CHAPTER 84 Ankle and foot
The ankle joint (talocrural joint) is a diarthrodial articulation involving the distal tibia and fibula and the body of the talus: it is the only example in the human body of a true mortise joint. The human foot is a complex structure adapted to allow orthograde bipedal stance and locomotion and is the only part of the body which is in regular contact with the ground. There are 28 separate bones in the human foot, including the sesamoid bones of the first metatarsophalangeal joint and 31 joints, including the ankle joint.
SKIN AND SOFT TISSUE
SKIN
Vascular supply and lymphatic drainage
The skin around the ankle is supplied by anterior lateral and anterior medial malleolar arteries from the anterior tibial artery, medial malleolar branches from the posterior tibial artery, and fasciocutaneous perforators from the anterior and posterior tibial and fibular arteries. The main blood supply to the medial side of the heel is from the medial calcaneal branches of the lateral plantar artery passing through the flexor retinaculum; the skin of the lateral side of the heel is supplied by calcaneal branches of the fibular artery and the lateral tarsal artery. The arterial supply to the skin of the foot is rich and is derived from branches of the dorsalis pedis (the direct continuation of the anterior tibial artery), posterior tibial and fibular arteries. The skin covering the dorsum of the foot is supplied by the dorsalis pedis artery, and by its continuation, the first dorsal metatarsal artery, with smaller contributions from the anterior perforating branch of the fibular artery and the marginal anastomotic arteries on the medial and lateral borders of the foot. The plantar skin is supplied by perforating branches of the medial and lateral plantar arteries (the terminal branches of the posterior tibial artery). The skin of the forefoot is supplied by cutaneous branches of the common digital arteries.
Cutaneous venous drainage is via dorsal and plantar venous arches, which drain into medial and lateral marginal veins. On the plantar aspect, a superficial venous network forms an intradermal and subdermal mesh that drains to the medial and lateral marginal veins. Branches that accompany the medial and lateral plantar arteries arise from a deep venous network. Uniquely within the lower limb, venous flow in the foot is bidirectional. However, when valves are present, flow is from the plantar to the superficial dorsal system. From here, blood leaves the foot in the superficial and deep veins of the lower limb.
Superficial lymphatic drainage is via vessels that accompany the long saphenous vein medially and the short saphenous vein laterally and drain via the inguinal lymph nodes. Deep lymphatic vessels accompany the dorsalis pedis, posterior tibial and fibular arteries and pass via the popliteal lymph nodes.
Cutaneous innervation
The skin covering the ankle and foot is supplied by the fourth and fifth lumbar and first sacral spinal nerves (see Figs 79.18, 79.20). Innervation of the dorsum of the foot is provided medially by the saphenous nerve, centrally by the superficial fibular nerve and laterally by the sural nerve; the deep fibular nerve supplies the dorsum of the first web space. Dorsal branches of the medial and lateral plantar nerves supply the nail beds. The plantar aspect of the foot is supplied by the medial and lateral plantar nerves, which arise as terminal branches of the tibial nerve. The medial plantar nerve supplies sensation to the plantar aspect of the hallux, the second, the third and the medial half of the fourth toes. The lateral plantar nerve supplies the remaining lateral aspect of the fourth and the entire fifth toe. The heel is innervated by calcaneal branches of the tibial nerve. Injury to any of these nerves can lead to painful neuromata and loss of protective sensation. The sural nerve is especially prone to neuroma formation.
SOFT TISSUES
The tendons that cross the ankle joint are all deflected to some degree from a straight course, and must therefore be held down by retinacula and enclosed in synovial sheaths.
Retinacula at the ankle
In the vicinity of the ankle joint, the tendons of the muscles of the leg are bound down by localized, band-shaped thickenings of the deep fascia termed retinacula which collectively serve to prevent bowstringing of the underlying tendons. There are superior and inferior extensor retinacula, superior and inferior fibular retinacula, and a flexor retinaculum.
Extensor retinacula
Superior extensor retinaculum
The superior extensor retinaculum binds down the tendons of tibialis anterior, extensor hallucis longus, extensor digitorum longus and fibularis tertius immediately proximal to the anterior aspect of the talocrural joint (see Fig. 83.7; Fig. 84.1). The anterior tibial vessels and deep fibular nerve pass deep to the retinaculum and the superficial fibular nerve passes superficially. The retinaculum is attached laterally to the distal end of the anterior border of the fibula and medially to the anterior border of the tibia. Its proximal border is continuous with the fascia cruris, and dense connective tissue connects its distal border to the inferior extensor retinaculum. The tendon of tibialis anterior is the only extensor tendon that possesses a synovial sheath at the level of the superior extensor retinaculum.
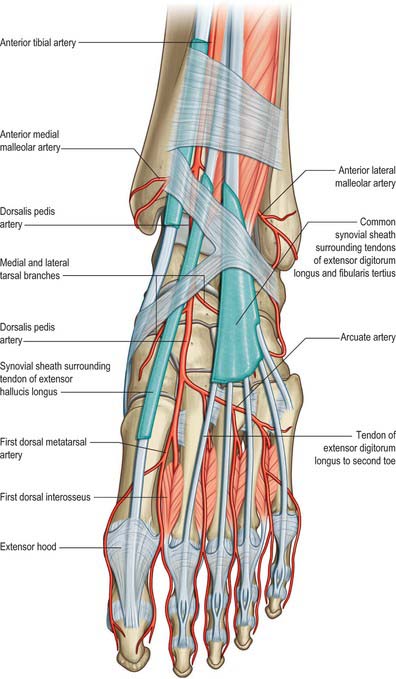
Inferior extensor retinaculum
The inferior extensor retinaculum is a Y-shaped band lying anterior to the talocrural joint (see Fig. 83.7; Fig. 84.2A,B). The stem of the Y is at the lateral end, where it is attached to the upper surface of the calcaneus, in front of the sulcus calcanei. The band passes medially, forming a strong loop around the tendons of fibularis tertius and extensor digitorum longus (Fig. 84.2A). From the deep surface of the loop, a band passes laterally behind the interosseous talocalcaneal ligament and the cervical ligament and is attached to the sulcus calcanei. At the medial end of the loop, two diverging limbs extend medially to complete the ‘Y’ shape of the retinaculum. The proximal of the two limbs consists of two layers. The deep layer passes deep to the tendons of extensor hallucis longus and tibialis anterior, but superficial to the anterior tibial vessels and deep fibular nerve, to reach the medial malleolus. The superficial layer crosses superficial to the tendon of extensor hallucis longus and then adheres firmly to the deep one; in some cases it continues superficial to the tendon of tibialis anterior, before blending with the deep layer. The distal limb extends downwards and medially and blends with the plantar aponeurosis. It is superficial to the tendons of extensor hallucis longus and tibialis anterior, the dorsalis pedis artery and the terminal branches of the deep fibular nerve.
Flexor retinaculum
The flexor retinaculum is attached anteriorly to the tip of the medial malleolus, distal to which it is continuous with the deep fascia on the dorsum of the foot (Fig. 84.2B). From its malleolar attachment it extends posteroinferiorly to the medial process of the calcaneus and the plantar aponeurosis. Proximally, there is no clear demarcation between its border and the deep fascia of the lower leg, especially the deep transverse layer of the deep fascia. Distally, its border is continuous with the plantar aponeurosis, and many fibres of abductor hallucis are attached to it. The flexor retinaculum converts grooves on the tibia and calcaneus into canals for the tendons, and bridges over the posterior tibial vessels and tibial nerve. As these structures enter the sole they are, from medial to lateral, the tendons of tibialis posterior and flexor digitorum longus, the posterior tibial vessels, the tibial nerve and the tendon of flexor hallucis longus (see Fig. 84.14).
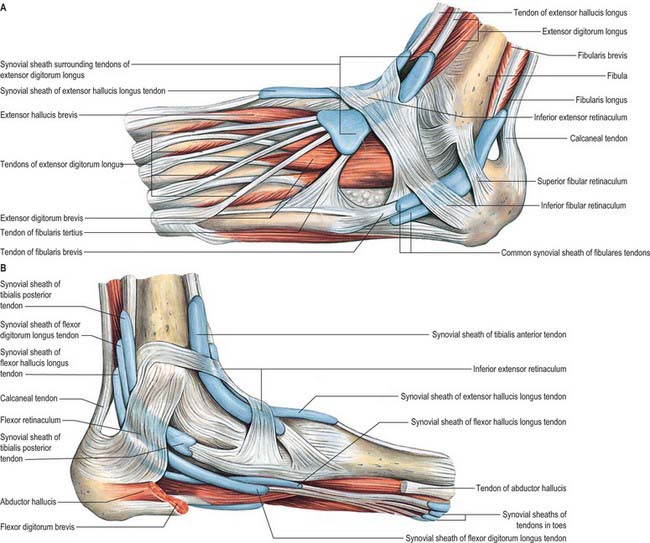
Fibular retinacula
The fibular retinacula are fibrous bands that retain the tendons of fibularis longus and brevis in position as these tendons cross the lateral aspect of the ankle region (Fig. 84.2A).
Superior fibular retinaculum
The superior fibular retinaculum is a short band which extends from the back of the lateral malleolus to the deep transverse fascia of the leg and the lateral surface of the calcaneus. Damage to the retinaculum can lead to instability of the fibular tendons.
Inferior fibular retinaculum
The inferior fibular retinaculum is continuous in front with the inferior extensor retinaculum, and is attached posteriorly to the lateral surface of the calcaneus. Some of its fibres are fused with the periosteum on the fibular trochlea (peroneal tubercle) of the calcaneus, forming a septum between the tendons of fibularis longus and brevis.
Synovial sheaths at the ankle
Anterior to the ankle, the sheath for tibialis anterior extends from the proximal margin of the superior extensor retinaculum to the interval between the diverging limbs of the inferior retinaculum (Figs 84.1, 84.2A,B). A common sheath encloses the tendons of extensor digitorum longus and fibularis tertius, starting just above the level of the malleoli, and reaching to the level of the base of the fifth metatarsal bone (Figs 84.1, 84.2A). The sheath for extensor hallucis longus starts at a level just distal to that for extensor digitorum longus and extends as far as the base of the first metatarsal bone (Figs 84.1, 84.2A,B).
Posteromedial to the ankle, the sheath for tibialis posterior starts approximately 4 cm above the medial malleolus and ends just proximal to the attachment of the tendon to the tuberosity of the navicular (Fig. 84.2B). The sheath for flexor hallucis longus starts at the level of the medial malleolus, and extends distally as far as the base of the first metatarsal bone (Fig. 84.2B). Occasionally, as a result of overuse, particularly in ballet dancers where balance on the tips of the toes en pointe involves sustained extreme plantar flexion of the ankle and first toe in the weight-bearing position, a fibrous nodule may develop in the tendon, just proximal to the tendon sheath. This may result in the thickened tendon being caught intermittently in the sheath, causing pain and ‘triggering’ of the great toe, a condition referred to as hallux saltans. Surgical opening of the sheath may be required. In athletes, the muscle belly of flexor hallucis longus may be abnormally large and may extend more distally than usual; it can also catch at the opening of the sheath. The sheath for flexor digitorum longus starts slightly above the level of the medial malleolus and ends at the level of the navicular (Fig. 84.2B).
Posterolateral to the ankle, the tendons of fibularis longus and brevis are enclosed in a sheath that is single proximally but double distally (Fig. 84.2A). From the tip of the lateral malleolus it extends for about 4 cm both proximally and distally.
Plantar fascia
The plantar fascia or aponeurosis is composed of densely compacted collagen fibres orientated mainly longitudinally, but also transversely (Fig. 84.3). Its medial and lateral borders overlie the intrinsic muscles of the hallux and fifth toe respectively, while its dense central part overlies the long and short flexors of the digits.
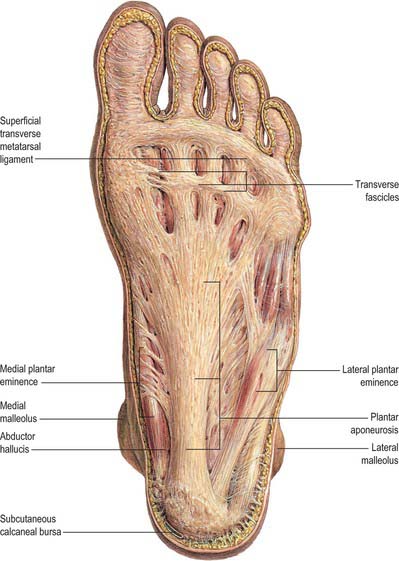
The central part is the strongest and thickest. The fascia is narrow posteriorly, where it is attached to the medial process of the calcaneal tuberosity proximal to flexor digitorum brevis, and traced distally it becomes broader and somewhat thinner. Just proximal to the level of the metatarsal heads it divides into five bands, one for each toe. As these five digital bands diverge below the metatarsal shafts, they are united by transverse fibres (Fig. 84.3). Proximal, plantar and a little distal to the metatarsal heads and the metatarsophalangeal joints, the superficial stratum of each of the five bands is connected to the dermis by skin ligaments (retinacula cutis). These ligaments reach the skin of the ball of the foot proximal to, and in the floors of, the furrows that separate the toes from the sole: Dupuytren’s disease may involve these ligaments resulting in contractures of the affected digits. The deep stratum of each digital band of the aponeurosis yields two septa that flank the digital flexor tendons and separate them from the lumbricals and the digital vessels and nerves. These septa pass deeply to fuse with the interosseous fascia, the deep transverse metatarsal ligaments (which run between the heads of adjacent metatarsals), the plantar ligaments of the metatarsophalangeal joints, and the periosteum and fibrous flexor sheaths at the base of each proximal phalanx. Pads of fat develop in the webs between the metatarsal heads and the bases of the proximal phalanges; they cushion the digital nerves and vessels from adjoining tendinous structures and extraneous plantar pressures. Just distal to the metatarsal heads, a plantar interdigital ligament (superficial transverse metatarsal ligament) blends progressively with the deep aspect of the superficial stratum of the plantar aponeurosis where it enters the toes (Fig. 84.3). The central part of the plantar aponeurosis thus provides an intermediary structure between the skin and the osteoligamentous framework of the foot via numerous cutaneous retinacula and deep septa that extend to the metatarsals and phalanges. The central part is also continuous with the medial and lateral parts: at the junctions, two intermuscular septa, medial and lateral, extend in oblique vertical planes between the medial, intermediate and lateral groups of plantar muscles to reach bone. Thinner horizontal intermuscular septa, derived from the vertical intermuscular septa, pass between the muscle layers.
The lateral part of the plantar aponeurosis, which covers abductor digiti minimi, is thin distally and thick proximally, where it forms a strong band, sometimes containing muscle fibres, between the lateral process of the calcaneal tuberosity and the base of the fifth metatarsal bone. It is continuous medially with the central part of the aponeurosis, and with the fascia on the dorsum of the foot around its lateral border. The medial part of the plantar aponeurosis, which covers abductor hallucis, is thin. It is continuous proximally with the flexor retinaculum, medially with the fascia dorsalis pedis, and laterally with the central part of the plantar aponeurosis.
