Chapter 2 Human Reproduction
Puberty begins when secondary sex characteristics (e.g., pubic hair) appear, typically between the ages of 12 and 15 years in females and 13 and 16 years in males. Menarche (the time of the first menstrual period) may occur as early as 8 years of age. Puberty in females is largely completed by age 16 years. In males, puberty ends when the first mature sperms are formed.
Reproductive Organs
Reproductive organs produce and transport germ cells (gametes) from the gonads (testes or ovaries) to the site of fertilization in the uterine tube (Fig. 2-1).
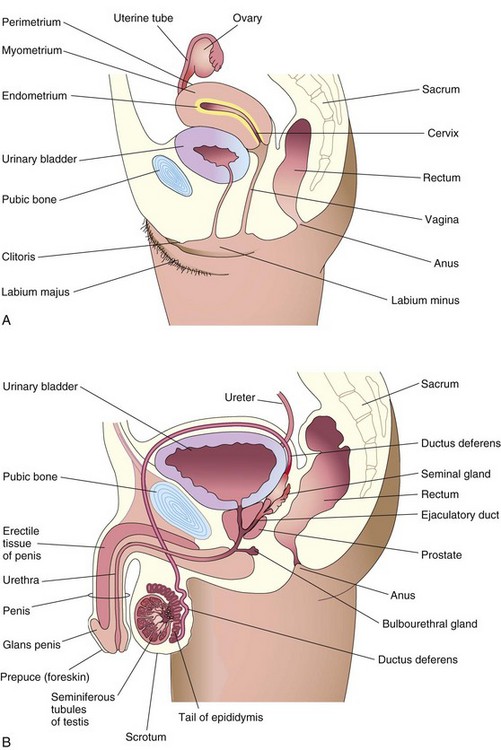
Female Reproductive Organs
Vagina
The vagina (see Fig. 2-1A) serves as the excretory passage for menstrual fluid, receives the penis (see Fig. 2-1B) during sexual intercourse, and forms the inferior part of the birth canal.
Uterus
The uterus is a thick-walled, pear-shaped organ (see Fig. 2-2A and B) and consists of two main parts:
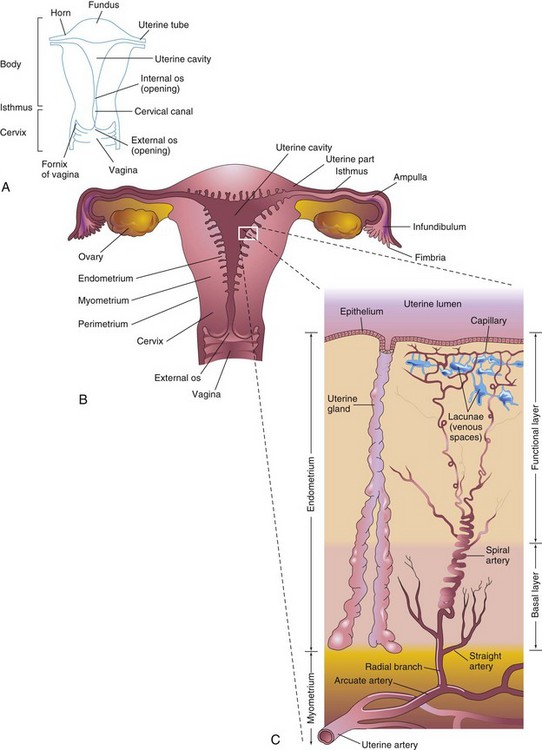
Figure 2–2 Female reproductive organs. A, Parts of the uterus. B, Diagrammatic frontal (coronal) section of the uterus, uterine tubes, and vagina. The ovaries are also shown. C, Enlargement of the area outlined in B. The functional layer of the endometrium is sloughed off during menstruation.
The fundus of the uterus is the rounded part of the uterine body that lies superior to the orifices of the uterine tubes. The body of the uterus narrows from the fundus to the isthmus, the constricted region between the body and the cervix (Fig. 2-2A). The lumen of the cervix, the cervical canal, has a constricted opening, the os (ostium), at each end. The internal os communicates with the cavity of the body of the uterus, whereas the external os communicates with the vagina. The walls of the body of the uterus consist of three layers:
At the peak of its development, the endometrium is 4 to 5 mm thick. During the luteal (secretory) phase of the menstrual cycle (see Fig. 2-8), three layers of the endometrium can be distinguished microscopically (see Fig. 2-2C) into the following:
• A compact layer, consisting of densely packed connective tissue around the neck of the uterine glands
• A spongy layer, composed of edematous connective tissue containing the dilated, tortuous bodies of the uterine glands
• A basal layer, containing the blind ends of the uterine glands
The compact and spongy layers, known collectively as the functional layer, disintegrate and are shed at menstruation and after parturition (childbirth). The basal layer of the endometrium has its own blood supply and is not cast off during menstruation.
