Chapter 17 Hearing
1. Sound waves are alternating phases of condensation and rarefaction (pressure waves) of molecules in the external environment.
2. Outer and middle ears funnel sound waves to the cochlea.
3. The cochlea is located in the inner ear.
4. The cochlea transduces sound waves to action potentials of the eighth cranial nerve.
5. Deciphering of sound wave frequency begins in the cochlea.
6. Action potentials from the cochlea are transmitted up through the brainstem to the cerebral cortex.
7. Deafness results from an interruption in the hearing process.
Our lives are enriched by music and conversation and altered by the sounds of danger. Many mammalian species have a particularly acute sense of hearing. Hearing depends on the remarkable properties of hair cell receptors in the cochlea that mediate transduction of sound into action potentials that are then sent to the brain. Fortunately, the auditory system is not usually a site of pathological lesions in veterinary medicine, except for occasional congenital defects and exposure to ototoxic drugs. Nevertheless, hearing is sufficiently important to warrant a brief discussion of its physiology.
Sound Waves Are Alternating Phases of Condensation and Rarefaction (Pressure Waves) of Molecules in the External Environment
Sound waves are longitudinal vibrations of molecules in the external environment characterized by alternating phases of condensation and rarefaction (increases and decreases in pressure). These alternating changes in pressure produce the sensation of sound when they strike the tympanic membrane and are subsequently transduced into neural signals that ultimately reach cerebral cortex. Sound waves reaching the tympanic membrane can be expressed as changes in sound pressure as a function of time (Figure 17-1).
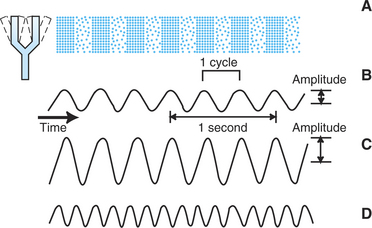
FIGURE 17-1 Characteristics of sound waves. A, Cyclical expansion and contraction of the tuning fork produce a cyclical compression and rarefaction of air molecules and a cyclical change in air pressure. B, Cyclical change in air pressure corresponding to a pure tone. The number of cycles per second is the frequency of the tone and is expressed in hertz (Hz). The frequency of the tone in B is 3 Hz. The amplitude of the wave reflects the magnitude of the pressure increase and is usually expressed in decibels (dB). C, Tone with a greater amplitude is perceived as louder than B. D, Tone with a greater frequency is perceived as having a higher pitch than B and C.
In general, the subjective loudness of the sound is correlated with the amplitude of a sound wave; the subjective pitch is correlated with the frequency of the wave. The amplitude of a sound is usually quantified according to the logarithmic decibel scale, which expresses the energy of the sound relative to the energy of a standard reference sound. This standard sound, representing 0 decibels (dB), is the threshold for human hearing where the sound of the background movement of air molecules can almost be heard. Normal conversation is about 60 dB, and the loudest tolerable sound for humans is about 120 dB, about 1 million times the threshold amplitude. Sound frequency, the number of pressure oscillation cycles per unit time, is usually expressed in units called hertz (Hz), where 1 Hz = 1 cycle per second.
Outer and Middle Ears Funnel Sound Waves to the Cochlea
The outer ear, composed of the fleshy part (pinna) and the ear canal, funnels sound waves to the tympanic membrane, or eardrum (Figure 17-2). Some animals can move the pinna to more effectively collect sound waves, and the natural shape of the pinna can act to filter selectively certain sound frequencies. The eardrum is a membrane between the outer and the middle ear. The middle ear is an air-filled cavity in the temporal bone and is connected to the nasopharynx by the auditory (eustachian) tube. Three tiny bones—the malleus, incus, and stapes—collectively called the ossicles, are connected to each other and are located in the middle ear. The malleus is connected to the eardrum, and the stapes is connected to the oval window, a membranous separation between the middle and the inner ear. The ossicles transfer vibration of the eardrum to the oval window in a manner that avoids a significant loss of energy as the sound wave is transferred from the air-filled outer ear to the fluid-filled inner ear. Two small skeletal muscles are also located in the middle ear, with one attached to the malleus and one attached to the stapes. Their contraction reduces the transfer of vibration between the eardrum and the oval window. This can function to protect the inner ear from very loud sounds.