Fascial compartments of the foot
There are four main compartments of the plantar aspect of the foot (Jones 1949) (Fig. 84.4). The medial compartment contains abductor hallucis and flexor hallucis brevis, and is bounded inferiorly and medially by the medial part of the plantar aponeurosis and its medial extension, laterally by an intermuscular septum, and dorsally by the first metatarsal. The central compartment contains flexor digitorum brevis, the lumbricals, flexor accessorius and adductor hallucis, and is bounded by the plantar aponeurosis inferiorly, the osseofascial tarsometatarsal structures dorsally and intermuscular septa medially and laterally. The lateral compartment contains abductor digiti minimi and flexor digiti minimi brevis, and its boundaries are the fifth metatarsal dorsally, the plantar aponeurosis inferiorly and laterally, and an intermuscular septum medially. The interosseous compartment contains the seven interossei and its boundaries are the interosseous fascia and the metatarsals.
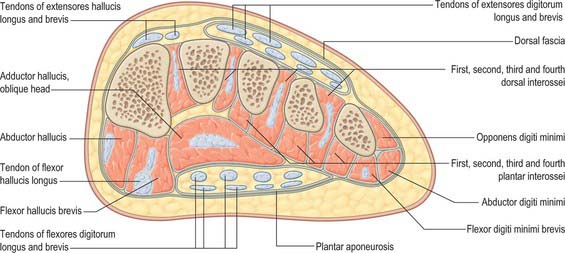
The dorsal aspect of the foot effectively contains a single compartment which is occupied by the extensor tendons and extensor digitorum brevis, and which is roofed by the deep dorsal fascia (see below).
Lateral intermuscular septum
The lateral intermuscular septum is incomplete, especially at its proximal end. Distally, its deep attachments are to the fibrous sheath of fibularis longus and to the fifth metatarsal bone.
Medial intermuscular septum
The medial intermuscular septum is incomplete and divides into three bands, proximal, intermediate and distal, each of which displays lateral and medial divisions as it approaches its deep attachment. The proximal band is attached laterally to the cuboid and blends medially with the tendon of tibialis posterior. The middle band is attached laterally to the cuboid and the long plantar ligament and medially to the medial cuneiform bone. The distal band divides to enclose the tendon of flexor hallucis longus and is attached to the fascia over flexor hallucis brevis.
Deep dorsal fascia
The deep fascia on the dorsum of the foot (fascia dorsalis pedis) is a thin layer, continuous above with the inferior extensor retinaculum; it covers the dorsal extensor tendons and extensor digitorum brevis.
Compartment syndrome in the foot
A compartment syndrome results from an increase in intracompartmental pressure sufficient to impair venous outflow from that compartment. As blood enters at arterial pressure, the compartment pressure increases further until it exceeds arterial pressure, at which point inflow of arterial blood ceases, leading to muscle and nerve ischaemia. Failure to relieve the increased pressure in the compartment surgically results in necrosis of the soft tissues within the compartment. The most common cause of compartment syndrome in the foot is trauma, usually of high-energy (high impact) type: crush injuries, calcaneal fractures and disruption of the tarsometatarsal joints are the usual antecedents associated with compartment syndrome in the foot.
Specialized adipose tissue (heel and metatarsal pad)
The heel pad is subject to repeated high impacts and is anatomically adapted to withstand these pressures. The adult heel pad has an average thickness of 18 mm and a mean epidermal thickness of 0.64 mm (dorsal epidermal thickness averages 0.069 mm). The heel pad contains elastic adipose tissue organized as spiral fibrous septa anchored to each other, to the calcaneus and to the skin. The septa are U-shaped fat-filled columns designed to resist compressive loads and are reinforced internally with elastic diagonal and transverse fibres, which separate the fat into compartments.
In the forefoot, the subcutaneous tissue consists of fibrous lamellae arranged in a complex whorl containing adipose tissue attached via vertical fibres to the dermis superficially and the plantar aponeurosis deeply. The fat is particularly thick in the region of the metatarsophalangeal joints, which cushions the foot during the toe-off phase of gait (see below). Like the heel pad, the metatarsal fat pad is designed to withstand compressive and shearing forces. Atrophy of either may be a cause of persistent pain in the distal plantar region.
BONE
Functionally, the skeleton of the foot may be divided into tarsus, metatarsus and phalanges. With regard to nomenclature of the surfaces of the foot, the terms ‘plantar’ and ‘dorsal’ are used, to denote the inferior and superior surfaces respectively. The terms ‘proximal’ and ‘distal’ are used with the same significance as in limbs generally. Rotation of the limb buds that occurs in the early stages of the development of the limbs results in a laterally directed thumb in the hand, and a medially directed great toe in the foot.
DISTAL TIBIA
The distal end of the tibia has anterior, medial, posterior, lateral and distal surfaces, and projects inferomedially as the medial malleolus (see Figs 83.2A,B; 83.3A,B). The distal surface articulates with the talus and is wider anteriorly than posteriorly. It is concave sagittally and slightly convex transversely and continues medially into the malleolar articular surface. The medial malleolus is short and thick and has a smooth lateral surface with a crescentic facet that articulates with the medial surface of the talar body. The distal end of the tibia, including its ossification, is described in detail in Chapter 83.
No muscles are attached to the distal tibia. The interosseous ligament, the deltoid ligament, and the anterior and posterior tibiofibular ligaments are attached to the distal tibia.
DISTAL FIBULA
The distal end of the fibula or lateral malleolus projects distally and posteriorly relative to the medial malleolus (see Figs 83.2A,B; 83.3A,B). Its lateral aspect is subcutaneous, the posterior surface has a broad groove with a prominent lateral border, and the anterior surface is rough and somewhat rounded and articulates with the anteroinferior aspect of the tibia. The medial surface has a triangular articular facet, vertically convex with its apex directed distally. It articulates with the lateral talar surface. Behind the facet is a rough malleolar fossa. The distal end of the fibula, including its ossification, is described in detail in Chapter 83.
No muscles are attached to the distal fibula below the level of the interosseous ligament. The ligamentous attachments are those of the lateral ligament complex, i.e. the anterior talofibular, the calcaneofibular and the posterior talofibular ligaments. The interosseous ligament is attached on its medial aspect.
TARSUS
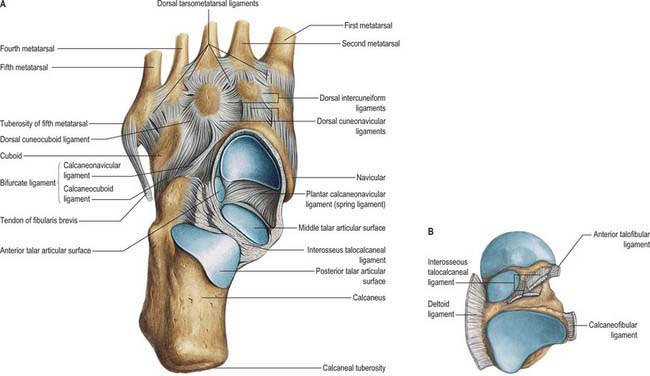
The seven tarsal bones occupy the proximal half of the foot (Figs 84.5A,B; 84.6). The tarsus and carpus are homologous, but the tarsal elements are larger, reflecting their role in supporting and distributing body weight. As in the carpus, tarsal bones are arranged in proximal and distal rows, but medially there is an additional single intermediate tarsal element, the navicular. The proximal row is made up of the talus and calcaneus; the long axis of the talus is inclined anteromedially and inferiorly, its distally directed head is medial to the calcaneus and at a higher level. The distal row contains, from medial to lateral, the medial, intermediate and lateral cuneiforms and the cuboid. Collectively these bones display an arched transverse alignment that is dorsally convex. Medially, the navicular is interposed between the head of the talus and the cuneiforms. Laterally, the calcaneus articulates with the cuboid.
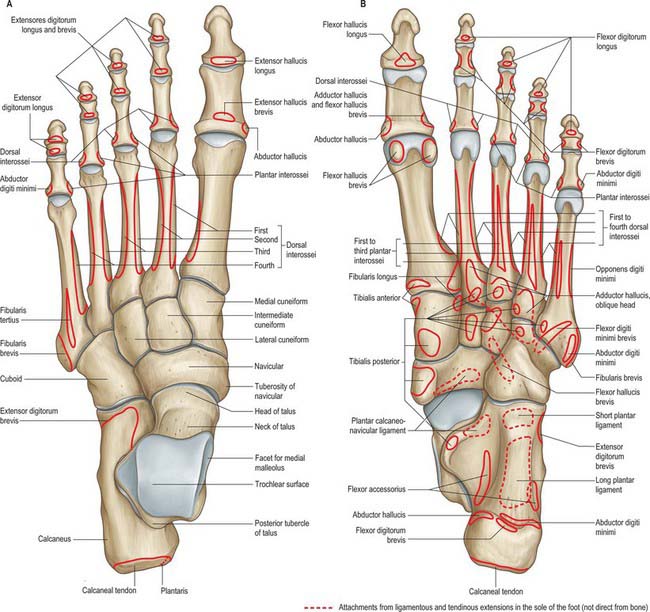
Fig. 84.5 Skeleton of the left foot, with muscle attachments. A, Dorsal aspect. B, Plantar aspect. The attachments of tibialis posterior to the metatarsals vary; those to the third and fifth are sometimes absent.
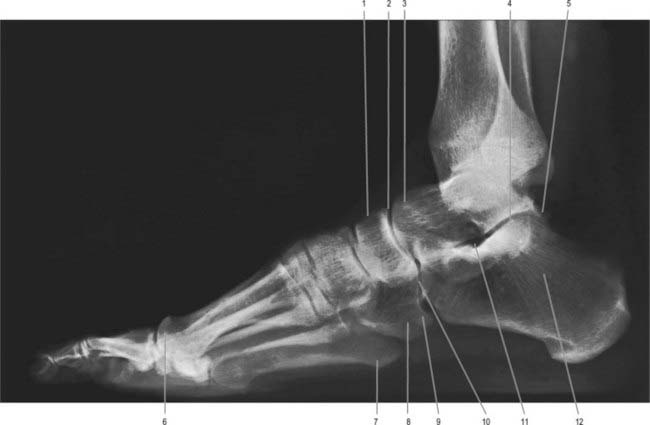
Fig. 84.6 Lateral radiograph of adult ankle and foot in full plantigrade contact with the ground, during symmetrical standing, in a man aged 44 years. 1. Navicular. 2. Talonavicular joint. 3. Head of talus. 4. Subtalar joint. 5. Os trigonum. 6. Head of first metatarsal. 7. Tuberosity on base of fifth metatarsal. 8. Cuboid. 9. Sesamoid bone in tendon of fibularis longus. 10. Calcaneocuboid joint. 11. Sinus tarsi. 12. Calcaneus (note trabecular pattern).
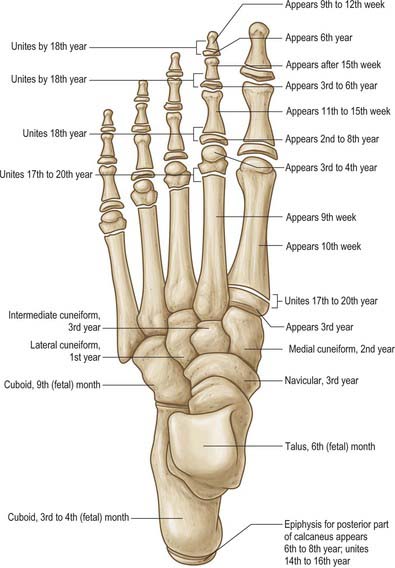
The tarsus and metatarsus are arranged to form intersecting longitudinal and transverse arches. Hence thrust and weight are not transmitted from the tibia to the ground (or vice versa) directly through the tarsus, but are distributed through the tarsal and metatarsal bones to the ends of the longitudinal arches. For the purposes of description, each tarsal bone is arbitrarily considered to be cuboidal in form, with six surfaces. The ossification sites and dates are summarized in Fig. 84.7.
Talus
The talus is the link between the foot and leg, through the ankle joint (see Figs 84.16 and 84.18).
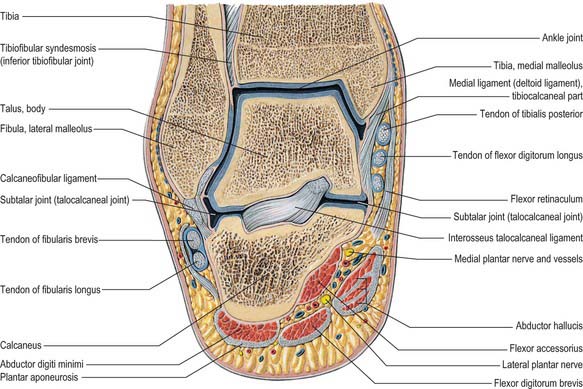
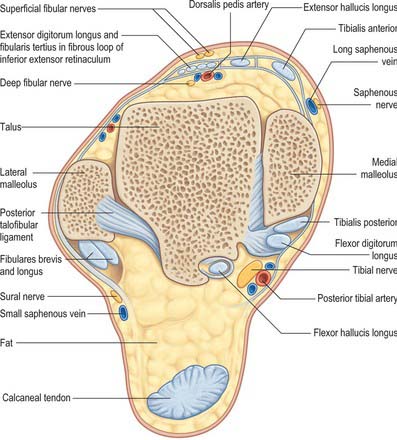
Fig. 84.16 Coronal section through the left ankle and talocalcaneal joint (seen from behind).
(From Sobotta 2006.)

Directed distally and somewhat inferomedially, the head has a distal surface which is ovoid and convex; its long axis is also inclined inferomedially to articulate with the proximal navicular surface. The plantar surface of the head has three articular areas, separated by smooth ridges. The most posterior and largest is oval, slightly convex and rests on a shelf-like medial calcaneal projection, the sustentaculum tali. Anterolateral to this and usually continuous with it, a flat articular facet rests on the anteromedial part of the dorsal (proximal) calcaneal surface; distally it continues into the navicular surface. Between the two calcaneal facets, a part of the talar head, covered with articular cartilage, is in contact with the plantar calcaneonavicular ligament, which is covered here, superiorly, by fibrocartilage (see Fig. 84.15A,B). When the foot is inverted passively, the dorsolateral aspect of the head is visible and palpable approximately 3 cm distal to the tibia; it is hidden by extensor tendons when the toes are dorsiflexed.
The neck is the narrow, medially inclined region between the head and body. Its rough surfaces are for ligaments. The medial plantar surface has a deep sulcus tali which, when the talus and calcaneus are articulated, forms a roof to the sinus tarsi, which is occupied by inter-osseous talocalcaneal and cervical ligaments.
The long axis of the neck, inclined downwards, distally and medially, makes an angle of approximately 150° with that of the body; it is smaller (130–140°) at birth, accounting in part for the inverted foot in young children. The dorsal talonavicular ligament and ankle articular capsule are attached distally to its dorsal surface. Thus the proximal part of this surface lies within the capsule of the ankle joint. The medial articular facet of the talar body and part of the trochlear surface may extend onto the neck. The anterior talofibular ligament is attached on the lateral aspect of the neck, spreading along the adjacent anterior border of the lateral surface. The interosseous talocalcaneal and cervical ligaments are attached to the inferior surface of the neck. A dorsolateral, so-called ‘squatting facet’ is commonly present on the talar neck in those individuals who habitually adopt the squatting position: it articulates with the anterior tibial margin in extreme dorsiflexion and may be double.
