Proprioception
Proprioception, mechanoreception and the anatomy of fascia
It is likely that the connective tissue continuum of fasciae and fascial structures serves as a body-wide mechanosensitive signaling system with an integrating function analogous to that of the nervous system (Langevin 2006). Without doubt fasciae and fascial structures play a substantial role in the process of proprioception (Langevin 2006; Stecco et al., 2007b; Benjamin 2009). Fascial components like membranes and septa or deep and superficial fascia are an intricate and integrated part of the locomotor apparatus (Wood Jones 1944; Standring 2005). To play that functional, role in proprioception, the fascial structure should be equipped with adequate neuroanatomical substrate (‘proprioceptors’). For the quality of the centripetal information, however, how the mechanical architecture of the connective tissue structure at stake relates to the skeletal and muscular tissue in a given area is an important feature (Benjamin 2009; Van der Wal 2009). Only if a given fascial structure has a mechanical architectural relationship with muscular or skeletal elements, is it able to provide the mechanoreceptive information needed for proprioception. This means that the aptitude of a fascial structure to provide centripetal mechanoreceptive information depends on its architecture and structural relationship with muscular and skeletal tissue and not simply on its topography (Van der Wal 2009).
In this chapter proprioception is defined in the neurophysiologic way – as the ability to sense the position and location, orientation, and movement of the body and its parts. In a more strict sense, proprioception could be defined as the process of conscious and subconscious sensing of joint position and/or motion (Skoglund 1973; Fix 2002). Here, the more explicit meaning of the psychological definition of ‘proprioception’ as sometimes applied, as the notion ‘body image sense’ or ‘body awareness’ is not meant. Proprioception, in this context, has to be discriminated from exteroception, relating us with the outer world, as well as from interoception, informing about visceral and metabolic processes (see elsewhere in this book).
The morphological substrate of proprioception – encapsulated or unencapsulated mechanosensitive sensory nerve endings (mechanoreceptors) and related afferent neurons (see Fig. 2.2.1) – is considered to provide the centripetal information needed for the control of locomotion or for the maintenance of posture (Barker 1974). On the level of the brain this information is integrated with information originating from other sources (e.g., more specific proprioceptive sense organs like the labyrinth or skin receptors) to the overall conscious and subconscious awareness of position and motion (kinesthesis and statesthesis).
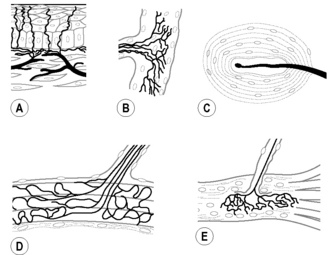
Fig. 2.2.1 • Types of mechanoreceptors.
(A) Free nerve ending (FNE); (B) Ruffini corpuscle (RC) or spraylike endings; (C) lamellated corpuscle (LC) or Paciniform ending; (D) Golgi tendon organ (GTO); (E) muscle spindle (MS).
In this context, mechanoreception is not synonymous with proprioception. Proprioception relates to mechanoreception as seeing relates to the retina. The mechanoreceptive information needed for the process of proprioception originates not only from fasciae and other connective tissue structures but also from mechanoreceptive or even tactile information from skin, muscles, joint surfaces, and joint structures. Mechanoreceptors are triggered by mechanical deformation like squeezing, stretching or compression. In order to understand their contribution to the proprioceptive information it is not only important to know their topography (where and in which elements of the locomotor apparatus they are located) but also how they are spatially and mechanically related with the various (tissue) components of the system. Proprioception in the fascia is not only provided by the mechanoreceptors that are located within or are immediately attached to the fascial structures, but also the architecture of the fascia plays an instrumental role in the process of proprioception. It can do so by mediating forces that cause deformation of receptors (which in fact represents the main stimulus for mechanoreceptors) that are not directly attached to the fascia itself. The term “ectoskeleton” has been proposed by some authors (Benjamin 2009) to capture the idea that fascia could serve as a significant site of muscle attachment, constituting a kind of “soft tissue skeleton”. Mechanoreceptors situated within muscles as anatomical units may orient as to their distribution and spatial organization to the fascial layers to which the muscle fascicles insert and between which muscular tissue is interposed in the process of force transmission. In such cases, the fascial architecture plays an instrumental role in the process of proprioception without the necessity for the connective tissue structures themselves being directly equipped with mechanoreceptive substrate (Van der Wal 2009).
