CHAPTER 17 Vascular supply and drainage of the brain
The brain is a highly vascular organ, its profuse blood supply characterized by a densely branching arterial network. It has a high metabolic rate that reflects the energy requirements of constant neural activity. It receives about 15% of the cardiac output and utilizes 25% of the total oxygen consumption of the body. The brain is supplied by two internal carotid arteries and two vertebral arteries that form a complex anastomosis (circulus arteriosus, circle of Willis) on the base of the brain. Vessels diverge from this anastomosis to supply the various cerebral regions. In general, the internal carotid arteries and the vessels arising from them supply the forebrain, with the exception of the occipital lobe of the cerebral hemisphere, and the vertebral arteries and their branches supply the occipital lobe, the brain stem and the cerebellum. Venous blood from the brain drains into sinuses within the dura mater (see Ch. 27). Acute interruption of the blood supply to the brain for more than a few minutes causes permanent neurological damage. Such ischaemic strokes along with intracranial haemorrhage are major contemporary sources of morbidity and mortality.
ARTERIAL SUPPLY OF THE BRAIN
The arterial supply of the brain is derived from the internal carotid and vertebral arteries, which lie, together with their proximal branches, within the subarachnoid space at the base of the brain.
INTERNAL CAROTID ARTERY
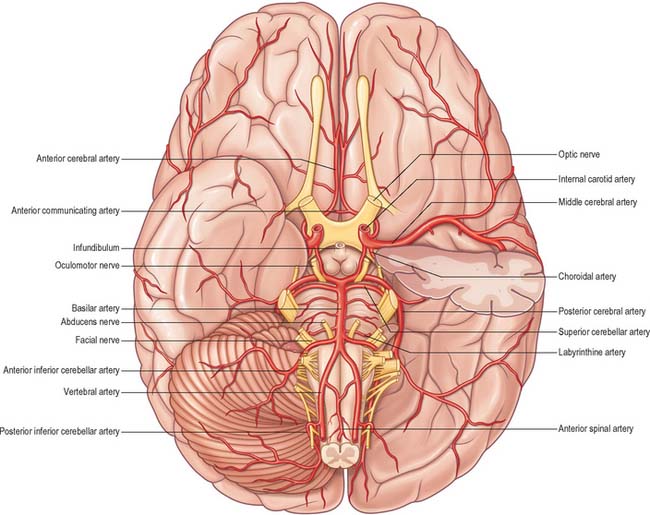
The internal carotid arteries (Fig. 17.1) and their major branches (the internal carotid system or ‘anterior’ circulation) supply blood to the majority of the forebrain. Some parts of the occipital and temporal lobes are supplied by branches of the vertebrobasilar system (see Fig. 17.5).
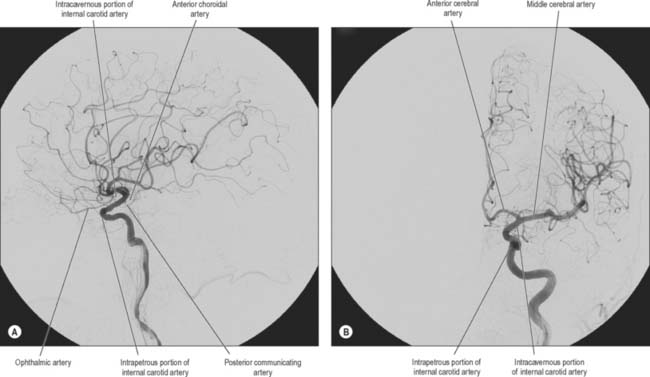
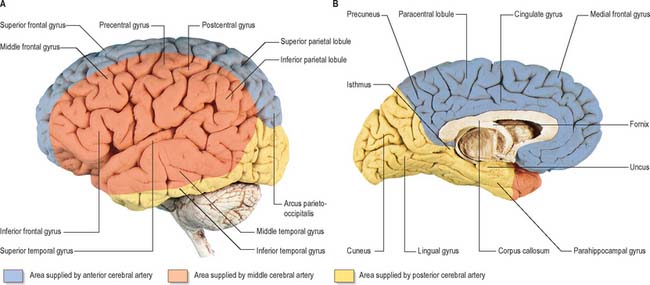
Fig. 17.5 The arteries supplying the left cerebral hemispheres. A, Lateral surface. B, Medial surface.
The internal carotid artery arises from the bifurcation of the common carotid artery, ascends in the neck and enters the carotid canal of the temporal bone. Its subsequent course is said to have petrous, cavernous and intracranial parts.
Petrous part
The petrous part of the internal carotid artery ascends in the carotid canal, curves anteromedially and then superomedially above the cartilage that fills the foramen lacerum, and enters the cranial cavity. It lies at first anterior to the cochlea and tympanic cavity, and is separated from the latter and the pharyngotympanic tube by a thin, bony lamella that is cribriform in the young and partly absorbed in old age. Further anteriorly, it is separated from the trigeminal ganglion by the thin roof of the carotid canal, although this is often deficient. The artery is surrounded by a venous plexus and by the carotid autonomic plexus, derived from the internal carotid branch of the superior cervical ganglion. The petrous part of the artery gives rise to two branches. The caroticotympanic artery is a small, occasionally double, vessel which enters the tympanic cavity by a foramen in the carotid canal and anastomoses with the anterior tympanic branch of the maxillary artery and the stylomastoid artery. The pterygoid artery is inconsistent: when present, it enters the pterygoid canal with the nerve of the same name, and anastomoses with a (recurrent) branch of the greater palatine artery.
Cavernous part
The cavernous part of the internal carotid artery ascends to the posterior clinoid process. It turns anteriorly to the side of the sphenoid within the cavernous sinus and then curves up medial to the anterior clinoid process, to emerge through the dural roof of the sinus. The oculomotor, trochlear, ophthalmic and abducens nerves are lateral to it within the cavernous sinus (see Fig. 27.9). Occasionally, the two clinoid processes form a bony ring round the artery.
This part of the artery gives off a number of small vessels. Cavernous branches supply the trigeminal ganglion, the walls of the cavernous and inferior petrosal sinuses and the nerves contained therein. A minute meningeal branch passes over the lesser wing of the sphenoid to supply the dura mater and bone in the anterior cranial fossa and also anastomoses with a meningeal branch of the posterior ethmoidal artery. Numerous small hypophysial branches supply the neurohypophysis, and are of particular importance because they form the pituitary portal system (Fig. 21.11).