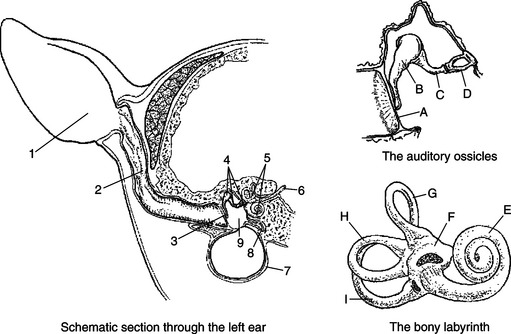
FIGURE 17-2 Schematic sections through the left ear, the auditory ossicles, and the bony labyrinth. 1, Pinna; 2, ear canal; 3, tympanic membrane; 4, auditory ossicles; 5, bony labyrinth; 6, eighth cranial nerve; 7, tympanic bulla; 8, eustachian tube; 9, middle ear; A, tympanic membrane; B, malleus; C, incus; D, stapes; E, cochlea; F,utricle; G, H, and I, semicircular canals.
(From Getty R: Atlas for applied veterinary anatomy, ed 2, Ames, 1964, Iowa State University Press.)
The Cochlea Is Located in the Inner Ear
The inner ear (labyrinth) contains the receptor organs of two sensory systems: the vestibular system, which detects acceleration and static tilt of the head (see Chapter 11), and the auditory system which detects and analyzes sound. The inner ear consists of the bony labyrinth and, within the bony labyrinth, the membranous labyrinth. The bony labyrinth is a series of tunnels within the petrous temporal bone. Inside these tunnels, surrounded by a fluid called perilymph, is the membranous labyrinth. The membranous labyrinth follows the contour of the bony labyrinth and contains endolymph. The vestibular and auditory portions of the inner ear are contiguous, and the “membranous tunnel within a bony tunnel” design is an anatomical feature of both parts. The auditory portion of this inner ear complex is called the cochlea (see Figure 11-1).
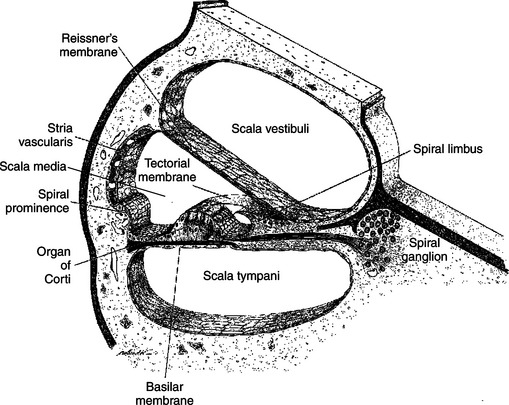
The cochlear portion of the labyrinth is coiled like a snail shell. If we could mentally uncoil this arrangement to a linear form and then take a transverse section through it, perpendicular to the long axis (like cutting a salami and then looking at the cut end), we would see two membranes, the basilar and Reissner’s, divide the cochlea into three chambers, or scalae (Figure 17-3). The dorsally located scala vestibuli and ventrally located scala tympani contain perilymph. The flexible middle scala, or scala media (cochlear duct), is formed by the membranous portion of the labyrinth and contains endolymph. The basilar membrane is the floor of the scala media, and atop this membrane lies the hair cell receptor organ for hearing called the organ of Corti. An anchored, gel-coated ridge, called the tectorial membrane, lies just atop the hair cells of the organ of Corti. The morphological organization just described is virtually the same all along the length of the cochlea, except that the scala vestibuli and scala tympani connect with each other at the distal end (farthest from the oval window).
The Cochlea Transduces Sound Waves to Action Potentials of the Eighth Cranial Nerve
The organ of Corti mediates the transduction of sound waves into action potentials. The hair cell receptors of the organ of Corti are similar in structure and function to the hair cells that form the vestibular sensory organs. The hair cells synapse on sensory neurons that form the cochlear portion of the eighth cranial (vestibulocochlear) nerve, which projects to the brainstem’s cochlear nuclei. The cell bodies of these sensory neurons reside in the spiral ganglion. Sound-induced bending of the hair cell cilia changes the frequency of action potentials on the eighth nerve fibers.
Sound waves in the external environment are collected by the outer ear and cause vibrations of the tympanic membrane. These vibrations are transmitted through the middle ear by movement of the ossicles and result in similar vibrations of the oval window of the cochlea. As the oval window vibrates, sound energy is transferred down through the perilymph of the scala vestibuli and down through the endolymph of the scala media to the basilar membrane. This energy produces a series of traveling waves that begin near the base of the basilar membrane (closest to the oval window) and move along its length. The situation is analogous to whipping the free end of a rope that is stationary at the opposite end. A diagram of this transmission is shown in Figure 17-4. The movement of the traveling wave causes portions of the flexible basilar membrane to move up and down. Because the organ of Corti sits atop the basilar membrane, this up-and-down motion causes the hair cell cilia to be sheared back and forth against the anchored, overlying tectorial membrane (Figure 17-5). This, in turn, changes the release of transmitter from the hair cells onto the eighth nerve neurons, which in turn alters the action potential firing rate of these neurons. It is at this point that the organ of Corti has transduced the sound wave energy into neural activity. As the amplitude of an environmental sound wave increases (normally perceived as a louder sound), a longer area of the basilar membrane is displaced. This results in a larger number of hair cells shearing against the tectorial membrane, which in turn affects the activity of a larger number of eighth nerve neurons. This is one way that sound intensity is coded by the nervous system.