Uterine Tubes
The uterine tubes, measuring 10 cm long and 1 cm in diameter, extend laterally from the horns of the uterus (see Fig. 2-2A). Each tube opens into a horn of the uterus at its proximal end and into the peritoneal cavity at its distal end. The uterine tube is divided into the following parts: the infundibulum, the ampulla, the isthmus, and the uterine part. The tubes carry oocytes from the ovaries to the fertilization site in the ampulla (see Fig. 2-2B). The uterine tube then conveys the dividing zygote to the uterine cavity.
Ovaries
The ovaries are almond-shaped glands located on each side of the uterus that produce oocytes (see Fig. 2-1A and B). When released from the ovary at ovulation, the secondary oocyte passes into one of two uterine tubes. These tubes open into the uterus, which protects and nourishes the embryo and fetus until birth. The ovaries also produce estrogen and progesterone, the hormones responsible for the development of secondary sex characteristics and regulation of pregnancy.
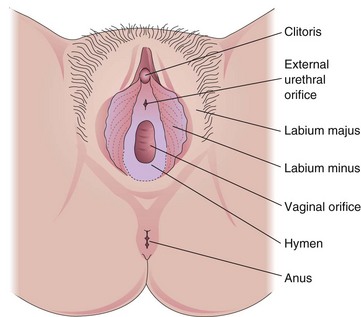
Female External Sex Organs
The female external sex organs are known collectively as the vulva (Fig. 2-3). The labia majora, fatty external folds of skin, conceal the vaginal orifice, the opening of the vagina. Inside these labia are two smaller folds of mucous membrane, the labia minora. The clitoris, a small erectile organ, is situated at the superior junction of these folds. The vagina and urethra open into a cavity, the vestibule (the cleft between the labia minora). The vaginal orifice varies with the condition of the hymen, a fold of mucous membrane that surrounds the vaginal orifice.
Male Reproductive Organs
The male reproductive organs (see Fig. 2-1B) include the penis, testes, epididymis, ductus deferens (vas deferens), prostate, seminal glands, bulbourethral glands, ejaculatory ducts, and urethra. The two oval testes are located in the cavity of the scrotum. Each testis consists of many highly coiled seminiferous tubules that produce sperms. Immature sperms pass from the testis into a single, complexly coiled tube, the epididymis, where they are stored. From the epididymis, the ductus deferens carries the sperms to the ejaculatory duct. This duct descends into the pelvis, where it fuses with the ducts of the seminal glands to form the ejaculatory duct, which enters the urethra.
The urethra is a tube that leads from the urinary bladder through the penis to the outside of the body. Within the penis, erectile tissue surrounds the urethra. During sexual excitement, this tissue fills with blood, causing the penis to erect. Semen consists of sperms mixed with seminal fluid produced by the seminal glands, bulbourethral glands, and prostate.
Gametogenesis
The sperm and oocyte are highly specialized gametes, or germ cells (Fig. 2-4). Each of these cells contains half the number of required chromosomes (i.e., 23 instead of 46). The number of chromosomes is reduced during a special type of cell division called meiosis. This type of cell division occurs during gametogenesis (formation of germ cells). In males, this process is termed spermatogenesis; in females, it is known as oogenesis (Fig. 2-5).
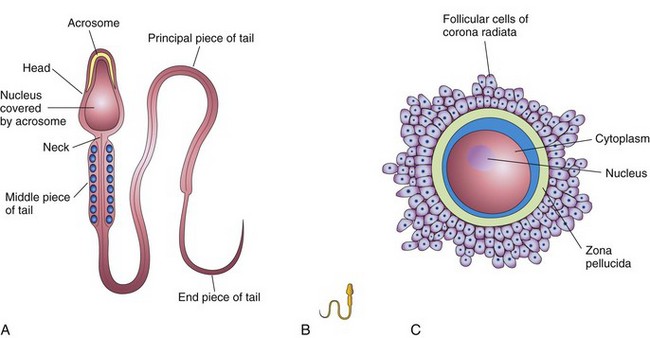
Figure 2–4 Male and female gametes or germ cells. A, The parts of a human sperm (×1250). The head, composed mostly of the nucleus, is partly covered by the acrosome, an organelle containing enzymes. B, A sperm drawn to approximately the same scale as the oocyte. C, A human secondary oocyte (×200), shown surrounded by the zona pellucida and corona radiata.