The body is cuboidal, covered dorsally by a trochlear surface articulating with the distal end of the tibia. It is anteroposteriorly convex, gently concave transversely, widest anteriorly and, therefore, sellar. The triangular lateral surface is smooth and vertically concave for articulation with the lateral malleolus. Superiorly, it is continuous with the trochlear surface; inferiorly its apex is a lateral process. Proximally, the medial surface is (posterosuperiorly) covered by a comma-shaped facet, which is deeper in front and articulates with the medial malleolus. Distally, this surface is rough and contains numerous vascular foramina. The small posterior surface features a rough projection termed the posterior process. The process is marked by an oblique groove between two tubercles which lodges the tendon of flexor hallucis longus. The lateral tubercle is usually larger; the medial is less prominent and immediately behind the sustentaculum tali (Fig. 84.8A). The plantar surface articulates with the middle one-third of the dorsal calcaneal surface by an oval concave facet, its long axis directed distolaterally at an angle of approximately 45° with the median plane. The medial edge of the trochlear surface is straight, but its lateral edge inclines medially in its posterior part and is often broadened into a small elongated triangular area which is in contact with the posterior tibiofibular ligament in dorsiflexion.
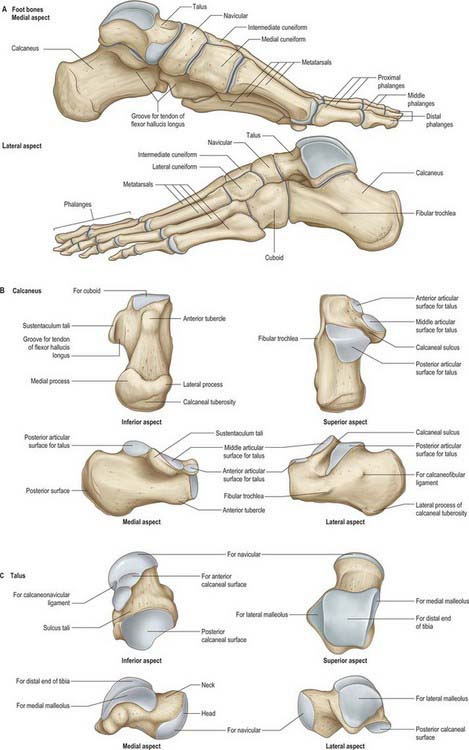
Fig. 84.8 Skeleton of the foot. A, Foot bones. B, Calcaneus. C, Talus.
(From Drake, Vogl, Mitchell, Tibbitts and Richardson 2008.)
The posterior talofibular ligament is attached to the lateral tubercle of the posterior process. Its attachment extends up to the groove, or depression, between the process and posterior trochlear border. The posterior talocalcaneal ligament is attached to the plantar border of the posterior process. The groove between the tubercles of the process contains the tendon of flexor hallucis longus and continues distally into the groove on the plantar aspect of the sustentaculum tali. The medial talocalcaneal ligament is attached below to the medial tubercle, whereas the most posterior superficial fibres of the deltoid ligament are attached above the tubercle. The deep fibres of the deltoid ligament are attached still higher to the rough area immediately below the comma-shaped articular facet on the medial surface (Fig. 84.8A,C).
No muscles are attached to the talus. However, many ligaments are attached to the bone, and these confer stability to the ankle, subtalar and talocalcaneonavicular joints (Fig. 84.8A–C).
The talar blood supply is rather tenuous because of the lack of muscle attachments. The first comprehensive account of talar blood supply was provided by Wildenauer in 1950. The extraosseous blood supply is via the posterior tibial, dorsalis pedis and fibular arteries (Fig. 84.9). The ‘artery of the tarsal canal’ arises from the posterior tibial artery approximately 1cm proximal to the origin of the medial and lateral plantar arteries (Fig. 84.10) and passes anteriorly between the sheaths of flexor digitorum longus and flexor hallucis longus to enter the tarsal canal in which it lies anteriorly, close to the talus. (The ‘tarsal canal’ is the term that is commonly used to describe the tunnel-shaped medial end of the sinus tarsi.) Branches from the arterial network in the tarsal canal enter the talus. The artery continues through the tarsal canal into the lateral part of the tarsal sinus, where it anastomoses with the artery of the tarsal sinus, forming a vascular sling under the talar neck. A branch of the artery of the tarsal canal known as the deltoid branch passes deep to the deltoid ligament and supplies part of the medial aspect of the talar body. Sometimes it arises from the posterior tibial artery; rarely, it arises from the medial plantar artery. In talar fractures it may be the only remaining arterial supply to the talus to maintain the viability of the talar body. The dorsalis pedis artery supplies branches to the superior aspect of the talar neck and also gives off the artery of the tarsal sinus. This large vessel is always present and anastomoses with the artery of the tarsal canal. The artery of the tarsal sinus receives a contribution from the anterior perforating branch of the fibular artery and supplies direct branches to the talus. The fibular artery provides small branches which form a plexus of vessels posteriorly with branches of the posterior tibial artery, however, the contribution that the fibular artery makes to the talar blood supply is thought to be insignificant.
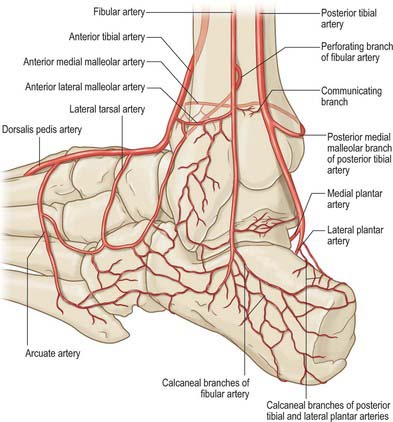
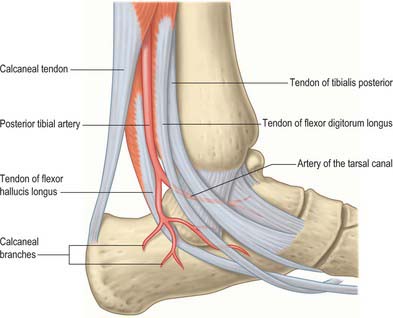
The intraosseous blood supply of the talar head comes medially from branches of the dorsalis pedis and laterally via vessels that arise from the anastomosis between the arteries of the tarsal canal and tarsal sinus. The middle one-third of the talar body, other than its most superior aspect, and the lateral one-third, other than its posterior aspect, are supplied mainly by the anastomotic arcade in the tarsal canal. The medial one-third of the talar body is supplied by the deltoid branch of the artery of the tarsal canal.
The talus is innervated by branches from the deep fibular, posterior tibial, saphenous and sural nerves.
A single ossification centre appears prenatally at 6 months (Fig. 84.7). The posterior process (Stieda’s process) is a separate bone in 5% of individuals and arises from a separate ossification centre, which appears between 8 and 11 years. In athletes and dancers, it may be susceptible to impingement against the posterior tibia, resulting in pain and sometimes requires surgical removal. Another accessory bone (although rare) is the os supratalare, which lies on the dorsal aspect of the talus; it rarely measures more than 4 mm in length.
The talus has no muscular attachments and 70% of its surface is covered by articular cartilage: displaced talar neck fractures, where the blood supply to the talar body is interrupted, can therefore result in avascular necrosis and non-union. In general, these complications do not occur in undisplaced fractures. If a subchondral lucency is seen in the talar dome on radiographs at 8 weeks after fracture of the talar neck (Hawkin’s sign), it may be assumed that vascularity to the talar body is intact and that the fracture is likely to heal satisfactorily.
Calcaneus
The calcaneus is the largest tarsal bone and projects posterior to the tibia and fibula as a short lever for muscles of the calf attached to its posterior surface. It is irregularly cuboidal, its long axis being inclined distally upwards and laterally (Fig 84.8A,B). The superior or proximal surface is divisible into three areas. The posterior one-third is rough and concavo-convex; the convexity is transverse and supports fibroadipose tissue (Kager’s fat pad) between the calcaneal tendon and ankle joint. The middle one-third carries the posterior talar facet, which is oval and convex anteroposteriorly. The anterior one-third is partly articular; distal (anterior) to the posterior articular facet, a rough depression, the sulcus calcanei, narrows into a groove on the medial side and completes the sinus tarsi with the talus. (The sinus tarsi is a conical hollow bounded by the talus medially, superiorly and laterally, with the superior surface of the calcaneus below. Its medial end is narrow and tunnel-shaped, and is often referred to as the tarsal canal.) Distal and medial to this groove, an elongated articular area covers the sustentaculum tali and extends distolaterally on the body of the bone. This facet is often divided into middle and anterior talar facets by a non-articular interval at the anterior limit of the sustentaculum tali (the incidence of this subdivision varies with sex, race and occupation). Rarely, all three facets on the upper surface of the calcaneus are fused into one irregular area. A detailed analysis of patterns of anterior talar articular facets in a series of 401 Indian calcanei revealed four types. Type I (67%) showed one continuous facet on the sustentaculum extending to the distomedial calcaneal corner; type II (26%) presented two facets, one sustentacular and one distal calcaneal; type III (5%) possessed only a single sustentacular facet; and type IV (2%) showed confluent anterior and posterior facets.
The anterior surface is the smallest, and is an obliquely set concavo-convex articular facet for the cuboid. The posterior surface is divided into three regions: a smooth proximal (superior) area separated from the calcaneal tendon by a bursa and adipose tissue; a middle area, which is the largest, limited above by a groove and below by a rough ridge for the calcaneal tendon; a distal (inferior) area inclined downwards and forwards, vertically striated, which is the subcutaneous weight-bearing surface.
The plantar surface is rough, especially proximally as the calcaneal tuberosity, the lateral and medial processes of which extend distally, separated by a notch. The medial process is longer and broader (Fig. 84.8B). Further distally, an anterior tubercle marks the distal limit of the attachment of the long plantar ligament.
The lateral surface is almost flat. It is proximally deeper and palpable on the lateral aspect of the heel distal to the lateral malleolus. Distally, it presents the fibular trochlea (Fig. 84.8A,B), which is exceedingly variable in size and palpable 2 cm distal to the lateral malleolus when well developed. It bears an oblique groove for the tendon of fibularis longus and a shallower proximal groove for the tendon of fibularis brevis. About 1 cm or more behind and above the fibular trochlea, a second elevation may exist for attachment of the calcaneofibular part of the lateral ligament.
The medial surface is vertically concave, and its concavity is accentuated by the sustentaculum tali, which projects medially from the distal part of its upper border (Fig. 84.8B). Superiorly, the process bears the middle talar facets and inferiorly a groove which is continuous with that on the talar posterior surface for the tendon of flexor hallucis longus (Fig. 84.8A,B). The medial aspect of the sustentaculum tali can be felt immediately distal to the tip of the medial malleolus; occasionally it is also grooved by the tendon of flexor digitorum longus.
Muscle and ligament attachments
The interosseous talocalcaneal and cervical ligaments and the medial root of the inferior extensor retinaculum are attached in the calcaneal sulcus. The non-articular area distal to the posterior talar facet is the site of attachment of extensor digitorum brevis (in part), the principal band of the inferior extensor retinaculum and the stem of the bifurcate ligament.
Abductor hallucis and the superficial part of the flexor retinaculum and, distally, the plantar aponeurosis and flexor digitorum brevis, are all attached to the medial process of the calcaneal tuberosity at its prominent medial margin. Abductor digiti minimi is attached to the lateral process, extending medially to the medial process. The long plantar ligament is attached to the rough region between the processes proximally, and extends to the anterior tubercle distally. The short plantar ligament is attached to the tubercle and the area distal to it. The lateral tendinous head of flexor accessorius is attached distal to the lateral process near the lateral margin of the long plantar ligament. Plantaris is attached to the posterior surface near the medial side of the calcaneal tendon. The anterior part of the lateral surface is crossed by the fibular tendons, but is largely subcutaneous. The calcaneofibular ligament is attached 1–2 cm proximal to the fibular trochlea, usually to a low, rounded elevation.
The dorsal surface of the sustentaculum tali is part of the talocalcaneonavicular joint; its plantar surface is grooved by the tendon of flexor hallucis longus and margins of the groove give attachment to the deep part of the flexor retinaculum. The plantar calcaneonavicular ligament is attached distally to the medial margin of the sustentaculum, which is narrow, rough and convex. A slip from the tendon of tibialis posterior, and superficial fibres of the deltoid ligament and medial talocalcaneal ligaments, are attached proximally. Distal to the attachment of the deltoid ligament, the tendon of flexor digitorum longus is related to the margin of the sustentaculum and may groove it. The large medial head of flexor accessorius is attached distal to the groove for flexor hallucis longus.
The calcaneus receives its arterial supply from the medial and lateral calcaneal arteries (arising from the posterior tibial and fibular arteries, respectively), fibular artery, posterior calcaneal anastomosis (formed from the posterior tibial and fibular arteries), medial and lateral plantar arteries, artery of the tarsal sinus and tarsal canal, branches of the lateral tarsal artery and perforating fibular arteries.
The calcaneus is the only tarsal bone that always has two ossification centres (Fig. 84.7). In addition to the main ossification centre, there is a scale-like posterior apophysis that covers most of the posterior, and part of the plantar, surfaces. The main centre appears prenatally in the third month, whereas the posterior apophysis appears in the sixth year in females and the eighth in males, fusing in the 14th and 16th years, respectively.
An os calcaneus secondarius, an accessory bone rather than a secondary ossification centre, occurs in 2% of individuals. When present, it is located on the dorsal beak or anterior process of the calcaneus in an interval between the anteromedial aspect of the calcaneus, the proximal ends of the cuboid and navicular, and the head of the talus. Other rare accessory bones of the calcaneus include the calcaneus accessorius in the region of the fibular trochlea, the os sustentaculi on the posterior aspect of the sustentaculum, the os subcalcis on the plantar aspect of the calcaneus slightly posterior to the origin of the plantar aponeurosis, and the os aponeurosis plantaris, which lies within the plantar aponeurosis in close proximity to the medial process of the calcaneal tuberosity.
Navicular
The navicular articulates with the talar head proximally and with the cuneiform bones distally (Figs 84.8A,B; 84.11). Its distal surface is transversely convex and divided into three facets (the medial being the largest) for articulation with the cuneiforms. The proximal surface is oval and concave and articulates with the talar head. The dorsal surface is rough and convex. The medial surface is also rough and projects proximally as a prominent tuberosity, palpable approximately 2.5 cm distal and plantar to the medial malleolus. The plantar surface, rough and concave, is separated from the tuberosity medially by a groove. The lateral surface is rough, irregular and often bears a facet for articulation with the cuboid.
The facet for articulation with the medial cuneiform is roughly triangular, its rounded apex is medial and its ‘base’, facing laterally, is often markedly curved; the articular facets for the intermediate and lateral cuneiforms are also triangular, with plantar facing apices. The facet for the lateral cuneiform may approach a wide crescent or a semicircle rather than a triangle (Fig. 84.11). Dorsal talonavicular, cuneonavicular and cubonavicular ligaments are attached to the dorsal navicular surface.
Muscle and ligament attachments
The navicular tuberosity is the main attachment of tibialis posterior and a groove lateral to it transmits part of the tendon distally to the cuneiforms and middle three metatarsal bases. The plantar calcaneonavicular ligament is attached to a slight projection lateral to the groove and adjacent to the proximal surface. The calcaneonavicular part of the bifurcate ligament is attached to the rough part of the lateral surface (Fig. 84.5B).
The dorsal aspect of the navicular is supplied either from a branch of, or directly from, the dorsalis pedis artery. Its plantar aspect is supplied from the medial plantar artery, and the tuberosity is supplied by an anastomosis from the dorsalis pedis and medial plantar arteries.