Intracranial part
After piercing the dura mater, the internal carotid artery turns back below the optic nerve to run between it and the oculomotor nerve. It reaches the anterior perforated substance at the medial end of the lateral fissure and terminates by dividing into the anterior and middle cerebral arteries.
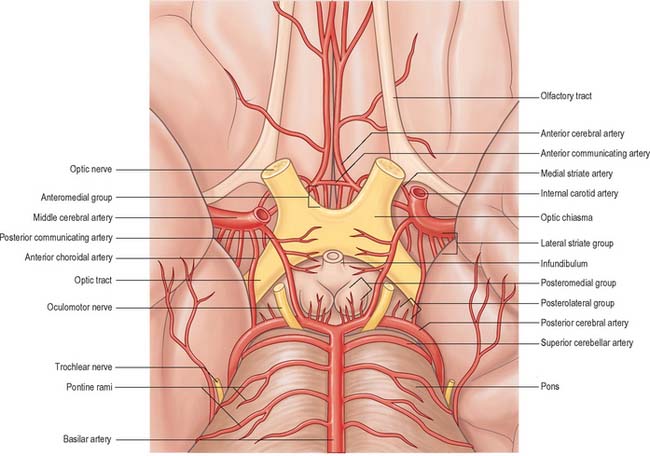
Several preterminal vessels leave the cerebral portion of the internal carotid. The ophthalmic artery arises from the anterior part of the internal carotid as it leaves the cavernous sinus, often at the point of piercing the dura, and enters the orbit through the optic canal. The posterior communicating artery (Fig. 17.2, Fig. 17.3) runs back from the internal carotid above the oculomotor nerve, and anastomoses with the posterior cerebral artery (a terminal branch of the basilar artery), thereby contributing to the circulus arteriosus around the interpeduncular fossa. The posterior communicating artery is usually very small. However, sometimes it is so large that the posterior cerebral artery is supplied via the posterior communicating artery rather than from the basilar artery (‘fetal posterior communicating artery’); it is often larger on one side only. Small branches from its posterior half pierce the posterior perforated substance together with branches from the posterior cerebral artery. Collectively they supply the medial thalamic surface and the walls of the third ventricle. The anterior choroidal artery leaves the internal carotid near its posterior communicating branch and passes back above the medial part of the uncus. It crosses the optic tract to reach and supply the crus cerebri of the midbrain, then turns laterally, recrosses the optic tract, and gains the lateral side of the lateral geniculate body, which it supplies with several branches. It finally enters the inferior horn of the lateral ventricle via the choroidal fissure and ends in the choroid plexus. This small, but important, vessel also contributes to the blood supply of the globus pallidus, caudate nucleus, amygdala, hypothalamus, tuber cinereum, red nucleus, substantia nigra, posterior limb of the internal capsule, optic radiation, optic tract, hippocampus and the fimbria of the fornix.
ANTERIOR CEREBRAL ARTERY
The anterior cerebral artery is the smaller of the two terminal branches of the internal carotid (Fig. 17.3).
Surgical nomenclature divides the vessel into three parts: A1 – from the termination of the internal carotid artery to the junction with the anterior communicating artery; A2 – from the junction with the anterior communicating artery to the origin of the callosomarginal artery; and A3 – distal to the origin of the callosomarginal artery; this segment is also known as the pericallosal artery.
The anterior cerebral artery starts at the medial end of the stem of the lateral fissure. It passes anteromedially above the optic nerve to the great longitudinal fissure where it connects with its fellow by a short transverse anterior communicating artery. The anterior communicating artery is about 4 mm in length and may be double. It gives off numerous anteromedial central branches that supply the optic chiasma, lamina terminalis, hypothalamus, para-olfactory areas, anterior columns of the fornix and the cingulate gyrus.
The two anterior cerebral arteries travel together in the great longitudinal fissure. They pass around the curve of the genu of the corpus callosum and then along its upper surface to its posterior end, where they anastomose with posterior cerebral arteries (Fig. 17.4). They give off cortical and central branches.
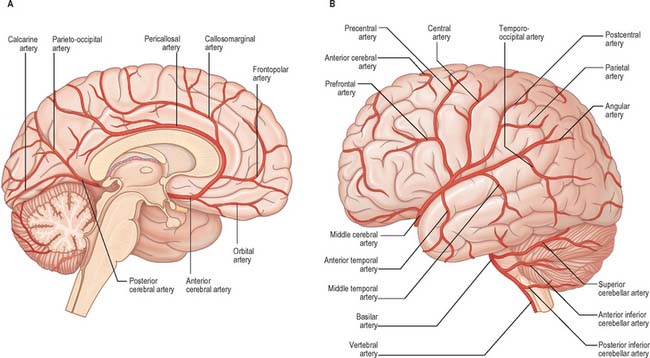
The cortical branches of the anterior cerebral artery are named according to their distribution. Two or three orbital branches ramify on the orbital surface of the frontal lobe and supply the olfactory cortex, gyrus rectus and medial orbital gyrus. Frontal branches supply the corpus callosum, cingulate gyrus, medial frontal gyrus and paracentral lobule. Parietal branches supply the precuneus, while the frontal and parietal branches both send twigs over the superomedial border of the hemisphere to supply a strip of territory on the superolateral surface (Fig. 17.5). Cortical branches of the anterior cerebral artery therefore supply the areas of the motor and somatosensory cortices that represent the lower limb.
Central branches of the anterior cerebral artery arise from its proximal portion and enter the anterior perforated substance (Fig. 17.3) and lamina terminalis. Collectively, they supply the rostrum of the corpus callosum, the septum pellucidum, the anterior part of the putamen, the head of the caudate nucleus and adjacent parts of the internal capsule. Immediately proximal or distal to its junction with the anterior communicating artery, the anterior cerebral artery gives rise to the medial striate artery which supplies the anterior part of the head of the caudate nucleus and adjacent regions of the putamen and internal capsule.
MIDDLE CEREBRAL ARTERY
The middle cerebral artery is the larger terminal branch of the internal carotid.
Surgical nomenclature divides the vessel into four parts: M1 – from the termination of the internal carotid artery to the bi/trifurcation, this segment is also known as the sphenoidal; M2 – the segment running in the lateral (Sylvian) fissure, also known as the insular; M3 – coming out of the lateral fissure, also known as the opercular; and M4 – cortical portions.
The middle cerebral artery runs at first in the lateral fissure, then posterosuperiorly on the insula, and divides into branches distributed to the insula and the adjacent lateral cerebral surface (Fig. 17.3, Fig. 17.4, Fig. 17.5). Like the anterior cerebral artery, it has cortical and central branches.
