Chapter 10 Hormonal cycles
fertilization and early development
Monthly physiological changes take place in the ovaries and the uterus, regulated by hormones produced by the hypothalamus, pituitary gland and ovaries. These cycles commence at puberty and occur simultaneously and together are known as the female reproductive cycle. The functions of the cycle are to prepare the egg often referred to as the gamete or oocyte for fertilization by the spermatozoon (sperm), and to prepare the uterus to receive and nourish the fertilized egg. If fertilization has not taken place the inner lining of the uterus or endometrium and the egg are shed and bleeding occurs per vagina, and the cyclic events begin again.
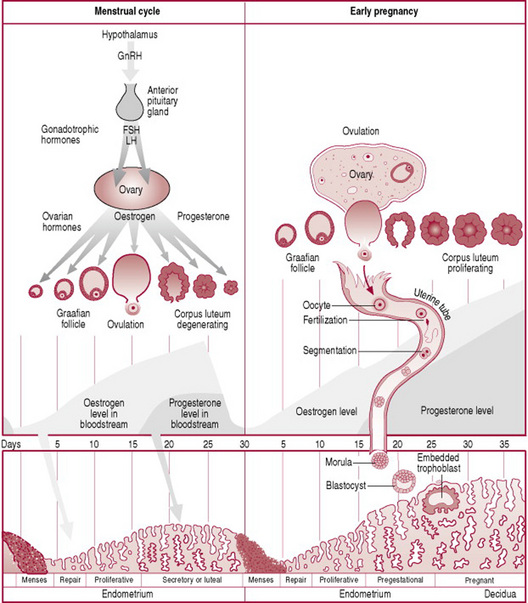
The first-ever occurrence of cyclic events is termed menarche, meaning the first menstrual bleeding. The average age of menarche is 12 years, although between the ages 8 and 16 is considered normal. Factors such as heredity, diet and overall health can accelerate or delay menarche. Interference with the hormonal-organ relationship during the reproductive years is likely to cause menstrual cycle dysfunction which may result in failure to ovulate. The cessation of cyclic events is referred to as the menopause, and signifies the end of reproductive life. Each woman has an individual reproductive cycle that varies in length, although the average cycle is normally 28 days long, and recurs regularly from puberty to the menopause except when pregnancy intervenes (Fig. 10.1).
The ovarian cycle
The ovarian cycle (Fig. 10.2) is the name given to the physiological changes that occur in the ovaries essential for the preparation and release of an oocyte. The ovarian cycle consists of three phases, all of which are under the control of hormones.
The follicular phase
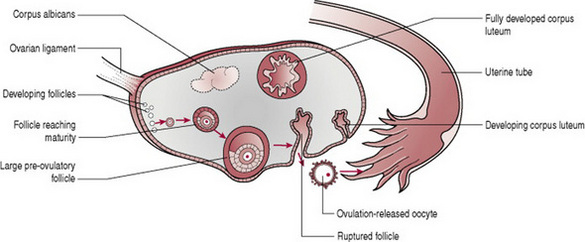
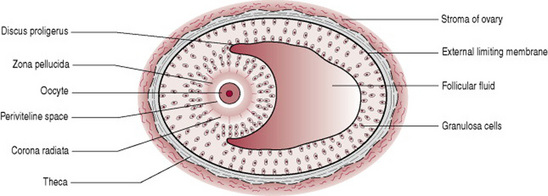
Throughout the year small primordial follicles containing primary oocytes, have been developing into large preovulatory or Graafian follicles containing secondary oocytes (Fig. 10.3), in a process known as folliculogenesis (oocytes undergo mitosis (cell division) prior to birth and until puberty in a process known as oogenesis).
Low levels of oestrogen and progesterone stimulate the hypothalamus to produce gonadotrophin releasing hormone (GnRH). This releasing hormone causes the production of follicle stimulating hormone (FSH) and luteinizing hormone (LH) by the anterior pituitary gland. FSH controls the growth and maturity of the Graafian follicles. The Graafian follicles begin to secrete oestrogen, which comprises oestradiol, oestrone and oestriol. Rising levels of oestradiol cause a surge in LH. When oestradiol reaches a certain peak, the secretion of FSH is inhibited. The reduced FSH secretion causes a slowing in follicle growth and eventually leads to follicle death, known as atresia. The largest and dominant follicle secretes inhibin, which further suppresses FSH. This dominant follicle prevails and forms a bulge near the surface of the ovary, and soon becomes competent to ovulate. The time from the growth and maturity of the Graafian follicles to ovulation is normally around 1 week, day 5–14 of a 28-day cycle of events. Occasionally the follicular phase may take longer if the dominant follicle does not ovulate and the phase will begin again.
