Chapter 11 The Vestibular System
1. The vestibular system is a bilateral receptor system located in the inner ear.
2. Specialized regions of the vestibular system contain receptors.
3. The semicircular ducts detect rotary acceleration and deceleration of the head.
4. The utricle and saccule detect linear acceleration and deceleration and static tilt of the head in space.
5. The vestibular system provides sensory information for reflexes involving spinal motor neurons, the cerebellum, and extraocular muscles of the eye.
6. Vestibular reflexes coordinate head and eye movements to maximize visual acuity during movements of the head.
To coordinate posture and locomotion, the brain needs to know not only what movement it intends to command, but also the orientation of the body and what movement the body is actually performing. Chapter 8 describes the muscle spindle, an important source of information for the brain about body position and movement. Another important source of information is the vestibular system. This bilateral receptor system is located in the inner ear and informs the brain about the position and motion of the head in space.
The vestibular system provides the organism with its sense of equilibrium or balance. It supplies information about the body’s orientation or tilt with respect to gravity and about acceleration of the body through space. It does this by detecting (1) static tilt of the head (e.g., the head is held stationary at 5 degrees from vertical), (2) linear acceleration of the head (e.g., the head accelerates in a straight line as an organism begins to run or as your elevator begins to rise), and (3) rotary acceleration of the head (e.g., the head accelerates in a circular fashion as an organism begins to turn its head toward a target or as someone begins to spin you in an office chair). This information is used most often to make automatic, unconscious postural adjustments to keep the organism from falling over as a result of self-imposed or environmentally imposed changes in body orientation. The vestibular system also helps to keep the eyes fixated on a relevant target in the face of such changes in body orientation.
The vestibular system is a common site of pathological lesions. In most veterinary species, lesions of the vestibular system cause a syndrome characterized by head tilt, compulsive rotary movements such as circling or rolling, and spontaneous nystagmus, which is an oscillating movement of the eyes.
To understand how such clinical signs arise and the importance of the vestibular system to the physiology of movement, you need to study its anatomy and function first.
The Vestibular System Is a Bilateral Receptor System Located in the Inner Ear
The inner ear, or labyrinth, is made up of two parts: the bony labyrinth and the membranous labyrinth. The bony labyrinth is a system of caverns and tunnels through the petrous temporal bone of the skull. The bony labyrinth houses the receptor organs of the vestibular system as well as the receptor organ for hearing, the cochlea (Figure 11-1) (see Chapter 17). These receptor organs are part of the membranous labyrinth, which consists of thin membranes of epithelium and lies within the bony labyrinth. This epithelial membrane is specialized at some locations to become the sensory receptor cells that form the vestibular and auditory receptor organs. The membranous labyrinth is filled with a fluid called endolymph and is separated from the bony labyrinth by a fluid called perilymph. The vestibular portion of the membranous labyrinth consists of two major sets of structures: (1) three semicircular ducts, located at approximately right angles to each other, and (2) a pair of saclike structures called the utricle and saccule, sometimes called the otolith organs. As discussed later, each major set of structures is respectively involved in transducing a different major class of vestibular stimulus.
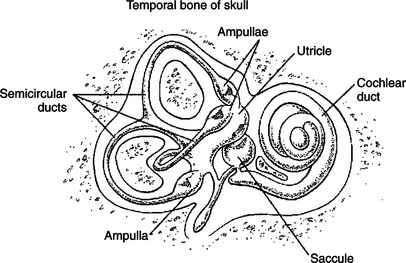
FIGURE 11-1 The bilateral inner ear contains receptor systems for hearing (cochlea) and for detecting the orientation and acceleration/deceleration of the head (vestibular system). The peripheral vestibular apparatus on each side of the head contains a utricle, a saccule, and three semicircular ducts, each with an ampulla at one end.
Specialized Regions of the Vestibular System Contain Receptors
Each vestibular structure of the membranous labyrinth has a region of epithelial lining that has become specialized into a set of secondary receptor cells (see Chapter 7) called hair cells (Figure 11-2). These hair cells form the basis of a sensory receptor organ within each vestibular structure. Each hair cell has several hairlike cilia at its apex that are arranged in order according to size. At its base the hair cell synapses with a sensory neuron that carries action potentials to the brainstem. The cell bodies of these sensory neurons are located in Scarpa’s ganglia, and their axons collectively form the vestibular portion of the vestibulocochlear nerve (cranial nerve VIII). The hairlike cilia from all the hair cells within any one vestibular structure project into a gelatinous mass; displacement of this gelatinous mass in a given direction causes all the hair cell cilia to bend in that direction.
