 CHAPTER 6
CHAPTER 6The Nervous System
The nervous system may be divided into the central nervous system, consisting of the brain and spinal cord, and the peripheral nervous system, composed of cranial, spinal, and named nerves.
MENINGES
The brain and spinal cord are covered by three membranes of connective tissue, the meninges (Figs. 6-1, 6-20, 6-22). The dura mater, or pachymeninx, is the thickest of these and the most external. Throughout most of the vertebral canal, the dura is separated from the periosteum of the bony canal by the loose connective tissue of the epidural space, which often contains fat.
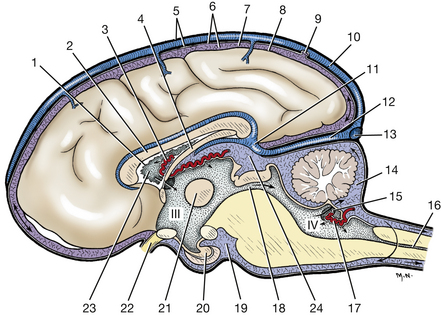
Fig. 6-1 Meninges and ventricles of brain, median plane.(Arrows indicate flow of cerebrospinal fluid.)
As the spinal cord approaches the brain stem, the dura adheres to the periosteum in the first one or two cervical vertebrae and to the atlanto-occipital membrane. Inside the cranial cavity, the dura and periosteum are fused. Starting at the dorsal margin, free the hemisectioned brain from the skull of the sagittally split head. On one half of the head, a fold of dura will be found extending ventrally from the midline in the longitudinal cerebral fissure between the two cerebral hemispheres. This is the falx cerebri, which contains the dorsal sagittal sinus. Remove the falx cerebri to allow reflection of the cerebral hemisphere.
The pia mater and arachnoid (the leptomeninges) are the other two connective tissue coverings of the central nervous system. The pia mater adheres to the external surface of the nervous tissue. The arachnoid in the live animal is attached to the dura and sends delicate trabeculae to the pia. These trabeculae closely invest the blood vessels that course on the surface of the pia. The space between the pia and the arachnoid is the subarachnoid space, which is filled with cerebrospinal fluid that surrounds the entire central nervous system. In the embalmed specimen, the arachnoid breaks free from the dura and collapses onto the pia of the central nervous system.
Subarachnoid cisterns occur in areas where the arachnoid and pia are more widely separated. The largest cistern is the cerebellomedullary cistern, located in the angle between the cerebellum and medulla. Cerebrospinal fluid may be obtained from this cistern (Fig. 6-1) by means of a needle puncture through the atlanto-occipital membrane.
ARTERIES
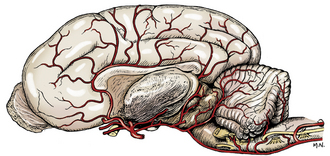
Examine the arteries to the brain on the latex-injected specimen (Figs. 5-50, 5-51, 6-2, 6-3). The arteries to the cerebrum and cerebellum are branches from the vessels on the ventral surface of the brain. These major arteries are not accompanied by veins. The basilar artery is formed by the terminal branches of the vertebral arteries, which enter the floor of the vertebral canal through the lateral vertebral foramina of the atlas.
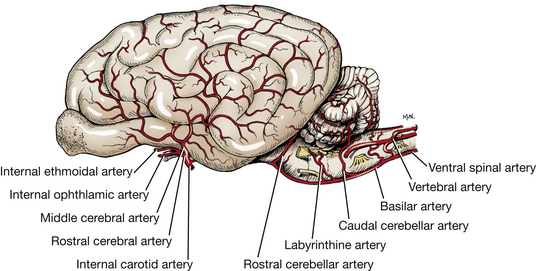
It is continuous caudally with the ventral spinal artery of the spinal cord. The basilar artery courses along the midline of the ventral surface of the medulla and pons and then divides into two branches that form the caudal portion of the arterial circle of the brain.
The internal carotid arteries are the other main source of blood to the arterial circle of the brain. After traversing the carotid canal in the tympanic part of the temporal bone and forming a loop at the foramen lacerum, each internal carotid artery enters the middle cranial fossa under the rostral end of the petrous temporal bone. It courses rostrally through the cavernous venous sinus beside the hypophyseal fossa and hypophysis. Between the hypophysis and the optic chiasm, it emerges through the dura over the sinus entering the subarachnoid space and divides into a middle cerebral, rostral cerebral, and caudal communicating artery. The small caudal communicating arteries from each internal carotid course caudally and join the terminal branches of the basilar artery. Rostrally, the two rostral cerebral arteries anastomose, completing the arterial circle on the ventral surface of the brain.
The cerebral arterial circle surrounds the pituitary gland, which receives small branches from the circle as well as directly from the internal carotid artery. Put the two halves of the latex-injected brain together to observe this arterial circle. Using the hemisectioned latex-injected brain, identify and trace the following vessels.
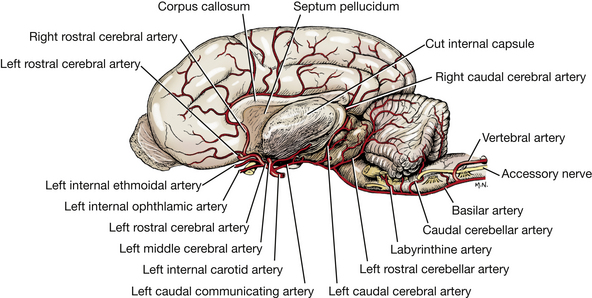
The rostral cerebral artery is a terminal branch of the internal carotid artery at the rostral aspect of the circle. It courses dorsally, lateral to the optic chiasm, and continues dorsally between the two frontal lobes in the longitudinal cerebral fissure. It courses dorsally over the fibers that connect the two cerebral hemispheres (corpus callosum) and caudally along the dorsal surface of the corpus callosum adjacent to the cerebral gyri, which these vessels supply.
Internal ethmoidal and internal ophthalmic arteries branch from the rostral cerebral artery. The internal ethmoidal artery anastomoses with the external ethmoidal artery and leaves the cranial cavity through the cribriform plate of the ethmoid bone to supply structures in the nasal cavity. The internal ophthalmic artery anastomoses with a branch of the external ophthalmic on the optic nerve. This is the source of the long ciliary arteries that follow the optic nerve to the eyeball, which they supply. Do not dissect these arteries.
The middle cerebral artery arises from the arterial circle at the level of the rostral aspect of the pituitary gland. It courses laterally, rostral to the piriform lobe on the ventral surface of the olfactory peduncle. It continues dorsolaterally over the cerebral hemisphere, where it branches to supply the lateral surface.
The caudal cerebral artery arises from the caudal communicating artery at the level of the caudal aspect of the pituitary gland, rostral to the oculomotor nerve. The artery courses caudodorsally, following the optic tract over the lateral aspect of the thalamus to the longitudinal fissure. It passes rostrally on the corpus callosum to supply the medial surface of the caudal portion of the cerebral hemisphere. It also supplies the diencephalon and the rostral mesencephalon.
The rostral cerebellar artery leaves the caudal third of the arterial circle caudal to the oculomotor nerve and courses dorsocaudally along the pons and the middle cerebellar peduncle to the cerebellar hemisphere. It supplies the caudal midbrain and the rostral half of the cerebellum.
The caudal cerebellar artery is a branch of the basilar artery near the middle of the medulla. It courses dorsally to supply the caudal portion of the cerebellum.
VEINS
The venous sinuses (Figs. 6-4 through 6-6) of the cranial dura mater are large venous passageways located within the dura or within bony canals in the skull unaccompanied by arteries. These sinuses receive the veins draining the brain and the bones of the skull. They convey venous blood to the paired maxillary, internal jugular, and vertebral veins and to the ventral internal vertebral venous plexuses (Fig. 6-4). The following venous sinuses should be located.
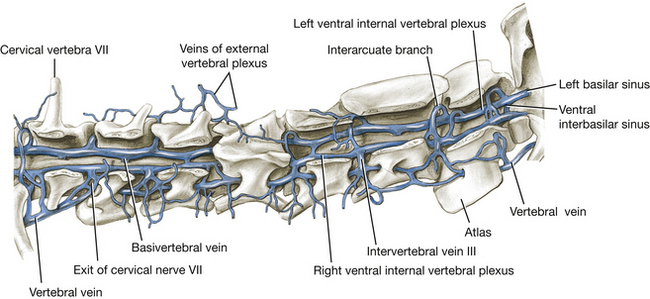
Fig. 6-4 Cervical vertebral veins, right lateral aspect. (From Reinhard K, Miller M, Evans H: The craniovertebral veins and sinuses of the dog, Am J Anat 11:67-87, 1962. Copyright © 1962 Wiley-Liss. Reprinted by permission of Wiley-Liss, Inc., a subsidiary of John Wiley & Sons, Inc.)