Cortical branches send orbital vessels to the inferior frontal gyrus and the lateral orbital surface of the frontal lobe. Frontal branches supply the precentral, middle and inferior frontal gyri. Two parietal branches are distributed to the postcentral gyrus, the lower part of the superior parietal lobule and the whole inferior parietal lobule. Two or three temporal branches supply the lateral surface of the temporal lobe. Cortical branches of the middle cerebral artery therefore supply the motor and somatosensory cortices that represent the whole of the body (other than the lower limb), the auditory area and the insula.
Small central branches of the middle cerebral artery, the lateral striate or lenticulostriate arteries, arise at its origin and enter the anterior perforated substance together with the medial striate artery. Lateral striate arteries ascend in the external capsule over the lower lateral aspect of the lentiform complex, then turn medially, traverse the lentiform complex and the internal capsule and extend as far as the caudate nucleus.
VERTEBRAL ARTERY
The vertebral arteries and their major branches (sometimes referred to as the ‘vertebrobasilar system’) essentially supply blood to the upper spinal cord, the brain stem and cerebellum and a significant but variable part of the posterior cerebral hemispheres (Fig. 17.6).
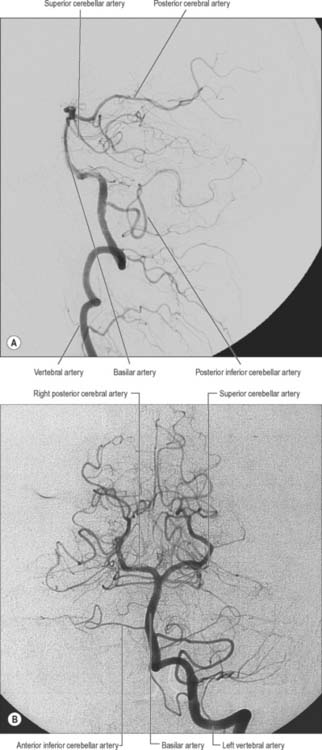
The vertebral arteries are derived from the subclavian arteries (see Ch. 30). They ascend through the neck in the foramina transversaria of the upper six cervical vertebrae and enter the cranial cavity through the foramen magnum, close to the anterolateral aspect of the medulla (Fig. 17.2). They converge medially as they ascend the medulla and unite to form the midline basilar artery at approximately the level of the junction between the medulla and pons.
One or two meningeal branches arise from the vertebral artery near the foramen magnum and ramify between the bone and dura mater in the posterior cranial fossa. They supply bone, diploë and the falx cerebelli.
A small anterior spinal artery arises near the end of the vertebral artery, and descends anterior to the medulla oblongata to unite with its fellow from the opposite side at mid-medullary level. The single trunk then descends on the ventral midline of the spinal cord, and is reinforced sequentially by small spinal rami from the vertebral, ascending cervical, posterior intercostal and first lumbar arteries, which all enter the vertebral canal via intervertebral foramina. Branches from the anterior spinal arteries and the beginning of their common trunk are distributed to the medulla oblongata.
The largest branch of the vertebral artery is the posterior inferior cerebellar artery (Fig. 17.6A). It arises near the lower end of the olive and then ascends behind the roots of the glossopharyngeal and vagus nerves to reach the inferior border of the pons. Here it curves and descends along the inferolateral border of the fourth ventricle before it turns laterally into the cerebellar vallecula between the hemispheres, and divides into medial and lateral branches. The medial branch runs back between the cerebellar hemisphere and inferior vermis, and supplies both. The lateral branch supplies the inferior cerebellar surface as far as its lateral border and anastomoses with the anterior inferior and superior cerebellar arteries (from the basilar artery). The trunk of the posterior inferior cerebellar artery supplies the medulla oblongata dorsal to the olivary nucleus and lateral to the hypoglossal nucleus and its emerging nerve roots. It also supplies the choroid plexus of the fourth ventricle and sends a branch lateral to the cerebellar tonsil to supply the dentate nucleus. The posterior inferior cerebellar artery is sometimes absent.
A posterior spinal artery usually arises from the posterior inferior cerebellar artery, but may originate directly from the vertebral artery near the medulla oblongata. It passes posteriorly and descends as two branches which lie anterior and posterior to the dorsal roots of the spinal nerves. These are reinforced by spinal twigs from the vertebral, ascending cervical, posterior intercostal and first lumbar arteries, all of which reach the vertebral canal by the intervertebral foramina, and sustain the posterior spinal arteries to the lower spinal levels.
Minute medullary arteries arise from the vertebral artery and its branches and are distributed widely to the medulla oblongata.
BASILAR ARTERY
The basilar artery is a large median vessel formed by the union of the vertebral arteries at the mid-medullary level (Figs 17.2, 17.3, 17.6). It lies in the pontine cistern, and follows a shallow median groove on the ventral pontine surface, extending to the upper border of the pons. It ends by dividing into two posterior cerebral arteries at a variable level behind the dorsum sellae, usually in the interpeduncular cistern.
Numerous small pontine branches arise from the front and sides of the basilar artery along its course and supply the pons. The long and slender labyrinthine (internal auditory) artery has a variable origin. It usually arises from the anterior inferior cerebellar artery, but variations in its origin include the lower part of the basilar artery, the superior cerebellar artery or, occasionally, the posterior inferior cerebellar artery. The labyrinthine artery accompanies the facial and vestibulocochlear nerves into the internal acoustic meatus and is distributed to the internal ear.
The anterior inferior cerebellar artery (Fig. 17.3) is given off from the lower part of the basilar artery and runs posterolaterally, usually ventral to the abducens, facial and vestibulocochlear nerves. It commonly exhibits a loop into the internal acoustic meatus below the nerves, and when this occurs, the labyrinthine artery may arise from the loop. The anterior inferior cerebellar artery supplies the inferior cerebellar surface anterolaterally and anastomoses with the posterior inferior cerebellar branch of the vertebral artery. A few branches supply the inferolateral parts of the pons and occasionally also supply the upper medulla oblongata.
The superior cerebellar artery (Figs 17.3, 17.6) arises near the distal portion of the basilar artery, immediately before the formation of the posterior cerebral arteries. It passes laterally below the oculomotor nerve, which separates it from the posterior cerebral artery, and curves round the cerebral peduncle below the trochlear nerve to gain the superior cerebellar surface. Here it divides into branches which ramify in the pia mater and supply this aspect of the cerebellum, and also anastomose with branches of the inferior cerebellar arteries. The superior cerebellar artery supplies the pons, pineal body, superior medullary velum and tela choroidea of the third ventricle.