The navicular ossification centre appears during the third year (Fig. 84.7). It is sometimes affected by avascular necrosis between the ages of 4 and 7 years (Köhler’s disease). An accessory navicular bone, which is considered an anatomic variant, occurs in approximately 5% of individuals. It arises from a separate ossification centre in the region of the navicular tuberosity. There are three distinct types of accessory navicular. Type I is probably a sesamoid bone within the plantar aspect of the tendon of tibialis posterior at the level of the inferior calcaneonavicular ligament. In type II, the accessory bone is separated from the body of the navicular by a synchondrosis. Type III is commonly called ‘the cornuate navicular,’ where the accessory bone is united to the navicular by a bony ridge, and may represent the possible end stage of type II. An accessory navicular may be the source of pain in athletes. Type II is the most commonly symptomatic variant: it has been suggested that the pull of the tendon of tibialis posterior, the degree of foot pronation, and the location of the accessory bone in relation to the undersurface of the navicular may produce tension, shear, and/or compression forces on the synchondrosis.
Rarely, the navicular is bipartite and it arises from two distinct centres of ossification. This can lead to premature degeneration within the talocalcaneonavicular joint (Muller–Weiss disease). Occasionally, a small bone is found within the talocalcaneonavicular joint on its dorsal aspect. Referred to as an os talonaviculare dorsale, it represents either a separate accessory bone or a fractured osteophyte of the proximal dorsal aspect of the navicular.
Cuboid
The cuboid, the most lateral bone in the distal tarsal row, lies between the calcaneus proximally and the fourth and fifth metatarsals distally (Fig. 84.11). Its dorsolateral surface is rough for the attachment of ligaments. The plantar surface is crossed distally by an oblique groove for the tendon of fibularis longus and bounded proximally by a ridge that ends laterally in the tuberosity of the cuboid, the lateral aspect of which is faceted for a sesamoid bone or cartilage that is frequently found in the tendon of fibularis longus. Proximal to its ridge, the rough plantar surface extends proximally and medially because of the obliquity of the calcaneocuboid joint, making its medial border much longer than the lateral. The lateral surface is rough; the groove for fibularis longus extends from a deep notch on its plantar edge. The medial surface, which is much more extensive and partly non-articular, bears an oval facet for articulation with the lateral cuneiform, and proximal to this another facet (sometimes absent) for articulation with the navicular: the two form a continuous surface separated by a smooth vertical ridge. The distal surface is divided vertically into a medial quadrilateral articular area for the fourth metatarsal base and a lateral triangular area, its apex lateral, for the fifth metatarsal base. The proximal surface, triangular and concavo-convex, articulates with the distal calcaneal surface; its medial plantar angle projects proximally and inferior to the distal end of the calcaneus.
Muscle (and ligament) attachments
The dorsal calcaneo-cuboid, cubonavicular, cuneocuboid and cubometatarsal ligaments are attached to the dorsal surface. Deep fibres of the long plantar ligament are attached to the proximal edge of the plantar ridge. Slips from the tendons of tibialis posterior and flexor hallucis brevis are attached to the projecting proximomedial part of the plantar surface. Interosseous, cuneocuboid and cubonavicular ligaments are attached to the rough part of the medial cuboidal surface. Proximally, the medial calcaneocuboid ligament, which is the lateral limb of the bifurcate ligament, is also attached to this surface.
The cuboid is supplied by deep branches of the medial and lateral plantar arteries and by branches from the dorsal arterial network.
The cuboid frequently begins to ossify before birth, the primary ossification centre appearing just before birth (Fig. 84.7). An os cuboides secundarium is a rare accessory bone situated on the plantar aspect of the cuboid.
Cuneiforms
The wedge-like cuneiform bones articulate with the navicular proximally and with the bases of the first to third metatarsals distally; the medial cuneiform is the largest, the intermediate the smallest. The dorsal surfaces of the intermediate and lateral cuneiforms form the base of the wedge. The wedge is reversed in the medial cuneiform, which is a prime factor in shaping the transverse arch. The proximal surfaces of all three form a concavity for the distal surface of the navicular. The medial and lateral cuneiforms project distally beyond the intermediate cuneiform and so form a recess for the second metatarsal base.
Medial cuneiform
The medial cuneiform (Figs 84.5A,B; 84.11) articulates with the navicular and first metatarsal base. It has a rough, narrow dorsal surface. The distal surface is a reniform facet for the first metatarsal base, its ‘hilum’ being lateral. The proximal surface bears a piriform facet for the navicular, which is concave vertically and dorsally narrowed. The medial surface, rough and subcutaneous, is vertically convex; its distal plantar angle carries a large impression which receives the principal attachment of the tendon of tibialis anterior (Fig. 84.5B). The lateral surface is partly non-articular; there is a smooth right-angled strip along its proximal and dorsal margins for the intermediate cuneiform. Its distal dorsal area is separated by a vertical ridge from a small, almost square, facet for articulation with the dorsal part of the medial surface of the second metatarsal base. Plantar to this, the medial cuneiform is attached to the medial side of the second metatarsal base by a strong ligament. Proximally, an interosseous intercuneiform ligament connects this surface to the intermediate cuneiform. The distal and plantar area of the surface is roughened by attachment of part of the tendon of fibularis longus (Fig. 84.5B).
The plantar surface receives a slip from the tendon of tibialis posterior, in addition to part of the insertion of the tendon of fibularis longus. The medial surface receives the attachment of most of the tendon of tibialis anterior.
The medial cuneiform is supplied via its dorsal, medial and lateral surfaces, mainly from the dorsal arterial network.
The medial cuneiform may have two separate ossification centres, which appear during the second year of life (Fig. 84.7). Very rarely, the medial cuneiform is bipartite and there is a horizontal cleavage plane between the two moieties.
The os cuneo-1 metatarsale-I plantare is a rare accessory bone that occurs on the plantar aspect of the foot at the base of the first metatarsal and articulates with the plantar base of the first metatarsal and the medial cuneiform.
Intermediate cuneiform
The intermediate cuneiform articulates proximally with the navicular and distally with the second metatarsal base (Figs 84.5A; 84.11). It has a narrow, plantar surface that receives a slip from the tendon of tibialis posterior. The distal and proximal surfaces are both triangular articular facets and articulate with the second metatarsal base and the navicular, respectively. The medial surface is partly articular: it articulates via a smooth, angled strip that is occasionally double with the medial cuneiform along its proximal and dorsal margins. The lateral surface is also partly articular: along its proximal margin a vertical strip, usually indented, abuts the lateral cuneiform. Strong interosseous ligaments connect non-articular parts of both surfaces to the adjacent cuneiforms.
The intermediate cuneiform is supplied via its dorsal, medial and lateral surfaces, mainly from the dorsal arterial network.
The ossification centre appears during the third year of life (Fig. 84.7). The os cuneo-2 metatarsale-II dorsale, a rare accessory bone, lies on the dorsal aspect of the joint between the intermediate cuneiform and the second metatarsal. It is wedge-shaped with its base orientated dorsally.
Lateral cuneiform
The lateral cuneiform is between the intermediate cuneiform and cuboid, and also articulates with the navicular and, distally, with the third metatarsal base (Figs 84.5A; 84.11). Like the intermediate cuneiform, its dorsal surface, which is rough and almost rectangular, is the base of a wedge. The plantar surface is narrow and receives a slip from tibialis posterior and sometimes part of flexor hallucis brevis. The distal surface is a triangular articular facet for the third metatarsal base. The proximal surface is rough on its plantar aspect, but its dorsal two-thirds articulate with the navicular by a triangular facet. The medial surface is partly non-articular and has a vertical strip, indented by the intermediate cuneiform, on its proximal margin; on its distal margin, a narrower strip (often two small facets) articulates with the lateral side of the second metatarsal base. The lateral surface, also partly non-articular, bears a triangular or oval proximal facet for the cuboid; a semilunar facet on its dorsal and distal margin articulates with the dorsal part of the medial side of the fourth metatarsal base. Non-articular areas of the medial and lateral surfaces receive intercuneiform and cuneocuboid ligaments, respectively, which are important in the maintenance of the transverse arch.
The plantar surface of the lateral cuneiform receives a slip from the tendon of tibialis posterior and, occasionally, part of flexor hallucis brevis.
The lateral cuneiform is supplied via its dorsal, medial and lateral surfaces, mainly from the dorsal arterial network.
Tarsal coalition
Tarsal coalition is a hereditary condition in which there is a fibrous, cartilaginous or osseous union of two or more tarsal bones, and is believed to arise as a result of a failure of segmentation of primitive mesenchyme. Harris & Beath (1948) were the first to recognize an association between tarsal coalitions and ‘peroneal (fibular) spastic flat foot’. The two most common examples are talocalcaneal and calcaneonavicular coalitions, which usually present with symptoms early in the second decade of life. They are often, but not invariably, associated with flat feet (see below). A talonavicular coalition is rare, but when present is often associated with a ‘ball and socket’ ankle joint. Surgical resection of tarsal coalitions may eradicate associated pain but seldom improves the range of movement.
METATARSALS
The five metatarsal bones lie in the distal half of the foot and connect the tarsus and phalanges. Like the metacarpals, they are miniature long bones, and have a shaft, proximal base and distal head. Except for the first and fifth, the shafts are long and slender, longitudinally convex dorsally, and concave on their plantar aspects. Prismatic in section, they taper distally. Their bases articulate with the distal tarsal row and with adjacent metatarsal bases. The line of each tarsometatarsal joint, except the first, inclines proximally and laterally, metatarsal bases being oblique relative to their shafts. The heads articulate with the proximal phalanges, each by a convex surface that passes farther on to its plantar aspect, where it ends on the summits of two eminences. The sides of the heads are flat, with a depression surmounted by a dorsal tubercle for a collateral ligament of the metatarsophalangeal joint.
On occasion, an os intermetatarseum is encountered between the medial cuneiform and the bases of the first and second metatarsal bones and represents a rare accessory bone in this region.
Individual metatarsals
First metatarsal
The first metatarsal (Fig. 84.5A,B; 84.11) is the shortest and thickest, and has a strong shaft, of marked prismatic form. The base sometimes has a lateral facet or ill-defined smooth area as a result of contact with the second metatarsal. Its large proximal surface, usually indented on the medial and lateral margins, articulates with the medial cuneiform. Its circumference is grooved for tarsometatarsal ligaments and, medially, part of the tendon of tibialis anterior is attached; its plantar angle has a rough, oval, lateral prominence for the tendon of fibularis longus. The medial head of the first dorsal interosseous is attached to the flat lateral surface of the shaft. The large head has a plantar elevation, the crista, which separates two grooved facets (of which the medial is larger), on which sesamoid bones glide.
The first metatarsal receives attachments from the tendon of tibialis anterior medially, and the tendon of fibularis longus on its plantar aspect. It gives origin to the medial head of the first dorsal interosseus on the proximal aspect of the lateral surface.
The first metatarsal is supplied by the first dorsal and first plantar metatarsal arteries and a superficial branch of the medial plantar artery, which together form a periosteal network. A nutrient artery enters the lateral surface of the mid-diaphysis. The head receives a medial, lateral and plantar supply from these arteries.
The first metatarsal has two centres of ossification, one in the shaft, the other in the base (unlike the other metatarsals, in which the secondary ossification centre is distal). They appear during the tenth week of prenatal life and the third year of life, respectively (Fig. 84.7) and fuse between the 17th and 20th years. There may be a third centre in the first metatarsal head.
Second metatarsal
The second metatarsal is the longest (Figs 84.5A,B; 84.11). Its cuneiform base bears four articular facets. The proximal one, concave and triangular, is for the intermediate cuneiform. The dorsomedial one, for the medial cuneiform, is variable in size and usually continuous with that for the intermediate cuneiform. Two lateral facets, dorsal and plantar, are separated by non-articular bone, each divided by a ridge into distal demifacets, which articulate with the third metatarsal base, and a proximal pair (sometimes continuous) for the lateral cuneiform. The areas of these facets vary, particularly the plantar facet, which may be absent. An oval pressure facet, caused by contact with the first metatarsal, may appear on the medial side of the base, plantar to that for the medial cuneiform. Because of its length, its steep inclination, and the position of its base recessed in the tarsometatarsal joint, it is at risk of stress overload; perhaps this is why it is a common site for stress fractures in athletes and an avascular phenomenon in its head (Freiberg’s infraction).
The lateral head of the first dorsal interosseous and the medial head of the second, are attached to the medial and lateral surfaces of the shaft, respectively.
The blood supply of the second, third and fourth metatarsals follows the same pattern as that described for the first metatarsal, i.e. the bones are all supplied by branches of the dorsal and plantar metatarsal arteries. The nutrient artery enters the diaphysis on its lateral side near the metatarsal base. A constant plantar vessel supplies the head.
The second metatarsal is innervated by branches of the deep fibular and branches of medial plantar nerve.
There are two centres of ossification, one in the shaft and one distally in the metatarsal head (Fig. 84.7). Ossification of the shaft starts during the ninth prenatal week and ossification of the metatarsal head starts between the third and fourth years; fusion occurs between the 17th and 20th years.
Third metatarsal
The third metatarsal (Figs 84.5A,B; 84.11) has a flat triangular base, articulating proximally with the lateral cuneiform, medially with the second metatarsal, via dorsal and plantar facets, and laterally, via a single facet, with the dorsal angle of the fourth metatarsal. The medial plantar facet is frequently absent. The third tarsometatarsal joint is relatively immobile and predisposes the third metatarsal to stress fracture.
The lateral head of the second dorsal interosseous and first plantar are attached to the medial surface of the shaft. The medial head of the third dorsal interosseous is attached to its lateral surface.
The blood supply of the third metatarsal is the same as that of the second metatarsal, described above.
There are two centres of ossification, one in the shaft and one distally in the metatarsal head (Fig. 84.7). Ossification of the shaft starts during the ninth prenatal week and ossification of the metatarsal head starts between the third and fourth years; fusion occurs between the 17th and 20th years.
Fourth metatarsal
The fourth metatarsal is smaller than the third (Figs 84.5A,B; 84.11). Its base has proximally, an oblique quadrilateral facet for articulation with the cuboid; laterally, a single facet for the fifth metatarsal; medially, an oval facet for the third metatarsal. The latter is sometimes divided by a ridge, in which case the proximal part articulates with the lateral cuneiform.
The lateral head of the third dorsal and second plantar interossei are attached to the medial surface. The medial head of the fourth dorsal interosseous is attached to the lateral surface.
The blood supply of the fourth metatarsal is the same as that of the second and third metatarsals, described above.
There are two centres of ossification, one in the shaft and one distally in the metatarsal head (Fig. 84.7). Ossification of the shaft starts during the ninth prenatal week and ossification of the metatarsal head commences between the third and fourth years; fusion occurs between the 17th and 20th years.