So, to evaluate the significance of fascial structures as to the proprioceptive input from a certain body region it is not only important to know the anatomy of the given fascia (‘where’) but also its architecture, i.e., functional relationship (‘how’). Many fascial structures play a direct or indirect role in force transmission. Most anatomy textbooks, however, describe the locomotor apparatus as a system built up from discrete elements involved in positioning, motion, and force transmission: i.e., muscles (with tendons and aponeuroses) and ligaments. In this outdated concept, muscles represent the main elements in the system, which in atlases are often presented as discrete anatomic structures with the surrounding and ‘enveloping’ connective tissue layers removed. When connective tissue is met as a layer, a membrane, a fascia covering a body structure or organ or region, is usually given a name derived from the anatomic substrate that the layer covers. Fasciae therefore are most often defined as a suborganization of the ‘primary’ anatomy of organs (e.g., muscles) This is all related to the ‘dissectional mind’ that still prevails in the anatomy atlases and textbooks and considers the locomotor apparatus as built up from anatomical elements. When Schleip (2003a, b) mentions the fascia as “the dense irregular connective tissue that surrounds and connects every muscle, even the tiniest myofibril, and every single organ of the body forming continuity throughout the body” or as the “organ of form” (Varela & Frenk 1987), he actually presents fascia as an important integrative element in human posture and organization of movement (locomotor apparatus). Therefore an analytical and ‘dissectional’ approach of the ‘anatomy’ of the fascia cannot do justice to the role of fascial tissue and structures in proprioception.
Connectivity and continuity
The primary connective tissue of the body is the embryonic mesoderm. The mesoderm represents the matrix and environment within which the organs and structures of the body have been differentiated and in fact are ‘embedded’. Blechschmidt (2004) distinguished the mesoderm, as germinal layer, as an ‘inner tissue’ in opposition to the ectoderm and endoderm as ‘limiting tissues and proposed not to call it a ‘derm’ but to call it ‘inner tissue’. The primary “inner tissue” is the undifferentiated connective tissue mesenchyme, which in principle is organized in three components: cells, intercellular space (interstitial substances), and fibers. In the functional development and differentiation of the primary connective tissue, there are two patterns of ‘connection’. The first pattern is the development of ‘intercellular space’, which represents a fissure functioning as a sliding and slipping space. This is seen in the formation of coelom (body cavities), joint cavities, and also in bursa-like gliding spaces between adjacent tendons or muscle bellies. In this pattern spatial separation is ensured, and in this way motion is enabled. The second pattern is the formation of a binding medium. That can be fibers (e.g., as in regular dense connective tissue structures like the desmal sutures in the skull, interosseous membranes, and ligaments) or interstitial substrate and matrix (for example, in cartilaginous joints). In osteopathic circles, the continuum and continuity of the ‘connective tissue apparatus’ in the human is emphasized. Such a view is in harmony with the view that the principal function of mesoderm as “inner tissue” is ‘mediating’ in the sense of ‘connecting’ (binding) and ‘disconnecting’ (shaping space and enabling movement).
This view of two patterns of connectivity is also applicable to the anatomy of fasciae. In general, fasciae in the musculoskeletal system exhibit two different mechanical and functional aspects:
• Fasciae of muscles adjacent to spaces that are filled with loose, areolar, connective tissue (‘sliding tissue’) and sometimes with adipose tissue. They enable the sliding and gliding of muscles (and tendons) against each other or against other structures. In such splits globular or oval mechanoreceptors triggered by compression (see below) could inform the brain about the displacement and movement of fascial tissue and related structures.