POSTERIOR CEREBRAL ARTERY
The posterior cerebral artery (Figs 17.2, 17.3, 17.4) is a terminal branch of the basilar artery.
Surgical nomenclature divides the vessel into three parts: P1 – from the basilar bifurcation to the junction with the posterior communicating artery; P2 – from the junction with the posterior communicating artery to the portion in the perimesencephalic cistern; and P3 – the portion that runs in the calcarine fissure.
The posterior cerebral artery is larger than the superior cerebellar artery, from which it is separated near its origin by the oculomotor nerve, and, lateral to the midbrain, by the trochlear nerve. It passes laterally, parallel with the superior cerebellar artery, and receives the posterior communicating artery. It then winds round the cerebral peduncle and reaches the tentorial cerebral surface, where it supplies the temporal and occipital lobes. Like the anterior and middle cerebral arteries, the posterior cerebral artery has cortical and central branches.
The cortical branches of the posterior cerebral artery are named according to their distribution. Temporal branches, usually two, are distributed to the uncus, parahippocampal, medial and lateral occipitotemporal gyri. Occipital branches supply the cuneus, lingual gyrus and posterolateral surface of the occipital lobe. Parieto-occipital branches supply the cuneus and precuneus. The posterior cerebral artery supplies the visual areas of the cerebral cortex and other structures in the visual pathway.
The central branches supply subcortical structures. Several small posteromedial central branches arise from the beginning of the posterior cerebral artery (Fig. 17.3) and, together with similar branches from the posterior communicating artery, pierce the posterior perforated substance to supply the anterior thalamus, subthalamus, lateral wall of the third ventricle and globus pallidus. One or more posterior choroidal branches pass over the lateral geniculate body and supply it before entering the posterior part of the inferior horn of the lateral ventricle via the lower part of the choroidal fissure. Branches also curl round the posterior end of the thalamus and pass through the transverse fissure, or go to the choroid plexus of the third ventricle, or traverse the upper choroidal fissure. Collectively these branches supply the choroid plexuses of the third and lateral ventricles and the fornix. Small posterolateral central branches arise from the posterior cerebral artery beyond the cerebral peduncle and supply the peduncle and the posterior thalamus, superior and inferior colliculi, pineal gland and medial geniculate body.
CIRCULUS ARTERIOSUS
The circulus arteriosus (circle of Willis) is a large arterial anastomosis which unites the internal carotid and vertebrobasilar systems (Figs 17.3, 17.6B). It lies in the subarachnoid space within the interpeduncular cistern, and surrounds the optic chiasma and infundibulum. Anteriorly, the anterior cerebral arteries, derived from the internal carotid arteries, are linked by the small anterior communicating artery. Posteriorly, the two posterior cerebral arteries, formed by the division of the basilar artery, are joined to the ipsilateral internal carotid artery by a posterior communicating artery.
There is considerable individual variation in the pattern and calibre of vessels that make up the circulus arteriosus. Although a complete circular channel almost always exists, one vessel is usually sufficiently narrowed to reduce its role as a collateral route and the circle is rarely functionally complete. Cerebral and communicating arteries individually may all be absent, variably hypoplastic, double or even triple. The haemodynamics of the circle are influenced by variations in the calibre of communicating arteries and in the segments of the anterior and posterior cerebral arteries which lie between their origins and their junctions with the corresponding communicating arteries. The greatest variation in calibre between individuals occurs in the posterior communicating artery, which is normally very small, so that only limited flow is possible between the anterior and posterior circulations. Commonly, the diameter of the precommunicating part of the posterior cerebral artery is larger than that of the posterior communicating artery, in which case the blood supply to the occipital lobes is mainly from the vertebrobasilar system. However, sometimes the diameter of the precommunicating part of the posterior cerebral artery is smaller than that of the posterior communicating artery, in which case the blood supply to the occipital lobes is mainly from the internal carotids via the posterior communicating arteries. Since the primary purpose of the vascular circle is to provide anastomotic channels if one vessel is occluded, it is important to note that a normal-sized posterior communicating artery cannot usually fulfil this role. Agenesis or hypoplasia of the initial segment of the anterior cerebral artery are more frequent than anomalies in the anterior communicating artery and contribute to defective circulation in about a third of individuals.
Aneurysms are balloon-like swellings which occur on arteries as a result of defects in the vessel wall. They are most commonly found on the vessels of the circulus arteriosus, particularly at or near the junctions of vessels. Aneurysms on the internal carotid artery near its termination may compress the lateral aspect of the optic chiasma, and compromise axons derived from the temporal side of the ipsilateral retina, which causes a defect in the nasal visual field. Aneurysms in the vicinity of the oculomotor nerve, e.g. on the posterior communicating artery, superior cerebellar artery, or the tip of the basilar artery, can cause third nerve palsy by compression (Fig. 17.7).
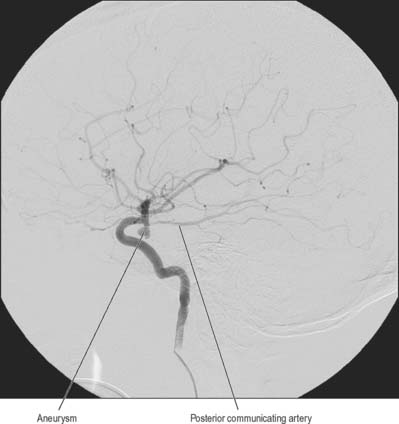
Bleeding into the subarachnoid compartment, subarachnoid haemorrhage, is the most common pathology that involves the subarachnoid space. There are many causes; in adults the commonest is rupture of an aneurysm of the intracranial vessels that run within the subarachnoid space. A person who has had a subarachnoid haemorrhage usually complains of a very sudden onset of headache that is frequently described as being their ‘worst ever headache’ or ‘like being hit on the head with a hammer’. When this is suspected the first investigation is an X-ray CT examination which has a very high sensitivity for detecting fresh haemorrhage. Blood will be seen in the basal cisterns and entering the depths of the cortical sulci, i.e. delineating the anatomy of the subarachnoid space, a presentation that permits an accurate diagnosis (Fig. 17.8). In marked contrast, the spread of blood in a subdural haemorrhage is limited by the arachnoid mater on its deep surface, and the blood therefore remains on the surface of the brain.