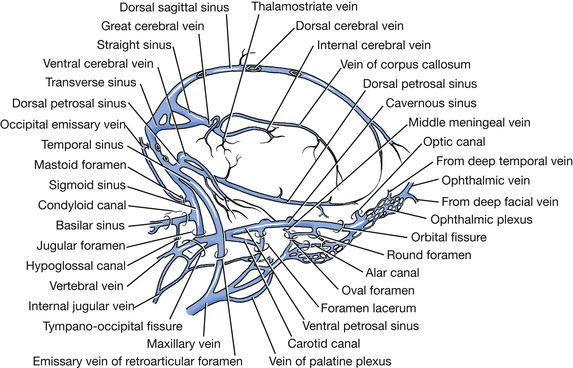
Fig. 6-5 Cranial venous sinuses, right lateral aspect. (From Reinhard K, Miller M, Evans H: The craniovertebral veins and sinuses of the dog, Am J Anat 11:67-87, 1962. Copyright © 1962 Wiley-Liss. Reprinted by permission of Wiley-Liss, Inc., a subsidiary of John Wiley & Sons, Inc.)
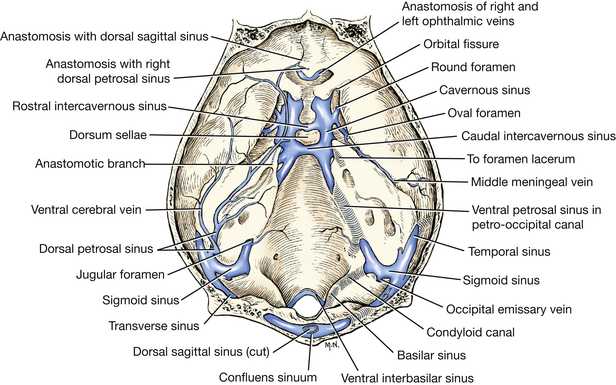
Fig. 6-6 Cranial venous sinuses, dorsal aspect, calvaria removed. (From Reinhard K, Miller M, Evans H: The craniovertebral veins and sinuses of the dog, Am J Anat 11:67-87, 1962.)
The dorsal sagittal sinus is located in the attached edge of the falx cerebri, which is a fold of dura extending ventrally into the longitudinal fissure between the two cerebral hemispheres. Caudally, this sinus enters the foramen for the dorsal sagittal sinus in the occipital bone dorsal to where the tentorium attaches. There it joins the right and left transverse sinuses.
Each transverse sinus runs laterally through the transverse canal and sulcus. At the distal end of the sulcus, at the dorsal border of the petrosal part of the temporal bone, the sinus divides into a temporal and a sigmoid sinus. Coursing caudolateral to the petrosal part of the temporal bone, the temporal sinus extends to the retroarticular foramen, where it emerges as the emissary vein of the retroarticular foramen and joins the maxillary vein.
Each sigmoid sinus forms an S-shaped curve as it courses over the dorsomedial side of the petrous part of the temporal bone. It passes through the jugular foramen into the tympano-occipital fissure. Within the fissure the ventral petrosal sinus enters rostrally from the petro-occipital canal and joins the sigmoid sinus. From this anastomosis the vertebral and internal jugular veins arise and leave the tympano-occipital fissure and course caudally. The vertebral vein descends the neck through the transverse foramina of the cervical vertebrae. The internal jugular vein was seen previously in the carotid sheath. The basilar sinus is a branch of the sigmoid sinus that continues caudally through the condyloid canal to the ventral internal vertebral venous plexus in the vertebral canal.
The cavernous sinus (Fig. 6-6) lies on each side of the floor of the middle cranial fossa from the orbital fissure to the petro-occipital canal. Emissary veins connect each cavernous sinus with the ophthalmic plexus of veins rostrally and with the maxillary vein laterally. These sinuses are each continued caudally by the ventral petrosal sinus, which lies in the petro-occipital canal. Two or three intercavernous sinuses connect the left and right cavernous sinuses rostral aud caudal to the pituitary gland. The internal carotid artery is seen coursing through the cavernous sinus.
The ventral internal vertebral venous plexuses are paired vessels that lie on the floor of the vertebral canal in the epidural connective tissue. They extend from the venous sinuses of the cranium throughout the vertebral canal. At each intervertebral foramen, intervertebral veins connect the vertebral venous plexus with the vertebral veins of the neck, intercostal veins of the thorax (azygos and costocervical veins), and lumbar veins (caudal vena cava) in the abdomen. This plexus will be seen later when the spinal cord is removed from the vertebral canal.
There is a continuous venous pathway from the angularis oculi vein (Fig. 5-40), through the external ophthalmic vein, ophthalmic plexus, cavernous sinus, ventral petrosal sinus, sigmoid sinus, and the basilar sinus in the condyloid canal, to the ventral internal vertebral venous plexus. This pathway can be demonstrated radiographically by compressing the external jugular veins and injecting a radiopaque solution into the angularis oculi vein on the side of the face near the medial angle of the eye. This procedure can be used clinically to diagnose space-occupying lesions that interfere with this pathway. It can also be used anatomically to inject the venous system with various materials.
BRAIN
The brain is composed of the embryologically segmented brain stem and two suprasegmental portions, the cerebrum (telencephalon) and the cerebellum (dorsal metencephalon). The brain stem includes the myelencephalon (medulla), the ventral metencephalon (pons), the mesencephalon (midbrain), and the diencephalon (interbrain epithalamus, thalamus, and hypothalamus).
Dissect and identify the following structures on the intact brain that has been provided.
Cerebrum—Surface Structures
The cerebrum is divided into two cerebral hemispheres by the longitudinal fissure. Each cerebral hemisphere has outward folds (convolutions) called gyri and inward folds called sulci. Identify the following gyri and sulci (Figs. 6-7, 6-8): the rostral and caudal parts of the lateral rhinal sulcus, the pseudosylvian fissure, the rostral and caudal sylvian gyri, the ectosylvian sulcus and gyrus, the suprasylvian sulcus and gyrus, the cruciate sulcus, the postcruciate and precruciate gyri, the coronal sulcus, the ectomarginal sulcus and gyrus, and the marginal sulcus and gyrus.
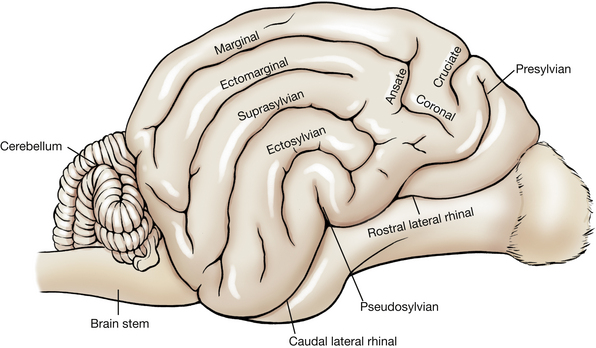
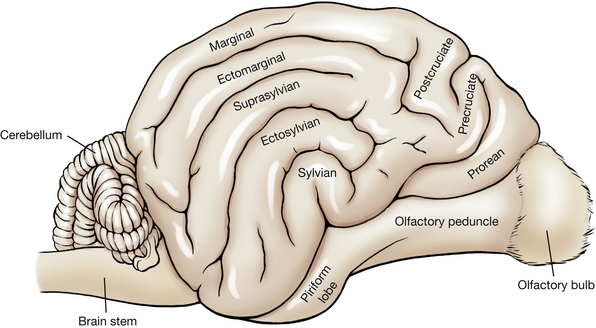
Each cerebral hemisphere may be divided into lobes named for that portion of the calvaria that covers them. The relationship is not precise and varies among species.
The frontal lobe is that portion of each cerebral hemisphere rostral to the cruciate sulcus. The precruciate gyrus is part of this lobe and functions as part of the motor cortex. The parietal lobe is caudal to the cruciate sulcus and dorsal to the sylvian gyri. It extends caudally to approximately the caudal third of the cerebral hemisphere. The postcruciate and rostral suprasylvian gyri are found in this lobe and function as part of the motor and somesthetic sensory cerebral cortex. The occipital lobe includes the caudal third of the cerebral hemisphere. Caudal portions of this lobe on both medial and lateral sides function as the visual cortex. The temporal lobe is composed of the gyri and sulci on the ventrolateral aspect of the cerebral hemisphere. Parts of the sylvian gyri are located here and function as the auditory cortex.
The rhinal sulcus separates the phylogenetically new cerebrum or neopallium, above, from the older olfactory cerebrum, the paleopallium, below. Portions of the paleopallium that are visible are the olfactory bulb, which rests on the cribriform plate, and the olfactory peduncle, which joins the bulb to the cerebral hemisphere. The olfactory peduncle (Figs. 6-8, 6-9) courses caudally with a band of fibers on its ventral surface. Caudally, this band divides into lateral and medial olfactory tracts. Observe the lateral olfactory tract passing caudally to the piriform lobe, which forms a ventral bulge just lateral to the pituitary gland and medial to the temporal lobe of the neopallium. The medial olfactory tract cannot be observed.
