Introduction to physiology
 Objectives
ObjectivesIn this chapter, you will learn to:
• Describe in detail the structure and function of the mammalian cell and the different cell organelles
• Relate the structure of the above to their function
• Discuss how energy is formed within the cell, e.g. Krebs cycle
• Outline the structure and function of DNA and RNA
• Describe in detail the sequence of events involved in protein synthesis
• Describe the different components that make up the cell membrane
• Discuss the different forms of transport involved in the movement of molecules across cell membranes
OVERVIEW OF BODY SYSTEMS
About 50-65% of the body consists of liquids, termed ‘fluid’. Two-thirds of this fluid is contained within cells (i.e. it is intracellular) and one-third is outside the cells (extracellular). The different types of fluid have different constituents (Fig. 1.1).
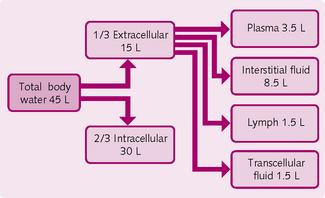
Extracellular fluid is a medium from which cells take up nutrients and into which they discharge metabolic waste products. About 75% of the extracellular fluid is found in between the cells, where it is called interstitial fluid; once it passes into the lymphatic vessels its name changes to lymph. Other extracellular fluids include cerebrospinal fluid and pericardial fluid; plasma, which is found in the blood vessels, comprises the remainder.
As blood flows through the capillaries, some of the plasma passes through the capillary pores to become the interstitial fluid that bathes the cells. The continual bidirectional movement of extracellular fluid from plasma into the interstitial fluid permits the exchange of nutrients and waste products.
Nutrient intake
The nutrients in the extracellular system originate from the:
• Respiratory system: blood flowing through the lung picks up oxygen via diffusion from the alveoli.
• Gastrointestinal system: nutrients from the gut, such as sugars, amino acids and fatty acids, are absorbed into the blood.
• Liver: the liver chemically modifies substances absorbed from the gut to facilitate their use by the body. Other body tissues or organs store some of these nutrients (e.g. fat cells, endocrine glands) until they are required.
• Musculoskeletal system: moves the body to enable it to physically obtain food and avoid adverse surroundings.
Removal of metabolic end products
• Lungs: carbon dioxide diffuses into the alveoli and is expelled into the atmosphere via exhalation.
• Kidneys: the kidneys remove the majority of waste products not needed by cells (e.g. urea, uric acid, excess ions and water), but not CO2. At the same time, the kidneys reabsorb those products needed by the body (glucose, electrolytes, amino acids and water).
Regulation of body functions
The nervous and hormonal systems work together to regulate muscular, secretory and metabolic activity and so achieve stability within the body.
Nervous system
The nervous system comprises three parts.
1.: Central nervous system The central nervous system (CNS) consists of the brain and spinal cord. It stores information and processes thoughts and emotions. Signals generated by the CNS are transferred via the motor output of the peripheral nervous system to initiate an appropriate response.
Hormonal secretion
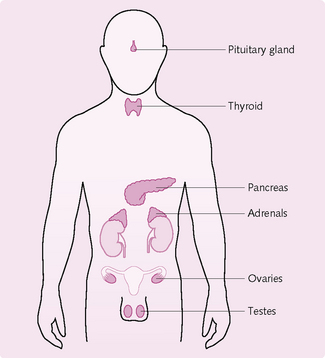
Eight primary endocrine glands secrete substances called hormones (Fig. 1.2) into the extracellular fluid, which transports them to their site of action. Hormones regulate metabolic processes, e.g. the thyroid gland secretes the hormone thyroxine, which increases the rate of many reactions in the body.
Homeostasis
This term refers to the maintenance of constant conditions within the body; all the tissues and organs of the body work together to achieve homeostasis. In general, it depends on a negative feedback loop so that, when a change occurs in the body, the body aims to restore the status quo by reversing the direction of change. For example, if you are suddenly moved from a cold environment to somewhere hot, your body temperature will increase. As the body must keep its temperature within certain limits to optimize function, it sets in motion a number of actions to reduce its temperature. These actions include:
• Vasodilatation of the surface vessels: so that heat is lost by convection.
Such responses tend to keep the body’s internal environment constant i.e. they sub-serve homeostasis.