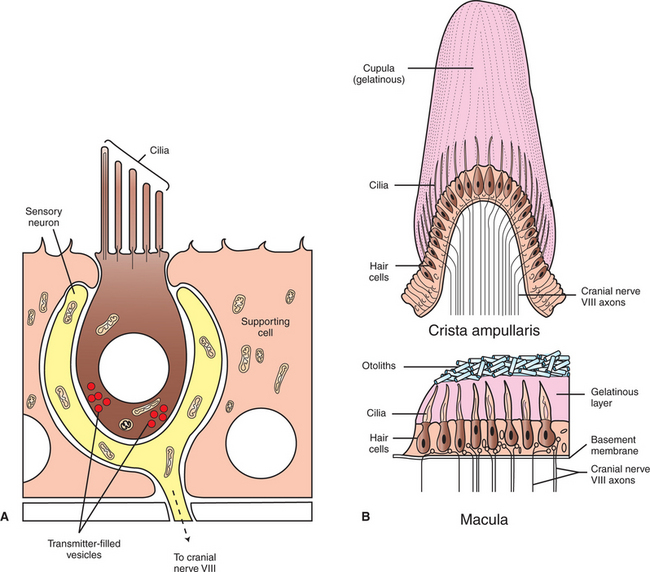
FIGURE 11-2 Each structure of the peripheral vestibular apparatus contains a region of hair cells that form the basis of a sensory receptor organ. A, Each hair cell has several cilia at its apex, arranged in size order, and synapses on a sensory neuron of cranial nerve VIII at its base. B, In each ampulla of the semicircular ducts, there is a crest of hair cells whose cilia project into a gelatinous mass called the cupula, forming a receptor organ called the crista ampullaris. The hair cell receptor organ in the utricle and saccule is the macula, a layer of hair cells whose cilia project up into a gelatinous layer, on top of which lies a layer of calcium carbonate crystals called otoliths.
At rest, when the cilia are not deflected, the sensory neurons that synapse with the vestibular hair cells transmit action potentials spontaneously at about 100 per second (Figure 11-3). When the hair cell cilia are bent in a direction toward the largest cilium, the hair cells depolarize, the release of transmitter from the hair cell onto the sensory neuron increases, and the neuron’s action potential frequency increases. When the cilia are bent in the opposite direction, toward the smaller cilia, hair cell membranes hyperpolarize, transmitter release decreases, and the action potential frequency of sensory neurons decreases. Therefore, displacement of the hair cell cilia in either of these directions can be detected by the brain as an increase or a decrease from the resting action potential frequency. Deflections in other directions are much less effective. How the brain uses this information to detect the direction of head movement is described later.
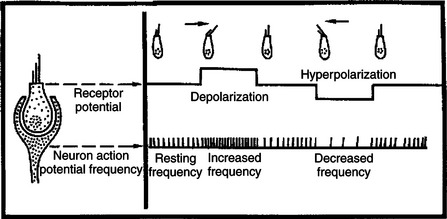
FIGURE 11-3 At rest, sensory neurons on which vestibular hair cells synapse transmit action potentials spontaneously at a rate of about 100 per second. When hair cell cilia are deflected in one direction, the action potential frequency increases; when cilia are deflected in the opposite direction, the frequency decreases.
The Semicircular Ducts Detect Rotary Acceleration and Deceleration of the Head
Three membranous semicircular ducts are located within corresponding semicircular canals of each bony labyrinth (Figure 11-4). They are positioned at approximately right angles to each other, and both ends of each fluid-filled duct terminate in the utricle. Each semicircular duct has an enlargement at one end, called the ampulla, near its junction with the utricle. The ampulla contains a hair cell receptor organ called the crista ampullaris (see Figure 11-2). This is a ridge of hair cells that, at their base, synapse on sensory neurons projecting to the central nervous system (CNS) and whose cilia are embedded in an overlying gelatinous mass. This mass, called the cupula, attaches to the roof of the ampulla (Figure 11-5). All hair cells of a given crista ampullaris are oriented in the same direction with respect to their cilia. Together, the hair cell ridge and overlying cupula span the diameter of the ampulla.