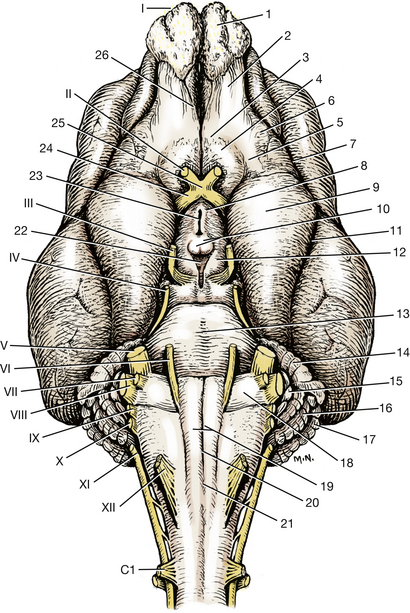
Fig. 6-9 Ventral view of brain, cranial nerves, and brain stem.
Each gyrus contains gray matter superficially and white matter in its center. The gray matter, or cerebral cortex of the neopallium, is composed of six layers of neuronal cell bodies. The white matter, corona radiata, contains the processes of neurons coursing to and from the overlying cortex.
Cerebellum
The cerebellum is derived from the dorsal portion of the metencephalon and lies caudal to the cerebrum and dorsal to the fourth ventricle. The transverse cerebral fissure separates it from the cerebrum. The dural and osseous tentorium cerebelli is located in this fissure. The cerebellum is connected to the brain stem by three cerebellar peduncles on each side of the fourth ventricle and by portions of the roof of the fourth ventricle.
The choroid plexus (Fig. 6-1) is a compact mass of pia, blood vessels, and ependyma. A choroid plexus develops where neural tube neuroepithelium did not proliferate to form parenchyma but remained as a single layer of the neuroepithelial cells, a roof plate. These areas are found in the medulla (roof plate of the fourth ventricle), the diencephalon (roof plate of the third ventricle), and telencephalon (roof plate of the lateral ventricle). At these sites the vessels in the pia covering the single layer of neuroepithelial cells proliferate to form a dense plexus of capillaries intimately related to the neuroepithelial cells. These cells and blood vessels are involved in passive and active secretion of cerebrospinal fluid into the ventricular system. The choroid plexus of the fourth ventricle protrudes into the lumen of the fourth ventricle and is visible caudolateral to the cerebellum on the dorsal surface of the medulla.
Identify the transverse fibers of the pons on the ventral surface of the brain stem. Follow these fibers laterally as they course dorsocaudally into the cerebellum on each side as the middle cerebellar peduncle (Figs. 6-9, 6-10, 6-17). At the point where they merge into the cerebellum, cut this peduncle with a scalpel. Continue the cut slightly rostral to cut the rostral cerebellar peduncle. Gently lift the caudal part of the cerebellum from the medulla. Remove the choroid plexus so you can see the caudal cerebellar peduncle connecting the medulla and cerebellum. Cut this peduncle and detach the cerebellum from the pons on that side. Cut these peduncles on the opposite side and remove the cerebellum. The rostral cerebellar peduncle contains mainly efferent axons from the cerebellum to the brain stem. Afferent axons to the cerebellum from the brain stem and spinal cord pass primarily through the middle and caudal cerebellar peduncles.
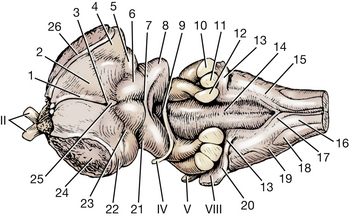
Fig. 6-10 Dorsal view of brain stem.
7. Commissure of caudal colliculus
9. Crossing of trochlear nerve fibers in rostral medullary velum
10. Middle cerebellar peduncle
11. Caudal cerebellar peduncle
12. Rostral cerebellar peduncle
13. Dorsal cochlear nucleus in acoustic stria
14. Median sulcus in fourth ventricle
18. Spinal tract of trigeminal nerve
19. Superficial arcuate fibers
20. Left ventral cochlear nucleus
21. Brachium of caudal colliculus
23. Brachium of rostral colliculus
24. Cut internal capsule between cerebral hemisphere and brain stem
II
IV
V
VIII
The cerebellum is composed of lateral cerebellar hemispheres and a middle portion, the vermis. The convolutions of the cerebellum are known as folia. These are grouped into three lobes and numerous cerebellar lobules that have specific names. The vermis comprises the entire middle portion of the cerebellum directly above the fourth ventricle. Some of its lobules are found on the ventral surface of the cerebellum facing the roof plate of the fourth ventricle. Each hemisphere projects over the cerebellar peduncles and the adjacent brain stem. A lateral component lies in the cerebellar fossa of the petrosal part of the temporal bone.
Make a median incision through the vermis, hemisectioning the cerebellum. Examine the cut surface. Note the pattern of white matter as it branches and arborizes from the medulla of the cerebellum into the folia (Figs. 6-15, 6-17, 6-19). The medulla of the cerebellum is the white matter in its central portion that contains the cerebellar nuclei and connects with all the folia and the cerebellar peduncles. Observe the laminae of foliate white matter and the cerebellar cortex. Make a transverse section of one half of the cerebellum through its medulla to observe the lateral extent of the medullary white matter and its nuclei (Fig. 6-15).
Brain Stem—Surface Structures
To expose the dorsal surface structures of the brain stem, the left cerebral hemisphere will be removed by the following dissection.
Gently separate the two cerebral hemispheres at the longitudinal fissure. Expose the band of fibers that course transversely from one hemisphere to the other in the depth of the fissure. This structure is the corpus callosum. Completely divide the corpus callosum longitudinally along the median plane in the depth of the longitudinal fissure. Cut deep enough to include the hippocampal commissure and body of the fornix, but do not cut into the thalamus (Figs. 6-11, 6-12, 6-17). Continue the cut rostrally and ventrally through the rostral commissure just dorsal to the optic chiasm and rostral to the thalamus. On the ventral surface, follow the optic tract in a dorsocaudal direction from the optic chiasm and cut the fibers of the internal capsule rostral and medial to this tract. This is an oblique cut. The fibers of the internal capsule attach the cerebral hemisphere to the brain stem. Gently lift the medial side of the cerebrum off the thalamus and continue this separation over the dorsal aspect of the diencephalon. Cut any remaining attachments, and remove the cerebral hemisphere from the diencephalon.
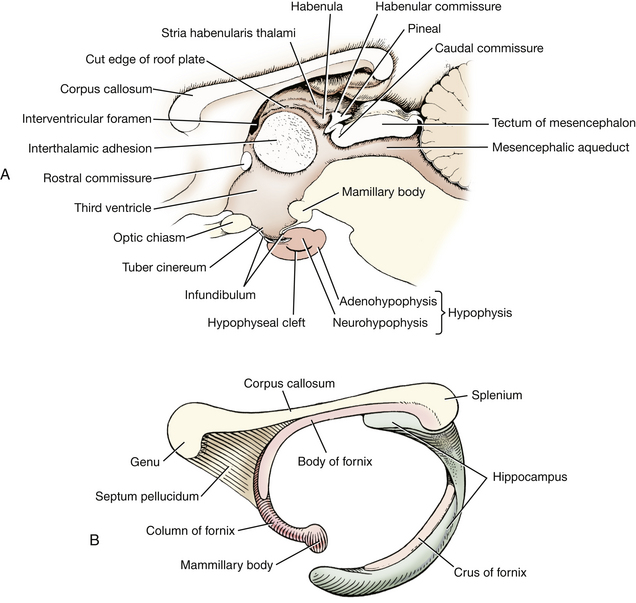
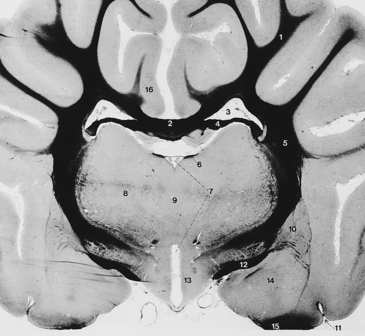
Fig. 6-12 Diencephalon and cerebral hemispheres.
(In this and the following transverse sections, the white matter is stained with iron hematoxylin and appears black in the photographs.)
Examine Fig. 6-12 and recognize the structures you have cut to remove the cerebral hemisphere. The internal capsule is the only structure connecting the cerebral hemisphere with the brain stem. It consists of neuronal processes projecting from the cerebral hemisphere to the brain stem and neuronal processes projecting from the thalamus to the cerebral hemisphere. This will be dissected further later.
Examine the surface of the brain stem and locate the following structures (Figs. 6-9, 6-10).
Diencephalon
The diencephalon consists of a large, centrally located thalamus; a smaller hypothalamus below; and a very small epithalamus on the dorsal midline. It is the most rostral part of the brain stem.
The optic or second cranial nerves form the optic chiasm of the diencephalon rostral to the hypophysis or pituitary gland (Figs. 6-9, 6-10). The optic tracts course laterally and dorsocaudally from the chiasm, pass over the lateral surface of the diencephalon, and enter the lateral geniculate nucleus of the thalamus. In this pathway, each tract curves around the caudal edge of the internal capsule (Figs. 6-12, 6-17, 6-19).
Caudal to the optic chiasm on the median plane is the hypophysis (pituitary gland), which is attached by the infundibulum to the tuber cinereum of the hypothalamus. If the gland is missing, the lumen of the infundibulum will be evident. This lumen communicates with the overlying third ventricle of the diencephalon.