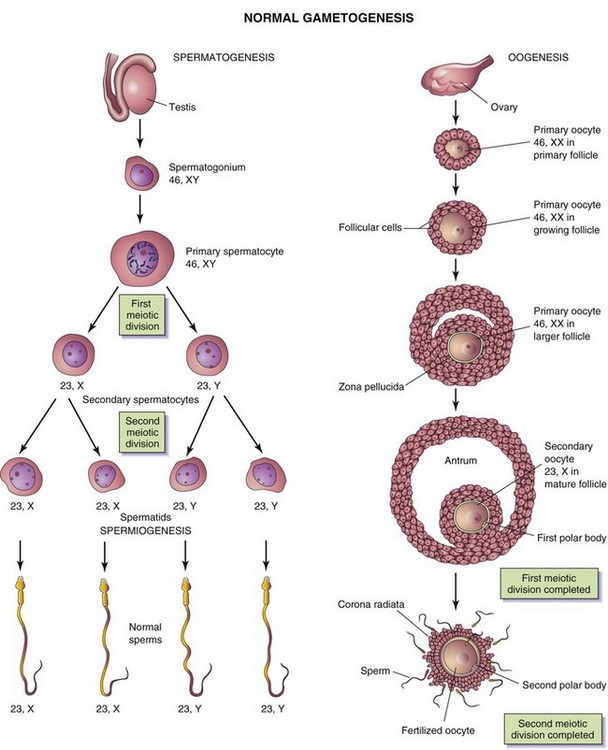
Figure 2–5 Normal gametogenesis: conversion of germ cells into gametes. The illustrations compare spermatogenesis and oogenesis. Oogonia are not shown in this figure because they differentiate into primary oocytes before birth. The chromosome complement of the germ cells is shown at each stage. The number designates the total number of chromosomes, including sex chromosome(s) (shown after the comma). Note: (1) After the two meiotic divisions, the diploid number of chromosomes, 46, is reduced to the haploid number, 23; (2) four sperms form from one primary spermatocyte, whereas only one secondary oocyte results from maturation of a primary oocyte; (3) the cytoplasm is conserved during oogenesis to form one large cell, the oocyte.
Meiosis
Meiosis consists of two meiotic cell divisions (Fig. 2-6), during which the chromosome number of the germ cells is reduced to half (23, the haploid number) the number present in other cells in the body (46, the diploid number).
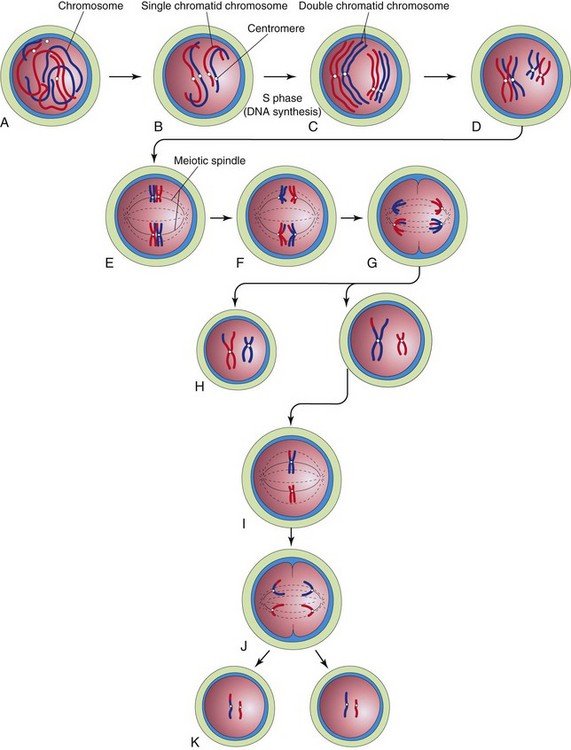
Figure 2–6 Diagrammatic representation of meiosis. Two chromosome pairs are shown. A to D, Stages of prophase of the first meiotic division. The homologous chromosomes approach each other and pair; each member of the pair consists of two chromatids. Observe the single crossover in one pair of chromosomes, resulting in the interchange of chromatid segments. E, Metaphase. The two members of each pair become oriented on the meiotic spindle. F, Anaphase. G, Telophase. The chromosomes migrate to opposite poles. H, Distribution of parental chromosome pairs at the end of the first meiotic division. I to K, Second meiotic division, which is similar to mitosis, except that the cells are haploid.
During the first meiotic division, the chromosome number is reduced from diploid to haploid. Homologous chromosomes (one from each parent) pair during prophase and then separate during anaphase, with one representative of each pair randomly going to each pole of the meiotic spindle. The spindle connects to the chromosome at the centromere (Fig. 2-6B). At this stage, they are double chromatid chromosomes. The X and Y chromosomes are not homologs; however, they have homologous segments at the tips of their short arms. They pair in these regions only. By the end of the first meiotic division, each new cell formed (secondary spermatocyte or secondary oocyte) has the haploid chromosome number of double chromatid chromosomes; therefore, each cell contains half the number of chromosomes of the preceding cell (primary spermatocyte or primary oocyte). This separation, or disjunction, of paired homologous chromosomes is the physical basis of segregation, or separation, of allelic genes during meiosis.