• Intermuscular and epimysial fasciae that serve as areas of insertion for muscle fibers that in this way can mechanically reach a skeletal element without necessarily being attached directly to the bone. They appear as intermuscular septa but also as so-called superficial fasciae (like fascia cruris and fascia antebrachii,) providing a broad insertion area for muscle fibers. If provided with more stretch susceptible receptors, such fascial layers could inform about stresses of the fascial tissue in relation to the transmission of forces.
This indicates that fasciae exhibit a variety of mechanical relationships with neighboring tissue and therefore may play quite different functional roles as to proprioception as well. The fasciae of the organs and of muscles often represent the ‘gliding fascia’ type; in this context, coelomic cavities actually function as ‘joint spaces’ enabling motion of the organs. Many epimysial muscle fasciae function in a similar way, providing mobility between a muscle and its neighborhood. However, fasciae like the fascia cruris (tibial fascia) or the retinaculum patellae function as epimuscular aponeuroses.
Architecture is different and more than anatomy
To understand the mechanical and functional circumstances of the fascial role in connecting and in conveying stresses and in proprioception, it is therefore more important to know the architecture of the connective and muscle tissue than the regular anatomical order or topography. This applies to every fascial layer in the human body. One must know both where they are situated (anatomy) and how they are connecting and connected (architecture). Depending on the architectural relationship of the fascial connective tissue with the muscular tissue, not only juxta-articular connective tissue (like interosseous ligaments) may provide proprioceptive information about joint movement or joint position, but epimysial, intermuscular, aponeurotic, fascial layers can also play a functional role in such processes (directly or indirectly).
In this context, two views on the organization of connective and muscular tissue may be described. On the one hand, there is the well-known view that muscular and connective tissue structures have to be considered as discrete anatomical elements. In this concept muscles function as the dynamic force-transmitting structures and they are organized in parallel to ligamentous structures as the more passive force-transmitting elements. The non-ligamentous fascial connective tissue is considered as auxiliary to the muscle units as tendons and aponeuroses. Areolar fascial connective tissue shapes, space in between the anatomical elements, providing the opportunity for sliding and mobility. Such architecture is exhibited clearly in the distal regions of the limbs, where separate muscle entities (bellies with tendons) function in parallel to underlying joint capsules with or without capsular reinforcing ligaments. Here, mechanoreceptive substrate in the fascia serves the (unconscious) perception of this sliding and movement.
On the other hand, a pattern can be described where connective tissue and muscular tissue are organized mainly in series with each other in a more ‘transmuscular’ organization. Huijing et al. (2003) point out that, often, muscles which from an anatomical perspective are considered as morphologically discrete elements cannot be considered as isolated units controlling forces and movements. Detailed studies of the lateral cubital region of man and rat showed such architecture quite clearly (Van der Wal 2009). Nearly all the deep and superficial regular dense connective tissue (RDCT) layers are organized in series with muscle fascicles (presented as muscle compartment walls). Collagenous fibers that run from bone to bone – thought to be stressed passively by displacement of the articulating bones – hardly occur. Instead, there occur broad aponeurotic layers of RDCT to which relatively short muscle fascicles insert, which, on the opposite side, are directly attached to skeletal elements. Such configurations of muscle fascicles attached to the periosteum of one articulating bone, and via a layer of RDCT indirectly attached to another articulating bone, could be considered ‘dynamic ligaments’. Such ‘dynaments’ are not necessarily situated directly beside the joint cavity or in the deep part of the joint region (Van der Wal 2009).