FIGURE 11-4 Three semicircular ducts, each positioned at approximately right angles to the other two, are located on each side of the head and work to detect rotary acceleration and deceleration of the head.
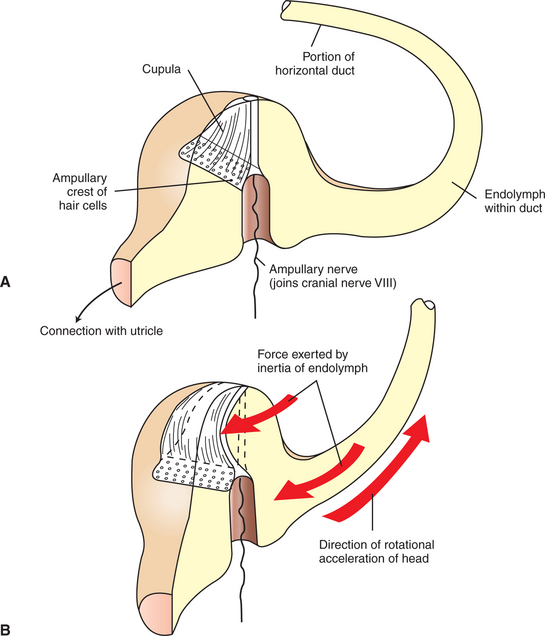
FIGURE 11-5 Ampullae of the semicircular ducts contain a crista ampullaris, which transduces rotational acceleration/deceleration of the head. The ducts are filled with endolymph. A, crista ampullaris of the horizontal canal when the head is at rest. B, on rotational acceleration of the head in the indicated direction, the relative inertia of the endolymphatic fluid displaces the cupula, and thus the hair cell cilia, in the opposite direction.
(Modified from Kandel ER, Schwartz JH, editors: Principles of neural science, ed 2, New York, 1985, Elsevier Science Publishing.)
The semicircular ducts, together with the ampulla and its contents, are involved in transducing rotary acceleration and deceleration of the head. When the head begins to accelerate in a rotary fashion, the semicircular duct and its receptor organ rotate with the head, but the endolymph’s acceleration lags behind because of inertia. This relative difference in the rate of acceleration of the semicircular duct and its enclosed endolymph causes the crista ampullaris to “crash into” the slower-moving endolymph. This results in a displacement of the gelatinous cupula in the direction opposite that of head rotation and a corresponding bending of the hair cells. This in turn changes the firing rate of the sensory neurons projecting to the CNS. The opposite happens with deceleration because the semicircular duct and crista ampullaris slow immediately along with the head while inertia continues to carry the endolymph forward. Stimulation of the crista ampullaris occurs only on rotary acceleration or deceleration of the head, not during constant velocity, since the movement of the endolymph will eventually catch up with the movement of the semicircular ducts, and the hair cells will no longer be bent.
Semicircular ducts located on opposite sides of the head, but in approximately the same plane (co-planar), work as a pair to provide the brain with information about the direction and nature of head movement. For instance, a clockwise rotary acceleration of the head would cause bending of the directionally sensitive hair cell cilia in each member of a co-planar pair of semicircular ducts on opposite sides of the head. However, the sensory axons leaving the crista ampullaris from the duct on one side of the head would carry an increased action potential frequency, whereas those from the duct of the other side would carry a decreased action potential frequency. The brain interprets such reciprocal changes in sensory action potential frequency as resulting from clockwise or counterclockwise acceleration or deceleration in a given plane of movement. In reality, rotary acceleration/deceleration in any given plane usually affects all three sets of paired semicircular ducts, but each pair to different degrees. In this way, the bilateral system of six semicircular ducts detects the direction of both rotary acceleration and deceleration of the head and activates or inhibits particular CNS structures to produce the appropriate reflex response.
The Utricle and Saccule Detect Linear Acceleration and Deceleration and Static Tilt of the Head in Space
In the utricle and saccule, the hair cell receptor organ is called the macula (see Figure 11-2). It is an oval patch of hair cells with a primarily horizontal orientation on the floor of the utricle and a primarily vertical orientation on the wall of the saccule. The cilia of the macular hair cells extend up into a gelatinous layer atop the hair cells. Embedded at the top of this gelatinous mass is a layer of calcium carbonate crystals called otoliths. This otolith layer is heavier and denser than the endolymph and other surrounding materials.