Ovulation
Ovulation is the process whereby the dominant follicle ruptures and discharges the secondary oocyte into the uterine tube where it awaits fertilization. Ovulation is stimulated by a sudden surge in LH which matures the oocyte and weakens the wall of the follicle. This LH surge occurs around day 12–13 of a 28-day cycle and lasts 48 hrs. Stringy clear mucus appears in the cervix, ready to accept the sperm from intercourse. During ovulation some women experience varying degrees of abdominal pain known as mittelschmerz, which can last several hours. There may be some light bleeding caused by the hormonal changes taking place. Following ovulation the fertilized or unfertilized oocyte travels to the uterus.
The luteal phase
The luteal phase is the process whereby the cells of the residual ruptured follicle proliferate and form a yellow irregular structure known as the corpus luteum. The corpus luteum produces oestrogen and progesterone for approximately 2 weeks, to develop the endometrium of the uterus, which awaits the fertilized oocyte. The corpus luteum continues its role until the placenta is adequately developed to take over. In the absence of fertilization the corpus luteum degenerates and becomes the corpus albicans (white body), and progesterone and oestrogen, and inhibin levels decrease. In response to low levels of oestrogen and progesterone the hypothalamus produces GnRH. The rising levels of GnRH stimulate the anterior pituitary gland to produce FSH and the ovarian cycle commences again (Stables and Rankin 2004).
The menstrual or endometrial cycle
The menstrual cycle is the name given to the physiological changes that occur in the uterus, and which are essential to receive the fertilized oocyte. The menstrual cycle consists of three phases.
The menstrual phase
This phase is often referred to as menstruation, bleeding, menses, or a period. Physiologically this is the terminal phase of the reproductive cycle of events and is simultaneous with the beginning of the follicular phase of the ovarian cycle. The spiral arteries of the endometrium go into spasm withdrawing the blood supply to it, and the endometrium dies, referred to as necrosis. The endometrium is shed down to the basal layer along with blood from the capillaries and the unfertilized oocyte. Failure to menstruate is an indication that a woman may have become pregnant. The term eumenorrhea denotes normal, regular menstruation that lasts for typically 3–5 days, although 2–7 days is considered normal. The average blood loss during menstruation is 50–150 mL. The blood is inhibited from clotting due to the enzyme plasmin contained in the endometrium. The term menorrhagia denotes heavy bleeding.
Some women experience uterine cramps caused by muscular contractions to expel the tissue. Severe uterine cramps are known as dysmenorrhea.
The proliferative phase
This phase follows menstruation, is simultaneous with the follicular phase and lasts until ovulation. There is the formation of a new layer of endometrium in the uterus, referred to as the proliferative endometrium. This phase is under the control of oestradiol and other oestrogens secreted by the Graafian follicle and consist of the re-growth and thickening of the endometrium in the uterus. During the first few days of this phase the endometrium is re-forming, described as in the regenerative phase. At the completion of this phase the endometrium consists of three layers. The basal layer lies immediately above the myometrium and is approximately 1 mm thick. It contains all the necessary rudimentary structures for building new endometrium. The functional layer, which contains tubular glands, is approximately 2.5 mm thick, and lies on top of the basal layer. It changes constantly according to the hormonal influences of the ovary. The layer of cuboidal ciliated epithelium covers the functional layer. It dips down to line the tubular glands of the functional layer. If fertilization occurs, the fertilized oocyte implants itself within the endometrium.
The secretory phase
This phase follows the proliferative phase and is simultaneous with ovulation. It is under the influence of progesterone and oestrogen secreted by the corpus luteum. The functional layer of the endometrium thickens to approximately 3.5 mm and becomes spongy in appearance because the glands are more tortuous. The blood supply to the area is increased and the glands produce nutritive secretions such as glycogen. These conditions last for approximately 7 days, awaiting the fertilized oocyte.
