orbital fissure. Beyond the anterior end of the fissure the zygomatic bone completes the floor of the bony orbit.
Posteriorly, the orbital process of the palatine bone makes a small contribution to the floor of the bony orbit near the junction of the maxilla, ethmoid, and sphenoid bones.
Lateral wall
The lateral wall of the bony orbit consists of contributions from two bones—anteriorly, the zygomatic bone and posteriorly, the greater wing of the sphenoid bone (Fig. 8.70).
Eyelids
The upper and lower eyelids are anterior structures that, when closed, protect the surface of the eyeball.
The space between the eyelids, when they are open, is the palpebral fissure.
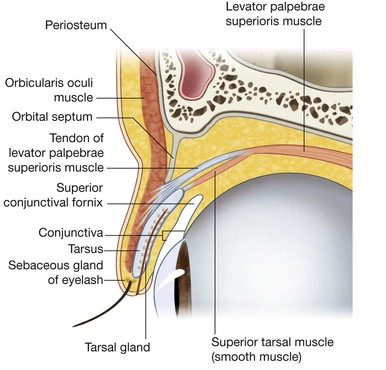
The layers of the eyelids, from anterior to posterior, consist of skin, subcutaneous tissue, voluntary muscle, the orbital septum, the tarsus, and conjunctiva (Fig. 8.71).
The upper and lower eyelids are basically similar in structure except for the addition of two muscles in the upper eyelid.
Skin and subcutaneous tissue
The skin of the eyelids is not particularly substantial and only a thin layer of connective tissue separates the skin from the underlying voluntary muscle layer (Fig. 8.71).
The thin layer of connective tissue and its loose arrangement account for the accumulation of fluid (blood) when an injury occurs.
Orbicularis oculi
The muscle fibers encountered next in an anteroposterior direction through the eyelid belong to the palpebral part of orbicularis oculi (Fig. 8.71). This muscle is part of the larger orbicularis oculi muscle, which consists primarily of two parts—an orbital part, which surrounds the orbit, and the palpebral part, which is in the eyelids (Fig. 8.72). Orbicularis oculi is innervated by the facial nerve [VII] and closes the eyelids.
The palpebral part is thin and anchored medially by the medial palpebral ligament, which attaches to the anterior lacrimal crest, and laterally blends with fibers from the muscle in the lower eyelid at the lateral palpebral ligament.
A third part of the orbicularis oculi muscle that can be identified consists of fibers on the medial border, which pass deeply to attach to the posterior lacrimal crest. These fibers form the lacrimal part of the orbicularis oculi, which may be involved in the drainage of tears.
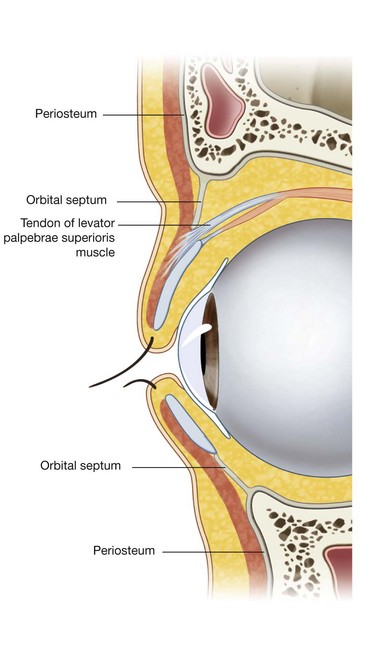
Orbital septum
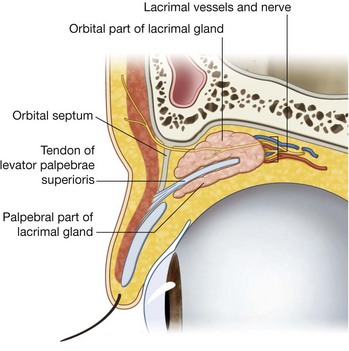
Deep to the palpebral part of the orbicularis oculi is an extension of periosteum into both the upper and lower eyelids from the margin of the orbit (Fig. 8.71). This is the orbital septum, which extends downward into the upper eyelid and upward into the lower eyelid and is continuous with the periosteum outside and inside the orbit (Fig. 8.73). The orbital septum attaches to the tendon of levator palpebrae superioris muscle in the upper eyelid and attaches to the tarsus in the lower eyelid.
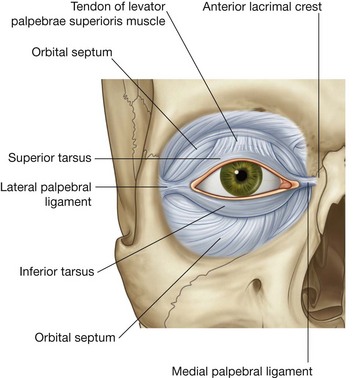
Tarsus and levator palpebrae superioris
Providing major support for each eyelid is the tarsus (Fig. 8.71). There is a large superior tarsus in the upper eyelid and a smaller inferior tarsus in the lower eyelid (Fig. 8.74). These plates of dense connective tissue are attached medially to the anterior lacrimal crest of the maxilla by the medial palpebral ligament and laterally to the orbital tubercle on the zygomatic bone by the lateral palpebral ligament.
Although the tarsal plates in the upper and lower eyelids are generally similar in structure and function, there is one unique difference. Associated with the tarsus in the upper eyelid is the levator palpebrae superioris muscle (Fig. 8.74), which raises the eyelid. Its origin is from the posterior part of the roof of the orbit, just superior to the optic foramen, and it inserts into the anterior surface of the superior tarsus, with the possibility of a few fibers attaching to the skin of the upper eyelid. It is innervated by the oculomotor nerve [III].
In companion with the levator palpebrae superioris muscle is a collection of smooth muscle fibers passing from the inferior surface of the levator to the upper edge of the superior tarsus (Fig. 8.71). Innervated by postganglionic sympathetic fibers from the superior cervical ganglion, this muscle is the superior tarsal muscle.
Loss of function of either the levator palpebrae superioris muscle or the superior tarsal muscle results in a ptosis or drooping of the upper eyelid.
Conjunctiva
The structure of the eyelid is completed by a thin membrane (the conjunctiva), which covers the posterior surface of each eyelid (Fig. 8.71). This membrane covers the full extent of the posterior surface of each eyelid before reflecting onto the outer surface (sclera) of the eyeball. It attaches to the eyeball at the junction between the sclera and the cornea. With this membrane in place, a conjunctival sac is formed when the eyelids are closed, and the upper and lower extensions of this sac are the superior and inferior conjunctival fornices (Fig. 8.71).
Glands
Embedded in the tarsal plates are tarsal glands, which empty onto the free margin of each eyelid. These glands are modified sebaceous glands and secrete an oily substance that increases the viscosity of the tears and decreases the rate of evaporation of tears from the surface of the eyeball.
Blockage and inflammation of a tarsal gland is a chalazion and is on the inner surface of the eyelid.
The tarsal glands are not the only glands associated with the eyelids. Associated with the eyelash follicles are sebaceous and sweat glands.
Blockage and inflammation of either of these is a stye and is on the edge of the eyelid.
Vessels
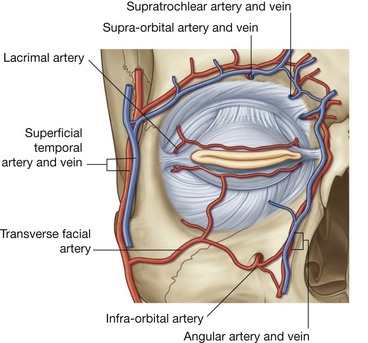
The arterial supply to the eyelids is from the numerous vessels in the area. They include:
the supratrochlear, supra-orbital, lacrimal, and dorsal nasal arteries from the ophthalmic artery;
the angular artery from the facial artery;
the transverse facial artery from the superficial temporal artery; and
branches from the superficial temporal artery itself (
Fig. 8.75).
Venous drainage follows an external pattern through veins associated with the various arteries and an internal pattern moving into the orbit through connections with the ophthalmic veins.
Lymphatic drainage is primarily to the parotid nodes, with some drainage from the medial corner of the eye along lymphatic vessels associated with the angular and facial arteries to the submandibular nodes.
Innervation
Innervation of the eyelids includes both sensory and motor components.
The sensory nerves are all branches of the trigeminal nerve [V]. Palpebral branches arise from:
the supra-orbital, supratrochlear, infratrochlear, and lacrimal branches of the ophthalmic nerve [V
1]; and
the infra-orbital branch of the maxillary nerve [V
2] (
Fig. 8.76).
Motor innervation is from:
the facial nerve [VII], which innervates the palpebral part of the orbicularis oculi;
the oculomotor nerve [III], which innervates the levator palpebrae superioris;
sympathetic fibers, which innervate the superior tarsal muscle.
Loss of innervation of the orbicularis oculi by the facial nerve [VII] causes an inability to close the eyelids tightly and the lower eyelid droops away, resulting in a spillage of tears.
Loss of innervation of the levator palpebrae superioris by the oculomotor nerve causes an inability to open the superior eyelid voluntarily, producing a complete ptosis.
Loss of innervation of the superior tarsal muscle by sympathetic fibers causes a constant partial ptosis.
In the clinic
Horner’s syndrome
Horner’s syndrome is caused by a lesion in the sympathetic trunk in the neck that results in sympathetic dysfunction. It is characterized by three typical features:
pupillary constriction due to paralysis of the dilator pupillae muscle;
partial ptosis (drooping of the upper eyelid) due to paralysis of the superior tarsal muscle of the levator palpebrae superioris;
absence of sweating on the ipsilateral side of the face and the neck due to absence of innervation of the sweat glands.
Secondary changes may also include:
ipsilateral vasodilation due to loss of the normal
sympathetic control of the subcutaneous blood vessels; and
enophthalmos (sinking of the eye)—believed to result from paralysis of the orbitalis muscle, though this is an uncommon feature of Horner’s syndrome.
The orbitalis muscle spans the inferior orbital fissure and helps maintain the forward position of orbital contents.
The commonest cause for Horner’s syndrome is a tumor eroding the cervicothoracic ganglion, which is typically an apical lung tumor.
Surgically induced Horner’s syndrome
A surgically induced Horner’s syndrome may be necessary for patients who suffer severe hyperhidrosis (sweating). This often debilitating condition may be so severe that patients are confined to their home for fear of embarrassment. Treatment is relatively straightforward and somewhat ingenious. The patient is anesthetized and a bifurcate endotracheal tube is placed into the left and right main bronchi. A small incision is made in the intercostal space on the appropriate side and a surgically-induced pneumothorax is created. The patient is ventilated through the contralateral lung.