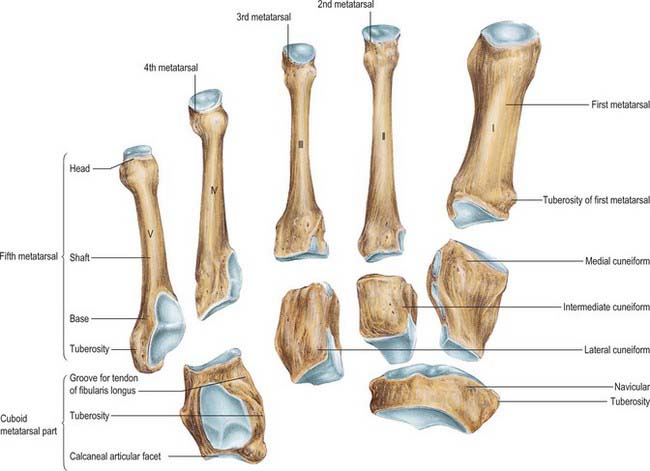
Fifth metatarsal
The fifth metatarsal has a tuberosity (styloid process) on the lateral side of its base (Figs 84.5A,B; 84.11). The base articulates proximally with the cuboid by a triangular, oblique surface, and medially with the fourth metatarsal. The tuberosity can be seen and felt midway along the lateral border of the foot; in acute inversion it may be fractured. The metaphysial–diaphysial junction of the fifth metatarsal base is prone to traumatic or stress fractures, and these have a tendency to delayed and non-union, and often require surgical fixation. It is believed that fractures at this level damage the nutrient artery and the extraosseous plexus, resulting in compromised vascularity of the fracture site and consequent poor fracture healing.
The tendon of fibularis tertius is attached to the medial part of the dorsal surface and medial border of the shaft, and that of fibularis brevis to the dorsal surface of the tuberosity. A strong band of the plantar aponeurosis, sometimes containing muscle, connects the apex of the tuberosity to the lateral process of the calcaneal tuberosity. It is this attachment, and not that of fibularis brevis, which is responsible for avulsion fractures of the tuberosity. The plantar surface of the base is grooved by the tendon of abductor digiti minimi, and flexor digiti minimi brevis is attached here. The lateral head of the fourth dorsal and the third plantar interossei are attached to the medial side of the shaft.
The fifth metatarsal is supplied by dorsal and plantar metatarsal arteries and an inconstant fibular marginal artery. The nutrient artery enters the diaphysis proximally and medially.
The fifth metatarsal is innervated by branches from the sural, superficial fibular and lateral plantar nerves.
There are three centres of ossification, one at the base in the region of the tuberosity (an apophysis), one in the shaft and one distally in the metatarsal head. Ossification of the shaft starts during the tenth prenatal week and ossification of the metatarsal head starts between the third and fourth years (Fig. 84.7). Fusion of the distal and shaft centres occurs between the 17th and 20th years, the proximal apophysis fuses earlier. An os vesalianum is a rare variant that should not be confused with the basal apophysis. (Fig. 84.12)
PHALANGES OF THE FOOT
In general, the phalanges of the foot resemble those of the hand: there are two in the hallux, and three in each of the other toes (Fig. 84.5A,B). On occasion there are only two phalanges in the little toe and, rarely, this is the case with the other lesser toes. The phalanges of the toes are much shorter than their counterparts in the hand and their shafts, especially those of the proximal set, are compressed from side to side. In the proximal phalanges, the compressed shaft is convex dorsally, with a plantar concavity. The base is concave for articulation with a metatarsal head and the head is a trochlea for a middle phalanx. Middle phalanges are small and short, but broader than their proximal counterparts. Distal phalanges resemble those in the fingers, but are smaller and flatter. Each has a broad base for articulation with a middle phalanx and an expanded distal end. A rough tuberosity on the plantar aspect of the latter supports the pulp of the toe, and provides a weight-bearing area.
Tendons of the long digital flexors and extensors are attached to the plantar and dorsal aspects of the bases of the distal phalanges of the lateral four toes. Flexor hallucis longus and extensor hallucis longus are similarly attached to the hallux. The bases of the middle phalanges receive the tendons of flexor digitorum brevis and extensor digitorum brevis. The proximal phalanges of the second, third, fourth and fifth toes each receive a lumbrical on their medial side; those of the second, third and fourth toes also receive an interosseous muscle on both sides. For further details of muscular, capsular and ligamentous arrangements in the toes refer to Fig. 84.5A,B. The terminal phalanx of the hallux normally shows a small degree of valgus (lateral) deviation, as may the proximal phalanx. This is presumed to be unrelated to footwear, because this degree of. deviation has also been observed in fetal specimens.
The proximal phalanges receive most of their blood supply from the dorsal digital arteries. The middle phalanges are supplied by plantar and dorsal digital arteries. The distal phalanges receive their supply mainly from plantar digital arteries.
Phalanges are ossified from a primary centre for the shaft and a basal epiphysis (Fig. 84.7). Primary centres for the distal phalanges appear between the ninth and twelfth prenatal weeks, somewhat later in the fifth digit. Primary centres for the proximal phalanges appear between the 11th and 15th weeks, and later for intermediate phalanges, but there is wide variation. Basal centres appear between the second and eighth years (usually second or third in the hallux), and union with the shaft occurs by the 18th year. There is considerable variation in ossification and fusion dates.
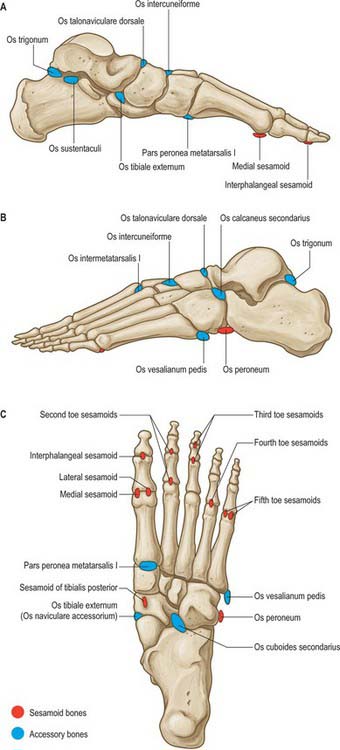
SESAMOIDS
Galen is believed to have been the first to use the term ‘sesamoid’ because of the resemblance of these bones to sesame seeds. Most sesamoid bones are only a few millimetres in diameter and their shape is variable. Some have a predictable location (see below), but many others vary in terms of location and frequency of occurrence (Fig. 84.12). Some sesamoids ossify, whereas others remain cartilaginous. Most sesamoid bones are embedded in tendons in close proximity to joints. Their precise role is not understood; it is believed that they may alter the direction of muscle pull, decrease friction and modify pressure.
Medial and lateral sesamoids of the first metatarsophalangeal joint
The two constant sesamoid bones within the foot are those of the first metatarsophalangeal articulation. The medial sesamoid is generally larger than the lateral sesamoid and lies slightly more distally. During dorsiflexion of the hallux, the sesamoids lie below the first metatarsal head, offering protection to the otherwise exposed plantar aspect of the first metatarsal head. The medial (tibial) sesamoid is approximately 10 mm wide and 14 mm long and the lateral (fibular) sesamoid is usually smaller (approximately 8 mm wide and 10 mm long): the overall sizes vary considerably.
The sesamoids are embedded within the double tendon of flexor hallucis brevis and articulate on their dorsal surfaces with the plantar facets of the first metatarsal head. They are separated by the crista or intersesamoidal ridge, which provides stability to the sesamoid complex. (This ridge can be eroded to the point of obliteration in severe cases of hallux valgus.) The sesamoids are connected to the plantar base of the proximal phalanx through the plantar plate, which is an extension of the tendon of flexor hallucis brevis. The plantar surface of each sesamoid is covered by a thin layer of the tendon of flexor hallucis brevis, whereas the dorsal or superior surface is covered by hyaline cartilage. The sesamoids are suspended by a sling-like mechanism made up of the collateral ligaments of the first metatarsophalangeal joint and the sesamoid ligaments on either side of the joint. The plantar aponeurosis also has an attachment to the sesamoids.
Approximately 30% of these sesamoid bones are bipartite. The medial is much more commonly affected and may have two, three or four parts, but the fibular sesamoid rarely has more than two. The condition may be bilateral. The sesamoids may be absent congenitally.
The medial sesamoid receives an attachment from abductor hallucis, which provides a medial stabilizing influence on the sesamoid complex. The lateral sesamoid receives some fibres from the tendon of adductor hallucis, and this provides lateral stabilization. The medial and lateral sesamoids are connected by the intersesamoid ligament, which forms the floor of the tendinous canal for the tendon of flexor hallucis longus.
There are three patterns of blood supply to the sesamoid bones. In 50% of cases the arterial supply is derived from the medial plantar artery and the plantar arch, in 25% of cases it is predominantly from the plantar arch and in 25% of cases it is from the medial plantar artery alone. The major arterial blood supply to the sesamoids enters from the proximal and plantar aspects, and only a minor contribution enters through the distal pole. The pattern of vascular supply determines the vulnerability of a sesamoid to avascular necrosis after injury to the bone.
Other sesamoid and accessory bones
Accessory or inconstant sesamoid bones may occur under any weight-bearing surface of the foot, but are most common under the second to fifth metatarsal heads. They are extremely variable in size and their incidence is difficult to determine.
A true sesamoid occurs in the tendon of tibialis posterior in about 10% of individuals. It lies on the plantar aspect of the navicular tuberosity within the tendon at the level of the inferior border of the calcaneonavicular ligament (see above, accessory navicular bones). Very rarely, a sesamoid is found within the tendon of tibialis anterior near its insertion at the level of the anteroinferior corner of the medial cuneiform, where there is an articular facet. An os peroneum is a sesamoid bone within the tendon of fibularis longus that articulates with the lateral surface of the calcaneum, the calcaneocuboid joint or, more frequently, the plantar aspect of the cuboid where there is an articular facet. The bone is situated where the tendon of fibularis longus angles around the plantar aspect of the lateral border of the cuboid and is probably present in 95% of cases; it is usually cartilaginous.
JOINTS
ANKLE (TALOCRURAL) JOINT
The talocrural joint is a hinge joint, approximately uniaxial. The lower end of the tibia and its medial malleolus, together with the lateral malleolus of the fibula and inferior transverse tibiofibular ligament, form a deep recess (‘mortise’) for the body of the talus. Although it appears to be a simple hinge, its axis of rotation is dynamic, shifting during dorsi- and plantar flexion. Starting from the plantigrade position the normal range of dorsiflexion is 10° when the knee is straight, and 30° with the knee flexed (when the calcaneal tendon will be relaxed). The range of normal plantar flexion is 30°. (The values of these ranges are all approximate.) Dorsiflexion results in the joint adopting the ‘close-packed’ position, with maximal congruence and ligamentous tension; from this position, all major thrusting movements are exerted, in walking, running and jumping. The malleoli grip the talus, and even in relaxation no appreciable lateral movement can occur without stretch of the inferior tibiofibular syndesmosis and slight bending of the fibula. The superior talar surface is broader in front, and in dorsiflexion the malleolar gap is increased by slight lateral rotation of the fibula, by ‘give’ at the inferior tibiofibular syndesmosis and gliding at the superior tibiofibular joint.
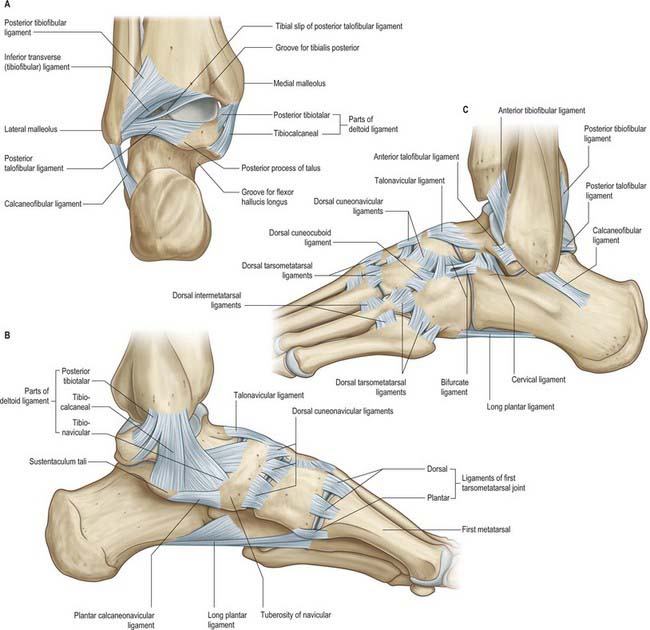
Articular surfaces are covered by hyaline cartilage. The talar trochlear surface, which is convex parasagittally and gently concave transversely, is wider in front; the distal tibial articular surface is reciprocally curved. The talar articular surface for the medial malleolus is a proximal area on the medial talar surface, and is fairly flat, comma-shaped and deeper anteriorly. The larger lateral talar articular surface is triangular and vertically concave, while the articular surface on the lateral malleolus is reciprocally curved. Posteriorly, the edge between the trochlear and fibular articular surfaces of the talus is bevelled to a narrow, flat triangular area that articulates with the inferior transverse tibiofibular ligament (Fig. 84.13A); all surfaces are contiguous. The bones are held together by a fibrous capsule, and by medial (deltoid), anterior and posterior talofibular and calcaneofibular ligaments.
Around the joint, the fibrous capsule is thin in front and behind. It is attached proximally to the borders of the tibial and malleolar articular surfaces, and distally to the talus near the margins of its trochlear surface, except in front where it reaches the dorsum of the talar neck. The capsule is strengthened by strong collateral ligaments. Its posterior part consists mainly of transverse fibres. It blends with the inferior transverse ligament and is thickened laterally where it reaches the fibular malleolar fossa.
The ligaments of the talocrural joint are the medial and lateral collateral ligaments.
Medial collateral ligament (deltoid ligament)
The medial collateral ligament (deltoid ligament) is a strong, triangular band, attached to the apex and the anterior and posterior borders of the medial malleolus (Fig. 84.13B). Of its superficial fibres, the anterior (tibionavicular) pass forwards to the navicular tuberosity, behind which they blend with the medial margin of the plantar calcaneonavicular ligament; inter-mediate (tibiocalcaneal) fibres descend almost vertically to the entire length of the sustentaculum tali; posterior fibres (posterior tibiotalar) pass posterolaterally to the medial side of the talus and its medial tubercle. The deep fibres (anterior tibiotalar) pass from the tip of the medial malleolus to the non-articular part of the medial talar surface. The ligament is crossed by the tendons of tibialis posterior and flexor digitorum longus. It is rarely injured alone, and when torn, is commonly associated with a fracture of the distal fibula. Chronic instability is rare.
The lateral ligament has three discrete parts. The anterior talofibular ligament extends anteromedially from the anterior margin of the fibular malleolus to the talus, attached in front of its lateral articular facet and to the lateral aspect of its neck (Fig. 84.13C). The posterior talofibular ligament runs almost horizontally from the distal part of the lateral malleolar fossa to the lateral tubercle of the posterior talar process (Fig. 84.13A); a ‘tibial slip’ of fibres connects it to the medial malleolus. The calcaneofibular ligament, a long cord, runs from a depression anterior to the apex of the fibular malleolus to a tubercle on the lateral calcaneal surface and is crossed by the tendons of fibularis longus and brevis (Fig. 8.13A,C). The lateral ligament complex is injured most commonly with inversion sprains, often during sport; the posterior talofibular ligament is almost always spared. Although the resulting increased laxity is tolerated in most cases, some require surgical reconstruction.
The joint is lined by synovial membrane which projects into the inferior (distal) tibiofibular joint.
Vascular supply and lymphatic drainage
The talocrural joint is supplied by malleolar branches of the anterior and posterior tibial and fibular arteries. Lymphatic drainage is via vessels accompanying the arteries and via the long and short saphenous veins superficially.
The talocrural joint is innervated by branches from the deep fibular, saphenous, sural and tibial nerves (or medial and lateral plantar nerves, depending on the level of division of the tibial nerve). Occasionally, the superficial fibular nerve also supplies the ankle joint.