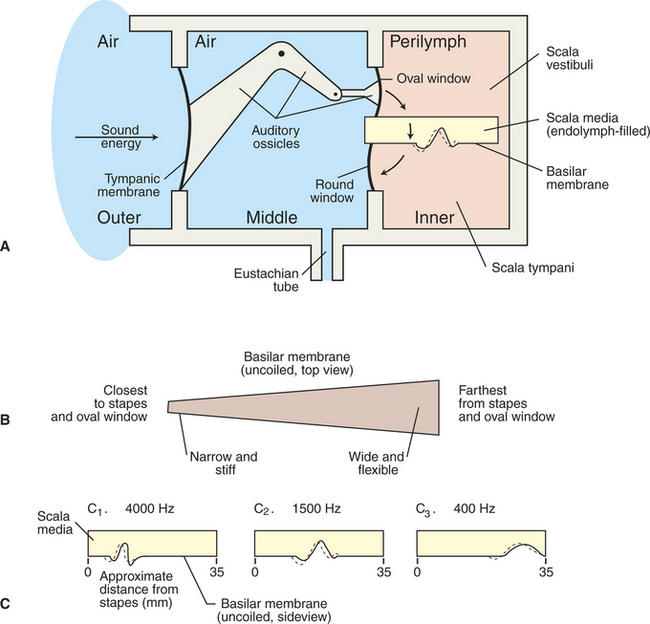
FIGURE 17-4 Schematic representation of the transmission of sound energy from the outer ear to the inner ear. A, Sound energy enters the air-filled outer ear and vibrates the tympanic membrane, which produces movement of the ossicles in the air-filled middle ear. Ossicular movement displaces the oval window of the fluid-filled inner ear, resulting in a traveling wave on the basilar membrane. B, Basilar membrane is narrow and stiff at the base and wider and more flexible farther from the base. C, As the frequency of a sound decreases, the region of maximum displacement of the basilar membrane, produced by the traveling wave, is located progressively farther from the base.
(Redrawn from Lippold OCJ, Winton FR: Human physiology, ed 6, New York, 1972, Churchill Livingstone.)
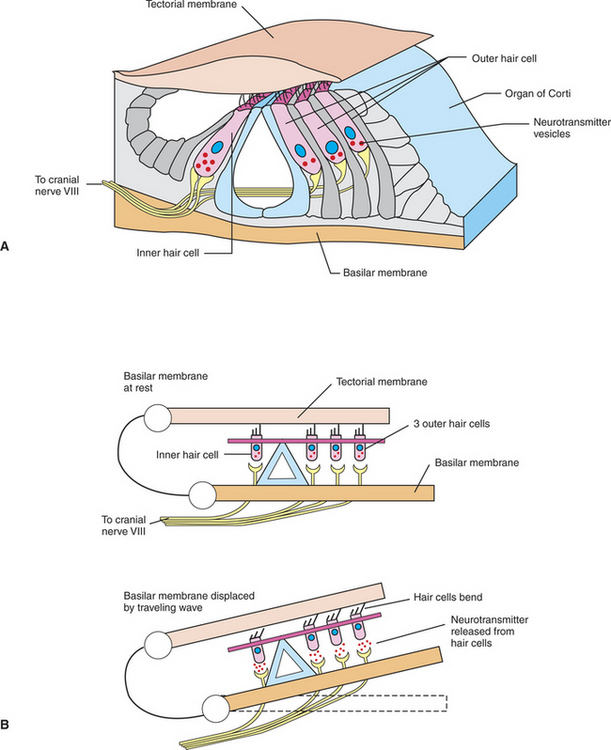
FIGURE 17-5 Transduction of sound into neural activity occurs at the organ of Corti. A, Structural organization of the organ of Corti and its hair cell receptors, sitting atop the basilar membrane. B, Traveling waves along the basilar membrane displace the membrane and organ of Corti upward, shearing hair cell cilia against the overlying tectorial membrane. This bending of the cilia induces the release of neurotransmitter from the hair cells onto sensory neurons of the eighth cranial nerve.
(Modified from Goldstein BE: Sensation and perception, ed 6, Pacific Grove, Calif, 2002, Wadsworth.)