The mamillary bodies of the hypothalamus bulge ventrally caudal to the tuber cinereum. They demarcate the most caudal extent of the hypothalamus on the ventral surface of the diencephalon.
The internal capsule bounds the diencephalon laterally and was cut when the left cerebral hemisphere was removed. The thalamus and epithalamus can be seen on the dorsal aspect of the diencephalon (Figs. 6-10, 6-11).
Three structures compose the epithalamus. They all are located adjacent to the median plane. The stria habenularis lies on either side of the midline, coursing dorsally and caudally from the rostroventral aspect of the hypothalamus over the thalamus to the dorsocaudal aspect of the diencephalon. Here the stria enters the habenular nucleus. The habenula nuclei on each side are connected by a small commissure. Caudal to the habenular nucleus is the small, unpaired pineal body. This caudal projection from the diencephalon is small in the dog but very prominent in larger domestic animals.
A space is located between the stria habenularis of each side. This is the dorsal part of the third ventricle (Figs. 6-10 through 6-12). It is covered by a thin remnant of the roof plate of the neural tube, a layer of ependyma that extends from one stria habenularis to the other. Branches of the caudal cerebral artery course over the diencephalon and form the choroid plexus of the third ventricle. This is usually pulled out when the calvaria is removed to expose the brain. Rostrally, the choroid plexus of the third ventricle is continuous with the choroid plexus of the lateral ventricle at the interventricular foramen. This foramen is caudal to the column of the fornix at the level of the rostral commissure. These structures will be seen in the dissection of the telencephalon.
The thalamus lies between the internal capsule on each side and dorsal to the hypothalamus. It is covered by pia, arachnoid trabeculations, and the subarachnoid space. It consists of a multitude of nuclei, two of which are readily recognized on the caudal surface. A lateral eminence on the caudodorsal surface of the thalamus is the lateral geniculate nucleus, which receives fibers of the optic tract and functions in the visual system. The lateral geniculate nucleus is connected with the rostral colliculus of the midbrain. Caudoventral to the lateral geniculate nucleus is the medial geniculate nucleus of the thalamus. This nucleus functions in the auditory system and is connected to the caudal colliculus of the midbrain by the brachium of the caudal colliculus.
In the third ventricle, between the stria habenularis of each side, observe the interthalamic adhesion between the right and left sides of the thalamus. This area appears round on median section because the third ventricle encircles it (Fig. 6-11). In transverse section the narrow, vertically oriented third ventricle appears as a perpendicular slit below the interthalamic adhesion (Fig. 6-12). Its lateral and ventral walls are formed by the hypothalamus. The dorsal portion of the third ventricle is small and tubular. It passes over the interthalamic adhesion, but its thin roof plate, which is attached on each side to the stria habenularis, cannot be observed grossly.
Mesencephalon
The mesencephalon is relatively short. It is nearly round on transverse section with a canal, the mesencephalic aqueduct (Figs. 6-13, 6-14), passing through it. The mesencephalon consists of a tectum or roof dorsal to the aqueduct, which is composed of four groups of neuronal cell bodies: the colliculi. That portion of the mesencephalon ventral to the aqueduct is the cerebral peduncle, which consists of a tegmentum (reticular formation), substantia nigra, and crus cerebri, from dorsal to ventral.
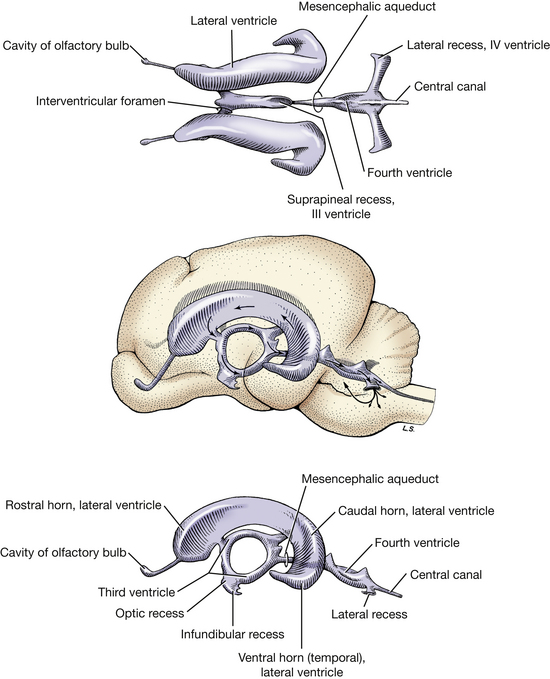
Fig. 6-13 Ventricles of brain. Direction of arrows indicates flow of cerebrospinal fluid. (From deLahunta A: Veterinary neuroanatomy and clinical neurology, Philadelphia, 1983, Saunders.)
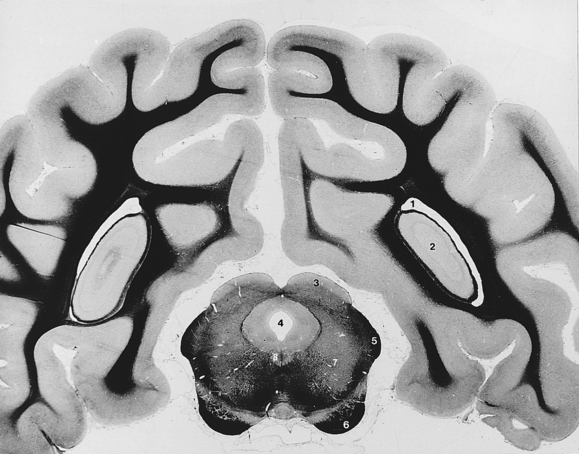
Fig. 6-14 Mesencephalon and cerebral hemispheres.
(The white matter is stained with iron hematoxylin and appears black in the photograph.)
Between the mamillary bodies of the hypothalamus and the transverse fibers of the pons is the ventral surface of the mesencephalon (midbrain). The transverse fibers of the pons cover part of the caudal mesencephalon ventrally. Descending tracts of projection processes that connect portions of the cerebral cortex with brain stem centers and the spinal cord course on the ventral surface of the midbrain. These are grouped together on each side as the crus cerebri (Figs. 6-9, 6-14). The oculomotor, or third cranial, nerve leaves the midbrain medial to the crus (Fig. 6-9).
The mesencephalic structures dorsal to the mesencephalic aqueduct compose the tectum of the midbrain (Fig. 6-11). The mesencephalic aqueduct is a short, narrow tube, derived from the neural canal in the midbrain, that connects the third ventricle rostrally with the fourth ventricle caudally. Four dorsal bulges, the corpora quadrigemina, are evident on the dorsal side. The rostral pair are the rostral colliculi, which function with the visual system. The smaller caudal pair are the caudal colliculi, which function in the auditory system (Fig. 6-10).
The trochlear, or fourth cranial, nerve courses laterally out of the roof of the fourth ventricle, the rostral medullary velum, adjacent to the caudal colliculus (Fig. 6-10). It continues rostroventrally on the lateral surface of the midbrain.
The lateral lemniscus (Figs. 6-17, 6-19) is a band of auditory system axons on the lateral side of the midbrain. It courses rostrodorsally from the level of the cochlear nucleus to the caudal colliculus and emerges medial to the middle cerebellar peduncle. Many of these fibers arise from the cochlear nucleus. The brachium of the caudal colliculus (Figs. 6-10, 6-17, 6-19) runs rostroventrally from the caudal colliculus to the medial geniculate nucleus of the thalamus. On the dorsal surface the commissure of the caudal colliculi (Fig. 6-10) can be seen crossing between these two structures. The rostral colliculus is connected to the lateral geniculate nucleus of the thalamus by a short brachium of the rostral colliculus (Fig. 6-10).
Ventral Metencephalon
The metencephalic portion of the rhombencephalon includes a segment of the brain stem, the pons, and the dorsal development, the cerebellum. The ventral surface of the pons includes the transverse fibers of the pons (Fig. 6-9), which course laterally into the middle cerebellar peduncles. This large band of transverse fibers borders the trapezoid body of the medulla caudally. Its rostral border covers part of the ventral surface of the midbrain. The trigeminal nerve is associated with the pons and can be found entering the pons along the caudolateral aspect of the transverse fibers (Figs. 6-9, 6-17). The descending fibers of the crus cerebri enter the pons dorsal to the transverse fibers, where they are called the longitudinal fibers of the pons. These longitudinal fibers are covered ventrally by the transverse fibers. The longitudinal fibers that do not terminate in pontine nuclei continue caudally on the ventral surface of the trapezoid body of the medulla as the pyramids. Many of the axons in the crus cerebri and the longitudinal fibers of the pons, and most of those in the transverse fibers of the pons, make up a large cerebropontocerebellar pathway. Synapse occurs in the pontine nuclei that are covered by the transverse fibers, and crossing occurs through the transverse fibers to enter the cerebellum through the middle cerebellar peduncle. Therefore impulses that arise in the left cerebrum are projected to the right cerebellar hemisphere.