The substrate of mechanoreception
Connective tissue and fasciae are richly innervated. (Stilwell 1957; Schleip 2003b; Stecco et al. 2007a; Benjamin 2009). Considerations such as ‘architecture versus anatomy’ mutatis mutandis may also apply for the spatial organization of mechanoreceptors, the morphologic substrate for proprioception. To study the role and function of mechanoreceptors in the process of proprioception, it may be important to know where they actually are located in such regions and how they are or are not connected with the relating tissue elements. In general, however, mechanoreceptors are often reported to occur either as muscle receptors or as joint receptors. Muscle receptors are mechanoreceptors present in the muscles, including their auxiliary structures such as tendons, aponeuroses, and fasciae. Muscle spindles and Golgi tendon organs (GTOs) are the best-known types of such receptors (Barker 1974). Joint receptors are considered to be situated in joint capsules and related structures, including reinforcing ligaments. These receptor types are usually ordered according to the (ultra)structure of the receptor itself, physiologic features, type of afferent nerve fiber, and other parameters (Freeman & Wyke 1967a, b).
Mechanoreceptors are in fact free nerve endings (FNEs), whether or not equipped with specialized end organs. The main stimulus for such receptors is deformation. Variation exists as to the micro-architecture of the ending. On the one hand, there exists the principle of lamellae around a relatively simple nerve ending. This represents the principle of the ball- or bean-shaped Vater Pacini or paciniform corpuscles, often called lamellated corpuscles (LC). On the other hand, there is the more spray-like organization of the nerve ending wrapping around and in between the deformable substrate such as connective tissue fibers. Those are the spindle-shaped Ruffini corpuscles (RC) or GTOs. These two types of microarchitecture roughly relate to the type of mechanical deformation that is at stake: compression for the lamellated bodies and traction and torsion for the spray-like type. Other varying parameters are threshold, adaptivity, and adjustability. In this general classification, the muscle spindle is a spindle-shaped spray-like ending organized around specialized muscle fibers equipped with the extra possibility of adjustable length (Strasmann et al. 1990).
As stated above, mechanoreceptors are in general reported to occur as either ‘muscle receptors’ or ‘joint receptors’. In this concept, muscle receptors are mechanoreceptors present in the muscles and joint receptors are considered to be mechanoreceptors situated in joint capsules and related structures like ligaments. In this context, the concept often prevails that joint receptors play the leading role in the process of monitoring joint position or movement for the purpose of statesthesis and kinesthesis, while muscle receptors are relegated to motor functions that operate at a subconscious or reflex level (Barker 1974).
Mechanoreceptors associated with muscles, including the muscle auxiliary structures such as tendons, are usually classified as follows (see Fig. 2.2.1):
• Muscle spindles (sensory endings with encapsulated intrafusal muscle fibers).
• GTOs (type III endings, relatively large spray-like endings, i.e., 100–600 μm diameter with high threshold and very slow-adapting).
The mechanoreceptors typically associated with joints are (see Fig. 2.2.1):
• LC (type II ending with a two- to five-layered capsule, less than 100 μm length, with low threshold and rapidly adapting).
Here, this term is preferred to the notion ‘paciniform corpuscle’.
• RC (type I ending, relatively small spray-like ending, up to 100 μm with low threshold and slow-adapting).
The functional role of architecture of the connective and muscular tissue in mechanoreception
In an extensive study of the spatial organization of the morphological substrate of proprioception in the proximal lateral cubital region of the rat (Van der Wal 2009), an inventory has been made of mechanoreceptors that may occur in direct or indirect relationships with the connective tissue layers in the joint region. A spectrum of mechanosensitive substrate occurs at the transitional areas between the regular dense connective tissue layers (organized as epimysial or intermuscular layers and septa) and the muscle fascicles organized in series with them. This substrate exhibits features of the mechanosensitive nerve terminals that usually are considered to be characteristic for joint receptors and for muscle receptors. At the so-called superficial antebrachial fascia, as well as at the intermuscular fascial layers RC as well as LC were present between the fascial layer and inserting muscle fibers. Sometimes, even, one pole of a muscle spindle was attached to those layers.