THE CELL
Microstructure of the cell
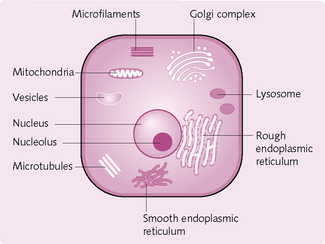
Protoplasm is the collective name given to the basic components of the cell, i.e. the material from which the cell is made. The protoplasm therefore includes the cell nucleus and cytoplasm (Fig. 1.3). The protoplasm is made up of:
• Water: comprises the majority of the cell (around 80%).
• Proteins: form 15% of the cell. There are two types:
• Lipids: there are three main types within the cell:
• Carbohydrates: stored as glycogen and provide the cell with energy.
• Electrolytes, which are used for cellular reactions (e.g. action potential): potassium ions (K+), magnesium ions (Mg2+), phosphate ions (PO43−), sulphate ions (SO42−), bicarbonate ions (HCO3−).
Organelles and their function
Within the cell are highly organized physical structures – organelles – which carry out specific functions.
Nucleus: This organelle is present in almost all cells and is the ‘control centre’ (Fig. 1.4). It stores and transmits genetic information (in the form of DNA) to the next generation. The genes within the cell’s nucleus determine the types of protein made by the cell.
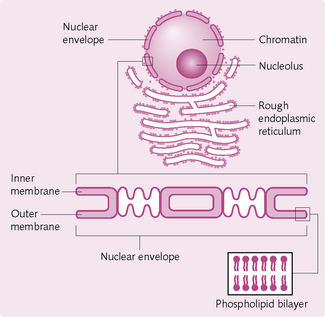
Nuclear membrane/envelope: These two membranes, which surround the nucleus, contain pores that regulate the entry and exit of molecules.
Nucleoli: The nucleoli are highly coiled structures that are not enveloped by a nuclear membrane. They contain RNA and protein components
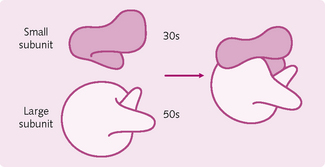
Ribosomes: These large particles are composed of about 70 proteins and several RNA molecules (Fig. 1.5). There are two subunits of different sizes – 30s and 50s – the former being smaller.
Protein molecules are synthesized by ribosomes from amino acids, using genetic information derived from DNA in the nucleus and carried to the ribosomes by RNA messenger molecules. The proteins formed are either released in the cytosol (the fluid part of the cell’s cytoplasm) or transferred to other organelles via the Golgi apparatus (see below).
Endoplasmic reticulum: The endoplasmic reticulum (ER) forms a network of membranes within the cell. There are two types:
1. Granular (or rough) ER: carries surface ribosomes. Proteins are synthesized on the attached ribosomes, enter the lumen of the ER and are either distributed within the cell or secreted to other cells.
2. Agranular (or smooth) ER: no ribosomes on the surface. This type of ER synthesizes fatty acids and regulates cellular levels of calcium (Ca2+), which controls many of the cell’s activities.
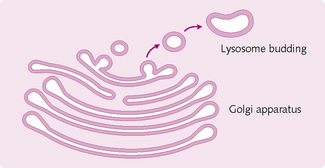
Golgi apparatus: These membranous sacs (Fig. 1.6) sort and modify proteins arriving from the granular ER, packaging them into vesicles before sending them to other organelles within the cell or secreting them.
Mitochondria: These are double-membraned, elongated, ovoid structures. The outer membrane is smooth and the inner is folded into tubes (cristae), thereby increasing the surface area. The cristae extend into the inside of the mitochondria (matrix) and contain DNA for mitochondrial protein synthesis.
The mitochondria function to make energy available to cells in the form of adenosine triphosphate (ATP).
Lysosomes: These single-membraned, oval organelles contain highly acidic digestive enzymes that break down bacteria, cell debris and dead organelles.
Peroxisomes: These single-membraned, oval organelles destroy the highly toxic compound hydrogen peroxide (H2O2), which is a by-product of certain cell reactions. Additionally, the perixosomes in the liver and kidney cells manufacture H2O2 and use it to detoxify various ingested molecules.