Using an endoscope the apex of the thoracic cavity can be viewed from inside and the cervicothoracic ganglion readily identified. Obliterative techniques include thermocoagulation and surgical excision. After the ganglion has been destroyed, the endoscope is removed, the lung is re-inflated, and the small hole is oversewn.
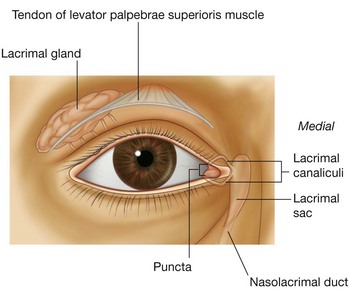
Lacrimal apparatus
The lacrimal apparatus is involved in the production, movement, and drainage of fluid from the surface of the eyeball. It is made up of the lacrimal gland and its ducts, the lacrimal canaliculi, the lacrimal sac, and the nasolacrimal duct.
The lacrimal gland is anterior in the superolateral region of the orbit (Fig. 8.77) and is divided into two parts by the levator palpebrae superioris (Fig. 8.78):
the larger
orbital part is in a depression, the lacrimal fossa, in the frontal bone;
the smaller
palpebral part is inferior to levator palpebrae superioris in the superolateral part of the eyelid.
Numerous ducts empty the glandular secretions into the lateral part of the superior fornix of the conjunctiva.
Fluid is continually being secreted by the lacrimal gland and moved across the surface of the eyeball from lateral to medial as the eyelids blink.
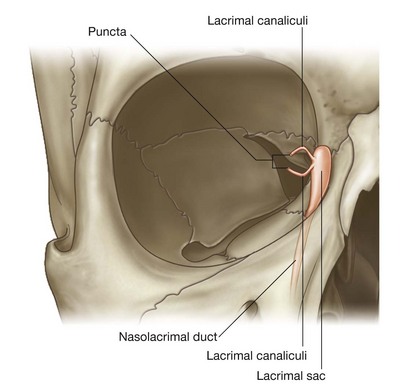
The fluid accumulates medially in the lacrimal lake and is drained from the lake by the lacrimal canaliculi, one canaliculus associated with each eyelid (Fig. 8.77). The lacrimal punctum is the opening through which fluid enters each canaliculus.
Passing medially, the lacrimal canaliculi eventually join the lacrimal sac between the anterior and posterior lacrimal crests, posterior to the medial palpebral ligament and anterior to the lacrimal part of the orbicularis oculi muscle (Figs. 8.79 and 8.80). When the orbicularis oculi muscle contracts during blinking, the small lacrimal part of the muscle may dilate the lacrimal sac and draw tears into it through the canaliculi from the conjunctival sac.
Innervation
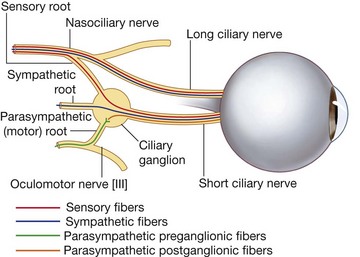
The innervation of the lacrimal gland involves three different components (Fig. 8.81).
Sensory innervation
Sensory neurons from the lacrimal gland return to the CNS through the lacrimal branch of the ophthalmic nerve [V1].
Secretomotor (parasympathetic) innervation
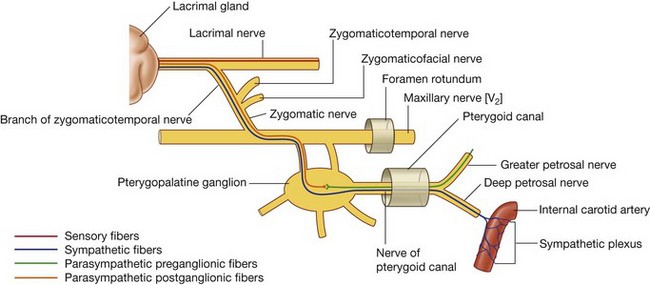
Secretomotor fibers from the parasympathetic part of the autonomic division of the PNS stimulate fluid secretion from the lacrimal gland. These preganglionic parasympathetic neurons leave the CNS in the facial nerve [VII], enter the greater petrosal nerve (a branch of the facial nerve [VII]), and continue with this nerve until it becomes the nerve of the pterygoid canal (Fig. 8.81).
The nerve of the pterygoid canal eventually joins the pteryatine ganglion where the preganglionic parasympathetic neurons synapse on postganglionic parasympathetic neurons. The postganglionic neurons join the maxillary nerve [V2] and continue with it until the zygomatic nerve branches from it, and travel with the zygomatic nerve until it gives off the zygomaticotemporal nerve, which eventually distributes postganglionic parasympathetic fibers in a small branch that joins the lacrimal nerve. The lacrimal nerve passes to the lacrimal gland.
Sympathetic innervation
Sympathetic innervation of the lacrimal gland follows a similar path as parasympathetic innervation. Postganglionic sympathetic fibers originating in the superior cervical ganglion travel along the plexus surrounding the internal carotid artery (Fig. 8.81). They leave this plexus as the deep petrosal nerve and join the parasympathetic fibers in the nerve of the pterygoid canal. Passing through the pteryatine ganglion, the sympathetic fibers from this point onward follow the same path as the parasympathetic fibers to the lacrimal gland.
Vessels
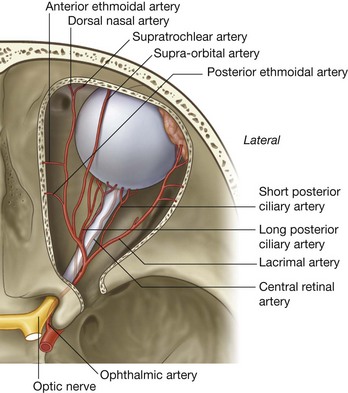
The arterial supply to the lacrimal gland is by branches from the ophthalmic artery and venous drainage is through the ophthalmic veins.
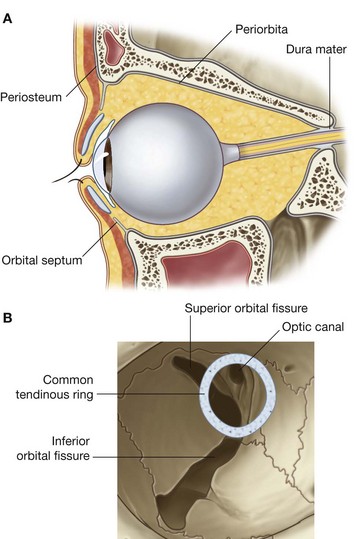
Fissures and foramina
Numerous structures enter and leave the orbit through a variety of openings (Fig. 8.82).
Optic canal
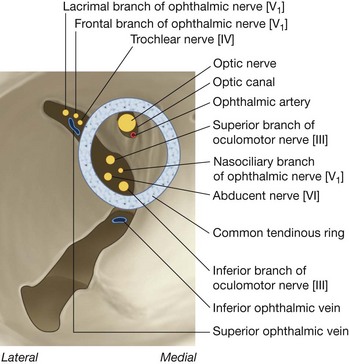
When the bony orbit is viewed from an anterolateral position, the round opening at the apex of the pyramidal-shaped orbit is the optic canal, which opens into the middle cranial fossa and is bounded medially by the body of the sphenoid and laterally by the lesser wing of the sphenoid. Passing through the optic canal are the optic nerve and the ophthalmic artery (Fig. 8.83).
Superior orbital fissure
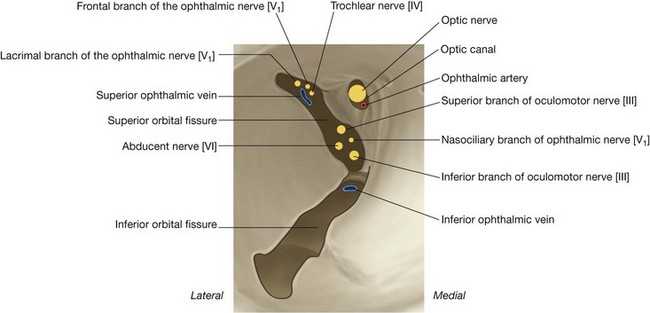
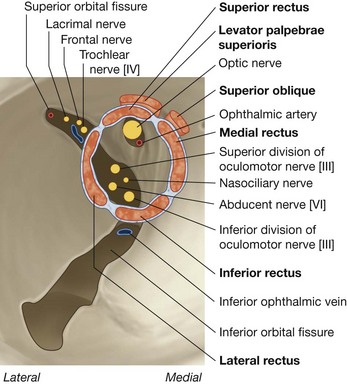
Just lateral to the optic canal is a triangular-shaped gap between the roof and lateral wall of the bony orbit. This is the superior orbital fissure and allows structures to pass between the orbit and the middle cranial fossa (Fig. 8.82).
Passing through the superior orbital fissure are the superior and inferior branches of the oculomotor nerve [III], the trochlear nerve [IV], the abducent nerve [VI], the lacrimal, frontal, and nasociliary branches of the ophthalmic nerve [V1], and the superior ophthalmic vein (Fig. 8.83).
Inferior orbital fissure
Separating the lateral wall of the orbit from the floor of the orbit is a longitudinal opening, the inferior orbital fissure (Fig. 8.82). Its borders are the greater wing of the sphenoid and the maxilla, palatine, and zygomatic bones. This long fissure allows communication between:
the orbit and the pteryatine fossa posteriorly;
the orbit and the infratemporal fossa in the middle; and
the orbit and the temporal fossa posterolaterally.
Passing through the inferior orbital fissure are the maxillary nerve [V2] and its zygomatic branch, the infra-orbital vessels, and a vein communicating with the pterygoid plexus of veins.
Infra-orbital foramen
Beginning posteriorly and crossing about two-thirds of the inferior orbital fissure, a groove (the infra-orbital groove) is encountered, which continues anteriorly across the floor of the orbit (Fig. 8.82). This groove connects with the infra-orbital canal that opens onto the face at the infra-orbital foramen.
The infra-orbital nerve, a branch of the maxillary nerve [V2], and vessels pass through this structure as they exit onto the face.
Other openings
Associated with the medial wall of the bony orbit are several smaller openings (Fig. 8.82).
The anterior and posterior ethmoidal foramina are at the junction between the superior and medial walls. These openings provide exits from the orbit into the ethmoid bone for the anterior and posterior ethmoidal nerves and vessels.
Completing the openings on the medial wall is a canal in the lower part of the wall anteriorly. Clearly visible is the depression for the lacrimal sac formed by the lacrimal bone and the frontal process of the maxilla. This depression is continuous with the nasolacrimal canal, which leads to the inferior nasal meatus. Contained within the nasolacrimal canal is the nasolacrimal duct, a part of the lacrimal apparatus.