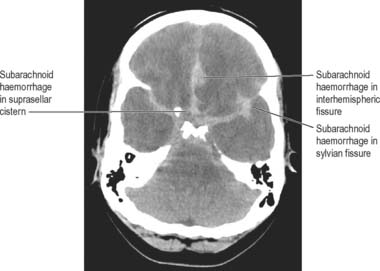
Fig. 17.8 Axial CT image (without intravenous contrast agent) at the level of posterior clinoids. There is abnormally high attenuation in the basal cisterns and subarachnoid space of the interhemispheric fissure, indicating an acute subarachnoid haemorrhage, which was subsequently shown to be due to a ruptured aneurysm on the anterior communicating artery.
CENTRAL OR PERFORATING ARTERIES
Numerous small central (perforating or ganglionic) arteries arise from the circulus arteriosus, or from vessels near it (Fig. 17.3). Many of these enter the brain through the anterior and posterior perforated substances. Central branches supply nearby structures on or near the base of the brain together with the interior of the cerebral hemisphere including the internal capsule, basal ganglia and thalamus. These branches form four principal groups. The anteromedial group arises from the anterior cerebral and anterior communicating arteries and passes through the medial part of the anterior perforated substance. These arteries supply the optic chiasma, lamina terminalis, anterior, preoptic and supraoptic areas of the hypothalamus, septum pellucidum, paraolfactory areas, anterior columns of the fornix, cingulate gyrus, rostrum of the corpus callosum and the anterior part of the putamen and the head of the caudate nucleus. The posteromedial group comes from the entire length of the posterior communicating artery and from the proximal portion of the posterior cerebral artery. Anteriorly, these arteries supply the hypothalamus and pituitary gland, and the anterior and medial parts of the thalamus via thalamoperforating arteries. Caudally, branches of the posteromedial group supply the mammillary bodies, subthalamus, the lateral wall of the third ventricle, including the medial thalamus, and the globus pallidus. The anterolateral group is mostly comprised of branches from the proximal part of the middle cerebral artery that are also known as striate, lateral striate or lenticulostriate arteries. They enter the brain through the anterior perforated substance and supply the posterior striatum, lateral globus pallidus and the anterior limb, genu and posterior limb of the internal capsule. The medial striate artery, derived from the middle or anterior cerebral arteries, supplies the rostral part of the caudate nucleus and putamen and the anterior limb and genu of the internal capsule. The posterolateral group is derived from the posterior cerebral artery distal to its junction with the posterior communicating artery, and supplies the cerebral peduncle, colliculi, pineal gland and, via thalamogeniculate branches, the posterior thalamus and medial geniculate body.
REGIONAL ARTERIAL SUPPLY OF THE BRAIN
Brain stem
The medulla oblongata is supplied by the branches of the vertebral, anterior and posterior spinal, posterior inferior cerebellar and basilar arteries, which enter along the anterior median fissure and the posterior median sulcus. Vessels that supply the central substance enter along the rootlets of the glossopharyngeal, vagus, accessory and hypoglossal nerves. There is an additional supply via a pial plexus from the same main arteries.
The pons is supplied by the basilar artery and the anterior inferior and superior cerebellar arteries. Direct branches from the basilar artery enter the pons along the ventral medial groove (basilar sulcus). Other vessels enter along the trigeminal, abducens, facial and vestibulcochlear nerves and from the pial plexus.
The midbrain is supplied by the posterior cerebral, superior cerebellar and basilar arteries. The crura cerebri are supplied by vessels entering on their medial and lateral sides. The medial vessels enter the medial side of the crus and also supply the superomedial part of the tegmentum, including the oculomotor nucleus, and lateral vessels supply the lateral part of the crus and the tegmentum. The colliculi are supplied by three vessels on each side from the posterior cerebral and superior cerebellar arteries. An additional supply to the crura, and the colliculi and their penduncles, comes from the posterolateral group of central branches of the posterior cerebral artery.
Cerebellum
The cerebellum is supplied by the posterior inferior, anterior inferior and superior cerebellar arteries. The cerebellar arteries form superficial anastomoses on the cortical surface. Anastomoses between deeper, subcortical, branches have been postulated.
The choroid plexus of the fourth ventricle is supplied by the posterior inferior cerebellar arteries.
Optic chiasma, tract and radiation
The blood supplies of the optic chiasma, tract and radiation are of considerable clinical importance. The chiasma is supplied in part by the anterior cerebral arteries but its median zone depends upon rami from the internal carotid arteries reaching it via the stalk of the hypophysis. The anterior choroidal and posterior communicating arteries supply the optic tract, and the optic radiation receives blood through deep branches of the middle and posterior cerebral arteries.
Diencephalon
The thalamus is supplied chiefly by branches of the posterior communicating, posterior cerebral and basilar arteries. A contribution from the anterior choroidal artery is often noted, but this has been disputed. The medial branch of the posterior choroidal artery supplies the posterior commissure, habenular region, pineal gland and medial parts of the thalamus, including the pulvinar. Small central branches, which arise from the circulus arteriosus and its associated vessels, supply the hypothalamus. The pituitary gland is supplied by hypophysial arteries derived from the internal carotid artery and the anterior cerebral and anterior communicating arteries supply the lamina terminalis.
The choroid plexuses of the third and lateral ventricles are supplied by branches of the internal carotid and posterior cerebral arteries.
Basal ganglia
The majority of the arterial supply to the basal ganglia comes from the striate arteries, which are branches from the roots of the anterior and middle cerebral arteries. They enter the brain through the anterior perforated substance and also supply the internal capsule. The caudate nucleus receives blood additionally from the anterior and posterior choroidal arteries. The posteroinferior part of the lentiform complex is supplied by the thalamostriate branches of the posterior cerebral artery. The anterior choroidal artery, a preterminal branch of the internal carotid artery, contributes to the blood supply of both segments of the globus pallidus and the caudate nucleus. Famously, the ligation of this vessel during a neurosurgical procedure on a patient suffering from Parkinson’s disease led to alleviation of the Parkinsonian symptoms, presumably as a consequence of infarction of the globus pallidus. This chance observation led to the initiation of pallidal surgery (pallidotomy) for this condition (see also Ch. 22).
Internal capsule
The internal capsule is supplied by central, or perforating, arteries which arise from the circulus arteriosus and its associated vessels. These include the lateral and medial striate arteries which come from the middle and anterior cerebral arteries and which also supply the basal ganglia. The lateral striate arteries supply the anterior limb, genu and much of the posterior limb of the internal capsule and are commonly involved in ischaemic and haemorrhagic stroke. One of the larger striate branches of the middle cerebral artery is known as ‘Charcot’s artery of cerebral haemorrhage’. The medial striate artery, a branch of the proximal part of either the middle or anterior cerebral artery, supplies the anterior limb and genu of the internal capsule and the basal ganglia. The anterior choroidal artery also contributes to the supply of the ventral part of the posterior limb and the retrolenticular (retrolentiform) part of the internal capsule.