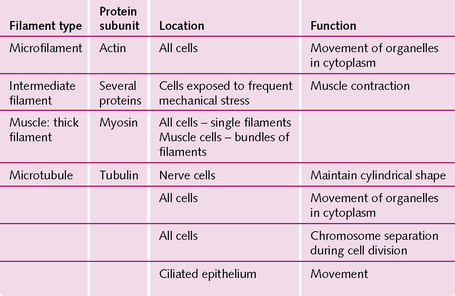
Filaments: The cytoplasm of most cells contains protein filaments – the cytoskeleton – which maintain the cell’s shape and produce cell movements. The four main types, and their functions, are outlined in Figure 1.7.
Energy production in the cell
Cells use oxygen to break down foodstuffs (carbohydrates → glucose, proteins → amino acids, fats → fatty acids) and form the compound adenosine triphosphate (ATP). ATP is a general source of energy for all intracellular metabolic reactions.
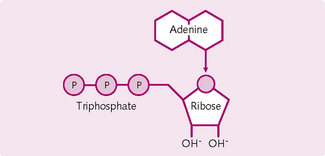
Structure of ATP: ATP is a nucleotide containing the base adenine, the pentose sugar ribose and three phosphate molecules (Fig. 1.8). The end two phosphate molecules are connected by a high-energy bond. When this bond is hydrolysed to adenosine diphosphate (ADP), large amounts of energy are liberated. Loss of another phosphate to form adenosine monophosphate (AMP) releases yet more energy. Each covalent bond broken liberates about 30.6 kJ/mol. This is about the same as the energy in a single peanut.
Formation of ATP: The majority (95%) of ATP is formed in the mitochondrial matrix (Fig. 1.9). There are four stages of production:
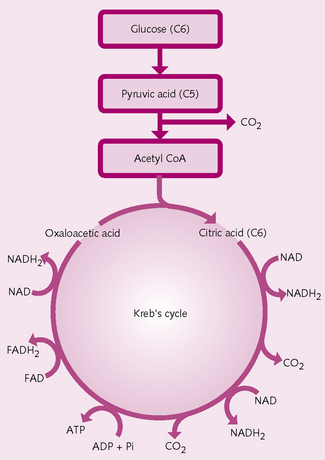
1.: Glycolysis Glycolysis is the formation of pyruvic acid (PA). Glucose, fatty acids and amino acids enter the cell cytoplasm and are converted to pyruvic acid, releasing energy for the formation of two molecules of ATP.
2.: Conversion of PA to acetyl-CoA PA enters the mitochondrial matrix, where enzymatic changes transform it into acetyl-CoA.
3.: Krebs cycle Krebs cycle is sometimes called the citric acid or tricarboxylic acid cycle. Acetyl-CoA enters a series of chemical reactions in the mitochondrial matrix in which it is split into acetyl and CoA. The acetyl portion enters the cycle and the CoA is used in the formation of more acetyl-CoA. During the Krebs cycle, for every glucose molecule:
• Seven molecules of water are added.
• Sixteen hydrogen atoms and four molecules of carbon dioxide (CO2) are released.
The CO2 diffuses out of the mitochondria and is eventually expired as a waste gas from the lungs.
4.: Oxidative phosphorylation The formation of large quantities of ATP (Fig 1.10). Only a small net quantity of ATP (two molecules) is formed during Krebs cycle; more is produced during oxidative phosphorylation. There are seven main steps:
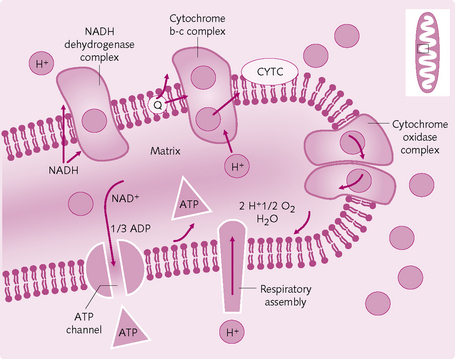
1. Hydrogen is split into a hydrogen ion (H+) and an electron.
2. The electron enters the electron transport chain on the inner mitochondrial membrane.
3. The electron is transported from electron acceptor to electron acceptor (i.e. flavoproteins and cytochromes B, C and A) until it reaches cytochrome A3 (cytochrome oxidase).