Given the organization of the receptor apparatus that lies within the utricle and saccule, these vestibular structures can transduce linear acceleration and deceleration of the head, as well as static head tilt (Figure 11-6). Considering the horizontally oriented macula of the utricle, if the head is accelerated forward in a straight line, movement of the dense otolith layer lags behind that of the macular hair cells. This produces a shearing force that bends the tips of the hair cell cilia, by way of the gelatinous layer, until constant velocity is achieved and the otolith layer catches up with the hair cell layer. In contrast to the crista ampullaris, not all the hair cell clusters of a given macula are oriented in the same direction with respect to their cilia (Figure 11-7). In addition, as previously noted, hair cells of the utricle are oriented in the horizontal plane, whereas those of the saccule are in the vertical plane. Therefore, linear acceleration in a specific direction will bend hair cells of a particular location and orientation in a way that will transiently increase the action potential firing rate of their associated sensory neurons; those of another location and orientation will be bent in a way that transiently decreases the firing rate; and those of yet another location and orientation will be bent in a way that has little or no effect on firing rate. This topographic pattern of hair cell bending and associated transient changes in action potential firing will be different for linear acceleration in a different direction. The CNS can decipher these various patterns of neural activity to determine the onset and direction of linear acceleration and to initiate an appropriate compensatory response.
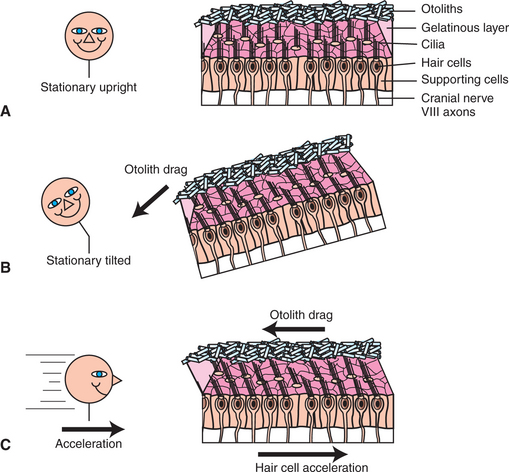
FIGURE 11-6 Utricle and saccule each contain a macula, which transduces static head tilt and linear acceleration/deceleration of the head. macula of the utricle is horizontally oriented. A, When the head is stationary and upright, there is little or no bending of the hair cell cilia. B, When the head tilts and remains tilted, the heavy otolith layer “falls over,” producing a drag. This bends the hair cell cilia, by way of the interposed gelatinous layer, in the direction of the tilt. C, When the head accelerates in a straight line, the hair cells accelerate in the same direction, but the heavy otolith layer lags behind, producing a drag in the opposite direction. This bends the hair cell cilia, by way of the interposed gelatinous layer, in the direction opposite the acceleration.
(Portions modified from Purves D, Augustine G, Fitzpatrick D, et al: Neuroscience, ed 3, Sunderland, Mass, 2004, Sinauer Associates.)
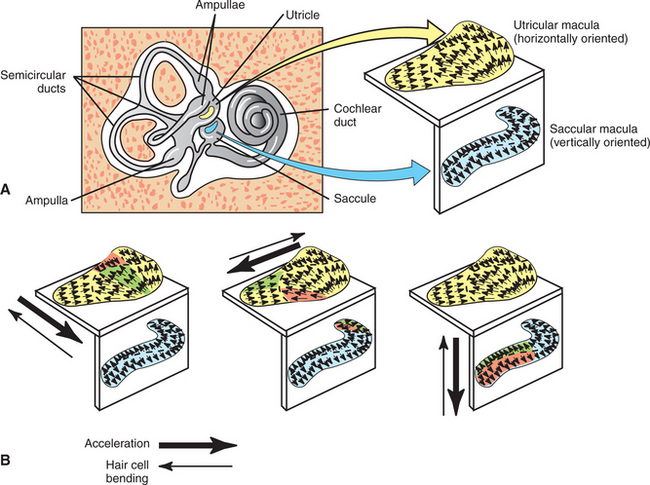
FIGURE 11-7 A, Macula of the utricle is horizontally oriented, and macula of the saccule is vertically oriented. Small arrows in a macula represent the approximate orientation of the hair cells in that region, with respect to their cilia. For a given hair cell, the arrow tip represents the position of the largest cilium, and the arrow tail represents the shortest cilium. B, Acceleration in a given direction (large thick arrows) results in bending of hair cell cilia in the opposite direction (large thin arrows) caused by otolith drag. Hair cells whose cilia are bent directly toward the largest cilium (green regions) will be depolarized the most and will produce the greatest increase in action potential frequency in their associated sensory neurons. Conversely, hair cells whose cilia are bent directly away from the largest cilium (red regions) will be hyperpolarized the most and will produce the greatest decrease in action potential frequency in their associated sensory neurons. Hair cells whose cilia are bent along other axes will be less significantly affected.