For a comprehensive account of the innervation of the ankle joint (and other foot joints) see Gardner & Gray (1968).
Anteriorly, from medial to lateral, are tibialis anterior, extensor hallucis longus, the anterior tibial vessels, deep fibular nerve, extensor digitorum longus and fibularis tertius; posteromedially from medial to lateral, are tibialis posterior, flexor digitorum longus, the posterior tibial vessels, tibial nerve, flexor hallucis longus; in the groove behind the lateral malleolus are the tendons of fibularis longus and brevis. The tendon of fibularis brevis lies anterior to the tendon of fibularis longus at this level (Fig. 84.14). The long saphenous vein and saphenous nerve cross the ankle joint medial to the tendon of tibialis anterior and anterior to the medial malleolus, the nerve lying posterior to the vein.
All of the above structures are at risk during surgery on the ankle: the main structures at risk are the neurovascular structures anteriorly and posteromedially. Branches of the superficial fibular nerve are at risk on the anterolateral aspect of the ankle, particularly during ankle arthroscopy.
Passive stability is conferred upon the ankle mainly by the medial and lateral ligament complexes, the distal tibiofibular ligaments, the tendons crossing the joint, the bony contours and the capsular attachments. Dynamic stability is conferred by gravity, muscle action and ground reaction forces. Stability requires the continuous action of soleus assisted by gastrocnemius: it increases when leaning forward, and decreases when leaning backwards. If backward sway takes the projection of the centre of gravity (‘weight line’) posterior to the transverse axes of the ankle joints, the plantar flexors relax and the dorsiflexors contract.
Failure of the fibular muscles can lead progressively to varus instability, whereas long-standing failure of the tendon of tibialis posterior, which is relatively common in the elderly female, can result in valgus instability of the ankle and particularly a planovalgus foot deformity.
Ankle fractures
Ankle fractures are common and of importance because failure to achieve accurate anatomical alignment in the treatment of ankle fractures often results in significant long term morbidity. Except for very simple and undisplaced fractures, most ankle fractures are associated with a ligamentous injury. The direction and nature of forces applied to the ankle correlate with the fracture pattern and concomitant ligament injury. The Lauge–Hansen classification, although slightly cumbersome, classifies injuries according to two components: the position of the foot at the time of injury and the direction of the force applied. In a supination/adduction type injury, the foot is supinated and an adduction force is applied, resulting in a transverse fracture of the distal fibula (stage 1), followed by an oblique fracture of the medial malleolus (stage 2). In a supination/external rotation type injury, the first structure to rupture is the anterior inferior tibiofibular ligament, followed in order by a spiral fibular fracture, a posterior malleolar fracture and then a medial malleolar fracture (or deltoid ligament rupture). In pronation/abduction injuries, a transverse medial malleolar fracture is followed by rupture of the anterior and posterior inferior tibiofibular ligaments and interosseous ligament, and then by an oblique fibular fracture. Pronation/external rotation injuries commence with a medial malleolar fracture (or deltoid ligament rupture), followed in turn by rupture of the anterior inferior tibiofibular ligament, the interosseous ligament, the interosseous membrane, a high fibular fracture and fracture of the posterior tibial plafond (‘posterior malleolus’).
Inferior (distal) tibiofibular joint
The inferior (distal) tibiofibular joint is usually considered a syndesmosis. It consists of the anterior and posterior tibiofibular ligaments and the interosseous ligament.
The distal tibiofibular joint is between the rough, medial convex surface on the distal end of the fibula and the rough concave surface of the fibular notch of the tibia. These surfaces are separated distally for approximately 4 mm by a synovial prolongation from the ankle joint, and may be covered by articular cartilage in their lowest parts.
The ligaments of the inferior (distal) tibiofibular joint are the anterior, interosseous and posterior ligaments.
Anterior tibiofibular ligament
The anterior tibiofibular ligament is a flat band which descends laterally between the adjacent margins of the tibia and fibula, anterior to the syndesmosis (Fig. 84.13C). Bassett’s ligament is a variant that presents as a low lying slip of the ligament which is inserted so far distally on the fibula that it may cause irritation of the lateral dome of the talus: it is amenable to arthroscopic removal.
Interosseous tibiofibular ligament
The interosseous tibiofibular ligament is continuous with the interosseous membrane and contains many short bands between the rough adjacent tibial and fibular surfaces; it is the strongest bond between the bones.
Posterior tibiofibular ligament
The posterior tibiofibular ligament is stronger than the anterior, and is disposed similarly on the posterior aspect of the syndesmosis (Fig. 84.13A). Its distal, deep part is the inferior transverse ligament, a thick band of yellow fibres which crosses from the proximal end of the lateral malleolar fossa to the posterior border of the tibial articular surface almost to the medial malleolus. The ligament projects distal to the bones, in contact with the talus. Its colour reflects its content of yellow elastic fibres.
The synovial membrane of the ankle joint projects into the distal 4 mm or so of the inferior tibiofibular joint.
Vascular supply and lymphatic drainage
The inferior tibiofibular joint is supplied by the perforating branch of the fibular artery and the medial malleolar branches of the anterior and posterior tibial arteries. Lymphatic drainage is via vessels corresponding to the arteries, and via vessels that accompany the long and short saphenous veins.
No significant structures pass anterior to the anterior aspect of the inferior tibiofibular joint, but the superficial fibular nerve is at risk during surgery to this area. Posteriorly, the fibular artery passes over the posterior tibiofibular ligament and is at risk in the posterolateral approach to the fibula.
Stability is maintained in part by the bone contours, but mainly by the dense anterior and posterior tibiofibular and the interosseous ligaments.
No muscles act on the inferior tibiofibular joint, which moves only slightly. Because of the varying slope of the lateral surface of the body of talus, the fibula undergoes a small degree of lateral rotation during dorsiflexion at the ankle, and this results in slight widening of the interval between the bones. Testing for a significant disruption of this joint is usually carried out with the patient under anaesthetic and involves applying an external rotation and abduction force, looking for abnormal widening of the syndesmosis. Excessive anterior/posterior glide of the fibula relative to the tibia also indicates a disruption of the joint.
Talocalcaneal joint
Anterior and posterior articulations between the calcaneus and talus form a functional unit often termed the ‘subtalar joint’. The posterior articulation is referred to as the talocalcaneal joint and the anterior articulation is regarded as part of the talocalcaneonavicular joint. The talocalcaneal joint is modified multiaxial and its permitted movements are considered together with those at other tarsal joints. The bones are connected by a fibrous capsule, and by lateral, medial, interosseous talocalcaneal and cervical ligaments.
The subtalar joint proper involves the concave posterior calcaneal facet on the posterior part of the inferior surface of the talus and the convex posterior facet on the superior surface of the calcaneus (Fig. 84.15A,B).
The ligaments of the talocalcaneal joint are the lateral, medial and interosseous talocalcaneal ligaments and the cervical ligament.
Lateral talocalcaneal ligament
The lateral talocalcaneal ligament is a short flat fasciculus that descends obliquely back from the lateral talar process to the lateral calcaneal surface and is attached anterosuperior to the calcaneofibular ligament.
The medial talocalcaneal ligament connects the medial talar tubercle to the back of the sustentaculum tali and adjacent medial surface of the calcaneus. Its fibres blend with the medial (deltoid) ligament of the ankle joint; the most posterior fibres line the groove for flexor hallucis longus between the talus and calcaneus.
Interosseous talocalcaneal ligament
The interosseous talocalcaneal ligament is a broad, flat, bilaminar transverse band in the sinus tarsi (Figs 84.15A; 84.16). It descends obliquely and laterally from the sulcus tali to the calcaneal sulcus. The posterior lamina of the ligament is associated with the talocalcaneal joint, and the anterior lamina with the talocalcaneonavicular joint. Its medial fibres are taut in eversion.
The cervical ligament is just lateral to the tarsal sinus and attached to the superior calcaneal surface (Fig. 84.13C). It is medial to the attachment of extensor digitorum brevis, from where it ascends medially to an inferolateral tubercle on the talar neck (Barclay-Smith 1896). It is considered to be taut in inversion.
The synovial cavity of the talocalcaneal joint is usually quite separate and does not communicate with those of other tarsal joints. However, direct communication with the ankle joint has been observed in rare instances.
The talocalcaneal joint is innervated by branches of the posterior tibial, medial plantar and sural nerves.
Posteromedially, in the region of the posterior aspect of the talocalcaneal joint, and viewed from anterior to posterior, lie the veins on either side of the posterior tibial artery, the tibial nerve and the tendon of flexor hallucis longus. These neurovascular structures are at risk in posteromedial approaches to the ankle and talocalcaneal joints. On the lateral side, the tendon of fibularis brevis lies anterior to the tendon of fibularis longus, both passing behind the fibular malleolus in proximity to the talocalcaneal joint. The sural nerve lies just posterior to the fibular tendons and is at risk during lateral exposure of the joint.
Stability is conferred by the bony contours of the hindfoot plus the above mentioned ligaments; it is not known which ligaments provide the most stability. An additional ligamentous restraint is provided by the calcaneofibular component of the lateral ligament complex. The tendons crossing the articulation also aid stability.
Heel inversion is controlled by tibialis anterior, tibialis posterior and the gastrocnemius–soleus complex via the calcaneal tendon; the long flexors of the toes also contribute. Heel eversion results from the pull of fibularis longus, brevis and tertius in addition to the long extensors of the toes.
Talocalcaneonavicular joint
An eloquent account of the talocalcaneonavicular joint was provided by Barclay-Smith in 1896. In terms of function, and in clinical practice, it is helpful to regard this complex joint as comprising two articulations, i.e. the anterior part of the ‘subtalar’ joint and the talonavicular joint. It is a compound, multiaxial articulation.
The ovoid talar head is continuous with the triple-faceted anterior area of its inferior surface. The whole fits the concavity formed collectively by the posterior surface of the navicular, the middle and anterior talar facets of the calcaneus, and the superior fibrocartilaginous surface of the plantar calcaneonavicular ligament (spring ligament). The bones are connected by a fibrous capsule and by the talonavicular and plantar calcaneonavicular ligaments, and the calcaneonavicular part of the bifurcate ligament.
The fibrous capsule is poorly developed, except posteriorly, where it is thick and blends with the anterior part of the interosseous ligament filling the tarsal sinus.
The ligaments of the talocalcaneonavicular joint are the talonavicular and plantar calcaneonavicular (spring) ligaments.
The talonavicular ligament is a broad, thin band (Fig. 84.13B,C). It connects the dorsal surfaces of the neck of the talus and the navicular, and is covered by extensor tendons. The plantar calcaneonavicular ligament and the calcaneonavicular part of the bifurcate ligament (Fig. 84.13C) are the plantar and lateral ligaments of the joint, respectively. Although the calcaneus and navicular do not articulate directly, they are connected by calcaneonavicular and plantar calcaneonavicular ligaments.
Plantar calcaneonavicular (spring) ligament
The plantar calcaneonavicular (spring) ligament is a broad, thick band connecting the anterior margin of the sustentaculum tali to the plantar surface of the navicular (Figs 84.13B; 84.15A; 84.17). It ties the calcaneus to the navicular below the head of the talus as part of its articular cavity and it sustains the medial longitudinal arch of the foot. The dorsal surface of the ligament has a triangular fibrocartilaginous facet on which part of the talar head rests (Fig. 84.15A). Its plantar surface is supported medially by the tendon of tibialis posterior and laterally by the tendons of flexors hallucis longus and digitorum longus; its medial border is blended with the anterior superficial fibres of the medial (deltoid) ligament. The calcaneonavicular ligament is described below as the medial band of the bifurcate ligament.
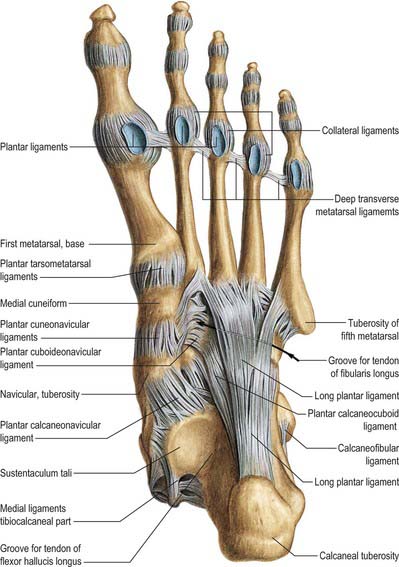
The talocalcaneonavicular joint is a synovial joint whose cavity sometimes communicates with that of the talocalcaneal joint.
On the medial side from dorsal to plantar lie the tendon of tibialis posterior and the tendon of flexor digitorum longus above the sustentaculum tali, and the flexor hallucis longus tendon below. At this point, flexor hallucis longus lies deep to the medial and lateral plantar branches of the posterior tibial artery and nerve. The latter structures are at risk during medial approaches to the joint, e.g. resection of talocalcaneal coalition. Dorsally from medial to lateral lie the tendons of tibialis anterior and extensor hallucis longus, the deep fibular nerve and the dorsalis pedis artery, the muscle belly of extensor hallucis brevis passing medially and deep to the tendons of extensor digitorum longus and fibularis tertius. Structures at risk during the dorsal approach to the talonavicular joint include the branches of the superficial fibular nerve superficially and the dorsalis pedis artery and deep fibular nerve deep to the inferior extensor retinaculum.
Calcaneocuboid joint
The calcaneocuboid joint is at the same level as the talonavicular joint and, together, they represent the transverse tarsal joint. It is a saddle (sellar) or biaxial joint with concavo-convex surfaces.
The articular surfaces of the calcaneocuboid joint, which is 2 cm proximal to the tubercle on the fifth metatarsal base, are between the anterior (distal) surface of the calcaneus and the posterior (proximal) surface of the cuboid.
The fibrous capsule is thickened dorsally as the dorsal calcaneocuboid ligament. The synovial cavity of this joint is separate, and does not communicate with those of other tarsal articulations (Fig. 84.6).
The ligaments of the calcaneocuboid joint are the bifurcate, long plantar and plantar calcaneocuboid ligaments.
The bifurcate ligament is a strong Y-shaped band (Fig. 84.13C). It is attached by its stem proximally to the anterior part of the upper calcaneal surface, and distally it divides into calcaneocuboid and calcaneonavicular parts. The (lateral) calcaneocuboid ligament extends to the dorsomedial aspect of the cuboid, forming a main bond between the two rows of tarsal bones; the (medial) calcaneonavicular ligament is attached to the dorsolateral aspect of the navicular.
The long plantar ligament is the longest ligament associated with the tarsus (Figs 84.13C; 84.17; 84.18). It extends from the plantar surface of the calcaneus (anterior to the processes of its tuberosity) and from its anterior tubercle, to the ridge and tuberosity on the plantar surface of the cuboid. Deep fibres are attached to the cuboid and more superficial fibres continue to the bases of the second to fourth, and sometimes fifth, metatarsals. This ligament, together with the groove on the plantar surface of the cuboid, makes a tunnel for the tendon of fibularis longus. It is a most powerful factor limiting depression of the lateral longitudinal arch.
Plantar calcaneocuboid ligament
This short plantar ligament (Fig. 84.17) is deeper than the long plantar ligament, from which it is separated by areolar tissue. It is a short, wide band of great strength, stretching from the anterior calcaneal tubercle and the depression anterior to it, to the adjoining part of the plantar surface of the cuboid; it also sustains the lateral longitudinal arch.