4. Cytochrome oxidase helps form ionic oxygen, which combines with the H+ to form water.
5. The large amounts of energy that are released during the transport of electrons pumps the H+ from the inner matrix of the mitochondrion to between the inner and outer membrane (outer chamber).
6. The high concentration of H+ in the outer chamber flows over the enzyme ATPase, which is attached to the inner mitochondrial membrane. This energy from the H+ flow is used by ATPase to convert ADP to ATP.
7. ATP is transferred – by facilitated diffusion – from the mitochondrion to the cell cytoplasm.
How much ATP is produced?: In total, 38 molecules of ATP are produced from one molecule of glucose:
• Two molecules during glycolysis.
• Two molecules during Krebs cycle: for each cycle, one ATP is formed. As glucose splits into two pyruvic acid molecules, two molecules of ATP are produced.
Other substrates (e.g. fats) can produce more ATP than carbohydrates. In fact, 1 g of fat can produce twice as much ATP as 1 g of carbohydrate. Protein is used as a substrate only when all the carbohydrate and fat reserves have been utilized, i.e. in a starvation state.
Genetic control of the cell
Genes control cell function. They determine what proteins – in the form of enzymes and structural proteins – are produced within the cell. Every gene is made of deoxyribonucleic acid (DNA), which regulates ribonucleic acid (RNA) to dictate the formation of the particular proteins.
DNA: DNA is a double helical chain composed of:
• Phosphoric acid, part of the backbone of the DNA molecule.
• A deoxyribose sugar, part of the backbone of the DNA molecule.
The bases connect the two DNA strands via hydrogen bonds:
This is known as complementary base pairing.
When DNA is split into its individual strands, the exposed bases provide the genetic code. Every three successive bases (triplet) codes for one amino acid. Chains of amino acids form proteins. The four bases can be arranged in 64 different three-letter combinations (4 × 4 × 4 = 64). As only 21 different amino acids are synthesized in the body, there are more than enough combinations and some amino acids are represented by more than one code. There are also codes for stop signals, which indicate that the end of a genetic message has been reached (Fig 1.11).
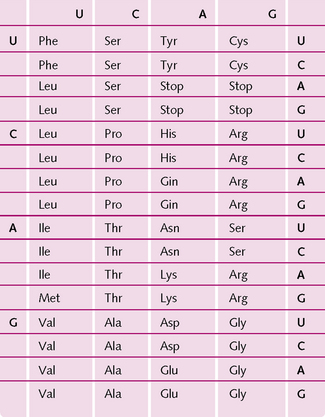
The remainder of the DNA has been classed as redundant DNA. However, new research is challenging this idea and hinting at the possibility of this ‘junk DNA’ having important functions.
Super-coiling: DNA is a very long molecule and must be packaged very well to fit into a cell. The double helix gives it a natural twist but further twisting packs it even tighter; this is super-coiling.
For proteins to form, the DNA code must be transferred to an RNA code. As DNA is in the cell nucleus and the majority of cell function occurs in the cytoplasm, the RNA acts as an intermediary to these processes.
Protein synthesis
Transcription: mRNA synthesis:
1. RNA polymerase attaches to the DNA promoter and moves along the helix (Fig. 1.12).
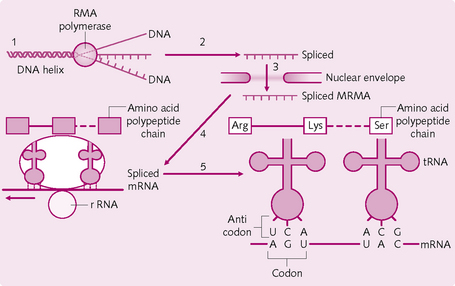
2. The DNA double helix unwinds and the strands separate.
3. RNA nucleotides attach – via hydrogen bonds – to the exposed bases on one DNA strand.
4. Covalent bonds are formed between the RNA nucleotide phosphate and ribose, producing mRNA.
5. When RNA polymerase comes across a DNA stop signal it breaks away from the DNA strand. The stop signal indicates that the mRNA is complete.
6. The hydrogen bonds holding the mRNA to the DNA strand break and the mRNA enters the nucleoplasm.
A three-base sequence in the mRNA transcript is known as a codon and is complementary to the three-base sequence in DNA (triplet).