Based upon the architecture of the connective tissue and upon the spatial distribution of the substrate of mechanoreception, it is assumed that the joint receptors here are also influenced by the activity of the muscle organized in series with the collagenous connective tissue near those receptors. This supports the idea that the stresses during joint positioning are conveyed mainly via those collagenous layers and also are involved in triggering the related mechanoreceptors. In the region studied there exists no basis in morphology for so-called joint receptors that are deformed exclusively by passive strain in collagenous connective tissue structures induced by displacement of the articulating bones. The substrate of proprioception that was found in and near the RDCT apparatus in the lateral cubital region has features of mechanoreceptors that are usually linked with ‘joint receptor’ substrate, as well as of mechanoreceptors usually present in muscles and related tendons. It is obvious that, in cases like this, the fascial layers together with the in series inserting muscular tissue function as a kind of ‘dynamic ligaments’ or ‘dynaments’.
Very often myofascial areas are richly innervated and covered by nerve plexuses. In the previously-mentioned study on rat and humans (Van der Wal 2009) it was shown that over the proximal (epimysial) antebrachial fascia, as well as over the fascia ‘covering’ the supinator muscle (in fact a supinator aponeurosis), extensive plexuses were present. Plexiform arrangements of peripheral nerves sprouting over tendons and ligaments are a consistent feature in the innervation pattern in the periarticular aponeuroses of the knee and elbow joint (Wilson & Lee 1986). Stilwell (1957) states that such networks terminate in small ‘paciniform corpuscles’ and in ‘freely ending axons’ on the surface connective tissue of tendons, aponeuroses, and muscles, and in periosteal connective tissue, nearly always in the vicinity of other mechanoreceptors. The type of axons present in the nerve fascicles of the plexuses studied here (Van der Wal 2009), as well as the demonstrated (or putative) origin of those axons from the substrate of mechanoreception in the studied material, support the notion that such peri- or juxta-articular nerve plexuses are not exclusively involved in nociceptive processes, as stated by Freeman and Wyke (1967a, b). This also means that the substrate of proprioception does not necessarily have to be situated within the fascial fibers to play a functional role in proprioception. Regarded mechanically, the intermediate zones between fascial dense connective tissue and adjacent muscle fibers and/or adjacent loose areolar connective tissue might be of interest as a source of mechanoreceptive information. In areas where the fascial connective tissue is so dense that it allows little dislocation or deformation, as is the case in most ligamentous structures, it seems logical that the innervation is more involved in nociception or sympathetic vascular regulation. In the latter respect, it is worth mentioning that there exist ligaments that are mechanically important yet poorly innervated and ligaments with a key role in sensory perception that are richly innervated (Hagert et al. 2007; Benjamin 2009). It all relates to the degree to which deformation is allowed (since it is deformation that forms the major stimulus for mechanoreceptive triggering), as well to the microscopic level (the kind of mechanoreceptor) and to the macroscopic level the (architecture of the fascia and related tissue).
Dynaments: more than ligaments or muscles
The findings in the studies described earlier regarding spatial distribution and the organization of so-called ‘muscle receptors’ were even more relevant to the concept that is brought forward here. Those receptors appeared not to be organized according to principles of anatomy and topography, but to cope with the functional architecture of the connective tissue complex of the epi-, inter- and submuscular RDCT layers in relation to muscular architecture. In all the antebrachial extensor muscles studied, the distribution of muscle spindles per muscle area is uneven. If the spatial distribution of muscle spindles is considered per muscle, it is difficult to detect a common distribution pattern in all muscles (see Plate 2.2.1). The spatial distribution of those receptors, however, becomes understandable from the regional functional architecture of the connective tissue and fascia, i.e., the RDCT structures. The muscular zones that are dense in muscle spindles and GTOs are the stress- and force-conveying zones of the muscle, which are in series with the connective tissue complex proximally and in series with the peripheral tendons distally. This arrangement provides a common principle that may explain many kinds of distribution patterns. Of course, sometimes architectural units coincide with specific topographic entities, as in this study the supinator muscle with its aponeurosis nearly represents the anatomy of a ‘dynament’.