The rostral medullary velum forms the roof of the fourth ventricle between the caudal colliculi of the mesencephalon rostrally and the midventral surface of the cerebellum caudally. The crossing fibers of the trochlear nerves course through this velum (Fig. 6-10). The velum in the preserved specimen lies on the floor of the fourth ventricle and covers the caudal opening of the mesencephalic aqueduct. Insert a probe into the slitlike fourth ventricle under the caudal cut edge of this velum and raise the velum to demonstrate its attachments and the continuity of the fourth ventricle with the aqueduct. This velum lies between the fourth ventricle, which it covers, and the rostral cerebellar vermis. Between the velum and the vermis is subarachnoid space.
Myelencephalon (Medulla)
The myelencephalon, or medulla, extends from the transverse fibers of the pons to the level of the ventral rootlets of the first cervical spinal nerve. The trapezoid body is the transverse band of fibers rostrally that course parallel but caudal to the transverse pontine fibers (Figs. 6-9, 6-17, 6-19). It is continuous with the vestibulocochlear nerve and cochlear nuclei laterally on the side of the medulla and functions in the auditory system. The pyramids are a pair of longitudinally coursing fiber bundles on either side of the ventral median plane. They emerge from the transverse fibers as the caudal continuations of axons from the longitudinal fibers of the pons that did not terminate in pontine nuclei. They course caudally across the trapezoid body to continue on the ventral surface of the medulla. They are separated by the ventral median fissure. This fissure can be followed caudally until it is obliterated over a short distance by the decussation of the pyramids located at the level of the emerging hypoglossal nerve fibers. The decussation itself is difficult to see because it occurs as the pyramidal fibers are passing dorsally into the parenchyma of the medulla. Pyramidal axons continue in the spinal cord as the corticospinal tracts. These are cerebral projection processes that project to the spinal cord.
The abducent, or sixth cranial, nerve leaves the medulla through the trapezoid body on the lateral border of each pyramid.
Cranial nerve VII, the facial nerve (Fig. 6-9), is smaller and leaves the lateral surface of the medulla through the trapezoid body caudal to the trigeminal nerve and rostroventral to the eighth cranial nerve.
Cranial nerve VIII, the vestibulocochlear nerve (Figs. 6-9, 6-10, 6-15), is on the lateral side of the medulla at the most lateral extent of the trapezoid body just dorsal to the facial nerve. The cochlear nuclei are located in this nerve where it attaches to the medulla (Figs. 6-10, 6-15, 6-17). All cochlear neurons in the vestibulocochlear nerve synapse in cochlear nuclei. The axons of these cell bodies in the cochlear nuclei continue into the medulla by one of two pathways. Some continue as the acoustic stria (Fig. 6-10) and pass over the dorsolateral surface of the medulla and caudal cerebellar peduncle to enter the medulla. Others pass on the ventral surface in the trapezoid body (Figs. 6-9, 6-15). The vestibular neurons in the vestibulocochlear nerve pass directly into the medulla, where most of them synapse in a vestibular nucleus (Fig. 6-15).
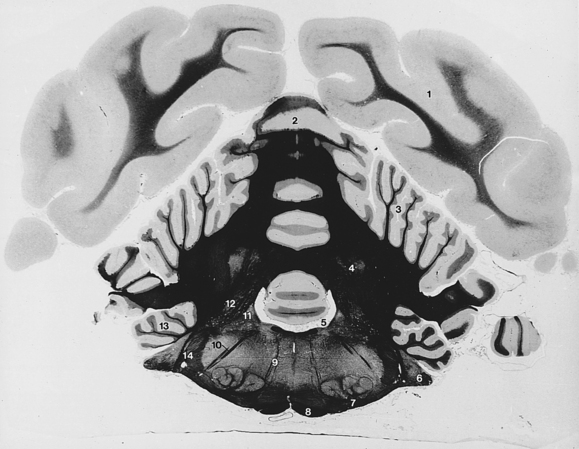
Fig. 6-15 Cerebellum and myelencephalon.
6. Cochlear nuclei and vestibulocochlear nerve
10. Descending facial nerve fibers
(The white matter is stained with iron hematoxylin and appears black in the photographs.)
The hypoglossal nerve, cranial nerve XII (Fig. 6-9), emerges as a number of fine rootlets from the ventrolateral surface of the myelencephalon caudal to the trapezoid body, lateral to the caudal portion of the pyramid. The junction of the myelencephalon and the spinal cord is between the hypoglossal fibers and the ventral rootlets of the first cervical spinal nerve. The hypoglossal nerve outside the skull is much larger, owing to the addition of connective tissue components.
Dorsolateral to the emergence of the hypoglossal fibers and the fibers of the ventral root of the first cervical spinal nerve, a nerve runs lengthwise along the lateral surface of the spinal cord. This is the eleventh cranial nerve, the accessory nerve (Figs. 6-9, 6-20). Its spinal rootlets emerge from the lateral surface of the spinal cord as far caudally as the seventh cervical segment. They emerge between the level of the dorsal and ventral rootlets of the cervical spinal nerves and course cranially within the subarachnoid space through the vertebral canal and foramen magnum. A few cranial rootlets emerge from the lateral side of the medulla caudal to the tenth cranial nerve and join the accessory nerve as it courses by the medulla (Fig. 6-9). These are usually torn off the medulla in removing the brain for dissection. The accessory nerve leaves the cranial cavity through the jugular foramen and tympano-occipital fissure along with the ninth and tenth cranial nerves.
Cranial nerves IX and X, the glossopharyngeal and vagal nerves, leave the lateral side of the myelencephalon caudal to the eighth cranial nerve and rostral to the accessory nerve. These rootlets are small and are rarely preserved on the brain when it is removed.
Examine the dorsal surface of the pons and medulla. On either side of the fourth ventricle are the cut ends of the three cerebellar peduncles. The rostral cerebellar peduncle is medial and courses rostrally into the mesencephalon; the middle cerebellar peduncle is lateral and arises from the transverse fibers on the lateral side of the pons; and the caudal cerebellar peduncle is in the middle, entering from the myelencephalon after passing beneath the acoustic stria.
The groove in the center of the floor of the fourth ventricle is the median sulcus. On the lateral wall the longitudinal groove is the sulcus limitans. Just lateral to this latter groove, at the level of the acoustic stria, there is a slight dorsal bulge of the medulla. This demarcates the location of the vestibular nuclei. Most of the vestibular neurons in the vestibulocochlear nerve terminate here.
The roof of the fourth ventricle caudal to the cerebellum is the caudal medullary velum. It is a thin layer composed of ependyma lining the ventricle and a supporting layer of vascularized pia. It attaches to the cerebellum rostrally. This attachment is in the midventral portion of the cerebellum between the rostral and caudal ventral portions of the vermis. This velum attaches to the caudal cerebellar peduncle and the fasciculus gracilis laterally and caudally. Its attachment caudally at the apex is known as the obex. At this level the fourth ventricle is continuous with the central canal of the spinal cord.
At the level of the eighth cranial nerve, dorsal to the acoustic stria, there is an opening in the caudal medullary velum known as the lateral aperture of the fourth ventricle (Fig. 6-1). The cerebrospinal fluid produced in the ventricular system communicates with the subarachnoid space of the meninges via this aperture. It then courses through the subarachnoid space over the entire surface of the brain and spinal cord and is absorbed into the venous system. Most of this absorption occurs where the arachnoid is in close apposition to the cerebral venous sinuses and where it has formed specialized structures known as arachnoid villi. Cerebrospinal fluid is also absorbed from the subarachnoid space, where the spinal nerves leave the vertebral canal through the intervertebral foramina, and along the olfactory and optic nerves.
The choroid plexus of the fourth ventricle bulges into the lumen of the ventricle on each side of the dorsal midline. Each plexus extends outward through the lateral recess and aperture, where it was seen caudal to the cerebellum before the latter was removed.
Examine the dorsal surface of the myelencephalon caudal to the fourth ventricle. The structures to be observed can be more easily recognized if the pia arachnoid is removed and the medulla is examined under a dissecting microscope. The median groove is the dorsal median sulcus. The narrow longitudinal bulge flanking the sulcus is the fasciculus gracilis (Fig. 6-10). This longitudinal tract ascends the entire length of the spinal cord in this position. At the caudal end of the myelencephalon, it ends at the nucleus gracilis. This nucleus is located at the caudal end of the fourth ventricle, where the fasciculus widens and ends. This fasciculus and nucleus function primarily in pelvic limb proprioception.
The groove lateral to the fasciculus gracilis is the dorsal intermediate sulcus. The longitudinal bulge lateral to this is the fasciculus cuneatus. This tract also ascends the dorsal aspect of the spinal cord, starting in the midthoracic region. The fasciculus cuneatus diverges laterally at the caudal end of the fourth ventricle and ends in a slight bulge. This bulge represents the lateral cuneate nucleus and is known as the cuneate tubercle. Rostrally, the lateral cuneate nucleus is continuous with the caudal cerebellar peduncle. This fasciculus and nucleus primarily function in thoracic limb proprioception.
The groove on the caudodorsal surface of the myelencephalon lateral to the fasciculus cuneatus is the dorsolateral sulcus.