Splicing: The primary mRNA transcript contains specific segments (exons) that code for amino acids; the remainder of the transcript does not code for any protein (introns). Splicing removes the introns and combines the remaining exons. This results in the final mRNA.
• The mRNA passes from the cell nucleus to the cytoplasm.
• rRNA binds to one end of the mRNA.
• The three-base anticodon in an amino-acid–tRNA complex pairs with its corresponding codon on the mRNA.
• The amino acid on the tRNA forms a covalent bond with the adjacent amino acid to elongate the polypeptide chain.
• The tRNA is liberated from the bound amino acid and released.
• The ribosome travels one codon along the mRNA and the procedure repeats until a termination sequence is reached.
Post-translational modifications: When the polypeptide chain has been assembled, various chemical groups might be attached and/or the protein might be split into several smaller side chains.
Cell movement
There are two types of cell movement (Figs 1.13 and 1.14):
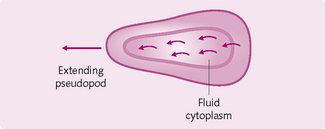

1. Amoeboid movement: a protrusion from the cell – a pseudopodium – attaches to the substrate across which the cell is moving. The remainder of the cell body drags itself towards the pseudopodium (e.g. white blood cells through tissues). Amoeboid movement is initiated by chemical substances produced by the tissues (chemotaxis). The cells can move towards (positive chemotaxis) or away from (negative chemotaxis) an area of chemotactic substances.
2. Ciliary movement: whip-like movements of cilia propel the cell. Such movement occurs on the inside surfaces of the respiratory airways and fallopian tubes. In the respiratory airways, ciliary movements propel mucus towards the trachea; in the fallopian tubes they propel the ovum towards the uterine cavity.
Membrane physiology
The cell membrane consists of three components: lipids, proteins and carbohydrates (Fig. 1.15).
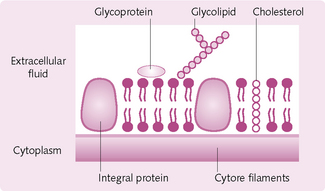
Lipids: Phospholipid molecules form a lipid bilayer, which envelops the entire cell and organelle. One part of the phospholipid molecule is hydrophilic (water loving) and the other is hydrophobic (water hating). This results in the molecules lining up with the hydrophilic portions back to back in contact with water. Because there is no chemical bond linking the phospholipids to each other they are free to move independently. This is sometimes termed ‘the fluid mosaic model’ and permits cells to change considerably without disruption to their structure.
The phospholipid bilayer regulates the movement of certain substances into and out of the cell. It prevents the entry of water-soluble substances (such as urea, glucose and ions) and allows the passage of fat-soluble substances (alcohol, oxygen and carbon dioxide).
Cholesterol, a steroid, inserts itself in the membrane with the same orientation as the phospholipid molecules. It functions to immobilize the first few hydrocarbon groups of the phospholipid molecules. If cholesterol was absent (as in a bacterium) a cell would need a cell wall. It also prevents crystallization of hydrocarbons and the membrane from shifting.
Proteins: There are two types of protein in the cell membrane:
1. Integral proteins: extend all the way through the bilayer (i.e. are transmembranous) and form structural channels through which ions and other water-soluble substances can permeate. They also form receptors for enzyme binding.
2. Peripheral proteins: located on the inside of the membrane and form enzymes.
Membranous junctions
Adjacent cells can be joined together by different types of junction: tight junctions, gap junctions and spot desmosomes (Fig. 1.16).
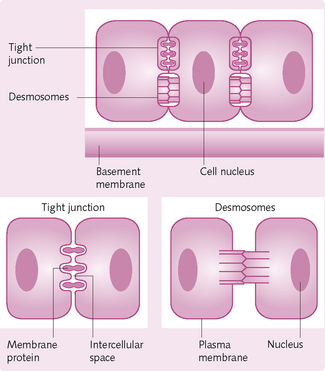
Tight junctions: These junctions form an impermeable bond between adjacent cells and direct the passage of substances through the cells by preventing passage between them. They are found in the epithelial cell sheet lining the small intestine.