As Huijing et al. (2003) pointed out, based upon mechanical arguments that the muscles are not isolated units controlling forces and movements as often thought, apparently also on the level of spinal sensorimotor control the muscles should no longer be considered the functional entity in the locomotor system (English & Letbetter 1982; English & Weeks 1984; Van der Wal 2009). Such considerations again match the task-dependent models of brain control well: motor units are not necessarily organized with respect to individual motor nuclei, but are organized according to behavioral tasks. The concept of the locomotor apparatus being built up by architectural units of muscular tissue in series with collagenous connective tissue is more consistent with such trans- or supramuscular models than is the concept in which muscles function as the entities that maintain joint integrity parallel to ligamentous structures.
Classification of mechanoreceptors in proprioception
In consequence of the identification of an in-series organization of muscular tissue and regular dense connective tissue structures in the locomotor apparatus (mainly tendons distally and muscle compartment walls proximally), three configuration types of mechanoreceptors were identified in the mentioned study (Van der Wal 2009). Mutatis mutandis this spectrum could also be considered to represent the substrate of proprioception of the fascia:
• Muscle spindles, GTO (RC), FNE, and LC are found in areas between muscular tissue and RDCT layers. This configuration coincides with the conventional muscle–tendon spectrum of sensory nerve endings (Barker 1974; Von Düring et al. 1984).
• LC and FNE are found in areas in which RDCT adjoins reticular connective tissue, gliding spaces. This configuration coincides mainly with the spectrum of sensory nerve endings usually indicated as articular receptors (Freeman & Wyke 1967a, b; Halata 1985).
• Only FNE are present in the transition to the skeletal attachment (periosteum). This configuration coincides with the endotenonial spectrum of sensory nerve endings with mainly (mechanoreceptive) FNE from group III and IV fibers (Von Düring et al. 1984).
Most plexuses in or near the regular dense connective tissue of fascial layers contain nerve fibers of type III and IV. Nerve fibers of group III (or A delta type) are afferent from mechanoreceptors; nerve fibers of group IV (or C-type) are afferents from FNE that are either nociceptive or mechanosensitive (strain).
In the above-mentioned configurations, RC are not indicated as a separate category but GTO and RC are considered to be the same receptor type, presenting gradual differences depending on the texture of the surrounding tissue. The quartet MS - GTO/RC–LC–FNE represents the complete spectrum of mechanoreceptors in the locomotor apparatus. In this way, the three main types of so-called muscle receptors (MS, GTO, and LC) are combined with the three types of so-called capsular (or joint) receptors (RC, LC, and FNE). Depending on the local situation, this quartet therefore represents the spectrum of mechanoreceptors involved in the proprioceptive function of fasciae and fascial structures. The activity and role of a mechanoreceptor is defined not only by its functional properties, but also by its architectural environment. It is the architecture of the fascial connective tissue in relation to the muscular tissue components and skeletal elements that plays a major role in the coding of the proprioceptive information that is provided.
References
Barker D. The morphology of muscle receptors. Barker D., Hunt C.C., McIntyre A.K., eds. Handbook of Sensory Physiology, Muscle receptors. Berlin, New York: Springer Verlag; 1974;vol. II.
Benjamin M. The fascia of the limbs and back – a review. J. Anat.. 2009;214:1–18.
Blechschmidt E. The ontogenetic basis of human anatomy. Berkeley, California USA: North Atlantic Books; 2004.
English A.W., Letbetter W.D. Anatomy and innervation patterns of cat lateral gastrocnemius and plantaris muscles. Am. J. Anat.. 1982;164(1):67–77.
English A.W., Weeks O.I. Compartmentalization of single muscle units in cat lateral gastrocnemius. Exp. Brain Res.. 1984;56(2):361–368.