The second meiotic division follows the first division, without a normal interphase (i.e., without an intervening step of DNA replication). Each double chromatid chromosome divides, and each half, or chromatid, is randomly drawn to a different pole of the meiotic spindle; thus, the haploid number of chromosomes (23) is retained. Each daughter cell formed by meiosis has the reduced haploid number of chromosomes, with one representative of each chromosome pair (now a single chromatid chromosome).
• Provides for constancy of the chromosome number from generation to generation by reducing the chromosome number from diploid to haploid, thereby producing haploid gametes.
• Allows random assortment of maternal and paternal chromosomes between the gametes.
• Relocates segments of maternal and paternal chromosomes by crossing over of chromosome segments, which “shuffles” the genes and produces a recombination of genetic material.
Spermatogenesis
Before puberty, primordial sperms (spermatogonia) remain dormant in the seminiferous tubules of the testes from the late fetal period. At puberty they begin to increase in number (see Fig. 2-5). After several mitotic cell divisions, the sperms grow and undergo gradual changes that transform them into primary spermatocytes—the largest germ cells in the seminiferous tubules. Each primary spermatocyte subsequently undergoes a reduction division—the first meiotic division—to form two haploid secondary spermatocytes, which are approximately half the size of primary spermatocytes. Subsequently, the secondary spermatocytes undergo a second meiotic division to form four haploid spermatids, which are approximately half the size of secondary spermatocytes. The spermatids are gradually transformed into four mature sperms during a process known as spermiogenesis. During this metamorphosis (change in form), the nucleus condenses, the acrosome forms, and most of the cytoplasm is shed and the tail forms. When spermiogenesis is complete, sperms enter the lumina (cavities) of the seminiferous tubules. The sperms then move to the epididymis (see Fig. 2-1B), where they are stored and become functionally mature. Spermatogenesis, including spermiogenesis, requires approximately 2 months for completion. Spermiogenesis normally continues throughout the reproductive life of a male.
When ejaculated, the mature sperms are free-swimming, actively motile cells consisting of a head and a tail (see Fig. 2-4A). The neck of the sperm is the junction between the head and tail. The head of the sperm, forming most of the bulk of the sperm, contains the nucleus. The anterior two thirds of the head are covered by the acrosome, a cap-like organelle containing enzymes that facilitate sperm penetration during fertilization. The tail provides the motility of the sperm, assisting with its transport to the site of fertilization in the ampulla of the uterine tube. The tail of the sperm consists of three parts: the middle piece, principal piece, and end piece. The middle piece contains the energy-producing mitochondria, which fuel the lashing movements of the tail.
Oogenesis
Oogenesis refers to the sequence of events by which oogonia (primordial oocytes) are transformed into oocytes. This maturation process begins during the fetal period, but is not completed until after puberty (see Fig. 2-5). During early fetal life oogonia proliferate by mitosis and enlarge to form primary oocytes. At birth, all primary oocytes have completed the prophase of the first meiotic division. These oocytes remain in prophase until puberty. Shortly before ovulation, a primary oocyte completes the first meiotic division (see Fig. 2-5). Unlike the corresponding stage of spermatogenesis, the division of cytoplasm is unequal. The secondary oocyte receives almost all the cytoplasm, whereas the first polar body receives very little, causing it to degenerate after a short time. At ovulation, the nucleus of the secondary oocyte begins the second meiotic division, but progresses only to metaphase.
If the secondary oocyte is fertilized by a sperm, the second meiotic division is completed and a second polar body is also formed (see Fig. 2-5). The secondary oocyte released at ovulation is surrounded by a covering of amorphous material known as the zona pellucida and a layer of follicular cells called the corona radiata (see Fig. 2-4C). The secondary oocyte is large, being just visible to the unaided eye.
Up to 2 million primary oocytes are usually present in the ovaries of a newborn infant. Most of these oocytes regress during childhood so that by puberty, no more than 40,000 remain. Of these, only approximately 400 mature into secondary oocytes and are expelled at ovulation.
Comparison of Male and Female Gametes
Compared with sperms, the oocytes are massive, immotile (see Fig. 2-4B and C), and have an abundance of cytoplasm. In terms of sex chromosome constitution, there are two kinds of normal sperms (see Fig. 2-5): 22 autosomes plus either an X sex chromosome (i.e., 23, X) or a Y sex chromosome (23, Y). There is only one kind of normal secondary oocyte: 22 autosomes plus an X sex chromosome (i.e., 23, X). The difference in sex chromosome complement forms the basis of primary sex determination.
Abnormal Gametogenesis
During gametogenesis, homologous chromosomes sometimes fail to separate—known as nondisjunction—and as a result, some gametes have 24 chromosomes and others only 22 (Fig. 2-7). If a gamete with 24 chromosomes unites with a normal one with 23 chromosomes, a zygote with 47 chromosomes results, as occurs in infants with Down syndrome (see Fig. 19-4). This condition is called trisomy 21 because of the presence of three representatives of a particular chromosome instead of the usual two. If a gamete with only 22 chromosomes unites with a normal gamete, a zygote with 45 chromosomes results. This condition is known as monosomy because only one representative of the particular chromosomal pair is present. Embryos with monosomy usually die.