Ischaemic stroke
Stroke refers to the clinical syndrome of a rapidly developing focal neurological deficit that is not due to seizure activity. If the cause is lack of, or reduced, blood supply to a portion of the brain then the term ischaemic stroke (see Fig. 17.9) is used, as opposed to haemorrhagic stroke e.g. subarachnoid haemorrhage. The lack of blood flow can be due to pathology in the vessel lumen, such as thrombosis or embolus (common), pathology outside the blood vessel, such as occlusion from mass effect of a tumour or haematoma (rare), or pathology of the vessel wall, such as inflammatory or infective arteritides (rare). The symptoms and signs of ischaemic stroke depend on the location and extent of the arterial infarction. In certain locations, even a small volume stroke can have devastating effects. For example, the internal capsule and most of the adjacent basal ganglia are supplied by small perforating arteries, which are a common site for ischaemic stroke. Corticobulbar and corticospinal motor pathways and third-order thalamocortical sensory fibres all pass through the internal capsule, so that it is effectively the only route connecting the cerebral cortex with other regions of the neuraxis. The neurological deficits that result from stroke in the internal capsule, therefore, may include contralateral spastic hemiparesis, contralateral hemisensory loss and psychological deficits.
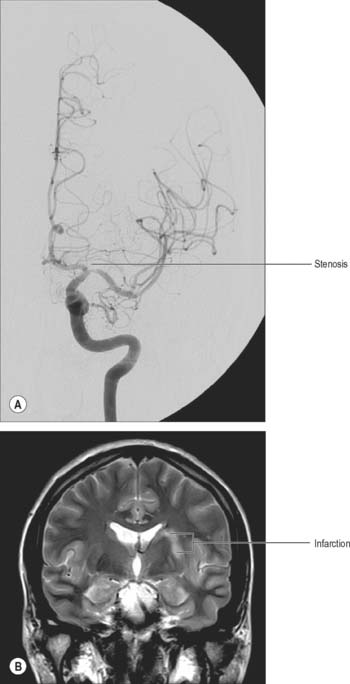
Fig. 17.9 A, Frontal projection of a catheter angiogram of the left internal carotid artery, showing a focal narrowing at the termination of the left internal carotid artery and the origin of the left middle cerebral artery. B, Coronal T2-weighted image of the brain at the level of the basal ganglia. There is abnormal high signal in the left putamen and adjacent white matter, consistent with infarction. It is likely that the infarction was due to occlusion of small perforating vessels originating in the area of the stenosis and supplying the basal ganglia.
It is possible for a major vessel, such as the internal carotid artery in the neck, to become occluded and the individual to remain asymptomatic, if an appropriate flow of blood can be recruited to supply the affected brain region from other sources (collateral supply). The primary role of the circle of Willis is to allow for this eventuality by redirecting blood from other sources such as the contralateral internal carotid and vertebrobasilar system in the example given above. If the circle of Willis is complete, two or even three of the four major arteries supplying the brain can occlude without producing an ischaemic stroke. Unfortunately the circle of Willis is rarely complete (a finding in approximately 15% of individuals): ischaemic stroke is therefore often caused because the collateral supply is inadequate. For example, an individual in whom neither anterior nor posterior communicating arteries are patent will suffer an ischaemic stroke if there is occlusion of one internal carotid artery, because the proximal anterior and middle cerebral arteries cannot receive collateral flow from the incomplete circle of Willis. Some collateral flow may be available from anastomoses between distal branches of the external carotid artery (e.g. middle meningeal artery), but invariably this is not sufficient to supply the whole hemisphere.
Somewhat paradoxically, the more distal the occlusion occurs in the cerebral vasculature the more likely it is to cause an ischaemic stroke (although the volume of infarcted tissue will be less). This is because the major capacity to form collateralization is proximal, mainly at the circle of Willis. For example, an embolus formed in the proximal internal carotid artery may travel to the M1 portion of the ipsilateral middle cerebral artery and occlude the vessel. There is very little scope for collateralisation at this site and an ischaemic stroke is unavoidable, unless the embolus can be cleared and flow restored in good time.
Cerebral cortex
The entire blood supply of the cerebral cortex comes from cortical branches of the anterior, middle and posterior cerebral arteries (Fig. 17.4, Fig. 17.5). In general, long branches traverse the cortex and penetrate the subjacent white matter for 3 or 4 cm without communicating. Short branches are confined to the cerebral cortex, and form a compact network in the middle zone of the grey matter, whereas the outer and inner zones are sparingly supplied. Although adjacent vessels anastomose on the surface of the brain, they become end arteries as soon as they enter it. In general, superficial anastomoses only occur between microscopic branches of the cerebral arteries, and there is little evidence that they can provide an effective alternative circulation after the occlusion of larger vessels.
The lateral surface of the hemisphere is mainly supplied by the middle cerebral artery. This includes the territories of the motor and somatosensory cortices which represent the whole of the body, apart from the lower limb, and also the auditory cortex and language areas. The anterior cerebral artery supplies a strip next to the superomedial border of the hemisphere, as far back as the parieto-occipital sulcus. The occipital lobe and most of the inferior temporal gyrus (excluding the temporal pole) are supplied by the posterior cerebral artery.
Medial and inferior surfaces of the hemisphere are supplied by the anterior, middle and posterior cerebral arteries. The area supplied by the anterior cerebral artery is the largest, and extends almost to the parieto-occipital sulcus and includes the medial part of the orbital surface. The rest of the orbital surface and the temporal pole are supplied by the middle cerebral artery, and the remaining medial and inferior surfaces are supplied by the posterior cerebral artery.
Near the occipital pole, the junctional zone between the territories of the middle and posterior cerebral arteries corresponds to the visual (striate) cortex which receives information from the macula. When the posterior cerebral artery is occluded, a phenomenon known as ‘macular sparing’ may occur in which vision with the central part of the retina is preserved. Collateral circulation of blood from branches of the middle cerebral artery into those of the posterior cerebral artery may account for this phenomenon. In addition, in some individuals, the middle cerebral artery may itself supply the macular area.