Fertilization
Human fertilization, known as conception, is the fusion of the sperm with the secondary oocyte, to form the zygote (Fig. 10.4). The process takes approximately 24 hrs and normally occurs in the ampulla of the uterine tube. Following ovulation, the oocyte, which is about 0.15 mm in diameter, passes into the uterine tube. The oocyte, having no power of locomotion, is wafted along by the cilia and by the peristaltic muscular contraction of the uterine tube. At the same time the cervix which is under the influence of oestrogen, secretes a flow of alkaline mucus that attracts the spermatozoa. In the fertile male at intercourse approximately 300 million sperm are deposited in the posterior fornix of the vagina. Those that reach the loose cervical mucus survive and propel themselves towards the uterine tubes while the rest are destroyed by the acid medium of the vagina. Once inside the uterine tubes the sperm undergo a process known as capacitation. Influenced by secretions from the uterine tube the sperm undergo changes to the plasma membrane, resulting in the removal of the glycoprotein coat. The acrosomal layer of the sperm becomes reactive and releases the enzyme hyaluronidase known as the acrosome reaction, which disperses the corona radiata (the outermost layer of the oocyte) allowing access to the zona pellucida (Fig. 10.4C). The first sperm that reaches the zona pellucida penetrates it (Fig. 10.4D). Penetration of the zona pellucida occurs with the aid of several enzymes processed by the sperm which break down the proteins of the zona layer. Upon penetration a chemical reaction known as the cortical reaction occurs. The cortical reaction alters the zona pellucida making it impermeable to other sperm. The plasma membranes of the sperm and oocyte fuse. The oocyte at this stage completes its second meiotic division, and becomes mature. The pronucleus now has 23 chromosomes, referred to as haploid. The tail and mitochondria of the sperm degenerate as the sperm penetrates the oocyte, and there is the formation of the male pronucleus. The male and female pronuclei fuse to form a new nucleus that is a combination of the genetic material from both the sperm and oocyte, referred to as a diploid cell. The male and the female gametes each contribute half the complement of chromosomes to make a total of 46 (Box. 10.1). This new cell is called a zygote.
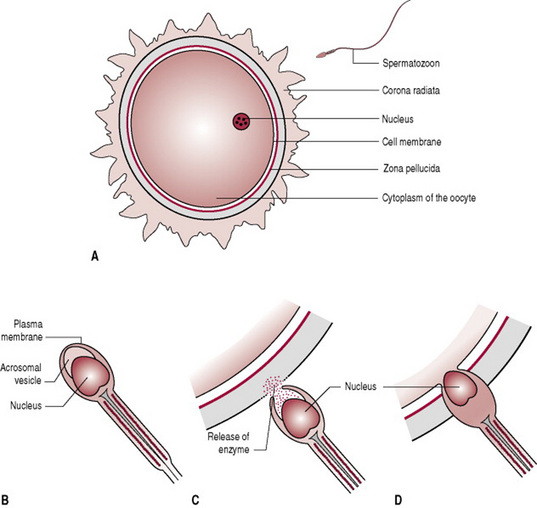
Figure 10.4 Fertilization. Diagrammatic representation of the fusion of the oocyte and the spermatozoon.
(Note that B, C and D are more greatly magnified than A.)
Each human cell has a complement of 46 chromosomes arranged in 23 pairs, of which two are sex chromosomes. The remainder are known as autosomes. During the process of maturation, both gametes shed half their chromosomes, one of each pair, during a reduction division called meiosis. Genetic material is exchanged between the chromosomes before they split up. In the male, meiosis starts at puberty and both halves redivide to form four sperm in all. In the female, meiosis commences during fetal life but the first division is not completed until many years later at ovulation. The division is unequal; the larger part will eventually go on to form the oocyte while the remainder forms the first polar body. At fertilization the second division takes place and results in one large cell, which is now mature, and a much smaller one, the second polar body. At the same time, division of the first polar body creates a third polar body.
When the gametes combine at fertilization to form the zygote, the full complement of chromosomes is restored. Subsequent division occurs by mitosis where the chromosomes divide to give each new cell a full set.
Sex determination
Females carry two similar sex chromosomes, XX; males carry two dissimilar sex chromosomes, XY. Each sperm will carry either an X or a Y chromosome, whereas the oocyte always carries an X chromosome. If the oocyte is fertilized by an X-carrying sperm a female is conceived, if by a Y-carrying one, a male.
Development of the zygote
The development of the zygote can be divided into three periods. The first 2 weeks after fertilization referred to as the pre-embryonic period includes the implantation of the zygote into the endometrium; weeks 2–8 are known as the embryonic period; and weeks 8 to birth, are known as the fetal period.
The pre-embryonic period
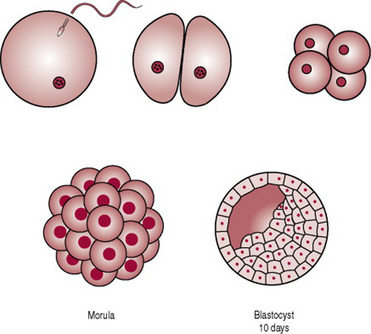
During the first week the zygote travels along the uterine tube towards the uterus. At this stage a strong membrane of glycoproteins called the zona pellucida surrounds the zygote. The zygote receives nourishment, mainly glycogen, from the goblet cells of the uterine tubes and later the secretory cells of the uterus. During the travel the zygote undergoes mitotic cellular replication and division referred to as cleavage, resulting in the formation of smaller cells known as blastomeres. The zygote divides into two cells at 1 day, then four at 2 days, eight by 2.5 days, 16 by 3 days, now known as the morula. The cells bind tightly together in a process known as compactation. Next cavitation occurs whereby the outermost cells secrete fluid into the morula and a fluid-filled cavity or blastocele appears in the morula. This results in the formation of the blastula or blastocyst, comprising 58 cells. The process from the development of the morula to the development of the blastocyst is referred to as blastulation and has occurred by around day 4 (Fig. 10.5). The surrounding zona pellucida begins to disintegrate and the volume of fluid increases. Around days 3–5 the blastocyst enters the uterus. The blastocyst possesses an inner cell mass or embryoblast, and an outer cell mass or trophoblast. The trophoblast becomes the placenta and chorion, while the embryoblast becomes the embryo, amnion and umbilical cord (Carlson 2004).