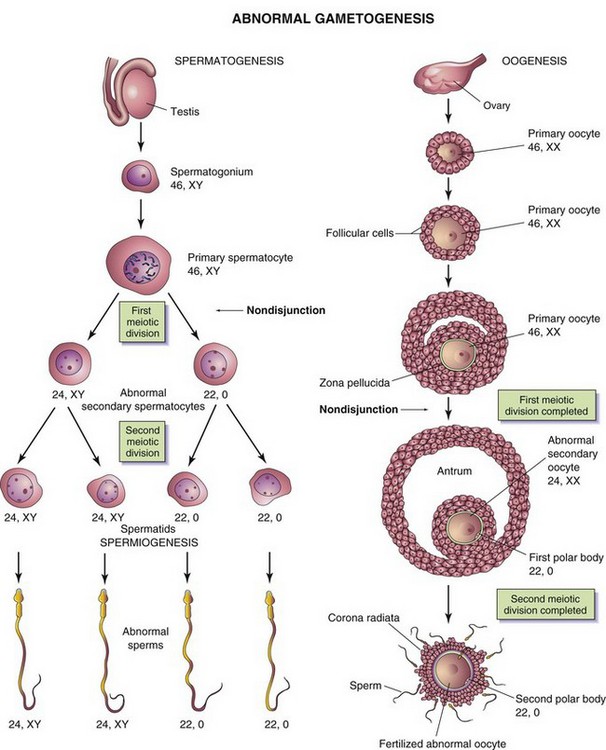
Figure 2–7 Abnormal gametogenesis. The illustrations show how nondisjunction, an error in cell division, results in an abnormal chromosome distribution in gametes. Although nondisjunction of sex chromosomes is illustrated, a similar defect may occur during the division of autosomes (any chromosomes other than sex chromosomes). When nondisjunction occurs during the first meiotic division of spermatogenesis, one secondary spermatocyte contains 22 autosomes plus an X and a Y chromosome, whereas the other one contains 22 autosomes and no sex chromosome. Similarly, nondisjunction during oogenesis may give rise to an oocyte with 22 autosomes and two X chromosomes (as shown) or one with 22 autosomes and no sex chromosome.
Female Reproductive Cycles
At menarche (the time of the first menstrual period), females undergo monthly reproductive cycles regulated by the hypothalamus, pituitary gland, and ovaries (Fig. 2-8). These cycles prepare the reproductive system for pregnancy. Gonadotropin-releasing hormone is synthesized by neurosecretory cells in the hypothalamus. It stimulates the release of two hormones (gonadotropins) that are produced by the anterior pituitary and act on the ovaries:
• Follicle-stimulating hormone (FSH) stimulates the development of the ovarian follicles and the production of estrogen by the follicular cells.
• Luteinizing hormone (LH) serves as the “trigger” for ovulation and stimulates the follicular cells and corpus luteum to produce progesterone.
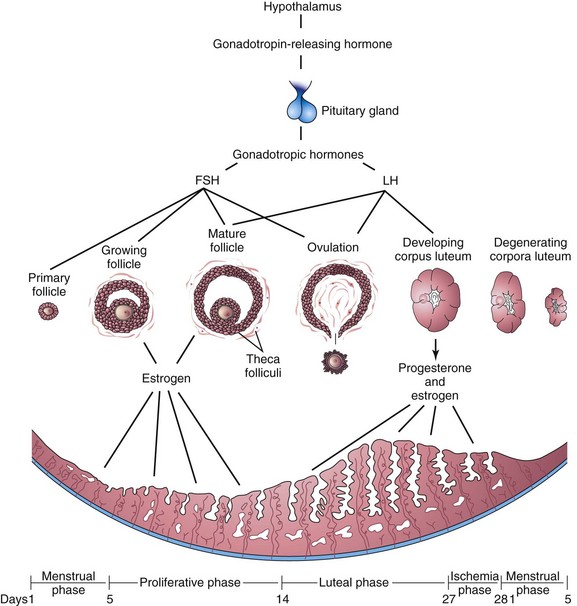
Figure 2–8 Illustrations of the interrelationships among the hypothalamus, pituitary gland, ovaries, and endometrium. One complete menstrual cycle and the beginning of another are shown.
These two ovarian hormones also produce growth of the endometrium.
Ovarian Cycle
Follicle-stimulating hormone (FSH) and luteinizing hormone (LH) produce cyclic changes in the ovaries (development of the ovarian follicles, ovulation, and formation of the corpus luteum), collectively known as the ovarian cycle. During each cycle, FSH promotes growth of several primary follicles (Figs. 2-8 and 2-9); however, only one of them usually develops into a mature follicle and ruptures, expelling its oocyte (Fig. 2-10).