Fascial specializations
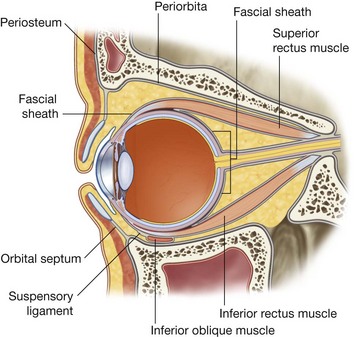
Periorbita
The periosteum lining the bones that form the orbit is the periorbita (Fig. 8.84). It is continuous at the margins of the orbit with the periosteum on the outer surface of the skull and sends extensions into the upper and lower eyelids (the orbital septa).
At the various openings where the orbit communicates with the cranial cavity the periorbita is continuous with the periosteal layer of dura mater. In the posterior part of the orbit, the periorbita thickens around the optic canal and the central part of the superior orbital fissure. This is the point of origin of the four rectus muscles and is the common tendinous ring.
Fascial sheath of the eyeball
The fascial sheath of the eyeball (bulbar sheath) is a layer of fascia that encloses a major part of the eyeball (Figs. 8.85 and 8.86):
posteriorly, it is firmly attached to the sclera (the white part of the eyeball) around the point of entrance of the optic nerve into the eyeball;
anteriorly, it is firmly attached to the sclera near the edge of the cornea (the clear part of the eyeball);
additionally, as the muscles approach the eyeball, the investing fascia surrounding each muscle blends with the fascial sheath of the eyeball as the muscles pass through and continue to their point of attachment.
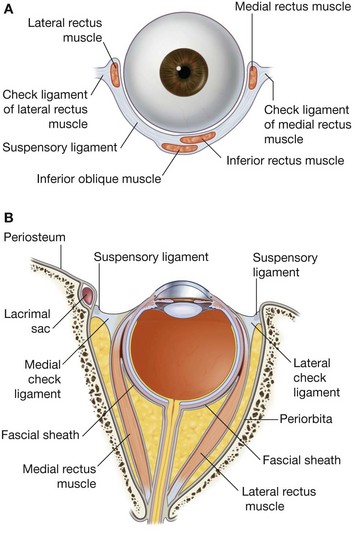
A specialized lower part of the fascial sheath of the eyeball is the suspensory ligament (Figs. 8.85 and 8.86), which supports the eyeball. This “sling-like” structure is made up of the fascial sheath of the eyeball and contributions from the two inferior ocular muscles and the medial and lateral ocular muscles.
Check ligaments of the medial and lateral rectus muscles
Other fascial specialization in the orbit are the check ligaments (Fig. 8.86). These are expansions of the investing fascia covering the medial and lateral rectus muscles, which attach to the medial and lateral walls of the bony orbit:
the medial check ligament is an extension from the fascia covering the medial rectus muscle and attaches immediately posterior to the posterior lacrimal crest of the lacrimal bone;
the lateral check ligament is an extension from the fascia covering the lateral rectus muscle and is attached to the orbital tubercle of the zygomatic bone.
Functionally, the positioning of these ligaments seems to restrict the medial and lateral rectus muscles, thus the names of the fascial specializations.
Muscles
There are two groups of muscles within the orbit:
extrinsic muscles of eyeball (
extra-ocular muscles) involved in movements of the eyeball or raising upper eyelids;
intrinsic muscles within the eyeball, which control the shape of the lens and size of the pupil.
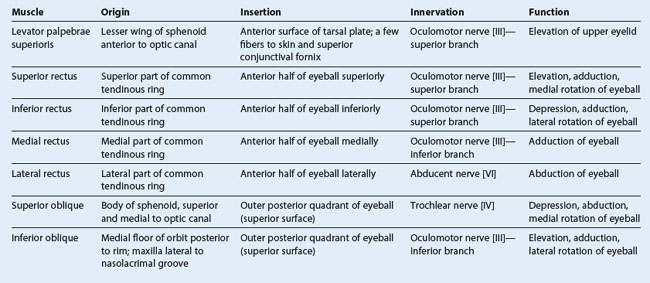
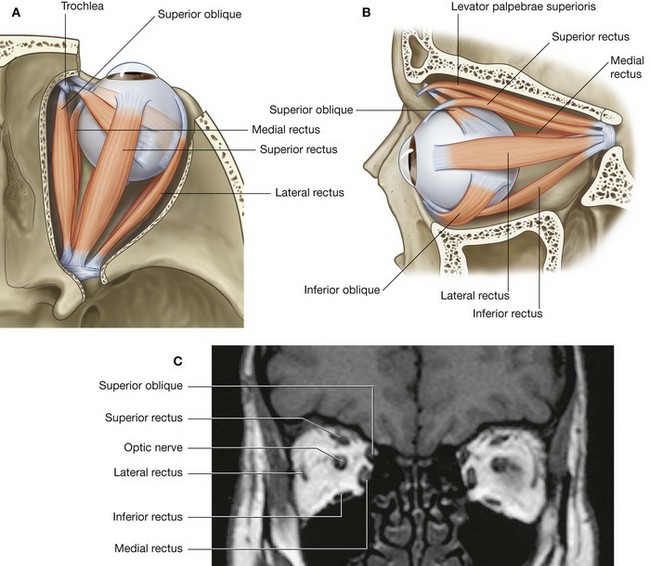
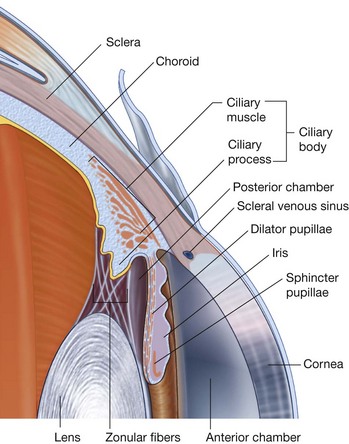
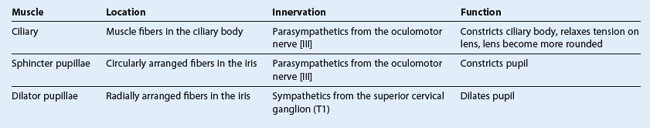
The extrinsic muscles include the levator palpebrae superioris, superior rectus, inferior rectus, medial rectus, lateral rectus, superior oblique, and inferior oblique.
The intrinsic muscles include the ciliary muscle, the sphincter pupillae, and the dilator pupillae.
Extrinsic muscles
Of the seven muscles in the extrinsic group of muscles, one raises the eyelids, whereas the other six move the eyeball itself (Table 8.8).
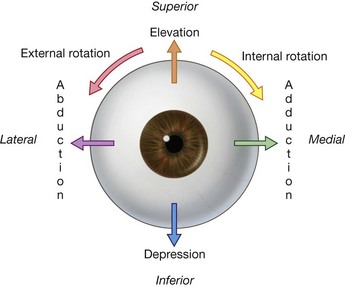
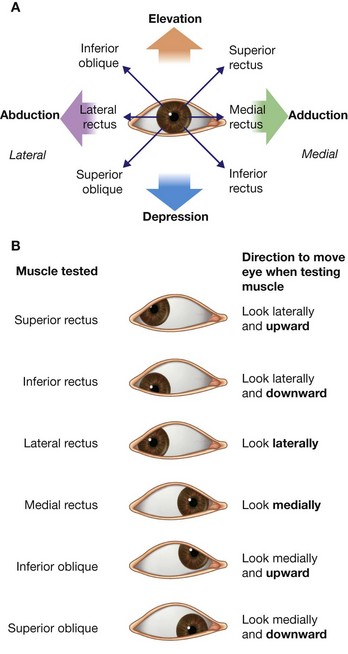
The movements of the eyeball, in three dimensions, (Fig. 8.87) are:
elevation—moving the pupil superiorly;
depression—moving the pupil inferiorly;
abduction—moving the pupil laterally;
adduction—moving the pupil medially;
internal rotation (intorsion)—rotating the upper part of the pupil medially (or toward the nose); and
external rotation (extorsion)—rotating the upper part of the pupil laterally (or toward the temple).
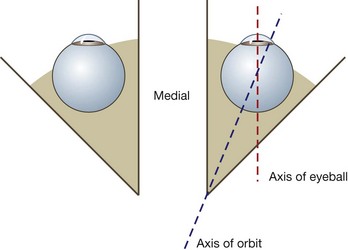
The axis of each orbit is directed slightly laterally from back to front, but each eyeball is directed anteriorly (Fig. 8.88). Therefore the pull of some muscles has multiple effects on the movement of the eyeball, whereas that of others has a single effect.
Levator palpebrae superioris
Levator palpebrae superioris raises the upper eyelid (Table 8.8). It is the most superior muscle in the orbit, originating from the roof, just anterior to the optic canal on the inferior surface of the lesser wing of the sphenoid (Fig. 8.89B). Its primary point of insertion is into the anterior surface of the superior tarsus, but a few fibers also attach to the skin of the upper eyelid and the superior conjunctival fornix.
Innervation is by the superior branch of the oculomotor nerve [III].
Contraction of the levator palpebrae superioris raises the upper eyelid.
A unique feature of levator palpebrae superioris is that a collection of smooth muscle fibers passes from its inferior surface to the upper edge of the superior tarsus (Fig. 8.71). This group of smooth muscle fibers (the superior tarsal muscle) help maintain eyelid elevation and are innervated by postganglionic sympathetic fibers from the superior cervical ganglion.
Loss of oculomotor nerve [III] function results in complete ptosis or drooping of the superior eyelid, whereas loss of sympathetic innervation to the superior tarsal muscle results in partial ptosis.
Rectus muscles
Four rectus muscles occupy medial, lateral, inferior, and superior positions as they pass from their origins posteriorly to their points of attachment on the anterior half of the eyeball (Fig. 8.89 and Table 8.8). They originate as a group from a common tendinous ring at the apex of the orbit and form a cone of muscles as they pass forward to their attachment on the eyeball.
Superior and inferior rectus muscles
The superior and inferior rectus muscles have complicated actions because the apex of the orbit, where the muscles originate, is medial to the central axis of the eyeball when looking directly forward:
the
superior rectus originates from the superior part of the common tendinous ring above the optic canal;
the
inferior rectus originates from the inferior part of the common tendinous ring below the optic canal (
Fig. 8.90).
As these muscles pass forward in the orbit to attach to the anterior half of the eyeball, they are also directed laterally (Fig. 8.89). Because of these orientations:
contraction of the superior rectus elevates, adducts, and internally rotates the eyeball (
Fig. 8.91A);
contraction of the inferior rectus depresses, adducts, and externally rotates the eyeball (
Fig. 8.91A).