The longitudinal bulge lateral to it is the spinal tract of the trigeminal nerve (Figs. 6-10, 6-17). The axons in this tract are from sensory cell bodies in the trigeminal ganglion that supply the head. The trigeminal tract extends caudally to the level of the first cervical segment of the spinal cord because of the large number of cell bodies serving its extensive peripheral distribution in the head. This tract emerges on the lateral surface of the myelencephalon caudal to a band of obliquely ascending fibers, the superficial arcuate fibers. The arcuate fibers connect structures in the medulla with the caudal cerebellar peduncle.
The dorsal rootlets of the spinal cord enter through the dorsolateral sulcus along the spinal cord.
Telencephalon (Cerebrum)
The left cerebral hemisphere was previously removed (Fig. 6-17) by cutting the rostral commissure, corpus callosum, and hippocampal commissure; separating the two halves of the body of the fornix on the median plane; and sectioning the internal capsule, which attached the cerebral hemisphere to the thalamus of the brain stem.
There are three commissural pathways crossing between the hemispheres, one for each phylogenetic division of the cerebrum. The corpus callosum connects the neopallial portion of each hemisphere and is the largest of the three pathways. The rostral commissure connects the paleopallial or olfactory components of each hemisphere. The hippocampal commissure is small and is located just caudal to the junction of the crus of each fornix. This connects the archipallial components of each hemisphere. The internal capsule consists of projection fibers that course between the brain stem and cerebral hemisphere. Association fibers remain within the hemisphere, coursing between adjacent or distant gyri.
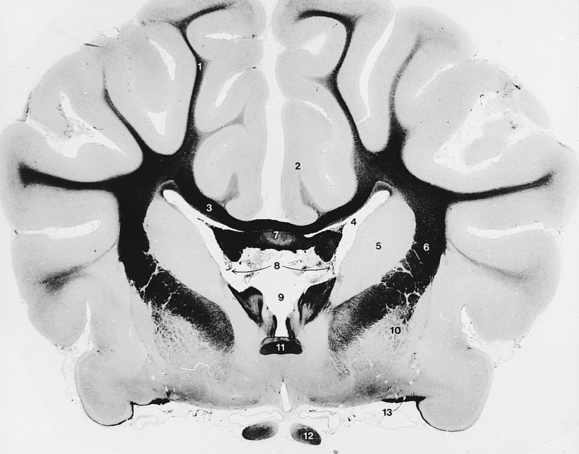
The corpus callosum (Figs. 6-11, 6-12, 6-16 through 6-18) consists of a rostral genu, middle body, and caudal splenium. It is the commissural pathway for axons crossing between the neopallium of each cerebral hemisphere. The internal capsule (Fig. 6-19) contains projection fibers that course between the telencephalon and the diencephalon and descend from the telencephalon to the brain stem and the spinal cord. The fibers that were previously seen in the mesencephalon as the crus cerebri descend through the internal capsule. The projection pathway of most sensory modalities to the cerebrum for conscious perception involves synapses in the thalamus. The axons of these thalamic cell bodies then enter the internal capsule to reach the cerebrum through thalamocortical fibers.
(The white matter is stained with iron hematoxylin and appears black in the photograph.)
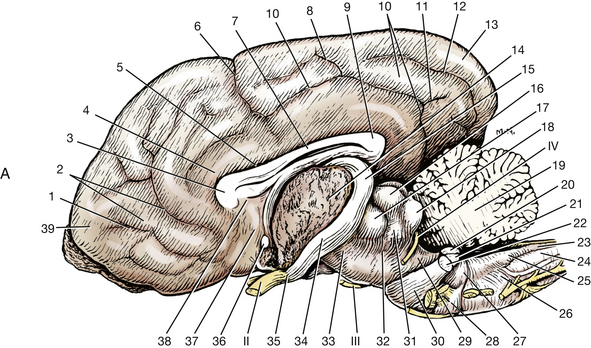
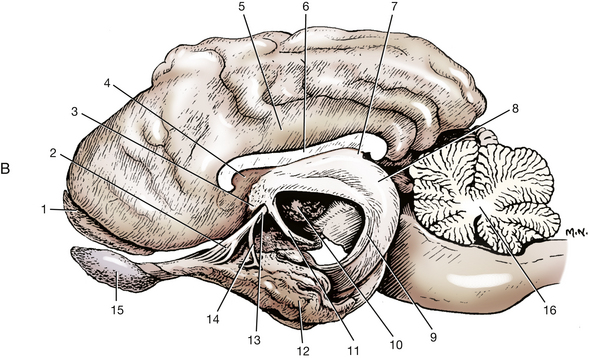
Fig. 6-17 A, Medial surface of right cerebral hemisphere and lateral surface of brain stem.
9. Splenium of corpus callosum
11. Caudal horizontal ramus of splenial sulcus
14. Cut internal capsule between cerebral hemisphere and brain stem
15. Optic tract at lateral geniculate nucleus
20. Rostral cerebellar peduncle
21. Caudal cerebellar peduncle
22. Middle cerebellar peduncle
24. Spinal tract of trigeminal nerve
26. Superficial arcuate fibers
31. Brachium of caudal colliculus
II
III
IV
(Adapted from Evans HE: Miller’s anatomy of the dog, ed 3, Philadelphia, 1993, Saunders.) B, Lateral view of a brain with the left half removed except for most of the left rhinencephalon.
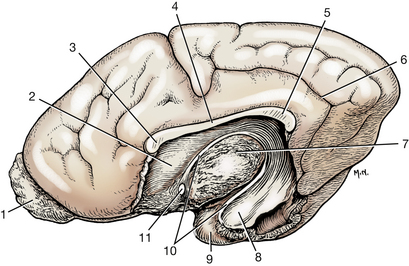
Fig. 6-18 Medial view of a right cerebral hemisphere with medial structures removed to show lateral ventricle. The rostral horn of the ventricle is bounded laterally by the caudate nucleus. (2). The distal part of the temporal horn is bounded laterally by the amygdala (8); elsewhere, the ventricle is bounded by white matter.
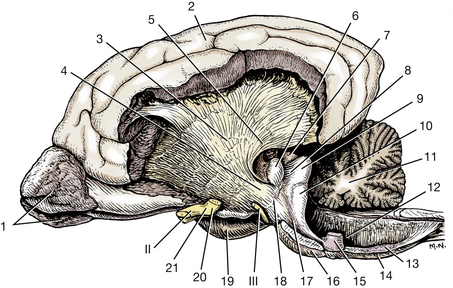
Fig. 6-19 Lateral view of the brain, internal capsule exposed.
Examine the isolated left cerebral hemisphere. On its medial surface, locate the portions of the corpus callosum and note the thin vertical sheet of tissue ventral to the corpus callosum. This is the septum pellucidum, which is more developed rostrally where it extends from the genu to the rostral commissure. A thickening in the septum dorsal and rostral to the rostral commissure represents the septal nuclei. Caudal to the rostral commissure, the septum attaches the corpus callosum to a column of fibers that courses rostrally and then descends in a rostroventral curve behind the rostral commissure. These fibers are part of the fornix. They connect the hippocampus with the diencephalon and rostral cerebrum. The hippocampus is a specialized region of cerebral cortex that will be seen shortly. Cut the septum pellucidum to separate the corpus callosum from the fornix. The fornix begins caudally by the accumulation of fibers on the lateral side of the hippocampus. These form the crus of the fornix. The crura join rostral to the hippocampus and dorsal to the thalamus to form the body of the fornix.
The body of the fornix courses rostrally and then descends rostroventrally as the column of the fornix. At the rostral commissure, some fibers course dorsal and rostral to the rostral commissure, but most descend caudal to the commissure and continue caudoventrally, lateral to the third ventricle, to reach the mamillary body of the hypothalamus. This descending column may be more evident on the intact right cerebral hemisphere. In the left cerebral hemisphere, pierce the septum and separate the corpus callosum from the column and body of the fornix.
The curved cavity exposed lateral to the septum pellucidum and ventral to the corpus callosum is the lateral ventricle. It communicates with the third ventricle of the diencephalon by the interventricular foramen (Figs. 6-11, 6-13, 6-16), which is located caudal to the column of the fornix at the level of the rostral commissure. The caudodorsal wall of this foramen is the roof plate and choroid plexus of the third ventricle. This is usually pulled free and lost when the brain is removed.
Locate the caudal part of the lateral rhinal sulcus in the left temporal lobe. Cut through this sulcus into the lateral ventricle. The smooth, curved bulge exposed in the wall of the ventricle is the caudal surface of the hippocampus as it ascends in a dorsorostral curve.
To remove the hippocampus intact from the left lateral ventricle, cut the column of the fornix dorsal to the rostral commissure on the medial side. Grasp the fornix with the forceps and, with the blunt end of the scalpel, gently roll the hippocampus out of the lateral ventricle. Its attachment ventrally in the temporal horn may be cut to free the hippocampus completely.
Phylogenetically, the cerebral cortex can be divided into three regions: paleopallium, neopallium, and archipallium. The paleopallium consists of the cortex of the olfactory peduncle. This is separated laterally from the neopallium by the rhinal sulcus. The neopallium includes all of the gyri on the external surface of the cerebrum. The hippocampus belongs to the archipallium and is an internal gyrus of the telencephalon that has been rolled into the lateral ventricle by the lateral expansion of the neopallium. Notice that the hippocampus begins ventrally in the temporal lobe and curves, first caudodorsally and then rostrodorsally, over the diencephalon to reach its caudodorsal aspect. At that point the hippocampus ends, and a hippocampal commissure connects the hippocampus of each cerebral hemisphere. The fibers of the hippocampus continue rostrally as the body and column of the fornix. Notice the crus of the fornix on its lateral surface. Place the previously removed hippocampus over the exposed left diencephalon to see its normal relationship with that structure. They are normally separated by arachnoid trabeculae and the subarachnoid space.