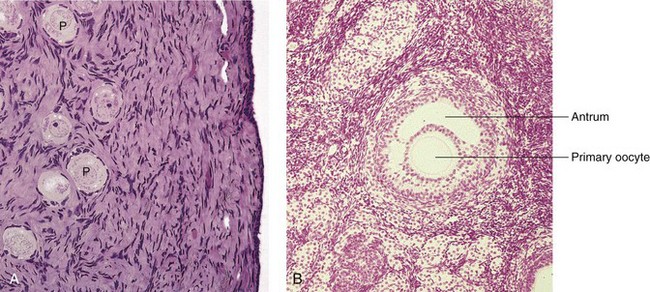
Figure 2–9 Photomicrographs of sections from adult human ovaries. A, Light micrograph of the ovarian cortex demonstrating primordial follicles (P), which are primary oocytes surrounded by follicular cells (×270). B, Light micrograph of a secondary follicle. Observe the primary oocyte and antrum containing the follicular fluid (×132).
(From Gartner LP, Hiatt JL: Color Textbook of Histology, 2nd ed. Philadelphia, WB Saunders, 2001.)
Follicular Development
Development of an ovarian follicle (Figs. 2-8 and 2-9) is characterized by:
• Growth and differentiation of a primary oocyte
• Proliferation of follicular cells
• Development of a connective tissue capsule surrounding the follicle—the theca folliculi. Thecal cells are believed to produce an angiogenic factor that promotes growth of blood vessels that provide nutritive support for follicular development.
Ovulation
The follicular cells divide actively, producing a stratified layer around the oocyte (see Fig. 2-9A and B). Subsequently, fluid-filled spaces appear around the follicular cells, which coalesce to form a single cavity, the antrum, containing follicular fluid. When the antrum forms, the ovarian follicle is called a secondary follicle (see Fig. 2-9B). The primary oocyte is surrounded by follicular cells, called the cumulus oophorus, which projects into the enlarged antrum. The follicle continues to enlarge and soon forms a bulge on the surface of the ovary (Fig. 2-10A). A small, oval, avascular spot, the stigma, soon appears on this bulge. Before ovulation, the secondary oocyte and some cells of the cumulus oophorus detach from the interior of the distended follicle (see Fig. 2-10B).
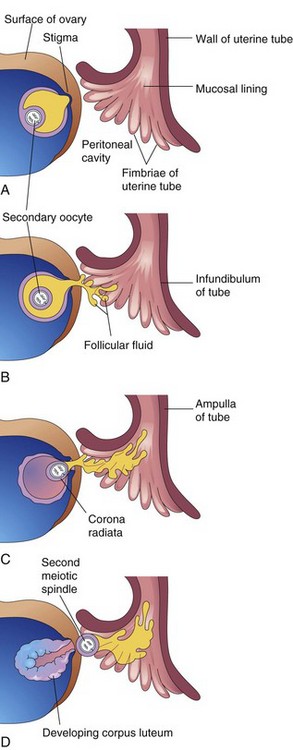
Figure 2–10 Diagrams (A–D) illustrating ovulation. When the stigma ruptures, the secondary oocyte is expelled from the ovarian follicle with the follicular fluid. After ovulation, the wall of the follicle collapses.
Ovulation follows within 24 hours of a surge of LH production that appears to be the result of signaling molecules from the granulosa cells. This surge, elicited by the high estrogen level in blood (Fig. 2-11), appears to cause the stigma to rupture, expelling the secondary oocyte along with the follicular fluid (see Fig. 2-10D). Plasmins and matrix metalloproteinases also appear to have some control over stigma rupture.
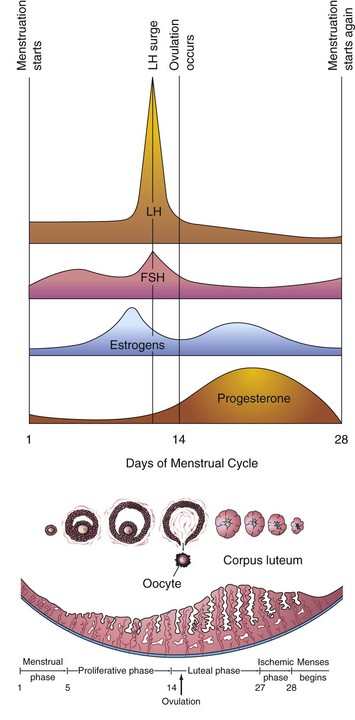
Figure 2–11 Blood levels of various hormones during the menstrual cycle. Follicle-stimulating hormone (FSH) stimulates the ovarian follicles to develop and produce estrogens. The level of estrogens rises to a peak just before the luteinizing hormone (LH) surge induces ovulation. Ovulation normally occurs within 24 hours after the LH surge. If fertilization does not occur, the blood levels of circulating estrogens and progesterone fall. This hormone withdrawal causes the endometrium to regress and menstruation to start again.