The superior branch of the oculomotor nerve [III] innervates the superior rectus, and the inferior branch of the oculomotor nerve [III] innervates the inferior rectus.
To isolate the function of and to test the superior and inferior rectus muscles, a patient is asked to track a physician’s finger laterally and then either upward or downward (Fig. 8.91B). The first movement brings the axis of the eyeball into alignment with the long axis of the superior and inferior rectus muscles. Moving the finger upward tests the superior rectus muscle and moving it downward tests the inferior rectus muscle (Fig. 8.91B).
Medial and lateral rectus muscles
The orientation and actions of the medial and lateral rectus muscles are more straightforward than those of the superior and inferior rectus muscles.
The medial rectus originates from the medial part of the common tendinous ring medial to and below the optic canal, whereas the lateral rectus originates from the lateral part of the common tendinous ring as the common tendinous ring bridges the superior orbital fissure (Fig. 8.90).
The medial and lateral rectus muscles pass forward and attach to the anterior half of the eyeball (Fig. 8.89). Contraction of each medial rectus adducts the eyeball, whereas contraction of each lateral rectus abducts the eyeball (Fig. 8.91A).
The inferior branch of the oculomotor nerve [III] innervates the medial rectus, and the abducent nerve [VI] innervates the lateral rectus.
To isolate the function of and test the medial and lateral rectus muscles, a patient is asked to track a physician’s finger medially and laterally, respectively, in the horizontal plane (Fig. 8.91B).
Oblique muscles
The oblique muscles are in the superior and inferior parts of the orbit, do not originate from the common tendinous ring, are angular in their approaches to the eyeball, and, unlike the rectus muscles, attach to the posterior half of the eyeball (Table 8.8).
Superior oblique
The superior oblique arises from the body of the sphenoid, superior and medial to the optic canal and medial to the origin of the levator palpebrae superioris (Figs. 8.89 and 8.90). It passes forward, along the medial border of the roof of the orbit, until it reaches a fibrocartilaginous pulley (the trochlea), which is attached to the trochlear fovea of the frontal bone.
The tendon of superior oblique passes through the trochlea and turns laterally to cross the eyeball in a posterolateral direction. It continues deep to the superior rectus muscle and inserts into the outer posterior quadrant of the eyeball.
Contraction of the superior oblique therefore directs the pupil down and out (Fig. 8.91A).
The trochlear nerve [IV] innervates the superior oblique along its superior surface.
To isolate the function of and to test the superior oblique muscle, a patient is asked to track a physician’s finger medially to bring the axis of the tendon of the muscle into alignment with the axis of the eyeball, and then to look down, which tests the muscle (Fig. 8.91B).
Inferior oblique
The inferior oblique is the only extrinsic muscle that does not take origin from the posterior part of the orbit. It arises from the medial side of the floor of the orbit, just posterior to the orbital rim, and is attached to the orbital surface of the maxilla just lateral to the nasolacrimal groove (Fig. 8.89).
The inferior oblique crosses the floor of the orbit in a posterolateral direction between the inferior rectus and the floor of the orbit, before inserting into the outer posterior quadrant just under the lateral rectus.
Contraction of the inferior oblique directs the pupil up and out (Fig. 8.91A).
The inferior branch of the oculomotor nerve innervates the inferior oblique.
To isolate the function of and to test the inferior oblique muscle, a patient is asked to track a physician’s finger medially to bring the axis of the eyeball into alignment with the axis of the muscle and then to look up, which tests the muscle (Fig. 8.91B).
Extrinsic muscles and eyeball movements
Six of the seven extrinsic muscles of the orbit are directly involved in movements of the eyeball.
For each of the rectus muscles, the medial, lateral, inferior, and superior, and the superior and inferior obliques, a specific action or group of actions can be described (Table 8.8). However, these muscles do not act in isolation. They work as teams of muscles in the coordinated movement of the eyeball to position the pupil as needed.
For example, although the lateral rectus is the muscle primarily responsible for moving the eyeball laterally, it is assisted in this action by the superior and inferior oblique muscles.
In the clinic
Examination of the eye
Examination of the eye includes assessment of the visual capabilities, the extrinsic musculature and its function, and disease processes that may affect the eye in isolation or as part of the systemic process.
Examination of the eye includes tests for visual acuity, astigmatism, visual fields, and color interpretation (to exclude color blindness) in a variety of circumstances. The physician also assesses the retina, the optic nerve and its coverings, the lens, and the cornea.
The extrinsic muscles are supplied by the abducent nerve [VI], the trochlear nerve [IV], and the oculomotor nerve [III].
The extrinsic muscles work synergistically to provide appropriate and conjugate eye movement:
lateral rectus—abducent nerve [VI];
superior oblique—trochlear nerve [IV];
remainder—oculomotor nerve [III].
The eye may be affected in systemic diseases. Diabetes mellitus typically affects the eye and may cause cataracts, macular disease, and retinal hemorrhage, all impairing vision.
Occasionally unilateral paralysis of the extra-ocular muscles occurs, and is due to brainstem injury or direct nerve injury, which may be associated with tumor compression or trauma. The paralysis of a muscle is easily demonstrated when the patient attempts to move the eye in the direction associated with normal action of that muscle. Typically the patient complains of double vision (diplopia).
Loss of innervation of the muscles around the eye
Loss of innervation of the orbicularis oculi by the facial nerve [VII] causes an inability to close the eyelids tightly, allowing the lower eyelid to droop away causing spillage of tears. This loss of tears allows drying of the conjunctiva, which may ulcerate, so allowing secondary infection.
Loss of innervation of the levator palpebrae superioris by oculomotor nerve [III] damage causes an inability of the superior eyelid to elevate, producing a ptosis. Usually, oculomotor nerve [III] damage is caused by severe head injury.
Loss of innervation of the superior tarsal muscle by sympathetic fibers causes a constant partial ptosis. Any lesion along the sympathetic trunk can induce this. An apical pulmonary malignancy should always be suspected because the ptosis may be part of Horner’s syndrome (see p. 863).
Vessels
Arteries
The arterial supply to the structures in the orbit, including the eyeball, is by the ophthalmic artery (Fig. 8.90). This vessel is a branch of the internal carotid artery, given off immediately after the internal carotid artery leaves the cavernous sinus. The ophthalmic artery passes into the orbit through the optic canal with the optic nerve.
In the orbit the ophthalmic artery initially lies inferior and lateral to the optic nerve (Fig. 8.92). As it passes forward in the orbit, it crosses superior to the optic nerve and proceeds anteriorly on the medial side of the orbit.
In the orbit the ophthalmic artery gives off numerous branches as follows:
the
lacrimal artery, which arises from the ophthalmic artery on the lateral side of the optic nerve, and passes anteriorly on the lateral side of the orbit, supplying the lacrimal gland, muscles, the anterior ciliary branch to the eyeball, and the lateral sides of the eyelid;
the central retinal artery, which enters the optic nerve, proceeds down the center of the nerve to the retina, and is clearly seen when viewing the retina with an ophthalmoscope—occlusion of this vessel or of the parent artery leads to blindness;
the
long and
short posterior ciliary arteries, which are branches that enter the eyeball posteriorly, piercing the sclera, and supplying structures inside the eyeball;
the
muscular arteries, which are branches supplying the intrinsic muscles of the eyeball;
the
supra-orbital artery, which usually arises from the ophthalmic artery immediately after it has crossed the optic nerve, proceeds anteriorly, and exits the orbit through the supra-orbital foramen with the supra-orbital nerve—it supplies the forehead and scalp as it passes across these areas to the vertex of the skull;
the
posterior ethmoidal artery, which exits the orbit through the posterior ethmoidal foramen to supply the ethmoidal cells and nasal cavity;
the
anterior ethmoidal artery, which exits the orbit through the anterior ethmoidal foramen, enters the cranial cavity giving off the anterior meningeal branch, and continues into the nasal cavity supplying the septum and lateral wall, and ending as the dorsal nasal artery;
the
medial palpebral arteries, which are small branches supplying the medial area of the upper and lower eyelids;
the
dorsal nasal artery, which is one of the two terminal branches of the ophthalmic artery, leaves the orbit to supply the upper surface of the nose;
the
supratrochlear artery, which is the other terminal branch of the ophthalmic artery and leaves the orbit with the supratrochlear nerve, supplying the forehead as it passes across it in a superior direction.
Veins
There are two venous channels in the orbit, the superior and inferior ophthalmic veins (Fig. 8.93).
The superior ophthalmic vein begins in the anterior area of the orbit as connecting veins from the supra-orbital vein and the angular vein join together. It passes across the superior part of the orbit, receiving tributaries from the companion veins to the branches of the ophthalmic artery and veins draining the posterior part of the eyeball. Posteriorly, it leaves the orbit through the superior orbital fissure and enters the cavernous sinus.
The inferior ophthalmic vein is smaller than the superior ophthalmic vein, begins anteriorly, and passes across the inferior part of the orbit. It receives various tributaries from muscles and the posterior part of the eyeball as it crosses the orbit.
The inferior ophthalmic vein leaves the orbit posteriorly by:
joining with the superior ophthalmic vein;
passing through the superior orbital fissure on its own to join the cavernous sinus; or
passing through the inferior orbital fissure to join with the pterygoid plexus of veins in the infratemporal fossa.
Because the ophthalmic veins communicate with the cavernous sinus, they act as a route by which infections can spread from outside to inside the cranial cavity.
Innervation
Numerous nerves pass into the orbit and innervate structures within its bony walls. They include the optic nerve [II], the oculomotor nerve [III], the trochlear nerve [IV], the abducent nerve [VI], and autonomic nerves. Other nerves such as the ophthalmic nerve [V1] innervate orbital structures and then travel out of the orbit to innervate other regions.
Optic nerve
The optic nerve [II] is not a true cranial nerve, but rather an extension of the brain carrying afferent fibers from the retina of the eyeball to the visual centers of the brain.
The optic nerve is surrounded by the cranial meninges, including the subarachnoid space, which extend as far forward as the eyeball.
Any increase in intracranial pressure therefore results in increased pressure in the subarachnoid space surrounding the optic nerve. This may impede venous return along the retinal veins, causing edema of the optic disc (papilledema), which can be seen when the retina is examined using an ophthalmoscope.
The optic nerve leaves the orbit through the optic canal (Fig. 8.94). It is accompanied in the optic canal by the ophthalmic artery.
Oculomotor nerve
The oculomotor nerve [III] leaves the anterior surface of the brainstem between the midbrain and the pons. It passes forward in the lateral wall of the cavernous sinus.