Fix J.D. Neuroanatomy. Hagerstown, MD: Lippincott Williams & Wilkins; 2002.
Freeman M.A.R., Wyke B.D. The innervation of the ankle joint. An anatomical and histological study in the cat. Acta Anat. (Basel). 1967;68(3):321–333.
Freeman M.A.R., Wyke B.D. The innervation of the knee joint. An anatomical and histological study in the cat. J. Anat.. 1967;101(3):505–532.
Hagert E., Garcia-Elias M., Forsgren S., Ljung B.O. Immunohistochemical analysis of wrist ligament innervation in relation to their structural composition. J. Hand Surg. [Am]. 2007;32:30–36.
Halata Z., Rettig T., Schulze W. The ultrastructure of sensory nerve endings in the human knee joint capsule. Anat. Embryol. (Berl). 1985;172(3):265–275.
Huijing P., Maas H., Baan G.C. Compartmental fasciotomy and isolating a muscle from neighbouring muscles interfere with myofascial force transmission within the rat anterior crural compartment. J. Morphol.. 2003;256:306–321.
Langevin H.M. Connective tissue: a body-wide signalling network? Med. Hypotheses. 2006;66:1074–1077.
Schleip R. Fascial plasticity – a new neurobiological explanation Part 1. J. Bodyw. Mov. Ther.. 2003;7(1):11–19.
Schleip R. Fascial plasticity – a new neurobiological explanation Part 2. J. Bodyw. Mov. Ther.. 2003;7(2):104–116.
Skoglund S. Joint receptors and kinaesthesis. Iggo A., ed. Handbook of Sensory Physiology, Handbook of Sensory Physiology. Berlin, Heidelberg, New York: Springer Verlag; 1973;vol. 2:111–136. vols.
Standring S. Gray’s Anatomy, thirty-ninth ed. London, New York: Elsevier; 2005.
Stecco C., Gagey O., Belloni A., et al. Tendinous muscular insertions onto the deep fascia of the upper limb. First part: anatomical study. Morphologie. 2007;91:29–37.
Stecco C., Gagey O., Belloni A., et al. Anatomy of the deep fascia of the upper limb. Second part: study of innervation. Morphologie. 2007;91:38–43.
Stilwell D.L. Regional variations in the innervation of deep fasciae and aponeuroses. Anat. Rec.. 1957;127:635–653.
Strasmann T.h., Van der Wal J.C., Halata Z., Drukker J. Functional topography and ultrastructure of periarticular mechanoreceptors in the lateral elbow region of the rat. Acta Anat. (Basel). 1990;138(1):1–14.
Van der Wal J.C. The architecture of connective tissue as parameter for proprioception - an often overlooked functional parameter as to proprioception in the locomotor apparatus. Int. J. Ther. Massage Bodywork. 2009;2(4):9–23.
Varela F.J., Frenk S. The organ of form: towards a theory of biological shape. J. Soc. Biol. Struct.. 1987;10:73–83.
Von Düring M., Andres K.H., Schmidt R.F. Ultrastructure of fine afferent terminations in muscle and tendon of the cat. In: Hamann W., Iggo A., eds. Sensory receptor mechanisms. Singapore: World Scientific Publishing, 1984.
Wilson A.S., Lee H.B. Hypothesis relevant to defective position sense in a damaged knee. J. Neurol. Neurosurg. Psychiatry. 1986;49:1462–1464.
Wood Jones F. Structure and function as seen in the foot. Tindall and Cox, London: Baillière; 1944.
Huijing P. Epimuscular myofascial force transmission between antagonistic and synergistic muscles can explain movement limitation in spastic paresis. J. Electromyogr. Kinesol.. 2007;17:708–724.
Huijing P. Muscular force transmission: a unified, dual or multiple system? A review and some explorative experimental results. Arch. Physiol. Biochem.. 1999;107:292–311.