(Portions modified from Fuchs AF: Peripheral motor control: the vestibular system. In Patton HD, Fuchs AF, Hille B, et al, editors: Textbook of physiology, ed 21, Philadelphia, 1989, Saunders.)
Again considering the horizontally oriented macula of the utricle, when the head tilts from the upright position, the heavy and dense otolith layer effectively “falls over” as it is pulled by gravity (see Figure 11-6, B). This bends the hair cell cilia, by way of the gelatinous layer, and keeps them bent as long as the head is tilted. This sustained bending (compared with the transient bending during linear acceleration) is translated into sustained changes in action potential firing frequency (compared to transient changes during linear acceleration) in particular populations of associated sensory neurons. In this way the utricle and saccule can inform the brain about a stationary tilt of the head. The direction of the tilt is detected by a similar mechanism as that for detecting the direction of linear acceleration. Astronauts in low gravitational settings receive relatively little information from their utricles and saccules about their stationary head position and must rely more heavily on visual and other sensory cues to detect head position.
The Vestibular System Provides Sensory Information for Reflexes Involving Spinal Motor Neurons, the Cerebellum, and Extraocular Muscles of the Eye
As noted earlier, vestibular hair cells synapse on sensory neurons whose axons form part of the eighth cranial (vestibulocochlear) nerve and that carry action potentials to the medulla. Almost all these axons synapse in the vestibular nuclear complex, a bilateral group of four distinct nuclei occupying a substantial portion of the medulla beneath the fourth ventricle. From here, second-order neurons (those on which cranial nerve VIII axons synapse) project to three important areas of the nervous system. Some of the neurons of the vestibular nuclear complex receive significant input from the utricle and saccule (the otolith organs), and their axons in turn form the lateral vestibulospinal tract. This tract provides excitatory facilitation to gamma (γ) and alpha (α) motor neurons of antigravity muscles of the trunk and limbs in response to linear acceleration/deceleration or static tilt of the head (see Chapter 10).
Other neurons of the vestibular nuclear complex receive significant sensory input from the cristae ampullaris of the semicircular ducts, and their axons in turn form a pathway that projects to cranial nerve nuclei that control eye movements. This pathway, called the medial longitudinal fasciculus (MLF), produces compensatory eye movements in response to rotary acceleration/deceleration of the head. The vestibular nuclear complex also sends projections to, and receives projections from, the cerebellum, especially the flocculonodular lobe. Through these reciprocal connections, the cerebellum can fine-tune the coordination of postural and visuomotor reflexes that are controlled by the vestibular system. Finally, some of the projections leaving the vestibular nuclear complex participate in neural circuits leading to cerebral cortex, resulting in conscious vestibular sensations.
Vestibular Reflexes Coordinate Head and Eye Movements to Maximize Visual Acuity During Movements of the Head
Vestibular reflex control of the extraocular muscles of the eye, known as the vestibuloocular reflex (VOR), coordinates eye and head movements so that as the head turns (rotates), the eyes remain fixed on the original field of vision for as long as possible. Imagine that a dog is seated on a piano stool and you rotated her clockwise to the right. As you rotate her slowly to the right, her eyes rotate in her head slowly to the left so that the eyes remain fixed on the same field of vision as long as possible. As the eyes reach the limit of their leftward excursion, they swiftly move to the right, in the direction of the head movement, until they fix on a new field of vision. If the head continues to rotate, the cycle is repeated until constant velocity is achieved. This allows the animal time to interpret a field of vision despite rotary acceleration of the head. When this pattern of eye movement occurs during the VOR, it is referred to as normal nystagmus or physiological nystagmus; the slow drift opposite head rotation followed by a fast flick back in the direction of head rotation. A transient postrotatory nystagmus, with the converse eye movement pattern to that just noted, can be seen if a spinning animal or person is suddenly stopped. Inertia of the endolymph causes it to continue rotating in the semicircular duct, pushing on the crista ampullaris, even though the head and duct have stopped moving.