Gap junctions: These are protein tunnels, which form between adjacent cells. They allow for the passage of small molecules and/or ions between the cells. Cells connected by gap junctions are capable of working synchronously as a unit instead of individually. In the heart, gap junctions allow the atria and the ventricles to contract in sequence.
Transport
The movement of molecules through the cell membrane occurs by diffusion/passive and active transport.
Diffusion: Diffusion is the movement of molecules from an area of higher concentration to an area of lower concentration as the result of random thermal molecular motion. There are several different types:
 It is important to differentiate between the different types and subtypes of diffusion. This is a commonly asked question in exams.
It is important to differentiate between the different types and subtypes of diffusion. This is a commonly asked question in exams.
Simple diffusion: Movement of molecules occurs through a membrane opening (Fig. 1.17). This opening either passes through the lipid bilayer (allowing the passage of lipid-soluble substances) or takes the form of protein channels, e.g. sodium and potassium (Na+ and K+) channels, through which water- and lipid-insoluble molecules pass. The protein channels regulate their permeability via a gating system.
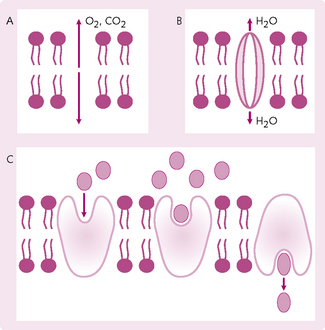
Fig. 1.17 Types of diffusion. (A) Simple diffusion. (B) Diffusion through channels: proteins and charged molecules are usually pulled through channels by water. (C) Facilitated diffusion: molecules (e.g. glucose) bind to protein, triggering a change in protein shape. This transports the glucose molecules across the membrane.
Voltage gating: The electrical potential across the cell membrane influences the entry of certain substances. For example, during an action potential an impulse passes down a neuron; the resulting reduction in the voltage causes sodium channels in the adjacent portion of the membrane to open. This allows the influx of sodium ions (Na+) into the neuron and hence the continuation of the nerve impulse
Chemical gating: Binding of another molecule permits entry. For example, acetylcholine opens the acetycholine channel, which allows the nerve signals to propagate.
Facilitated diffusion: A carrier protein is required to transport a substance across the membrane. A good example is found in the liver cells, which control the concentration of glucose in the blood. Liver cells store excess glucose as glycogen when blood sugar levels are high. The breakdown of glycogen is closely controlled by hormones. The breakdown products of glycogen are impermeable to the liver cell membrane and so a transport protein, which functions by facilitated diffusion, allows the movement of glucose.
Osmosis: This is the diffusion of water, across a semipermeable membrane (i.e. a membrane that allows the passage of certain, small molecules but which prevents the passage of large molecules). The water molecules diffuse from a region of higher water concentration to lower water concentration.
Note: a low water concentration implies a high solute concentration:
• The solutions inside and outside cells contain water; the cell wall is a semipermeable membrane. Thus osmosis applies to the movement of water molecules into and out of the cell.
• If the solution outside the cell contains a higher concentration of water molecules than the interior of the cell (i.e. the external solution is hypertonic for water), then water molecules will diffuse into the cell. This will cause the cell to swell and lyse (break).
• If the solution outside the cell contains a lower concentration of water molecules than the inside (i.e. the external solution is hypotonic for water), then water will diffuse out of the cell. This will cause the cell to shrink (crenate).
• If the concentrations of water molecules in the intracellular and extracellular concentrations are identical (isotonic), there is no net movement of water.
Active transport: This energy-dependent system transports ions or molecules across a membrane against an electrochemical gradient. It can thus maintain the concentration of a substance against a concentration gradient, e.g. the K+ concentration is high intracellularly and low extracellularly.
Active transport depends on two types of transmembrane carrier protein, which derive their energy sources in different ways:
• Primary transmembrane carrier proteins: receive their energy from ATP. They transport many different ions (e.g. Na+, K+, Ca2+ into and out of muscle cells; H+ and K+ into and out of the gastric glands). The most common type is the Na+/K+ pump system hereafter referred to as Na+/K+-ATPase, which is present in all cells of the body. This pumps Na+ out of the cell, K+ into the cell and maintains the electrical potential across the cell membrane (see Chapter 3)
• Secondary transmembrane carrier proteins: energy is generated from differences in ionic concentration between two sides of a membrane.