Just before entering the orbit the oculomotor nerve [III] divides into superior and inferior branches (Fig. 8.95). These branches enter the orbit through the superior orbital fissure, lying within the common tendinous ring (Fig. 8.94).
Inside the orbit the small superior branch passes upward over the lateral side of the optic nerve to innervate the superior rectus and levator palpebrae superioris muscles (Fig. 8.95).
The large inferior branch divides into three branches:
one passing below the optic nerve as it passes to the medial side of the orbit to innervate the medial rectus muscle;
a second descending to innervate the inferior rectus muscle;
the third descends as it runs forward along the floor of the orbit to innervate the inferior oblique muscle (
Fig. 8.95).
As the third branch descends, it gives off the branch to the ciliary ganglion. This is the parasympathetic root to the ciliary ganglion and carries preganglionic parasympathetic fibers that will synapse in the ciliary ganglion with postganglionic parasympathetic fibers. The postganglionic fibers are distributed to the eyeball through short ciliary nerves and innervate the sphincter pupillae and ciliary muscles.
Trochlear nerve
The trochlear nerve [IV] arises from the posterior surface of the midbrain, and passes around the midbrain to enter the edge of the tentorium cerebelli. It continues on an intradural path arriving in and passing through the lateral wall of the cavernous sinus just below the oculomotor nerve [III].
Just before entering the orbit, the trochlear nerve ascends, passing across the oculomotor nerve [III] and enters the orbit through the superior orbital fissure above the common tendinous ring (Fig. 8.94). In the orbit the trochlear nerve [IV] ascends and turns medially, crossing above the levator palpebrae superioris muscle to enter the upper border of the superior oblique muscle (Fig. 8.96).
Abducent nerve
The abducent nerve [VI] arises from the brainstem between the pons and medulla. It enters the dura covering the clivus and continues in a dural canal until it reaches the cavernous sinus.
The abducent nerve enters the cavernous sinus and runs through the sinus lateral to the internal carotid artery. It passes out of the sinus and enters the orbit through the superior orbital fissure within the common tendinous ring (Fig. 8.94). Once in the orbit it courses laterally to supply the lateral rectus muscle.
Postganglionic sympathetic fibers
Preganglionic sympathetic fibers arise from the upper segments of the thoracic spinal cord, mainly T1. They enter the sympathetic chain through white rami communicantes, and ascend to the superior cervical ganglion where they synapse with postganglionic sympathetic fibers.
The postganglionic fibers are distributed along the internal carotid artery and its branches.
The postganglionic sympathetic fibers destined for the orbit travel with the ophthalmic artery. Once in the orbit the fibers are distributed to the eyeball either by:
passing through the ciliary ganglion, without synapsing, and joining the short ciliary nerves, which pass from the ganglion to the eyeball; or
passing through long ciliary nerves to reach the eyeball.
In the eyeball postganglionic sympathetic fibers innervate the dilator pupillae muscle.
Ophthalmic nerve [V1]
The ophthalmic nerve [V1] is the smallest and most superior of the three divisions of the trigeminal nerve. This purely sensory nerve receives input from structures in the orbit and from additional branches on the face and scalp.
Leaving the trigeminal ganglion, the ophthalmic nerve [V1] passes forward in the lateral wall of the cavernous sinus inferior to the trochlear [IV] and oculomotor [III] nerves. Just before it enters the orbit it divides into three branches—the nasociliary, lacrimal, and frontal nerves (Fig. 8.97). These branches enter the orbit through the superior orbital fissure with the frontal and lacrimal nerves outside the common tendinous ring, and the nasociliary nerve within the common tendinous ring (Fig. 8.94).
Lacrimal nerve
The lacrimal nerve is the smallest of the three branches of the ophthalmic nerve [V1]. Once in the orbit it passes forward along the upper border of the lateral rectus muscle (Fig. 8.98). It receives a branch from the zygomaticotemporal nerve, which carries parasympathetic and sympathetic postganglionic fibers for distribution to the lacrimal gland.
Reaching the anterolateral aspect of the orbit, the lacrimal nerve supplies the lacrimal gland, conjunctiva, and lateral part of the upper eyelid.
Frontal nerve
The frontal nerve is the largest branch of the ophthalmic nerve [V1] and receives sensory input from areas outside the orbit. Exiting the superior orbital fissure, this branch passes forward between the levator palpebrae superioris and the periorbita on the roof of the orbit (Fig. 8.94). About midway across the orbit it divides into its two terminal branches—the supra-orbital and supratrochlear nerves (Figs. 8.97 and 8.98):
the supratrochlear nerve continues forward in an anteromedial direction, passing above the trochlea, exits the orbit medial to the supra-orbital foramen, and supplies the conjunctiva and skin of the upper eyelid and the skin on the lower medial part of the forehead;
the supra-orbital nerve is the larger of the two branches, continues forward, passing between the levator palpebrae superioris muscle and the periorbita covering the roof of the orbit (
Fig. 8.98), exits the orbit through the supra-orbital notch and ascends across the forehead and scalp, supplying the upper eyelid and conjunctiva, the forehead, and as far posteriorly as the middle of the scalp.
Nasociliary nerve
The nasociliary nerve is intermediate in size between the frontal and lacrimal nerves and is usually the first branch from the ophthalmic nerve (Fig. 8.97). It is most deeply placed in the orbit, entering the area within the common tendinous ring between the superior and inferior branches of the oculomotor nerve [III] (Fig. 8.94).
Once in the orbit, the nasociliary nerve crosses the superior surface of the optic nerve as it passes in a medial direction below the superior rectus muscle (Figs. 8.97 and 8.99). Its first branch, the communicating branch with the ciliary ganglion (sensory root to the ciliary ganglion), is given off early in its path through the orbit.
The nasociliary nerve continues forward along the medial wall of the orbit, between the superior oblique and the medial rectus muscles, giving off several branches. These include:
the
long ciliary nerves, which are sensory to the eyeball but may also contain sympathetic fibers for pupillary dilation;
the
posterior ethmoidal nerve, which exits the orbit through the posterior ethmoidal foramen to supply posterior ethmoidal cells and the sphenoidal sinus;
the
infratrochlear nerve, which distributes to the medial part of the upper and lower eyelids, the lacrimal sac, and skin of the upper half of the nose; and
the
anterior ethmoidal nerve, which exits the orbit through the anterior ethmoidal foramen to supply the anterior cranial fossa, nasal cavity, and skin of the lower half of the nose (
Figs. 8.97 and
8.99).
Ciliary ganglion
The ciliary ganglion is a parasympathetic ganglion of the oculomotor nerve [III]. It is associated with the nasociliary branch of the ophthalmic nerve [V1] and is the site where preganglionic and postganglionic parasympathetic neurons synapse as fibers from this part of the autonomic division of the PNS make their way to the eyeball. The ciliary ganglion is also traversed by postganglionic sympathetic fibers and sensory fibers as they travel to the eyeball.
The ciliary ganglion is a very small ganglion, in the posterior part of the orbit immediately lateral to the optic nerve and between the optic nerve and the lateral rectus muscle (Fig. 8.99). It is usually described as receiving at least two, and possibly three, branches or roots from other nerves in the orbit.
Parasympathetic root
As the inferior branch of the oculomotor nerve [III] passes the area of the ciliary ganglion, it sends a branch to the ganglion (the parasympathetic root). The parasympathetic branch carries preganglionic parasympathetic fibers, which enter the ganglion and synapse with postganglionic parasympathetic fibers within the ganglion (Fig. 8.100).
The postganglionic parasympathetic fibers leave the ganglion through short ciliary nerves, which enter the posterior aspect of the eyeball around the optic nerve.
In the eyeball the parasympathetic fibers innervate:
the sphincter pupillae muscle, responsible for pupillary constriction; and
the ciliary muscle, responsible for accommodation of the lens of the eye for near vision.
Sensory root
A second branch (the sensory root), passes from the nasociliary nerve to the ganglion (Fig. 8.100). This branch enters the posterosuperior aspect of the ganglion, and carries sensory fibers, which pass through the ganglion and continue along the short ciliary nerves to the eyeball. These fibers are responsible for sensory innervation to all parts of the eyeball.
Sympathetic root
The third branch to the ciliary ganglion is the most variable. This branch, when present, is the sympathetic root and contains postganglionic sympathetic fibers from the superior cervical ganglion (Fig. 8.100). These fibers travel up the internal carotid artery, leave the plexus surrounding the artery in the cavernous sinus, and enter the orbit through the common tendinous ring. In the orbit they enter the posterior aspect of the ciliary ganglion, cross the ganglion, and continue along the short ciliary nerves to the eyeball.
Sympathetic fibers to the eyeball may not enter the ganglion as a separate branch. The postganglionic sympathetic fibers may leave the plexus associated with the internal carotid artery in the cavernous sinus, join the ophthalmic nerve [V1] and distribute to the ciliary ganglion through the sensory root from the nasociliary nerve.
Whatever their path, postganglionic sympathetic fibers reach the eyeball and innervate the dilator pupillae muscle.
Eyeball
The globe-shaped eyeball occupies the anterior part of the orbit. Its rounded shape is disrupted anteriorly, where it bulges outward. This outward projection represents about one-sixth of the total area of the eyeball and is the transparent cornea (Fig. 8.101).
Posterior to the cornea and in order from front to back are the anterior chamber, the iris and pupil, the posterior chamber, the lens, the postremal (vitreous) chamber, and the retina.
Vascular layer of the eyeball
The vascular layer of the eyeball consists of three continuous parts—the choroid, the ciliary body, and the iris from posterior to anterior (Fig. 8.101).
Choroid
The choroid is posterior and represents approximately two-thirds of the vascular layer. It is a thin, highly vascular, pigmented layer consisting of smaller vessels adjacent to the retina and larger vessels more peripherally. It is firmly attached to the retina internally and loosely attached to the sclera externally.
Ciliary body
Extending from the anterior border of the choroid is the ciliary body (Fig. 8.101). This triangular-shaped structure, between the choroid and the iris, forms a complete ring around the eyeball. Its components include the ciliary muscle and the ciliary processes (Fig. 8.103).
The ciliary muscle consists of smooth muscle fibers arranged longitudinally, circularly, and radially. Controlled by parasympathetics traveling to the orbit in the oculomotor nerve [III], these muscle fibers, on contraction, decrease the size of the ring formed by the ciliary body.
The ciliary processes are longitudinal ridges projecting from the inner surface of the ciliary body (Fig. 8.103). Extending from them are zonular fibers attached to the lens of the eyeball, which suspend the lens in its proper position and collectively form the suspensory ligament of the lens.