The expelled secondary oocyte is surrounded by the zona pellucida, an acellular glycoprotein coat, and one or more layers of follicular cells, which are radially arranged to form the corona radiata (see Fig. 2-4C) and the cumulus oophorus.
Mittelschmerz and Ovulation
A variable amount of abdominal pain, called mittelschmerz, accompanies ovulation in some women. Mittelschmerz may be used as a sign of ovulation; however, there are better indicators, such as the slight drop in basal body temperature, followed by a sustained rise after ovulation.
Anovulation and Hormones
Some women do not ovulate because of an inadequate release of gonadotropins. In some of these women, ovulation can be induced by the administration of gonadotropins or an ovulatory agent, resulting in maturation of several ovarian follicles and multiple ovulations. The incidence of multiple pregnancy increases when ovulation is induced.
Anovulatory Menstrual Cycles
In anovulatory cycles, the endometrial changes are minimal; the proliferative endometrium develops as usual, but no ovulation occurs and no corpus luteum forms. Consequently, the endometrium does not progress to the luteal phase; it remains in the proliferative phase until menstruation begins. The estrogen in oral contraceptives, with or without progesterone, suppresses ovulation by acting on the hypothalamus and pituitary gland, inhibiting secretion of gonadotropin-releasing hormone, follicle-stimulating hormone, and luteinizing hormone.
Corpus Luteum
Shortly after ovulation, the ovarian follicle collapses (see Fig. 2-10D). Under the influence of LH, the walls of the follicle develop into a glandular structure, the corpus luteum, which secretes primarily progesterone, but also some estrogen.
If the oocyte is fertilized, the corpus luteum enlarges to form a corpus luteum of pregnancy and increases its hormone production. Degeneration of the corpus luteum is prevented by human chorionic gonadotropin (hCG) (see Chapter 4).
If the oocyte is not fertilized, the corpus luteum degenerates 10 to 12 days after ovulation. It is then called a corpus luteum of menstruation. The degenerated corpus luteum is subsequently transformed into white scar tissue in the ovary, forming the corpus albicans.
Menstrual Cycle
The menstrual cycle is the period during which the oocyte matures, is ovulated, and enters the uterine tube (Fig. 2-11). Estrogen and progesterone produced by the ovarian follicles and the corpus luteum cause cyclic changes in the endometrium of the uterus. These monthly changes in the uterine lining constitute the menstrual cycle. The average menstrual cycle is 28 days (ranging from 23 to 35 days). Day 1 of the cycle corresponds to the day on which menstruation begins.
Phases of the Menstrual Cycle
The menstrual cycle is divided into three main phases for descriptive purposes only (see Fig. 2-11). In actuality, the menstrual cycle is a continuous process; each phase gradually passes into the next one. The reproductive cycles normally continue until the permanent cessation of the menses—menopause—which usually occurs between the ages of 48 and 55 years.
Menstrual Phase
The first day of menstruation is the beginning of the menstrual phase. The functional layer of the uterine wall is sloughed off and discarded with the menstrual flow, which usually lasts 4 to 5 days. The menstrual flow, or menses, discharged through the vagina, consists of varying amounts of blood combined with small pieces of endometrial tissue. After menstruation, the eroded endometrium is thin (see Fig. 2-11).
Proliferative Phase
The proliferative phase, lasting approximately 9 days, coincides with growth of the ovarian follicles and is controlled by estrogen secreted by the follicles. There is a two- to three-fold increase in the thickness of the endometrium during this time. Early during this phase, the surface epithelium of the endometrium regenerates. The glands increase in number and length, and the spiral arteries elongate (see Fig. 2-2C).
Luteal Phase
The luteal (secretory) phase, lasting approximately 13 days, coincides with the formation, function, and growth of the corpus luteum. The progesterone produced by the corpus luteum stimulates the glandular epithelium to secrete a glycogen-rich, mucoid material. The glands become wide, tortuous, and saccular. The endometrium thickens because of the influence of progesterone and estrogen from the corpus luteum and the increase in fluid in the connective tissue.
If fertilization does not occur:
• The corpus luteum degenerates.
• Estrogen and progesterone levels decrease, and the endometrium enters an ischemic phase.
Ischemia (reduced blood supply) of the spiral arteries occurs by constriction resulting from the decrease in the secretion of progesterone. Hormone withdrawal also results in the stoppage of glandular secretions, a loss of interstitial fluid, and a marked shrinking of the endometrium. As the spiral arteries constrict for longer periods, venous stasis and patchy ischemic necrosis (death) in the superficial tissues occur. Eventually, rupture of vessel walls follows, and blood seeps into the surrounding connective tissue. Small pools of blood form and break through the endometrial surface, resulting in bleeding into the uterus and vagina.