These reflex eye movement patterns require normal sensory input from the semicircular ducts, an intact MLF in the brainstem, and normally functioning motor units of the extraocular muscles (as well as an intact cerebellum). The VOR occurs on rotation of the head in the horizontal plane or the vertical plane, or on clockwise or counterclockwise torsional rotation of the head. Voluntary control of the eyes is independent of vestibular reflexes and is controlled by the cerebral cortex.
Nystagmus may appear occasionally under pathological conditions of the vestibular system, even when the head is straight and at rest. This condition is known as spontaneous nystagmus. A persisting head tilt, falling, and compulsive circling or rolling often accompany spontaneous nystagmus in animals with acute vestibular disease. These actions often tend to be oriented in a consistent pattern with respect to the side of a peripheral lesion of the vestibular system. This is presumably a result of abnormal, asymmetric action potential inputs to the brainstem from the vestibular apparatus on the two sides of the head.
CLINICAL CORRELATIONS
Vestibular Syndrome
History.
A 3-year-old male cocker spaniel is brought to your clinic. The owner states that for the previous 2 days, the dog has held his right ear lower than his left ear. He also tends to walk in circles, clockwise to the right. You have treated this dog previously for an infection of the outer right ear.
Clinical Examination.
On physical examination of the dog, you find that the outer ear infection persists. You also confirm that the dog persistently tilts his head with the right ear down and circles to the right; you find that he has a spontaneous horizontal nystagmus. Results of the remaining physical and neurological examination are within normal limits.
Comment.
Head tilt, circling, and spontaneous nystagmus constitute a common constellation of clinical signs often called the vestibular syndrome. It results from abnormality in the vestibular system, usually in the membranous labyrinth. It is frequently caused by the extension of an infection from the outer and middle ear to the labyrinth of the inner ear. This results in an abnormal balance of action potential frequencies between the normal and abnormal sides of the vestibular system, causing asymmetric stimulation of the ocular and postural reflex mechanisms normally controlled by the vestibular nuclei.
Treatment.
When such labyrinthitis is caused by bacterial infection, treatment with appropriate antibiotics is often effective in eliminating the clinical signs by returning the peripheral receptor to its normal function. In cases of idiopathic vestibular syndrome in older cats and dogs, spontaneous recovery, without treatment, is common.
Bear MF, Connors BW, Paradiso MA. Neuroscience: exploring the brain, ed 3. Philadelphia: Lippincott Williams & Wilkins, 2007.
Brodal P. The central nervous system: structure and function, ed 3. New York: Oxford University Press, 2004.
Fuchs AF. Peripheral motor control: the vestibular system. Patton HD, Fuchs AF, Hille B, et al. Textbook of physiology, ed 21, Philadelphia: Saunders, 1989.
Guyton AC, Hall JE. Textbook of medical physiology, ed 11. Philadelphia: Saunders, 2006.
Haines DE. Fundamental neuroscience. New York: Churchill Livingstone, 1997.
Kandel ER, Schwartz JH, Jessell TM. Principles of neural science, ed 4, New York: McGraw-Hill, 2000.
Purves D, Augustine G, Fitzpatrick D, et al. Neuroscience, ed 3. Sunderland, Mass: Sinauer Associates, 2004.
PRACTICE QUESTIONS
1. The receptor organ detecting rotary acceleration and deceleration of the head is located in the:
2. Which two of the following are not generally associated with the macula?
3. You are presented with a dog with a head tilt, compulsive circling, and spontaneous nystagmus. The most likely site of this dog’s pathological lesion is the:
4. Which one of the following statements is false?
5. If a normal dog is sitting on a piano stool and I start to spin (accelerate) the stool to the right, which two of the following will be false regarding the observed nystagmus?