Regions of the brain lying at the extreme edges of the major cerebral arterial territories are called watershed areas: they are the first to be deprived of sufficient blood flow in the event of cerebral hypoperfusion. Ischaemic infarcts of the cortex and adjacent subcortical white matter in the border zones between these territories are known as watershed, boundary zone, or border zone infarcts. There is some evidence that a second arterial border zone, the internal arterial border zone, exists within the white matter of the centrum semiovale and corona radiata (between the lenticulostriate perforators and the deep penetrating branches of the distal middle cerebral artery).
Cerebral blood flow
The brain is devoid of either glucose stores or a means of storing oxygen and is, therefore, dependent minute-by-minute upon an adequate blood supply. It has a high metabolic rate in comparison to other organs, which reflects the metabolic demands of constant neural activity. The blood supply of grey matter is more copious than that of white matter.
Cerebral blood flow in the human brain is approximately 50 ml g−1 min−1. Global cerebral blood flow is autoregulated, i.e. it remains constant in normal individuals despite variations in mean arterial blood pressure over a range of 8.7–18.7 kPa (65–140 mmHg). If the blood pressure falls below this range, cerebral blood flow decreases. Alternatively, if the pressure rises above this range, cerebral blood flow may increase. Arterial and arteriolar intraluminal pressure directly control contraction of intramural muscle, so that an increase in arterial pressure, for example, causes arterial constriction, and blood flow remains constant.
Although autoregulation normally ensures that global cerebral blood flow remains constant, regional blood flow varies in response to the level of neural activity and, thus, to local metabolic demand. This has been demonstrated for many brain areas including the motor and sensory cortical regions, areas involved in convulsive activity and even cortical areas involved in complex thought processes. The principal local factors affecting regional blood flow are the local hydrogen ion (H+) or carbon dioxide concentration, which cause arterial dilatation by both a direct action on the endothelium and through indirect mediators, such as nitric oxide.
Not all substances circulating in arterial blood have access to the brain parenchyma. Particulate matter, such as bacteria, is excluded. In general, lipophilic molecules and small molecules, such as oxygen and carbon dioxide, can cross the blood–brain barrier but hydrophilic ones (excluding glucose) cannot. The cellular basis for the blood–brain barrier is discussed in Chapter 3.
VENOUS DRAINAGE OF THE BRAIN
The venous drainage of the brain occurs through a complex system of deep and superficial veins. These veins possess no valves and have thin walls devoid of muscular tissue. They pierce the arachnoid mater and the inner layer of the dura mater to open into the dural venous sinuses (see Chapter 27).
VENOUS DRAINAGE OF THE POSTERIOR FOSSA
The veins of the brain stem form a superficial venous plexus deep to the arteries.
Veins of the medulla oblongata drain into the veins of the spinal cord or the adjacent dural venous sinuses, or into variable radicular veins which accompany the last four cranial nerves to either the inferior petrosal or occipital sinuses, or to the superior bulb of the jugular vein. Anterior and posterior median medullary veins may run along the anterior median fissure and posterior median sulcus, to become continuous with the spinal veins in corresponding positions. Pontine veins, which may include a median vein and a lateral vein on each side, drain into the basal vein, cerebellar veins, the petrosal sinuses, transverse sinus or the venous plexus of the foramen ovale. Veins of the midbrain join the great cerebral vein or basal vein.
The veins of the cerebellum drain mainly into sinuses adjacent to them or, from the superior surface, into the great cerebral vein. The cerebellar veins course on the cerebellar surface, and consist of superior and inferior groups. Superior cerebellar veins either run anteromedially across the superior vermis to the straight sinus or great cerebral vein, or they run laterally to the transverse and superior petrosal sinuses. Inferior cerebellar veins include a small median vessel that runs backwards on the inferior vermis to enter the straight or sigmoid sinus. Laterally coursing vessels join the inferior petrosal and occipital sinuses.
VENOUS DRAINAGE OF THE CEREBRAL HEMISPHERE
External and internal cerebral veins drain the surfaces and the interior of the cerebral hemisphere (Figs 17.10, 17.11, 17.12, 17.13).
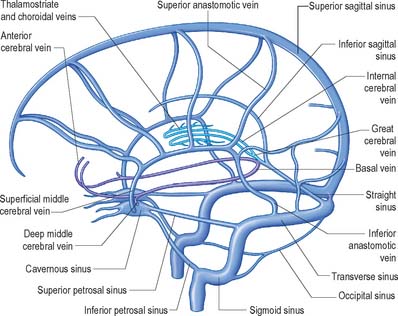
Fig. 17.10 The cerebral venous system showing the principal superficial and deep veins of the brain and their relationship to the dural venous sinuses, viewed from the left side.
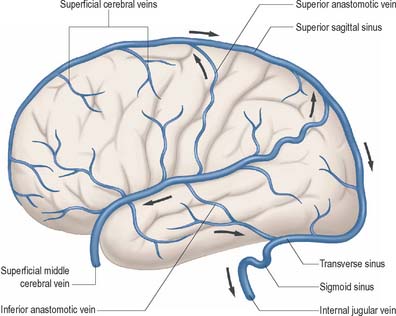
Fig. 17.11 The external (superficial) cerebral veins of the left hemisphere and their relationship to the dural venous sinuses.
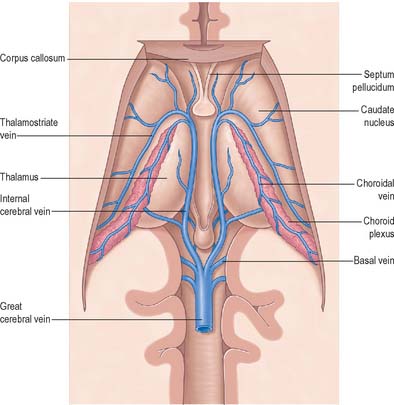
Fig. 17.12 The internal (deep) cerebral veins, viewed from above after removal of the central portion of the corpus callosum.
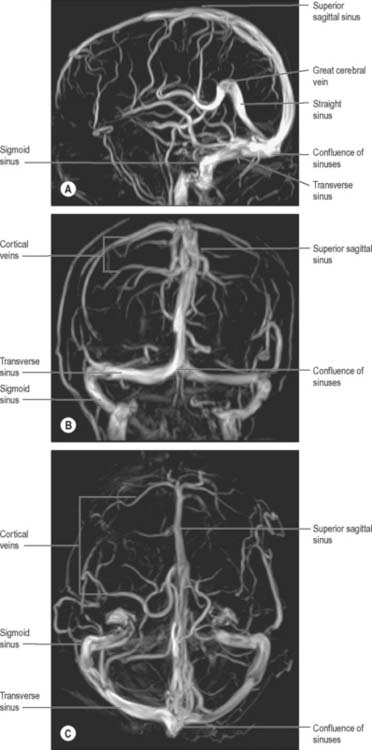
Fig. 17.13 Phase contrast magnetic resonance venograms taken at 3.0T in A, lateral; B, frontal and C, superior projections.