Attached to the lateral free edge of the crus of the fornix is a network of blood vessels covered by meninges and ependyma. This is the choroid plexus of the lateral ventricle. Its anatomical structure is the same as the choroid plexus of the third and fourth ventricles. The surface of the choroid plexus that faces the lumen of the ventricle is the ependymal layer derived from the embryonic neural tube. This layer of ependyma is attached on one side to the free edge of the fornix. To complete the wall of the lumen of the lateral ventricle, it must be attached on its other side. This attachment is to a small tract, the stria terminalis, located in the groove between the thalamus and the caudate nucleus (Figs. 6-10, 6-18). The stria terminalis is a small tract that connects the amygdala with the septal nuclei. This layer of ependyma from the stria terminalis to the fornix forms part of the medial wall of the lateral ventricle. Branches of the middle and caudal cerebral arteries covered by pia push this layer of ependyma into the lumen of the lateral ventricle. The result is the formation of a choroid plexus that is continuous with the choroid plexus of the third ventricle at the interventricular foramen (Fig. 6-1).
Examine the rostral commissure (Figs. 6-11, 6-16 through 6-18). On each side this commissure connects rostrally to the olfactory peduncles and caudally to the piriform lobes.
In each cerebral hemisphere the neuronal cell bodies are located either on the surface in the cerebral cortex or deep to the surface in a basal nucleus. Examine the floor of the lateral ventricle of the removed left hemisphere. The bulge that enlarges rostrally is the caudate nucleus. This is one of the subcortical basal nuclei of the telencephalon, a part of the corpus striatum. Its rostral extremity is the head. Caudal to this the body rapidly narrows into a small tail, which courses over the internal capsule fibers in the floor of the ventricle. With the blunt end of a scalpel, free the caudate nucleus from the medial side of the internal capsule.
Dorsolateral to the caudate nucleus, the internal capsule forms the lateral angle of the lateral ventricle. At the dorsolateral angle of the lateral ventricle, the fibers of the internal capsule meet those of the corpus callosum. Arising from this interdigitation of fibers, the corona radiata radiates in all directions to reach the gray matter of the cerebral cortex. A longitudinal section of the cerebral hemisphere in the dorsal plane at the level of this interdigitation reveals a mass of white matter in the center of the hemisphere. This mass is referred to as the centrum semiovale.
Remove the pia and arachnoid from the surface of the right cerebral hemisphere. Expose the corona radiata by removing the gray matter, the cerebral cortex, with the scalpel handle. Begin this removal of gray matter on the medial side of the hemisphere. The cingulate gyrus is located dorsal to the corpus callosum. Remove the gray matter of the cingulate gyrus to expose its fibers, which form the cingulum. Many of the fibers in the cingulum are long association fibers that course longitudinally from one end of the hemisphere to the other. Demonstrate this by freeing some fibers rostral to the genu of the corpus callosum and by stripping them caudally. Remove the cingulum and demonstrate the transverse course of the fibers of the corpus callosum by stripping these from their cut edge toward the hemisphere.
Remove the gray matter from the gyri on the lateral surface of the rostral half of the cerebral hemisphere. Examine the white matter of the gyri. Short association fibers, the arcuate fibers, course between adjacent gyri.
The internal capsule is situated lateral to the caudate nucleus. Expose the lateral surface of the internal capsule (Fig. 6-19).
When you have completed the dissection of the right cerebral hemisphere, all that remains are the corpus callosum, the internal capsule, and part of the corona radiata.
Remove the corpus callosum of the right cerebral hemisphere to expose the lateral ventricle, with the caudate nucleus on the floor rostrally and the hippocampus on the floor caudally. The internal capsule forms the lateral wall (Figs. 6-12, 6-14, 6-16, 6-19).
Make a median section of the brain stem from the optic chiasm through the medulla (Fig. 6-11). Observe the interthalamic adhesion and note the smooth surface of the third ventricle surrounding it. The ventricle is bounded dorsally by the roof plate between each stria habenularis. It connects rostrally with each lateral ventricle through the interventricular foramen, which is caudal to the column of the fornix and dorsal to the rostral commissure. Caudally, it is continuous with the mesencephalic aqueduct. Rostroventral to the interventricular foramen, the ventricle is bounded by the lamina terminalis, the most rostral extent of the embryonic neural tube on the median plane. (From this point each cerebral hemisphere develops laterally.) The third ventricle extends into the infundibulum of the pituitary gland as the infundibular recess (Fig. 6-11, 6-13).
Follow the ventricular system caudally from the third ventricle into the mesencephalic aqueduct and into the fourth ventricle. Note that the roof of the fourth ventricle consists of the rostral medullary velum, the cerebellum, and the caudal medullary velum. The choroid plexus of the fourth ventricle is formed in the caudal medullary velum. The fourth ventricle continues into the central canal of the spinal cord at the obex.
Functional and structural correlates of the nervous system are listed in Table 6-1.
Table 6-1
Functional and Structural Correlates of the Nervous System
| Neuroanatomical Structure | Function |
| Frontal lobe | Behavior |
| Frontoparietal lobe (sensorimotor cortex) | Upper motor neuron (central motor system) |
| Conscious perception of somatic, visceral, and proprioceptive sensations | |
| Temporal lobe | Behavior, hearing, balance |
| Occipital lobe | Vision |
| Olfactory bulb, peduncle, tract, piriform lobe | Olfaction—smell |
| Pons—transverse fibers | Pathway from cerebrum to cerebellum |
| Middle cerebellar peduncle | |
| Rostral cerebellar peduncle | Pathway from cerebellum to brain stem and cerebrum |
| Caudal cerebellar peduncle | Pathway from spinal cord and medulla with vestibular proprioceptive systems to cerebellum |
| Mamillary bodies | Limbic system (behavior) |
| Crus cerebri | Pathway from cerebrum to brain stem and spinal cord—upper motor neuron, and from cerebrum to cerebellum via pons |
| Internal capsule | Pathway between cerebrum and brain stem—upper motor neuron and thalamocortical projections—sensory and motor, and from cerebrum to cerebellum via pons |
| Hypothalamus | Visceral functions |
| Autonomic nervous system | |
| Limbic system | |
| Pituitary control | |
| Lateral geniculate nucleus | Visual system (conscious perception) |
| Rostral colliculus | Visual system (reflexes) |
| Medial geniculate nucleus | Auditory system (conscious perception) |
| Caudal colliculus | Auditory system (reflexes and conscious perception) |
| Brachium of caudal colliculus | |
| Lateral lemniscus | |
| Trapezoid body | |
| Acoustic stria | |
| Cochlear nuclei | |
| Pyramid | Upper motor neuron |
| Corticospinal tracts | |
| Fasciculus gracilis | Pelvic limb—conscious pathway of general proprioception |
| Nucleus gracilis | |
| Fasciculus cuneatus | Thoracic limb—general proprioception |
| Medial cuneate nucleus | Conscious perception pathway |
| Lateral cuneate nucleus | Cerebellar pathway |
| Spinal tract of trigeminal nerve | Projection pathway of somatic afferent sensation from head |
| Septal nuclei | Limbic system |
| Fornix—hippocampus | Limbic system |
| Cingulum | Limbic system |
| Caudate nucleus | Upper motor neuron |
| Lentiform nucleus | Upper motor neuron |
| Stria habenularis | Limbic system |
| Habenular nucleus | Limbic system |
SPINAL CORD
If a prepared dissection of the spinal cord is not available, do a complete laminectomy by removing all of the vertebral arches and the attached muscles from your specimen to expose the spinal cord. Observe the spinal cord with its dural covering. Between the dura of the spinal cord and the periosteum of the vertebrae is the epidural space, which contains loose connective tissue, fat, and blood vessels.
The spinal cord is divided into segments (Fig. 6-20). A group of dorsal and ventral rootlets leave each spinal cord segment on each side and combine, respectively, to form the spinal nerve at the level of the intervertebral foramen. Note the spinal ganglia in the intervertebral foramina (Figs. 6-21, 6-22).