Contraction of the ciliary muscle decreases the size of the ring formed by the ciliary body. This reduces tension on the suspensory ligament of the lens. The lens therefore becomes more rounded (relaxed) resulting in accommodation of the lens for near vision.
Ciliary processes also contribute to the formation of aqueous humor.
Iris
Completing the vascular layer of the eyeball anteriorly is the iris (Fig. 8.101). This circular structure, projecting outward from the ciliary body, is the colored part of the eye with a central opening (the pupil). Controlling the size of the pupil are smooth muscle fibers within the iris (Fig. 8.103):
fibers arranged in a circular pattern make up the
sphincter pupillae muscle (
Table 8.9), which is innervated by parasympathetics—contraction of its fibers decreases or constricts the pupillary opening;
fibers arranged in a radial pattern make up the
dilator pupillae muscle, which is innervated by sympathetics—contraction of its fibers increases or dilates the pupillary opening.
EAR
The ear is the organ of hearing and balance. It has three parts (Fig. 8.104):
the first part is the
external ear consisting of the part attached to the lateral aspect of the head and the canal leading inward;
the second part is the
middle ear—a cavity in the petrous part of the temporal bone bounded laterally, and separated from the external canal, by a membrane and connected internally to the pharynx by a narrow tube;
the third part is the
internal ear consisting of a series of cavities within the petrous part of the temporal bone between the middle ear laterally and the internal acoustic meatus medially.
The internal ear converts the mechanical signals received from the middle ear, which start as sound captured by the external ear, into electrical signals to transfer information to the brain. The internal ear also contains receptors that detect motion and position.
External ear
The external ear consists of two parts. The part projecting from the side of the head is the auricle (pinna) and the canal leading inward is the external acoustic meatus.
Auricle
The auricle is on the side of the head and assists in capturing sound. It consists of cartilage covered with skin and arranged in a pattern of various elevations and depressions (Fig. 8.105).
The large outside rim of the auricle is the helix. It ends inferiorly at the fleshy lobule, the only part of the auricle not supported by cartilage.
The hollow center of the auricle is the concha of auricle. The external acoustic meatus leaves from the depths of this area.
Just anterior to the opening of the external acoustic meatus, in front of the concha, is an elevation (the tragus). Opposite the tragus, and above the fleshy lobule, is another elevation (the antitragus). A smaller curved rim, parallel and anterior to the helix is the antihelix.
Muscles
Numerous intrinsic and extrinsic muscles are associated with the auricle:
the intrinsic muscles pass between the cartilaginous parts of the auricle and may change the shape of the auricle;
the extrinsic muscles, the anterior, superior, and posterior auricular muscles, pass from the scalp or skull to the auricle and may also play a role in positioning of the auricle.
Both groups of muscles are innervated by the facial nerve [VII].
Innervation
Sensory innervation of the auricle is from many sources (Fig. 8.106):
the outer more superficial surfaces of the auricle are supplied by the great auricular nerve (anterior and posterior inferior portions) and the lesser occipital nerve (posterior superior portion) from the cervical plexus and the auriculotemporal branch of the mandibular nerve [V
3] (anterior superior portion);
the deeper parts of the auricle are supplied by the vagus nerve [X] (the auricular branch) and the facial nerve [VII] (which sends a branch to the auricular branch of the vagus nerve [X]).
Vessels
The arterial supply to the auricle is from numerous sources. The external carotid artery supplies the posterior auricular artery, the superficial temporal artery supplies anterior auricular branches, and the occipital artery supplies a branch.
Venous drainage is through vessels following the arteries.
Lymphatic drainage of the auricle passes anteriorly into parotid nodes and posteriorly into mastoid nodes, and possibly into the upper deep cervical nodes.
External acoustic meatus
The external acoustic meatus extends from the deepest part of the concha to the tympanic membrane (eardrum), a distance of approximately 1 inch (2.5 cm) (Fig. 8.107). Its walls consist of cartilage and bone. The lateral one-third is formed from cartilaginous extensions from some of the auricular cartilages and the medial two-thirds is a bony tunnel in the temporal bone.
Throughout its length the external acoustic meatus is covered with skin, some of which contains hair and modified sweat glands producing cerumen (earwax). Its diameter varies, being wider laterally and narrow medially.
The external acoustic meatus does not follow a straight course. From the external opening it passes upward in an anterior direction, then turns slightly posteriorly still passing upward, and finally, turns again in an anterior direction with a slight descent. For examination purposes, observation of the external acoustic meatus and tympanic membrane can be improved by pulling the ear superiorly, posteriorly, and slightly laterally.
Innervation
Sensory innervation of the external acoustic meatus is from several of the cranial nerves. The major sensory input travels through branches of the auriculotemporal nerve, a branch of the mandibular nerve [V3] (anterior and superior walls), and in the auricular branch of the vagus nerve [X] (posterior and inferior walls). A minor sensory input may also come from a branch of the facial nerve [VII] to the auricular branch of the vagus nerve [X].
Tympanic membrane
The tympanic membrane separates the external acoustic meatus from the middle ear (Figs. 8.108 and 8.109). It is at an angle, sloping medially from top to bottom and posteriorly to anteriorly. Its lateral surface therefore faces inferiorly and anteriorly. It consists of a connective tissue core lined with skin on the outside and mucous membrane on the inside.
Around the periphery of the tympanic membrane a fibrocartilaginous ring attaches it to the tympanic part of the temporal bone. At its center, a concavity is produced by the attachment on its internal surface of the lower end of the handle of malleus, part of the malleus bone in the middle ear. This point of attachment is the umbo of tympanic membrane.
Anteroinferior to the umbo of tympanic membrane a bright reflection of light, referred to as the cone of light, is usually visible when examining the tympanic membrane with an otoscope.
Superior to the umbo in an anterior direction is the attachment of the rest of the handle of malleus (Fig. 8.109). At the most superior extent of this line of attachment a small bulge in the membrane marks the position of the lateral process of the malleus as it projects against the internal surface of the tympanic membrane. Extending away from this elevation, on the internal surface of the membrane, are the anterior and posterior malleolar folds. Superior to these folds the tympanic membrane is thin and slack (the pars flaccida), whereas the rest of the membrane is thick and taut (the pars tensa).
Innervation
Innervation of the external and internal surfaces of the tympanic membrane is by several cranial nerves:
sensory innervation of the skin on the outer surface of the tympanic membrane is primarily by the auriculotemporal nerve, a branch of the mandibular nerve [V
3] with additional participation of the auricular branch of the vagus nerve [X], a small contribution by a branch of the facial nerve [VII] to the auricular branch of the vagus nerve [X], and possibly a contribution from the glossopharyngeal nerve [IX];
sensory innervation of the mucous membrane on the inner surface of the tympanic membrane is carried entirely by the glossopharyngeal [IX] nerve.
In the clinic
Examination of the ear
The ear comprises three components—the external, middle, and internal ear.
Clinical examination is carried out to assess hearing and balance. Further examination involves use of an otoscope or other imaging techniques.
External ear
The external ear is easily examined. The external acoustic meatus and the tympanic membrane require otoscopic examination (Fig. 8.109B). An otoscope is a device through which light can be shone and the image magnified to inspect the external acoustic meatus and the tympanic membrane.
The examination begins by grasping the posterosuperior aspect of the ear and gently retracting it to straighten the external auditory meatus. The normal tympanic membrane is relatively translucent and has a gray–reddish tinge. The handle of the malleus is visible near the center of the membrane. In the 5 o’clock position a cone of light is always demonstrated.
Middle and inner ears
The middle ear is investigated by CT and MRI to visualize the malleus, incus, and stapes. The relationship of these bones to the middle ear cavity is determined and any masses identified.
The inner ear is also assessed by CT and MRI.
In the clinic
Tympanic membrane perforation
Although perforation of the tympanic membrane (eardrum) has many causes, trauma infection is still one of the commonest causes today.
Otitis media (infection of the middle ear) is common, but can usually be treated by antibiotics. If the infection persists, the chronic inflammatory change may damage the ossicular chain and other structures within the middle ear to produce deafness.
Most ruptures of the tympanic membrane tend to heal spontaneously, but surgical intervention may be necessary if the rupture is large.
From time to time it is necessary to enter the middle ear through the tympanic membrane. As the chorda tympani runs in the upper one-third of the tympanic membrane, incisions are always below this level. The richer blood supply to the posterior aspect of the tympanic membrane determines the standard surgical approach in the posterior-inferior aspect.
Middle ear
The middle ear is an air-filled, mucous membrane–lined space in the temporal bone between the tympanic membrane laterally and the lateral wall of the internal ear medially. It is described as consisting of two parts (Fig. 8.110):
the
tympanic cavity immediately adjacent to the tympanic membrane;
the
epitympanic recess superiorly.
The middle ear communicates with the mastoid area posteriorly and the nasopharynx (via the pharyngotympanic tube) anteriorly. Its basic function is to transmit vibrations of the tympanic membrane across the cavity of the middle ear to the internal ear. It accomplishes this through three interconnected but movable bones that bridge the space between the tympanic membrane and the internal ear. These bones are the malleus (connected to the tympanic membrane), the incus (connected to the malleus by a synovial joint), and the stapes (connected to the incus by a synovial joint, and attached to the lateral wall of the internal ear at the oval window).
Boundaries
The middle ear has a roof and a floor, and anterior, posterior, medial, and lateral walls (Fig. 8.111).
Tegmental wall
The tegmental wall (roof) of the middle ear consists of a thin layer of bone, which separates the middle ear from the middle cranial fossa. This layer of bone is the tegmen tympani on the anterior surface of the petrous part of the temporal bone.
Jugular wall
The jugular wall (floor) of the middle ear consists of a thin layer of bone that separates it from the internal jugular vein. Occasionally, the floor is thickened by the presence of mastoid air cells.
Near the medial border of the floor is a small aperture, through which the tympanic branch from the glossopharyngeal nerve [IX] enters the middle ear.
Membranous wall
The membranous (lateral) wall of the middle ear consists almost entirely of the tympanic membrane, but because the tympanic membrane does not extend superiorly into the epitympanic recess, the upper part of the membranous wall of the middle ear is the bony lateral wall of the epitympanic recess.
Mastoid wall
The mastoid (posterior) wall of the middle ear is only partially complete. The lower part of this wall consists of a bony partition between the tympanic cavity and mastoid air cells. Superiorly, the epitympanic recess is continuous with the aditus to the mastoid antrum (Figs. 8.111 and 8.112).
Associated with the mastoid wall are:
the pyramidal eminence, a small elevation through which the tendon of the stapedius muscle enters the middle ear; and
the opening through which the chorda tympani nerve, a branch of the facial nerve [VII], enters the middle ear.