External cerebral veins may be divided into three groups, namely superior, middle and inferior.
Eight to 12 superior cerebral veins drain the superolateral and medial surfaces of each hemisphere. They mainly follow the sulci, although some do pass across gyri. They ascend to the superomedial border of the hemisphere, where they receive small veins from the medial surface, and then open into the superior sagittal sinus. Superior cerebral veins in the anterior part of the hemisphere join the sinus almost at right angles. The larger posterior veins are directed obliquely forwards, against the direction of flow in the sinus, an arrangement which may resist their collapse when intracranial pressure is raised.
The superficial middle cerebral vein drains most of the lateral surface of the hemisphere, and follows the lateral fissure to end in the cavernous sinus. A superior anastomotic vein runs posterosuperiorly between the superficial middle cerebral vein and the superior sagittal sinus, thus connecting the superior sagittal and cavernous sinuses. An inferior anastomotic vein courses over the temporal lobe and connects the superficial middle cerebral vein to the transverse sinus. The deep middle cerebral vein drains the insular region and joins the anterior cerebral and striate veins to form a basal vein. Regions drained by the anterior cerebral and striate veins correspond approximately to those supplied by the anterior cerebral artery and the central branches which enter the anterior perforated substance. The basal veins pass back alongside the interpeduncular fossa and midbrain, receive tributaries from this vicinity and join the great cerebral vein.
Inferior cerebral veins on the orbital surface of the frontal lobe join the superior cerebral veins and thus drain to the superior sagittal sinus. Those on the temporal lobe anastomose with basal veins and middle cerebral veins, and drain to the cavernous, superior petrosal and transverse sinuses.
The basal vein begins at the anterior perforated substance by the union of a small anterior cerebral vein, which accompanies the anterior cerebral artery, a deep middle cerebral vein, which receives tributaries from the insula and neighbouring gyri and runs in the lateral cerebral fissure, and striate veins, which emerge from the anterior perforated substance. The basal vein passes back round the cerebral peduncle to the great cerebral vein and receives tributaries from the interpeduncular fossa, inferior horn of the lateral ventricle, parahippocampal gyrus and midbrain.
The internal cerebral vein drains the deep parts of the hemisphere and the choroid plexuses of the third and lateral ventricles. It is formed near the interventricular foramen, behind the column of the fornix, primarily by union of the thalamostriate and choroidal veins, although numerous smaller veins from surrounding structures also converge here. The thalamostriate vein runs anteriorly, between the caudate nucleus and thalamus, and receives many tributaries from both areas. The choroidal vein runs a convoluted course along the whole choroid plexus, and receives veins from the hippocampus, fornix, corpus callosum and adjacent structures. After their formation, the two internal cerebral veins travel back parallel to one another beneath the splenium of the corpus callosum, where they unite to form the great cerebral vein. The great cerebral vein is a short median vessel which curves sharply up around the splenium of the corpus callosum and opens into the anterior end of the straight sinus after receiving the right and left basal veins.
Intracranial venosinus thrombosis
One of the most potentially serious pathologies affecting the intracranial venous system is venosinus thrombosis. In this condition, blood clots within the dural sinuses and/or intracranial veins, and the resulting obstruction of venous drainage, may produce life-threatening complications. In many cases the cause of the venosinus thrombosis is not found: trauma involving the dural sinuses or infections in the vicinity of the sinuses (e.g. mastoiditis) present significant risk. Conditions that increase the coagulability of the blood, such as the oral contraceptive pill, pregnancy, dehydration and inborn abnormalities of the coagulation pathways, are also risk factors.
At its severest, individuals with intracranial venosinus thrombosis present moribund, in coma or with major seizure activity: in these situations the prognosis is poor. The clot within the dural sinuses and/or cortical veins obstructs the drainage of venous blood from the brain and causes venous hypertension. As the pressure in the venous system approaches the arterial pressure, the blood perfusing the affected parts of the brain decreases. If it decreases below a critical level the brain tissue will become ischaemic and may die (venous infarction). As the arterial supply to the brain is still present, the infarcted areas of the brain frequently undergo major haemorrhage and this is often the factor that kills the patient.
Although they may occur together, clinicians distinguish between thrombosis that affects the ‘superficial’ and ‘deep’ venous systems (Fig. 17.10). Superficial thrombosis usually occurs in the superior sagittal sinus (with or without involvement of the cortical veins), whilst deep intracranial thrombotic disease usually affects the vein of Galen and straight sinus. The parts of the cerebral hemispheres affected are different in the two situations; superficial thrombosis affects the cerebral hemispheres (particularly the white matter) and deep thrombosis affects the thalami and basal ganglia. However, one common feature is the tendency for bilateral involvement, because all of the main draining venous channels are in the midline and accept venous drainage from both hemispheres.
If the extent of the intracranial venosinus thrombosis is less severe, or if alternative venous drainage has been accommodated by reasonable collateral channels, the patient may have more modest symptoms such as headaches or visual disturbance. Those symptoms are due to raised intracranial pressure and if untreated may become chronic.
Andeweg J. The anatomy of collateral venous flow from the brain and its value in aetiological interpretation of intracranial pathology. Neurorad. 1996;38:621-628.
Includes a brief historical review..
Bogousslavsky J, Caplan L. Stroke Syndromes, 2nd edn., Cambridge: Cambridge University Press, 2002.
Duvernoy HM, Delon S, Vannson JL. Cortical blood vessels of the human brain. Brain Res Bull. 1981;7:519-579.
Duvernoy H, Delon S, Vannson JL. The vascularization of the human cerebellar cortex. Brain Res Bull. 1983;11:419-480.
Kaplan HA, Ford DH. The Brain Vascular System. Amsterdam: Elsevier, 1966.
Plets C, De Reuck J, Vander Eecken H, Van den Bergh R. The vascularization of the human thalamus. Acta Neurol Belg. 1970;70:687-770.
Puchades-Orts A, Nombela-Gomez M, Ortu-o-Pacheco G. Variation in form of the circle of Willis. Some anatomical and embryological considerations. Anat Rec. 1976;185:119-123.
Sengupta RP, McAllister VL. Subarachnoid Haemorrhage. Berlin: Springer-Verlag; 1986:9-31.