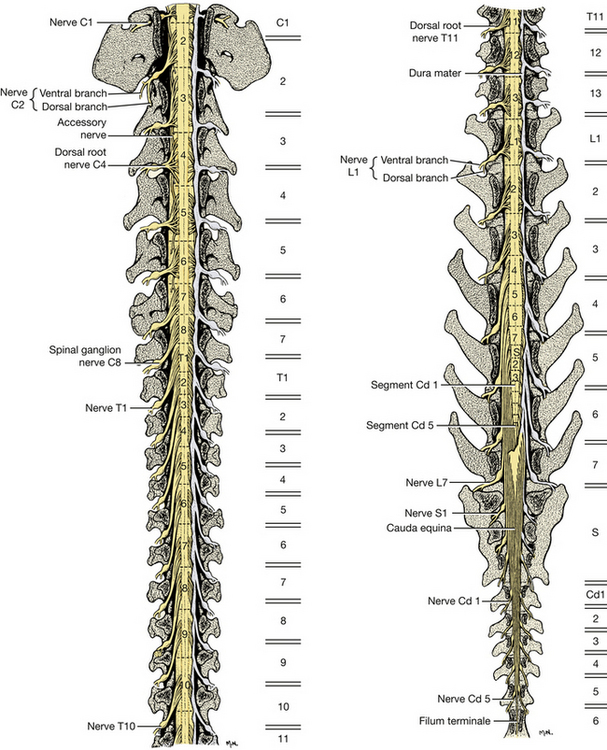
Fig. 6-20 Dorsal roots of spinal nerves and spinal cord segments. Dorsal view, vertebral arches removed. Dura removed on left side of both figures. (Figures on the right represent levels of vertebral bodies.)
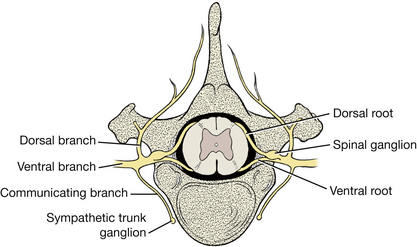
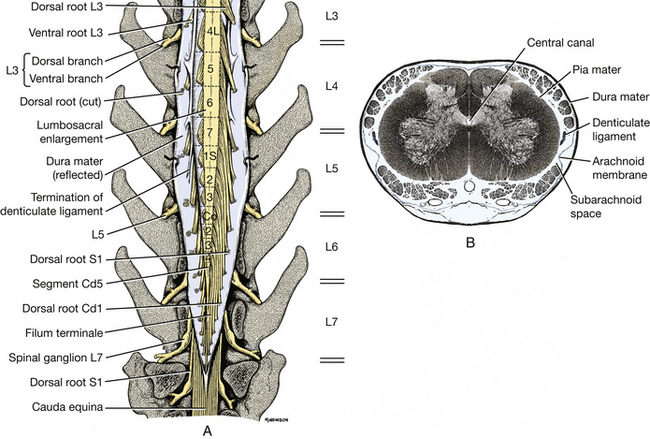
Fig. 6-22 A, Enlarged schematic view of terminal spinal cord with dura reflected. B, Enlarged transverse section of seventh lumbar segment. (Courtesy Dr. Thomas Fletcher.)
There are eight cervical spinal cord segments, thirteen thoracic, seven lumbar, three sacral, and about five caudal. The first cervical spinal nerves leave the vertebral canal through the lateral vertebral foramen in the arch of the atlas. The second cervical spinal nerves leave caudal to the atlas. The cervical spinal nerves of segments 3 through 7 leave the vertebral canal through the intervertebral foramina cranial to the vertebra of the same number. The spinal nerves of the eighth cervical segment pass caudal to the seventh (last) cervical vertebra. The spinal nerves of all the remaining spinal cord segments pass through the intervertebral foramina caudal to the vertebra of the same number.
In the caudal cervical region over the fifth to seventh cervical vertebrae, there is an enlargement of the spinal cord that nearly fills the vertebral canal. This is the cervical intumescence. Its presence is due to an increase in white matter and cell bodies that are associated with the innervation of the thoracic limb. The intumescence occurs from the sixth cervical segment of the spinal cord through the first thoracic segment. Another enlargement occurs in the midlumbar vertebral region for the innervation of the pelvic limb. The lumbar intumescence begins at about the fourth lumbar segment and gradually narrows caudally as the spinal cord comes to an end near the intervertebral space between the sixth and seventh lumbar vertebrae. The narrow caudal end of the parenchyma of the spinal cord is known as the conus medullaris. The spinal cord terminates in the filum terminale, which is a narrow cord of meninges that may include a long extension of the neural tube and central canal. This attaches the conus medullaris to the caudal vertebrae. The cauda equina includes the conus medullaris together with the adjacent caudal lumbar, sacral, and caudal roots that extend caudally in the vertebral canal.
Observe the relationship of the spinal cord segments to the corresponding vertebrae. The only spinal cord segments that are found entirely within their corresponding vertebrae are the last two thoracic and the first two (or occasionally three) lumbar segments. All other spinal cord segments reside in the vertebral canal cranial to the vertebra of the same number (Fig. 6-20). This is most pronounced in the caudal lumbar and sacrocaudal segments of the spinal cord. In general, the three sacral segments lie within the fifth lumbar vertebra and the caudal segments lie within the sixth lumbar vertebra. There is a breed variation in the length of the spinal cord. In small breeds it extends about one vertebra farther caudally, and in large breeds one vertebra farther cranially.
The nerve roots of the first 10 thoracic segments and those caudal to the third lumbar segment are long because of the distance between their origin at the spinal cord and their passage through the intervertebral foramen (Fig. 6-20).
Meninges
The thick, fibrous dura may be cut longitudinally along the entire length of the dorsal aspect of the spinal cord to reveal the dorsal rootlets and their length at different levels.
The thin arachnoid and pia are apposed to each other in the embalmed specimen and remain on the spinal cord. On the lateral surface of the spinal cord, the pia thickens and forms a longitudinal cord of connective tissue called the denticulate ligament. This ligament segmentally attaches to the arachnoid and dura laterally, midway between the roots of adjacent spinal cord segments (Figs. 6-20, 6-22).
Vessels
There is a longitudinal ventral spinal artery and one or two dorsal spinal arteries on the spinal cord. They are formed by spinal branches of the paired vertebral arteries in the cervical region, intercostal arteries in the thoracic region, and lumbar arteries in the lumbar region. The ventral spinal artery is continuous cranially with the basilar artery at the foramen magnum.
There are ventral internal vertebral venous plexuses on the floor of the vertebral canal in the epidural space (Fig. 6-4). These were seen previously in the atlanto-occipital region to be continuous cranially with the basilar sinus, a branch of the sigmoid sinus. Anastomoses occur between the plexus of each side and the branches of the vertebral and azygos veins and caudal vena cava.
Although this is a bilaterally symmetrical segmental system, there is often considerable variation in the size of various segmental vessels, including an occasional absence of one or more vessels.
Transverse Sections
Study transverse sections of the spinal cord at segments C4, C8, T4, T12, L2, L6, and S1 under a dissecting microscope. Compare the shape of the gray matter of these segments and relate it to their areas of innervation (Fig. 6-23).
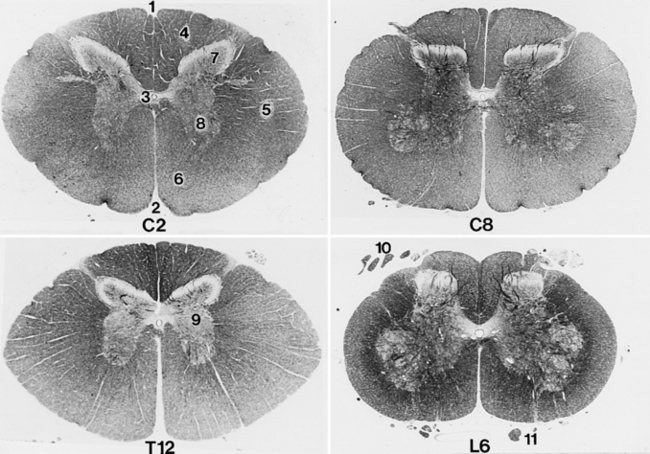
Fig. 6-23 Transverse section of spinal cord: second cervical segment (C2), eighth cervical segment (C8), twelfth thoracic segment (T12), and sixth lumbar segment (L6).
The gray matter of the spinal cord in transverse section is in the shape of a butterfly or the letter H. It consists primarily of neuronal cell bodies. The dorsal extremity on each side is the dorsal horn, which receives the entering dorsal (sensory) rootlets. The ventral extremity is the ventral horn, which sends axons out via the ventral (motor) rootlets. In the thoracolumbar region, the lateral horn projects laterally from the gray matter midway between the dorsal and ventral gray horns. This lateral horn contains the cell bodies of the preganglionic sympathetic neurons. In the center of the gray matter of the spinal cord is the central canal. This remnant of the embryonic neural tube is continuous rostrally with the fourth ventricle. At the caudal end of the conus medularis, there is a communication with the subarachnoid space.
The white matter of the spinal cord can be divided into three pairs of funiculi. Dorsally, a shallow, longitudinal groove extends the entire length of the spinal cord. This is the dorsal median sulcus. The longitudinal furrow along which the dorsal rootlets enter the spinal cord is the dorsolateral sulcus. Between these two sulci is the dorsal funiculus of the spinal cord.
Between the dorsolateral sulcus and the line of exit of the ventral rootlets, the ventrolateral sulcus, is located the lateral funiculus. The ventral funiculus is the white matter between the line of exit of the ventral rootlets and the longitudinal groove on the ventral side of the spinal cord, the ventral median fissure. In some species the funiculi have been subdivided topographically into specific cranially and caudally projecting tracts. Such anatomical information in domestic animals is still incomplete.