Anterior wall
The anterior wall of the middle ear is only partially complete. The lower part consists of a thin layer of bone that separates the tympanic cavity from the internal carotid artery. Superiorly, the wall is deficient because of the presence of:
a large opening for the entrance of the pharyngotympanic tube into the middle ear; and
a smaller opening for the canal containing the tensor tympani muscle.
The foramen for the exit of the chorda tympani nerve from the middle ear is also associated with this wall (Fig. 8.111).
Labyrinthine wall
The labyrinthine (medial) wall of the middle ear is also the lateral wall of the internal ear. A prominent structure on this wall is a rounded bulge (the promontory) produced by the basal coil of the cochlea, which is an internal ear structure involved with hearing (Fig. 8.111).
Associated with the mucous membrane covering the promontory is a plexus of nerves (the tympanic plexus), which consists primarily of contributions from the tympanic branch of the glossopharyngeal nerve [IX] and branches from the internal carotid plexus. It supplies the mucous membrane of the middle ear, the mastoid area, and the pharyngotympanic tube.
Additionally, a branch of the tympanic plexus (the lesser petrosal nerve) leaves the promontory and the middle ear, travels across the anterior surface of the petrous part of the temporal bone, and leaves the middle cranial fossa through the foramen ovale to enter the otic ganglion. Other structures associated with the labyrinthine wall are two openings, the oval and round windows, and two prominent elevations (Fig. 8.111):
the
oval window is posterosuperior to the promontory, is the point of attachment for the
base of stapes (
footplate), and ends the chain of bones that transfer vibrations initiated by the tympanic membrane to the cochlea of the internal ear;
the
round window is posteroinferior to the promontory;
posterior and superior to the oval window on the medial wall is the
prominence of facial canal, which is a ridge of bone produced by the facial nerve [VII] in its canal as it passes through the temporal bone;
just above and posterior to the prominence of facial canal is a broader ridge of bone (
prominence of lateral semicircular canal) produced by the lateral semicircular canal, which is a structure involved in detecting motion.
Mastoid area
Posterior to the epitympanic recess of the middle ear is the aditus to mastoid antrum, which is the opening to the mastoid antrum (Fig. 8.112).
The mastoid antrum is a cavity continuous with collections of air-filled spaces (the mastoid cells), throughout the mastoid part of the temporal bone, including the mastoid process. The mastoid antrum is separated from the middle cranial fossa above by only the thin tegmen tympani.
The mucous membrane lining the mastoid air cells is continuous with the mucous membrane throughout the middle ear. Therefore infections in the middle ear can easily spread into the mastoid area.
In the clinic
Mastoiditis
Infection within the mastoid antrum and mastoid cells is usually secondary to infection in the middle ear. The mastoid cells provide an excellent culture medium for infection. Infection of the bone (osteomyelitis) may also develop, spreading into the middle cranial fossa.
Drainage of the pus within the mastoid air cells is necessary and there are numerous approaches for doing this. When undertaking this type of surgery, it is extremely important that care is taken not to damage the mastoid wall of the middle ear to prevent injury to the facial nerve [VII]. Any breach of the inner table of the cranial vault may allow bacteria to enter the cranial cavity and meningitis will ensue.
Pharyngotympanic tube
The pharyngotympanic tube connects the middle ear with the nasopharynx (Fig. 8.113) and equalizes pressure on both sides of the tympanic membrane. Its opening in the middle ear is on the anterior wall, and from here it extends forward, medially, and downward to enter the nasopharynx just posterior to the inferior meatus of the nasal cavity. It consists of:
a
bony part (the one-third nearest the middle ear); and
a
cartilaginous part (the remaining two-thirds).
The opening of the bony part is clearly visible on the inferior surface of the skull at the junction of the squamous and petrous parts of the temporal bone immediately posterior to the foramen ovale and foramen spinosum.
Vessels
The arterial supply to the pharyngotympanic tube is from several sources. Branches arise from the ascending pharyngeal artery (a branch of the external carotid artery) and from two branches of the maxillary artery (the middle meningeal artery and the artery of the pterygoid canal).
Venous drainage of the pharyngotympanic tube is to the pterygoid plexus of veins in the infratemporal fossa.
Innervation
Innervation of the mucous membrane lining the pharyngotympanic tube is primarily from the tympanic plexus because it is continuous with the mucous membrane lining the tympanic cavity, the internal surface of the tympanic membrane, and the mastoid antrum and mastoid cells. This plexus receives its major contribution from the tympanic nerve, a branch of the glossopharyngeal nerve [IX].
Auditory ossicles
The bones of the middle ear consist of the malleus, incus, and stapes. They form an osseous chain across the middle ear from the tympanic membrane to the oval window of the internal ear (Fig. 8.114).
Muscles associated with the auditory ossicles modulate movement during the transmission of vibrations.
Malleus
The malleus is the largest of the auditory ossicles and is attached to the tympanic membrane. Identifiable parts include the head of malleus, neck of malleus, anterior and lateral processes, and handle of malleus (Fig. 8.114). The head of malleus is the rounded upper part of the malleus in the epitympanic recess. Its posterior surface articulates with the incus.
Inferior to the head of malleus is the constricted neck of malleus, and below this are the anterior and lateral processes:
the anterior process is attached to the anterior wall of the middle ear by a ligament;
the lateral process is attached to the anterior and posterior malleolar folds of the tympanic membrane.
The downward extension of the malleus, below the anterior and lateral processes, is the handle of malleus, which is attached to the tympanic membrane.
Incus
The second bone in the series of auditory ossicles is the incus. It consists of the body of incus, and long and short limbs (Fig. 8.114):
the enlarged body of incus articulates with the head of malleus and is in the epitympanic recess;
the long limb extends downward from the body, paralleling the handle of the malleus, and ends by bending medially to articulate with the stapes;
the short limb extends posteriorly and is attached by a ligament to the upper posterior wall of the middle ear.
Stapes
The stapes is the most medial bone in the osseous chain and is attached to the oval window. It consists of the head of stapes, anterior and posterior limbs, and the base of stapes (Fig. 8.114):
the head of stapes is directed laterally and articulates with the long process of the incus;
the two limbs separate from each other and attach to the oval base;
the base of stapes fits into the oval window on the labyrinthine wall of the middle ear.
Muscles associated with the ossicles
Two muscles are associated with the bony ossicles of the middle ear—the tensor tympani and stapedius (Fig. 8.115 and Table 8.10).
Tensor tympani
The tensor tympani muscle lies in a bony canal above the pharyngotympanic tube. It originates from the cartilaginous part of the pharyngotympanic tube, the greater wing of the sphenoid, and its own bony canal, and passes through its canal in a posterior direction, ending in a rounded tendon that inserts into the upper part of the handle of the malleus.
Innervation of the tensor tympani is by a branch from the mandibular nerve [V3].
Contraction of the tensor tympani pulls the handle of the malleus medially. This tenses the tympanic membrane, reducing the force of vibrations in response to loud noises.
Stapedius
The stapedius muscle is a very small muscle that originates from inside the pyramidal eminence, which is a small projection on the mastoid wall of the middle ear (Fig. 8.115). Its tendon emerges from the apex of the pyramidal eminence and passes forward to attach to the posterior surface of the neck of stapes.
Stapedius is innervated by a branch from the facial nerve [VII].
Contraction of the stapedius muscle, usually in response to loud noises, pulls the stapes posteriorly and prevents excessive oscillation.
Vessels
Numerous arteries supply the structures in the middle ear:
the two largest branches are the
tympanic branch of the maxillary artery and the
mastoid branch of the occipital or posterior auricular arteries;
smaller branches come from the middle meningeal artery, the ascending pharyngeal artery, the artery of the pterygoid canal, and tympanic branches from the internal carotid artery.
Venous drainage of the middle ear returns to the pterygoid plexus of veins and the superior petrosal sinus.
Innervation
The tympanic plexus innervates the mucous membrane lining the walls and contents of the middle ear, which includes the mastoid area and the pharyngotympanic tube. It is formed by the tympanic nerve, a branch of the glossopharyngeal nerve [IX], and from branches of the internal carotid plexus. The tympanic plexus occurs in the mucous membrane covering the promontory, which is the rounded bulge on the labyrinthine wall of the middle ear (Fig. 8.116).
As the glossopharyngeal nerve [IX] exits the skull through the jugular foramen, it gives off the tympanic nerve. This branch re-enters the skull through a small foramen and passes through the bone to the middle ear.
Once in the middle ear, the tympanic nerve forms the tympanic plexus, along with branches from the plexus of nerves surrounding the internal carotid artery (caroticotympanic nerves). Branches from the tympanic plexus supply the mucous membranes of the middle ear, including the pharyngotympanic tube and the mastoid area.
The tympanic plexus also gives off a major branch (the lesser petrosal nerve), which supplies preganglionic parasympathetic fibers to the otic ganglion (Fig. 8.116).
The lesser petrosal nerve leaves the area of the promontory, exits the middle ear, travels through the petrous part of the temporal bone, and exits onto the anterior surface of the petrous part of the temporal bone through a hiatus just below the hiatus for the greater petrosal nerve (Fig. 8.117). It continues diagonally across the anterior surface of the temporal bone before exiting the middle cranial fossa through the foramen ovale. Once outside the skull it enters the otic ganglion.
Internal ear
The internal ear consists of a series of bony cavities (the bony labyrinth) and membranous ducts and sacs (the membranous labyrinth) within these cavities. All these structures are in the petrous part of the temporal bone between the middle ear laterally and the internal acoustic meatus medially (Figs. 8.118 and 8.119).
The bony labyrinth consists of the vestibule, three semicircular canals, and the cochlea (Fig. 8.119). These bony cavities are lined with periosteum and contain a clear fluid (the perilymph).
Suspended within the perilymph but not filling all spaces of the bony labyrinth is the membranous labyrinth, which consists of the semicircular ducts, the cochlear duct, and two sacs (the utricle and the saccule). These membranous spaces are filled with endolymph.
The structures in the internal ear convey information to the brain about balance and hearing:
the cochlear duct is the organ of hearing;
the semicircular ducts, utricle, and saccule are the organs of balance.
The nerve responsible for these functions is the vestibulocochlear nerve [VIII], which divides into vestibular (balance) and cochlear (hearing) parts after entering the internal acoustic meatus (Fig. 8.119).
Bony labyrinth
The vestibule, which contains the oval window in its lateral wall, is the central part of the bony labyrinth (Fig. 8.120). It communicates anteriorly with the cochlea and posterosuperiorly with the semicircular canals.
A narrow canal (the vestibular aqueduct) leaves the vestibule, and passes through the temporal bone to open on the posterior surface of the petrous part of the temporal bone.