The calcaneocuboid joint is innervated on its plantar aspect by the lateral plantar nerve, and dorsally by the sural and deep fibular nerves.
Fibularis longus and abductor digiti minimi pass in proximity to the joint, and the lateral plantar nerve passes medially. Extensor digitorum brevis overlies the lateral aspect of the joint. The sural nerve and the tendon of fibularis longus are at risk during approaches to an os peroneum.
Naviculocuneiform joints
The naviculocuneiform joint is a compound joint, often described as plane.
The navicular articulates distally with the cuneiform bones where the distal navicular surface is transversely convex and divided into three facets by low ridges that are adapted to the proximal, slightly curved surfaces of the cuneiforms.
The fibrous capsule is continuous with those of the intercuneiform and cuneocuboid joints and it is also connected to the second and third cuneometatarsal joints and intermetatarsal joints between the second to fourth metatarsal bones.
The ligaments of the naviculocuneiform joint are the dorsal and plantar ligaments.
The dorsal and plantar ligaments connect the navicular to each cuneiform; of the three dorsal ligaments, one is attached to each cuneiform. The fasciculus from the navicular to the medial cuneiform is continued as the capsule of the joint around its medial aspect, and then blends medially with the plantar ligament. Plantar ligaments have similar attachments and receive slips from the tendon of tibialis posterior.
The naviculocuneiform joint is innervated dorsally by branches from the deep fibular nerve. The medial and intermediate naviculocuneiform joints receive a plantar innervation from the medial plantar nerve, and the lateral naviculocuneiform joint is innervated from the lateral plantar nerve.
The tendon of tibialis posterior sends fibres to the medial cuneiform and crosses the joint on its medial side; it is vulnerable to injury when a medial surgical approach is used, as are the dorsal venous arch and the saphenous nerve. The tendon of tibialis anterior passes over the dorsomedial aspect of the joint and the tendon of extensor hallucis longus is lateral. Running over the intermediate cuneiform, from medial to lateral, are the deep fibular nerve, the dorsalis pedis artery and the muscle belly of extensor hallucis brevis, all of which are vulnerable during dorsal exposure of the joint. On the lateral aspect of the joint, the superficial structures at risk are the superficial fibular nerve and the dorsal venous arch.
Movements at the naviculo-cuneiform, cuboideonavicular, intercuneiform and cuneocuboid joints are slight and subtle gliding and rotational movements that occur during pronation or supination of the foot: when alterations occur in a loaded foot in contact with the ground, they increase suppleness when the forefoot is stressed, e.g. in the initial thrust of running and jumping. The muscles responsible for these slight movements are tibialis anterior, tibialis posterior, fibularis longus and brevis and the long flexors and extensors of the toes.
Cuboideonavicular joint
The cuboideonavicular joint is usually a fibrous joint, the bones being connected by dorsal, plantar and interosseous ligaments. This syndesmosis is often replaced by a synovial joint that is almost plane; its articular capsule and synovial lining are continuous with that of the naviculocuneiform joint. The dorsal ligament extends distolaterally, the plantar nearly transversely from the cuboid to the navicular. The interosseous ligament is made of strong transverse fibres and connects non-articular parts of adjacent surfaces to the two bones (Fig. 84.13C).
Intercuneiform and cuneocuboid joints
The intercuneiform and cuneocuboid joints are all synovial and approximately plane or slightly curved. Their articular capsules and synovial linings are continuous with those of the naviculocuneiform joints. The bones are connected by dorsal, plantar and interosseous ligaments (Fig. 84.13B,C; 84.15A).
The ligaments of the intercuneiform and cuneocuboid joints are the dorsal, plantar and interosseous ligaments.
Tarsometatarsal articulations
Tarsometatarsal articulations are approximately plane synovial joints.
The first metatarsal articulates with the medial cuneiform; the second is recessed between the medial and lateral cuneiforms and articulates with the intermediate cuneiform; the third articulates with the lateral cuneiform; the fourth articulates with the lateral cuneiform and the cuboid; the fifth articulates with the cuboid. The joints are approximately on a line from the tubercle of the fifth metatarsal to the tarsometatarsal joint of the hallux, except for that between the second metatarsal and intermediate cuneiform, which is 2–3 mm proximal (Figs 84.5A; 84.11).
The hallucal joint has its own capsule. The articular capsules and cavities of the second and third are continuous with those of the intercuneiform and naviculocuneiform joints, but are separated from the fourth and fifth joints by an interosseous ligament between the lateral cuneiform and fourth metatarsal base.
The bones are connected by dorsal and plantar tarsometatarsal and interosseous cuneometatarsal ligaments.
The dorsal ligaments are strong and flat. The first metatarsal is joined to the medial cuneiform by an articular capsule and the other tarsometatarsal capsules blend with the dorsal and plantar ligaments. The second metatarsal receives a band from each cuneiform, the third from the lateral cuneiform, the fourth from the lateral cuneiform and the cuboid, the fifth from the cuboid alone.
The plantar ligaments are longitudinal and oblique bands, and are less regular than the dorsal ligaments. Those for the first and second metatarsals are strongest. The second and third metatarsals are joined by oblique bands to the medial cuneiform, the fourth and fifth by a few fibres to the cuboid.
Interosseous cuneometatarsal ligaments
There are three interosseous cuneometatarsal ligaments. One (the strongest) passes from the lateral surface of the medial cuneiform to the adjacent angle of the second metatarsal (Fig. 84.17). Known as Lisfranc’s ligament, it is crucial to the stability of the tarsometatarsal joint complex. Disruption of this ligament can lead to instability and deformity and subsequent degenerative changes. A second ligament connects the lateral cuneiform to the adjacent angle of the second metatarsal: it does not divide the joint between the second metatarsal and lateral cuneiform and is inconstant. A third ligament connects the lateral angle of the lateral cuneiform to the adjacent fourth metatarsal base.
The first tarsometatarsal joint has its own capsule, with a synovial lining. The other joints are similarly arranged, but there is a communication between the second and third and between the fourth and fifth tarsometatarsal joints.
The interosseous cuneometatarsal ligaments are innervated dorsally via the deep fibular nerve. The plantar aspects of the medial two joints are innervated from the medial plantar nerve, and the plantar aspects of the lateral joints are innervated by the lateral plantar nerve.
From medial to lateral, the following structures cross the dorsal aspect of the tarsometatarsal joints: the saphenous nerve, the dorsal venous arch, the tendon of extensor hallucis longus, the dorsalis pedis artery, the deep fibular nerve, extensor hallucis brevis, the tendons of extensor digitorum longus, extensor digitorum brevis, fibularis tertius and fibularis brevis. Branches of the superficial fibular nerve are variable in location on the dorsum. The sural nerve lies just inferior to the tendon of fibularis brevis in the subcutaneous tissues. Surgery involving the tarsometatarsal joints is performed through a dorsal approach, and therefore all of these structures are potentially at risk.
The major stabilizers of the tarsometatarsal joints are the associated ligaments. However, despite the strength of the ligaments, this joint complex is vulnerable to injury because of the lack of ligamentous connection between the first and second metatarsal bases. The medial interosseous ligament or Lisfranc’s ligament is responsible for the tell-tale avulsion fracture of the second metatarsal base, which may be the only radiographic indication of abnormality after injury to this part of the foot. It is a highly significant injury that is often missed: failure to recognize and treat this injury by surgical reduction and fixation can lead to long-term disability.
Movements between the tarsal and metatarsal bones are limited to flexion and extension, except in the first tarsometatarsal joint, where some abduction and rotation also occur. The muscles that produce this motion are tibialis anterior and fibularis longus. Flexion and extension are brought about by the long and short flexors and extensors of the toes. Movement between the medial cuneiform and first metatarsal, and between the fourth and fifth metatarsal bases and the cuboid, is moderate and allows the foot to adapt to uneven surfaces, whereas movement between the second and third metatarsal bases and their corresponding cuneiforms is very limited.
Intermetatarsal joints
The intermetatarsal interosseous ligaments are very strong and are present between all the lateral four metatarsals; they are absent between the first and second metatarsals. The base of the second metatarsal is joined to the first tarsometatarsal joint by the medial interosseous ligament (Lisfranc’s ligament) which connects the plantar aspect of the second metatarsal to the medial cuneiform.
The ligaments of the intermetatarsal joints include the dorsal and plantar intermetatarsal ligaments.
Dorsal and plantar intermetatarsal ligaments
As with their dorsal counterparts, the plantar intermetatarsal ligaments are longitudinal, oblique or transverse and they vary considerably in both number and organization. The plantar ligaments are significantly stronger than the corresponding dorsal ligaments. The strongest is the second oblique plantar ligament, which connects the medial cuneiform to the second and third bases (Fig. 84.17).
All the metatarsal heads are connected indirectly by deep transverse metatarsal ligaments. Dorsal and plantar ligaments pass transversely between adjacent bases; interosseous ligaments are strong transverse bands that connect non-articular parts of the adjacent surfaces (Fig. 84.17).
Metatarsophalangeal articulations
Metatarsophalangeal articulations are ovoid or ellipsoid joints between the rounded metatarsal heads and shallow cavities on the proximal phalangeal bases. They are usually 2.5 cm proximal to the webs of the toes.
Articular surfaces cover the distal and plantar, but not the dorsal, aspects of the metatarsal heads. The plantar aspect of the first metatarsal head has two longitudinal grooves separated by a ridge (the crista). Each articulates with a sesamoid bone embedded in the capsule of the joint, formed here by the two tendons of flexor hallucis brevis. The sesamoids are connected to each other by the intersesamoid ligament, which forms the floor of the tendinous canal for the tendon of flexor hallucis longus. The medial sesamoid receives an attachment from abductor hallucis and the lateral sesamoid receives an attachment from adductor hallucis.
Articular areas on the proximal phalangeal bases are concave. The ligaments are capsular, plantar, deep transverse metatarsal and collateral.
Fibrous capsules are attached to their articular margins. They are thin dorsally, and may be separated from the long extensor tendons by small bursae, or they may be replaced by the tendons, but they are inseparable from the plantar and collateral ligaments. The plantar aponeurosis blends with the plantar capsule to form the so-called ‘plantar plate’ which inserts distally into the base of the proximal phalanx via medial and lateral bundles. Proximally, the plate is attached to the metatarsal head via a thin synovial fold. It also receives an attachment from the accessory collateral ligament.
The ligaments of the metatarsophalangeal joints are the plantar, deep transverse metatarsal and collateral ligaments.
The plantar ligaments are thick and dense. They lie between and blend with the collateral ligaments, being loosely attached to the metatarsals, and firmly attached to the phalangeal bases. Their margins blend with the deep transverse metatarsal ligaments. Their plantar surfaces are grooved for the flexor tendons, the fibrous sheaths of which connect with the edges of the grooves, and their deep surfaces extend the articular areas for metatarsal heads.
Deep transverse metatarsal ligaments
The deep transverse metatarsal ligaments are four short, wide, flat bands that unite the plantar ligaments of adjoining metatarsophalangeal joints. The interossei are dorsal to, and the lumbricals, digital vessels and nerves are plantar to, these ligaments. They resemble the deep transverse metacarpal ligaments except that, in the foot, there is a deep transverse metacarpal ligament between the plantar ligament of the second metatarsophalangeal joint and that of the hallucal metatarsophalangeal joint (Fig. 84.17).
The collateral ligaments are strong cords flanking each joint. They are attached to the dorsal tubercles on the metatarsal heads and the corresponding side of the phalangeal bases and they slope downwards and forwards. The first metatarsophalangeal joint also contains metatarso-sesamoid ligaments. On either side, the ligaments arise from the metatarsal head with a narrow origin and then fan out to insert on the border of the proximal phalanx and the plantar plate. Each collateral ligament consists of the phalangeal collateral ligament, which inserts into the base of the proximal phalanx, and the accessory collateral ligament, which inserts into the plantar plate.
The main nerve supply of the metatarsophalangeal joints is from the plantar interdigital nerves, which supply the first, second, third and medial half of the fourth metatarsophalangeal joint on their plantar aspects. Digital branches of the lateral plantar nerve supply the lateral half of the fourth joint, and both medial and lateral aspects of the fifth joint, on their plantar sides. The dorsomedial side of the hallucal metatarsophalangeal joint is supplied by the medial dorsal cutaneous branch of the superficial fibular nerve. The deep fibular nerve supplies the dorsolateral side of the hallucal metatarsophalangeal joint and the medial side of the metatarsophalangeal joint of the second toe.
Dorsally, the tendon of extensor hallucis longus lies medial to the tendon of extensor hallucis brevis. The same arrangement occurs in the lesser toes with the tendons of extensor digitorum longus and brevis. The interossei are plantarmedial and plantarlateral, dorsal to the intermetatarsal ligament, whereas the lumbrical tendons and the digital artery and nerve are plantar to the intermetatarsal ligament. The long and short flexors lie on the plantar aspect of the joint in the midline. If approaching the metatarsophalangeal joint surgically from the plantar surface, it is important not to stray from the midline.
The first metatarsophalangeal joint owes its stability to its capsuloligamentous structures, and to flexor and extensor hallucis brevis, with a small contribution from flexor and extensor hallucis longus or the bony contours. The metatarsophalangeal joints of the lesser toe are stabilized principally by the collateral ligaments and plantar plates. Rupture of the plantar plate leads to hammer toe deformity.
The types of movements that occur at these joints are like those that occur at the corresponding joints in the hand, but the range of movement is quite different. In contrast to the metacarpophalangeal joints, the range of active extension that can occur at the metatarsophalangeal joints (50–60°) is greater than that of flexion (30–40°): this is an adaptation to the needs of walking, and is most marked in the hallucal joint, where flexion is a few degrees while extension may reach 90°. When the foot is on the ground, metatarsophalangeal joints are already extended to at least 25° because the metatarsals incline proximally in the longitudinal arches of the foot (Fig. 84.8A). The range of passive movements in these joints is 90° (extension) and 45° (flexion), according to Kapandji (1974). The following muscles produce movements at the metatarsophalangeal joints:
Flexor digitorum brevis, lumbricals and interossei, assisted by flexores digitorum longus and accessorius. In the fifth toe, flexor digiti minimi brevis assists. In the hallux, flexores hallucis longus and brevis are the only flexors.
Adductor hallucis; in the third to fifth toes, the first, second and third plantar interossei, respectively.
Abductor hallucis; in the second toe, the first and second dorsal interossei; in the third and fourth toes, the corresponding dorsal interossei; in the fifth toe, abductor digiti minimi.
Note: The line of reference for adduction and abduction is along the second digit, which has the least mobile metatarsal. The second toe may therefore be ‘abducted’ medially and laterally by the first and second dorsal interossei respectively.
Hallux valgus is a common condition, occurring mainly in individuals who have a genetic predisposition. Footwear is implicated in the condition, which presumably accounts for the greater incidence in females. Metatarsus primus varus, an adduction deformity of the first metatarsal, is commonly associated with hallux valgus.
The more spheroidal the shape of the first metatarsal head, the more likely it is to be unstable. Conversely, a flat metatarsal head is less likely to be associated with hallux valgus. No muscle inserts into the first metatarsal head, and therefore its position is determined by the position of the proximal phalanx. As the proximal phalanx moves laterally on the metatarsal head, it pushes the head medially. This leads to attenuation of the medial soft tissue structures and contracture of the lateral ones. The sesamoid sling, which is anchored laterally by adductor hallucis, remains in place as the head moves medially, displacing the sesamoids from beneath the metatarsal head. As this happens, the weakest point of the medial capsule fails, so that abductor hallucis slips under the metatarsal head. This leads to failure of the intrinsic muscles to stabilize the joint, and the pull of abductor hallucis leads to spinning of the proximal phalanx, which results in a pronation deformity. Failure to intervene surgically inevitably results in a progressive deformity.