During week 2, the trophoblast proliferates and differentiates into 2 layers: the outer syncytiotrophoblast or syncytium and the inner cytotrophoblast (cuboidal dividing cells) (Fig. 10.6). Implantation of the trophoblast layer into the endometrium now known as the decidua begins (see Ch. 11). Implantation is usually to the upper posterior wall. At the implantation stage the zona pellucida will have totally disappeared. The syncytiotrophoblast layer invades the decidua by forming finger-like projections called villi that make their way into the decidua and spaces called lacunae that fill up with the mother’s blood. The villi begin to branch, and contain blood vessels of the developing embryo, thus allowing gaseous exchange between the mother and embryo. Implantation is assisted by hydrolytic enzymes secreted by the syncytiotrophoblast cells that erode the decidua and assist with the nutrition of the embryo. The syncytiotrophoblast cells also produce human chorionic gonadotrophin (hCG), a hormone that prevents menstruation and maintains pregnancy by sustaining the function of the corpus luteum.
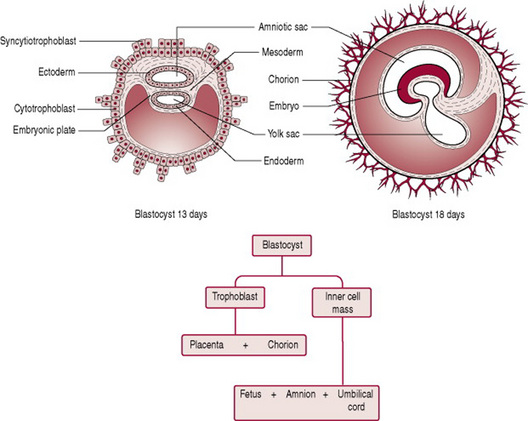
Simultaneously to implantation, the embryo is developing from the embryoblast. The cells of the embryoblast differentiate into two types of cells: the epiblast (closest to the trophoblast) and the hypoblast (closest to the blastocyst cavity). The epiblast cells give rise to cells of the embryo. Each layer of epiblast cells, of which there are three, will form particular parts of the embryo. The first appearance of these layers, collectively known as the primitive streak, is around day 15.
The epiblast separates from the trophoblast and forms a cavity, known as the amniotic cavity. The amniotic cavity derives from the ectoderm layer. The cavity is filled with fluid, and gradually enlarges and folds around the developing structures of the embryo to enclose it. The amnion forms from the lining of the cavity. It swells out into the chorionic cavity and eventually obliterates it when the amniotic and chorionic membranes come into contact.
The hypoblast layer of the embryoblast gives rise to extra-embryonic structures only, such as, the yolk sac. Hypoblast cells migrate along the inner cytotrophoblast lining of the blastocele secreting extracellular tissue which becomes the yolk sac. The yolk sac is lined with extraembryonic endoderm, which in turn is lined with extraembryonic mesoderm. The yolk sac serves as a primary nutritive function, carrying nutrients and oxygen to the embryo. The endoderm and mesoderm cells contribute to the formation of some organs, such as the primitive gut arising out of the endoderm cells; blood islands which later go on to develop blood cells arise from the mesodermal layer; the remainder resembles a balloon floating in front of the embryo until it atrophies by the end of the 6th week when blood forming activity transfers to embryonic sites. After birth, all that remains of the yolk sac is a vestigial structure in the base of the umbilical cord, known as the vitelline duct.
The pre-embryonic period is crucial in terms of initiation and maintenance of the pregnancy and early embryonic development. Inability to implant properly results in miscarriage. Additionally chromosomal defects and abnormalities in structure and organs can occur during this time (Moore & Persaud 2003).
Coad J, Dunstall M. Anatomy and physiology for midwives. Edinburgh: Mosby, 2001.
A very full and clear explanation of endocrine activity is given in Chapter 3. Chapter 4 addresses the reproductive cycles in similar detail with clear diagrams to assist the reader.
Johnson MH, Everitt BJ. Essential reproduction, 5th edn. Oxford: Blackwell Science, 2000.
This authoritative volume provides the interested reader with a much greater depth of information than is possible in the present book and is recommended for those who wish to study the hormonal patterns of reproduction in detail.