Deciphering of Sound Wave Frequency Begins in the Cochlea
The physical properties of the basilar membrane are not uniform along its length. It is narrower and relatively stiffer at its base (near the oval window), and it becomes progressively wider and more flexible toward its apex (see Figure 17-4, B). Thus the resonant properties of the membrane are not uniform. A region near the base is significantly displaced by a high-frequency sound, and as the sound frequency decreases, the region of significant displacement is located progressively closer to the apex (see Figure 17-4, C). Because the organ of Corti sits atop the basilar membrane, high-frequency sounds are most likely to affect hair cells and their associated eighth nerve neurons near the base of the membrane. As frequency decreases, the hair cells and neurons that are activated are located progressively closer to the apex. Given this orderly relationship between the frequency of a sound wave and the region of the cochlea that is activated by that frequency, the cochlea is said to have a tonotopic organization. Therefore a principal means by which the nervous system begins to decipher the frequency of a sound is through the location of the hair cells and neurons that are most affected by that sound.
Action Potentials from the Cochlea Are Transmitted Up Through the Brainstem to the Cerebral Cortex
Action potentials arising in the cochlea travel along the cochlear portion of the eighth cranial nerve to the cochlear nuclei in the medulla oblongata. From there, neural activity is synaptically relayed, in a sequential manner, to the superior olivary nucleus (near the pontomedullary border), the inferior colliculus of the midbrain, the medial geniculate nucleus of the thalamus, and finally to the auditory cortex of the temporal lobe. Conscious perception of sound occurs in the cerebral cortex. Because of extensive connections of central auditory neurons across the midline, information originating in the cochlear nuclei on one side can reach other auditory nuclei on both sides of the brain. Each nucleus in the auditory pathway has a tonotopic representation of sound frequency but is specialized to process particular features of sound. For example, the superior olivary nucleus plays a major role in determining which side of the head an environmental sound source is coming from, whereas the medial geniculate nucleus is specialized to detect certain combinations of frequencies, as well as timing patterns among sounds.
Deafness Results from an Interruption in the Hearing Process
Clinical deafness may result from a loss of sound transmission in the outer or middle ear, called conduction deafness, or from malfunction of the cochlear hair cells or eighth nerve fibers, called nerve deafness. Given that auditory information from one ear is significantly distributed to both sides of the central nervous system, unilateral damage to the auditory system in the brain is often difficult to detect or to localize with a traditional hearing test. In veterinary medicine, deafness in young animals is usually caused by a congenital defect in the cochlea, frequently associated with white coat coloration. Certain antibiotics, diuretics, and antineoplastic agents have ototoxic properties capable of damaging cochlear structures.
CLINICAL CORRELATIONS
Congenital Deafness
History.
An owner brings you an 8-week-old, almost completely white, male Dalmatian and reports that the pup does not appear to hear anything.
Clinical Examination.
Your physical examination reveals a seemingly normal, healthy Dalmatian pup except for his apparent deafness. He does not seem to respond to voice commands or loud noises. Vestibular and all other neurological reflexes are within normal limits. A brainstem auditory-evoked response is flat, which suggests that the brain has not received any signal from the cochlea.
Comment.
Congenital deafness is fairly common in dogs and other animals with white coat color. It is usually caused by the partial or complete absence of the cochlea and occasionally by the absence of other neural elements in the auditory pathway. This is known as “nerve deafness” and is usually present at birth (congenital). Why it is linked to white coat coloration is unknown, but the pattern suggests that this is a genetically determined failure, usually bilateral, of the cochlea to develop. This dog can have a relatively normal life as long as the owners are attentive.
Bear MF, Connors BW, Paradiso MA. Neuroscience: exploring the brain, ed 3. Philadelphia: Lippincott, Williams & Wilkins, 2007.
Brodal P. The central nervous system: structure and function, ed 3. New York: Oxford University Press, 2004.
Goldstein BE. Sensation and perception, ed 6. Pacific Grove, Calif: Wadsworth, 2002.
Guyton AC, Hall JE. Textbook of medical physiology, ed 11. Philadelphia: Saunders, 2006.
Haines DE. Fundamental neuroscience, ed 3. New York: Churchill Livingstone, 2006.
Kandel ER, Schwartz JH, Jessell TM. Principles of neural science, ed 4, New York: McGraw-Hill, 2000.
Purves D, Augustine G, Fitzpatrick D, et al. Neuroscience, ed 3. Sunderland, Mass: Sinauer Associates, 2004.
Strain GM, Myers LJ. Hearing and equilibrium. Reece WO, ed. Duke’s physiology of domestic animals, ed 12, Ithaca, NY: Comstock Cornell University Press, 2004.
PRACTICE QUESTIONS
1. Which one of the following is the first to be displaced by sound energy entering the ear?
2. Hair cells similar to those of the organ of Corti are important for the function of which two of the following sensory organs?
3. Which one of the following statements is false?
4. Which one of the following cranial nerves transmits sound to the brain?
5. Of the following brain nuclei, which one receives auditory information last?