As small pieces of the endometrium detach and pass into the uterine cavity, the torn ends of the spiral arteries bleed into the uterine cavity, resulting in an accumulated loss of 20 to 80 mL of blood. Over 3 to 5 days, the entire compact layer and most of the spongy layer of the endometrium are discarded.
• Cleavage of the zygote and formation of the blastocyst occur.
• The blastocyst begins to implant on approximately the sixth day of the luteal phase.
• hCG maintains secretion of estrogens and progesterone by the corpus luteum.
• The luteal phase continues and menstruation does not occur.
During pregnancy, the menstrual cycles cease and the endometrium passes into a pregnancy phase. With the termination of pregnancy, the ovarian and menstrual cycles resume after a variable amount of time.
Transportation of Gametes
Oocyte Transport
During ovulation, the fimbriated (fringed) end of the uterine tube comes in close proximity to the ovary (see Fig. 2-10). The fingerlike processes of the tube, the fimbriae, move back and forth over the ovary. The sweeping action of the fimbriae and the fluid currents produced by them “sweep” the secondary oocyte into the funnel-shaped infundibulum of the tube (see Fig. 2-2B). The oocyte then passes into the ampulla of the tube, primarily as a result of waves of peristalsis—movements of the wall of the tube characterized by alternate contraction and relaxation.
Sperm Transport
During ejaculation, sperms are rapidly transported, from their storage site in the epididymis (see Fig. 2-1B) to the urethra by peristaltic contractions of the ductus deferens. Secretions from the seminal glands, prostate, and bulbourethral glands and the sperms form the semen (ejaculate). The number of sperms ejaculated ranges from 200 to 600 million. The sperms pass slowly through the cervical canal by movements of their tails. Vesiculase, an enzyme produced by the seminal glands, coagulates some of the semen and forms a vaginal plug that may prevent backflow of semen into the vagina. At the time of ovulation, the amount of cervical mucus increases and becomes less viscid, making it more favorable for sperm transport. Prostaglandins in the semen stimulate uterine motility and help to move the sperms through the uterus to the site of fertilization in the uterine tube.
The sperms move 2 to 3 mm per minute. They move slowly in the acid environment of the vagina, but more rapidly in the alkaline environment of the uterus. Approximately 200 sperms reach the fertilization site in the uterine tube.
Sperm Counts
Semen analysis is an important part of evaluating patients for infertility. Sperms account for less than 5% of the volume of semen. The remainder of the ejaculate consists of the secretions of the seminal glands (60%), prostate (30%), and bulbourethral glands (5%). The ejaculate of normal males usually contains more than 100 million sperms per milliliter of semen. Although there is much variation in individual cases, men whose semen contains a minimum of 20 million sperms per milliliter, or 50 million in the total specimen, are probably fertile. A man with less than 10 million sperms per milliliter of semen is likely to be sterile, especially when the specimen contains immotile and abnormal sperms. For potential fertility, at least 40% of sperms should be motile after 2 hours, and some should be motile after 24 hours. Male infertility may result from endocrine disorders, abnormal spermatogenesis, or obstruction of a genital duct (e.g., the ductus deferens). Male infertility is found in 30% to 50% of involuntary childless couples.
Maturation of Sperms
Freshly ejaculated sperms are unable to fertilize oocytes. They must undergo a period of conditioning—capacitation—lasting approximately 7 hours. During this period, a glycoprotein coat and seminal proteins are removed from the surface of the sperm acrosome. Capacitation and the acrosome reaction are regulated by src kinase, a tyrosine kinase. Capacitated sperms show no morphologic changes, but they exhibit increased activity. Sperms are usually capacitated in the uterus or the uterine tubes by substances (including interleukin-6) secreted by these organs.
Viability of Oocytes and Sperms
Oocytes in the uterine tube are usually fertilized within 12 hours of ovulation. In vitro observations have shown that oocytes cannot be fertilized after 24 hours, and they degenerate shortly thereafter. Most sperms probably do not survive for more than 24 hours in the female genital tract. Some sperms are captured in folds of the mucosa of the cervix and are gradually released into the cervical canal and pass through the uterus into the uterine tubes. Semen and oocytes can be stored frozen for many years to be used in assisted reproduction.
Clinically Oriented Questions
1. There have been reports of a woman who claimed that she menstruated throughout her pregnancy. How could this happen?
2. If a woman forgets to take an oral contraceptive and then takes two, is she likely to become pregnant?
3. What is coitus interruptus? Some people believe that it is an effective method of birth control. Is this true?
4. What is the difference between spermatogenesis and spermiogenesis?
5. Some say that an intrauterine device (IUD) is a contraceptive. Is this correct?