Semicircular canals
Projecting in a posterosuperior direction from the vestibule are the anterior, posterior, and lateral semicircular canals (Fig. 8.120). Each of these canals forms two-thirds of a circle connected at both ends to the vestibule and with one end dilated to form the ampulla. The canals are oriented so that each canal is at right angles to the other two.
Cochlea
Projecting in an anterior direction from the vestibule is the cochlea, which is a bony structure that twists on itself two and one-half to two and three-quarter times around a central column of bone (the modiolus). This arrangement produces a cone-shaped structure with a base of cochlea that faces posteromedially and an apex that faces anterolaterally (Fig. 8.121). This positions the wide base of the modiolus near the internal acoustic meatus, where it is entered by branches of the cochlear part of the vestibulocochlear nerve [VIII].
Extending laterally throughout the length of the modiolus is a thin lamina of bone (the lamina of modiolus, or spiral lamina). Circling around the modiolus, and held in a central position by its attachment to the lamina of modiolus, is the cochlear duct, which is a component of the membranous labyrinth.
Attached peripherally to the outer wall of the cochlea, the cochlear duct creates two canals (the scala vestibuli and the scala tympani), which extend throughout the cochlea and are continuous with each other at the apex through a narrow slit (the helicotrema):
the scala vestibuli is continuous with the vestibule;
the scala tympani is separated from the middle ear by the secondary tympanic membrane covering the round window (
Fig. 8.122).
Finally, near the round window is a small channel (the cochlear canaliculus), which passes through the temporal bone and opens on its inferior surface into the posterior cranial fossa. This provides a connection between the perilymph-containing cochlea and the subarachnoid space (Fig. 8.122).
Membranous labyrinth
The membranous labyrinth is a continuous system of ducts and sacs within the bony labyrinth. It is filled with endolymph and separated from the periosteum that covers the walls of the bony labyrinth by perilymph.
Consisting of two sacs (the utricle and the saccule) and four ducts (the three semicircular ducts and the cochlear duct), the membranous labyrinth has unique functions related to balance and hearing:
the utricle, saccule, and three semicircular ducts are part of the vestibular apparatus (i.e., organs of balance);
the cochlear duct is the organ of hearing.
The general organization of the parts of the membranous labyrinth (Fig. 8.122) places:
the cochlear duct within the cochlea of the bony labyrinth, anteriorly;
the three semicircular ducts within the three semicircular canals of the bony labyrinth, posteriorly; and
the saccule and utricle within the vestibule of the bony labyrinth, in the middle.
Organs of balance
Five of the six components of the membranous labyrinth are concerned with balance. These are the two sacs (the utricle and the saccule) and three ducts (the anterior, posterior, and lateral semicircular ducts).
Utricle, saccule, and endolymphatic duct
The utricle is the larger of the two sacs. It is oval, elongated and irregular in shape and is in the posterosuperior part of the vestibule of the bony labyrinth.
The three semicircular ducts empty into the utricle. Each semicircular duct is similar in shape, including a dilated end forming the ampulla, to its complementary bony semicircular canal, only much smaller.
The saccule is a smaller, rounded sac lying in the anteroinferior part of the vestibule of the bony labyrinth (Fig. 8.122). The cochlear duct empties into it.
The utriculosaccular duct establishes continuity between all components of the membranous labyrinth and connects the utricle and saccule. Branching from this small duct is the endolymphatic duct, which enters the vestibular aqueduct (a channel through the temporal bone) to emerge onto the posterior surface of the petrous part of the temporal bone in the posterior cranial fossa. Here the endolymphatic duct expands into the endolymphatic sac, which is an extradural pouch that functions in resorption of endolymph.
Sensory receptors
Functionally, sensory receptors for balance are organized into unique structures that are located in each of the components of the vestibular apparatus. In the utricle and saccule the sense organ is the macula of utricle and the macula of saccule, respectively, and in the ampulla of each of the three semicircular ducts it is the crista.
The utricle responds to centrifugal and vertical acceleration, while the saccule responds to linear acceleration. In contrast, the receptors in the three semicircular ducts respond to movement in any direction.
Organ of hearing
Cochlear duct
The cochlear duct has a central position in the cochlea of the bony labyrinth dividing it into two canals (the scala vestibuli and the scala tympani). It is maintained in this position by being attached centrally to the lamina of modiolus, which is a thin lamina of bone extending from the modiolus (the central bony core of the cochlea), and peripherally to the outer wall of the cochlea (Fig. 8.123).
Thus, the triangular-shaped cochlear duct has:
an outer wall against the bony cochlea consisting of thickened, epithelial-lined periosteum (the
spiral ligament);
a roof (the
vestibular membrane), which separates the endolymph in the cochlear duct from the perilymph in the scala vestibuli and consists of a membrane with a connective tissue core lined on either side with epithelium; and
a floor, which separates the endolymph in the cochlear duct from the perilymph in the scala tympani and consists of the free edge of the lamina of modiolus, and a membrane (the
basilar membrane) extending from this free edge of the lamina of modiolus to an extension of the spiral ligament covering the outer wall of the cochlea.
The spiral organ is the organ of hearing, rests on the basilar membrane, and projects into the enclosed, endolymph-filled cochlear duct (Fig. 8.123).
Vessels
The arterial supply to the internal ear is divided between vessels supplying the bony labyrinth and the membranous labyrinth.
The bony labyrinth is supplied by the same arteries that supply the surrounding temporal bone—these include an anterior tympanic branch from the maxillary artery, a stylomastoid branch from the posterior auricular artery, and a petrosal branch from the middle meningeal artery.
The membranous labyrinth is supplied by the labyrinthine artery, which either arises from the anterior inferior cerebellar artery or is a direct branch of the basilar artery—whatever its origin, it enters the internal acoustic meatus with the facial [VII] and vestibulocochlear [VIII] nerves and eventually divides into:
a
cochlear branch, which passes through the modiolus and supplies the cochlear duct; and
one or two
vestibular branches, which supply the vestibular apparatus.
Venous drainage of the membranous labyrinth is through vestibular veins and cochlear veins, which follow the arteries. These come together to form a labyrinthine vein, which eventually empties into either the inferior petrosal sinus or the sigmoid sinus.
Innervation
The vestibulocochlear nerve [VIII] carries special afferent fibers for hearing (the cochlear component) and balance (the vestibular component). It enters the lateral surface of the brainstem, between the pons and medulla, after exiting the temporal bone through the internal acoustic meatus and crossing the posterior cranial fossa.
Inside the temporal bone, at the distal end of the internal acoustic meatus, the vestibulocochlear nerve divides to form:
the
cochlear nerve; and
the
vestibular nerve.
The vestibular nerve enlarges to form the vestibular ganglion, before dividing into superior and inferior parts, which distribute to the three semicircular ducts and the utricle and saccule (Fig. 8.119).
The cochlear nerve enters the base of the cochlea and passes upward through the modiolus. The ganglion cells of the cochlear nerve are in the spiral ganglion at the base of the lamina of modiolus as it winds around the modiolus (Fig. 8.121). Branches of the cochlear nerve pass through the lamina of modiolus to innervate the receptors in the spiral organ.
Facial nerve [VII] in the temporal bone
The facial nerve [VII] is closely associated with the vestibulocochlear nerve [VIII] as it enters the internal acoustic meatus of the temporal bone. Traveling through the temporal bone, its path and several of its branches are directly related to the internal and middle ears.
The facial nerve [VII] enters the internal acoustic meatus in the petrous part of the temporal bone (Fig. 8.124A). The vestibulocochlear nerve and the labyrinthine artery accompany it.
At the distal end of the internal acoustic meatus, the facial nerve [VII] enters the facial canal and continues laterally between the internal and middle ears. At this point the facial nerve [VII] enlarges and bends posteriorly and laterally. The enlargement is the sensory geniculate ganglion. As the facial canal continues, the facial nerve [VII] turns sharply downward and running in an almost vertical direction, it exits the skull through the stylomastoid foramen (Fig. 8.124A).
Branches
Greater petrosal nerve.
At the geniculate ganglion, the facial nerve [VII] gives off the greater petrosal nerve (Fig. 8.124A). This is the first branch of the facial nerve [VII]. The greater petrosal nerve leaves the geniculate ganglion, travels anteromedially through the temporal bone, and emerges through the hiatus for the greater petrosal nerve on the anterior surface of the petrous part of the temporal bone (Fig. 8.117). The greater petrosal nerve carries preganglionic parasympathetic fibers to the pteryatine ganglion.
Continuing beyond the bend, the position of the facial nerve [VII] is indicated on the medial wall of the middle ear by a bulge (Fig. 8.116).
Nerve to stapedius and chorda tympani.
Near the beginning of its vertical descent, the facial nerve [VII] gives off a small branch, the nerve to stapedius (Fig. 8.124), which innervates the stapedius muscle, and just before it exits the skull the facial nerve [VII] gives off the chorda tympani nerve.
The chorda tympani does not immediately exit the temporal bone, but ascends to enter the middle ear through its posterior wall, passing near the upper aspect of the tympanic membrane between the malleus and incus (Fig. 8.124B). It then exits the middle ear through a canal leading to the petrotympanic fissure and exits the skull through this fissure to join the lingual nerve in the infratemporal fossa.
Transmission of sound
A sound wave enters the external acoustic meatus and strikes the tympanic membrane moving it medially (Fig. 8.125). As the handle of malleus is attached to this membrane, the handle of malleus also moves medially. This moves the head of the malleus laterally. Because the heads of the malleus and incus articulate with each other, the head of the incus is also moved laterally. This pushes the long process of the incus medially. The long process articulates with the stapes, so its movement causes the stapes to move medially. In turn, because the base of stapes is attached to the oval window, the oval window is also moved medially.
This action completes the transfer of a large-amplitude, low-force, airborne wave that vibrates the tympanic membrane into a small-amplitude, high-force vibration of the oval window, which generates a wave in the fluid-filled scala vestibuli of the cochlea.
The wave established in the perilymph of the scala vestibuli moves through the cochlea and causes an outward bulging of the secondary tympanic membrane covering the round window at the lower end of the scala tympani (Fig. 8.125). This causes the basilar membrane to vibrate, which in turn leads to stimulation of receptor cells in the spiral organ.
The receptor cells send impulses back to the brain through the cochlear part of the vestibulocochlear nerve [VIII] where they are interpreted as sound.
If the sounds are too loud, causing excessive movement of the tympanic membrane, contraction of the tensor tympani muscle (attached to the malleus) and/or the stapedius muscle (attached to the stapes) dampens the vibrations of the ossicles and decreases the force of the vibrations reaching the oval window.