Interphalangeal articulations
Interphalangeal articulations are almost pure hinge joints, in which the trochlear surfaces on the phalangeal heads articulate with reciprocally curved surfaces on adjacent phalangeal bases. Each has an articular capsule and two collateral ligaments, as occurs in the metatarsophalangeal joints. The plantar surface of the capsule is a thickened fibrous plate, like the plantar metatarsophalangeal ligaments, and is often termed the plantar ligament.
The interphalangeal articulations are innervated by branches from the plantar interdigital nerves. The medial dorsal cutaneous branch of the superficial fibular nerve also supplies the hallucal interphalangeal joint. Branches of the deep fibular, intermediate dorsal cutaneous and sural nerves sometimes supply the joints of the lesser toes.
Movements are flexion and extension, which are greater in amplitude between the proximal and middle phalanges than between the middle and distal. Flexion is marked, but extension is limited by tension of the flexor muscles and plantar ligaments. Abduction, adduction and rotation occur to a minor extent.
ARCHES OF THE FOOT
Three main arches are recognized in the foot. They are the medial longitudinal, the lateral longitudinal and the transverse arch. The roles of the arches of the foot in standing, walking and running are discussed later in this chapter.
MEDIAL LONGITUDINAL ARCH
The medial margin of the foot arches up between the heel proximally and the medial three metatarsophalangeal joints to form a visible arch (Fig. 84.8A). It is made up of the calcaneus, talar head, navicular, the three cuneiforms and the medial three metatarsals. The posterior and anterior pillars are the posterior part of the inferior calcaneal surface and the three metatarsal heads, respectively. The bones themselves contribute little to the stability of the arch, whereas the ligaments contribute significantly. The most important ligamentous structure is the plantar aponeurosis, which acts as a tie beam between the supporting pillars (Hicks 1953). Dorsiflexion, especially of the hallux, draws the two pillars together, thus heightening the arch: the so-called ‘windlass’ mechanism. Next in importance is the spring ligament, which supports the head of the talus. If this ligament fails, the navicular and calcaneus separate, allowing the talar head, which is the highest point of the arch, to descend. The talocalcaneal ligaments and the anterior fibres of the deltoid ligament, from the tibia to the navicular, also contribute to the stability of the arch.
Muscles play a role in the maintenance of the medial longitudinal arch. Flexor hallucis longus acts as a bowstring. Flexor digitorum longus, abductor hallucis and the medial half of flexor digitorum brevis also contribute, but to a lesser extent. Tibialis posterior and anterior invert and adduct the foot, and so help to raise its medial border. The importance of tibialis posterior is manifest by the collapse of the medial longitudinal arch that accompanies failure of its tendon (see below).
LATERAL LONGITUDINAL ARCH
The lateral longitudinal arch is much less pronounced arch than the medial one. The bones making up the lateral longitudinal arch are the calcaneus, the cuboid and the fourth and fifth metatarsals: they contribute little to the arch in terms of stability (Fig. 84.8B). The pillars are the calcaneus posteriorly, and the lateral two metatarsal heads, anteriorly. Ligaments play a more important role in stabilizing the arch, especially the lateral part of the plantar aponeurosis and the long and short plantar ligaments. However, the most important contribution to the maintenance of the lateral arch is made by the tendon of fibularis longus. The lateral two tendons of flexor digitorum longus (and flexor accessorius), the muscles of the first layer (lateral half of flexor digitorum brevis and abductor digiti minimi) and fibularis brevis and peronius tertius also contribute to the maintenance of the lateral longitudinal arch.
TRANSVERSE ARCH
The bones involved in the transverse arch are the bases of the five metatarsals, the cuboid and the cuneiforms (Fig. 84.5B). The intermediate and lateral cuneiforms are wedge-shaped and thus adapted to maintenance of the transverse arch. The stability of the arch is mainly provided by the ligaments, which bind the cuneiforms and the metatarsal bases, and also by the tendon of fibularis longus, which tends to approximate the medial and lateral borders of the foot. A shallow arch is maintained at the metatarsal heads by the deep transverse ligaments, transverse fibres that bind together the digital slips of the plantar aponeurosis, and, to a lesser extent, by the transverse head of adductor hallucis.
PES PLANUS AND PES CAVUS
The term pes planus denotes an excessively flat foot. There is no precise degree of flatness that defines pes planus, but it may be either physiological or pathological. In physiological pes planus, the feet are flexible and rarely problematic. In marked contrast, pathological pes planus is often associated with stiffness and pain. The windlass (or ‘Jack’s great toe’) test involves passively dorsiflexing the hallux at the metatarsophalangeal joint. This tightens the plantar aponeurosis and, in flexible pes planus, results in accentuation of the medial longitudinal arch. In pathological pes planus, no accentuation of the arch is seen. This test can also be carried out by asking the individual to stand on tiptoe and viewing the hindfoot from behind. In flexible flat feet, the calcaneus swings into a varus position; in pathological pes planus it does not. Causes of pathological pes planus include tarsal coalition, disruption of the tendon of tibialis posterior, rupture of the spring ligament, tarsometatarsal arthritis (and subsequent collapse) and hindfoot (talo-calcaneal or subtalar joint) degenerative or inflammatory arthritis.
Pes cavus denotes an excessively high-arched foot. The majority of cases arise as a result of a neurological disorder (e.g. Charcot–Marie–Tooth disease, diastematomyelia, poliomyelitis). According to the anatomical location of the deformity, pes cavus may be classified into hindfoot, midfoot or forefoot cavus. When pes cavus involves all three parts of the foot it is called ‘global’ cavus.
In Charcot–Marie–Tooth disease, an overactive fibularis longus leads to plantar hyperflexion of the first metatarsal. To keep the forefoot in contact with the ground, the affected individual develops a progressive compensatory hindfoot varus. If ignored, the hindfoot varus, which is initially flexible, becomes fixed. The Coleman block test involves standing with the lateral part of the forefoot on a wooden block (Coleman & Chesnut 1977). This effectively balances the forefoot supination and the hindfoot deformity corrects itself spontaneously, provided the hindfoot varus is flexible. If the hindfoot is still flexible, surgery need only address the forefoot supination. If fixed, however, the hindfoot deformity also needs to be surgically corrected.
MUSCLES
The muscles acting on the foot may be divided into extrinsic and intrinsic groups.
EXTRINSIC MUSCLES
The extrinsic muscles are described in Chapter 83. Their tendons cross the ankle, and move and stabilize this joint. Distally, the tendons also act on the small joints of the foot and help to stabilize these joints. The muscles can be grouped according to their arrangement in the leg. The extensors arise in the anterior compartment of the leg and their tendons pass anterior to the ankle, where they are bound down by the extensor retinacula. The lateral group arises in the relatively narrow lateral compartment of the leg and their tendons pass posterior to the lateral malleolus, bound down by the fibular retinacula. The flexors arise in the posterior compartment of the leg and their tendons pass posterior to the ankle, where the tendons of the superficial group of flexors are inserted into the calcaneus (see below) and the tendons of the deep group of flexors are bound down by the flexor retinaculum.
Anterior group
Tibialis anterior, extensor hallucis longus, extensor digitorum longus and fibularis tertius are all described in Chapter 83.
Posterior group
Superficial group
Gastrocnemius and soleus are described in Chapter 83; the calcaneal tendon is described below.
Calcaneal (Achilles) tendon
The calcaneal tendon is the thickest and strongest tendon in the human body (see Fig. 82.3). It is approximately 15 cm long, and begins near the middle of the calf, although its anterior surface receives muscle fibres from soleus almost to its lower end. It gradually becomes more rounded until approximately 4 cm above the calcaneus, below this level it expands and becomes attached to the midpoint of the posterior surface of the calcaneus. The fibres of the calcaneal tendon are not aligned strictly vertically and they display a variable degree of spiralization (Cummins et al 1946).
The blood supply to the calcaneal tendon is poor: the predominant artery is a recurrent branch of the posterior tibial artery which mainly supplies peritendinous tissues (Taylor & Razaboni 1994). There is an additional supply proximally from intramuscular arterial branches and distally from the calcaneus. The vascularity of the mid-substance of the tendon is relatively poor, although some blood enters the tendon at this point through its anterior surface. During surgery, the paratenon is therefore best left intact in this region.
The calcaneal tendon is not the only plantar flexor of the ankle, which is one of the reasons ruptures of the calcaneal tendon may not always be clinically apparent. However, it is a frequent site of pathology because of its susceptibility to rupture, degenerative change (tendinosis) and inflammation (paratendonitis): the area of relative avascularity in the mid-substance of the tendon is where the majority of problems occur.
The calcaneal tendon is subcutaneous. The sural nerve crosses its lateral border about 10 cm above its insertion: the nerve is especially vulnerable here to iatrogenic injury during surgery. Distally, there are bursae superficial and deep to the tendon. The muscle belly of flexor hallucis longus lies deep to the deep fascia on the anterior surface of the tendon.
Plantar flexion of the ankle joint. The tendon fibres spiral laterally through 90° as they descend, so that the fibres associated with gastrocnemius come to insert on the bone more laterally, and those associated with soleus more medially.
There are three locations about the heel where bursae occur. The most common is the retrocalcaneal bursa, which lies between the calcaneal tendon and the posterior surface of the calcaneus. An almost constant finding, it has an anterior bursal wall composed of fibrocartilage and a thin posterior wall which blends with the thin epitenon (epitendineum) of the calcaneal tendon. Dorsiflexion of the ankle results in compression of the bursa. Less common are an adventitious bursa superficial to the calcaneal tendon, and a subcalcaneal bursa between the inferior surface of the calcaneus and the origin of the plantar aponeurosis. A prominent superolateral calcaneal tuberosity may impinge on the deep aspect of the calcaneal tendon where it inserts onto the calcaneus (Haglund’s disease). It is often associated with a retrocalcaneal bursa, and symptoms are exacerbated by dorsiflexion of the ankle, because this movement increases the pressure within the bursa and causes impingement of calcaneus against the tendon insertion.
Deep group
The deep flexors of the calf include popliteus, which acts on the knee joint, and flexor hallucis longus, flexor digitorum longus and tibialis posterior, which all act on the ankle joint and joints of the foot.
Flexor hallucis longus
Flexor hallucis longus is described in Chapter 83. The tendon of flexor hallucis longus is described below.
INTRINSIC MUSCLES
The intrinsic muscles, i.e. those contained entirely within the foot, follow the primitive limb pattern of plantar flexors and dorsal extensors.
The plantar muscles in the foot can be divided into medial, lateral and intermediate groups. The medial and lateral groups consist of the intrinsic muscles of the hallux and minimus, respectively, and the central or intermediate group includes the lumbricals, interossei and short digital flexors. It is customary to group the muscles in four layers, because this is the order in which they are encountered during dissection. However, in clinical practice and in terms of function, the former grouping is often more useful.
Plantar muscles of the foot: first layer
This superficial layer includes abductor hallucis, abductor digiti minimi and flexor digitorum brevis (Fig. 84.19A). All three extend from the calcaneal tuberosity to the toes, and all assist in maintaining the concavity of the foot.
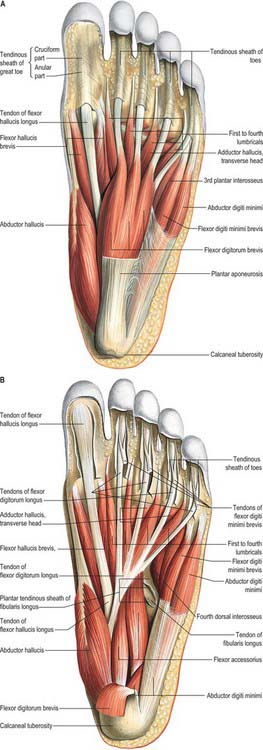
Abductor hallucis
Abductor hallucis arises principally from the flexor retinaculum, but also from the medial process of the calcaneal tuberosity, the plantar aponeurosis, and the intermuscular septum between this muscle and flexor digitorum brevis. The muscle fibres end in a tendon that is attached, together with the medial tendon of flexor hallucis brevis, to the medial side of the base of the proximal phalanx of the hallux. Some fibres are attached more proximally to the medial sesamoid bone of this toe. The muscle may also derive some fibres from the dermis along the medial border of the foot.
Abductor hallucis lies along the medial border of the foot and covers the origins of the plantar vessels and nerves (Fig. 84.20).
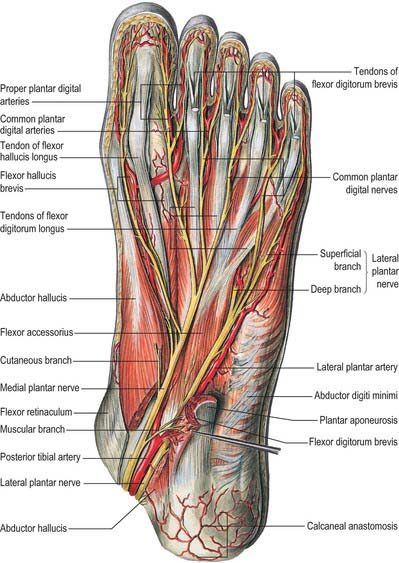
Abductor hallucis is supplied by the medial malleolar network, medial calcaneal branches of the lateral plantar artery (Fig. 84.9), the medial plantar artery (directly and via superficial and deep branches), the first plantar metatarsal artery and perforators from the plantar arterial arch.
Abductor hallucis is innervated by the medial plantar nerve, S1 and S2. Contraction of the muscle confirms an intact medial plantar nerve when the integrity of this nerve is in question.
Abductor hallucis is tested clinically by instructing the subject to resist a forcibly applied lateral deviation of the proximal phalanx.
The deep fascia has been implicated in chronic heel pain, because it can cause entrapment of the first branch of the lateral plantar nerve that lies immediately deep to it. The abductor hallucis fascia is strong and can be used in soft tissue augmentation following correction of hallux valgus. Rarely, persistent, exaggerated tonus in the muscle may be a cause of varus deformity of the foot, necessitating surgical intervention. An abductor hallucis flap is sometimes used for provision of soft tissue cover.
Flexor digitorum brevis
Flexor digitorum brevis arises by a narrow tendon from the medial process of the calcaneal tuberosity, from the central part of the plantar aponeurosis, and from the intermuscular septa between it and adjacent muscles. It divides into four tendons, which pass to the lateral four toes; the tendons enter digital tendon sheaths accompanied by the tendons of flexor digitorum longus, which lie deep to them. At the bases of the proximal phalanges, each tendon divides around the corresponding tendon of flexor digitorum longus; the two slips then reunite and partially decussate, forming a tunnel through which the tendon of flexor digitorum longus passes to the distal phalanx. The short flexor tendon divides again and attaches to both sides of the shaft of the middle phalanx. The way in which the tendons of flexor digitorum brevis divide and attach to the phalanges is identical to that of the tendons of flexor digitorum superficialis in the hand. The slip to a given toe may be absent, or it may be replaced by a small muscular slip from the long flexor tendon or from flexor accessorius. Conversely, the slip may be joined by a second, supernumerary slip.
Flexor digitorum brevis lies immediately deep to the central part of the plantar aponeurosis (Fig. 84.19A). Its deep surface is separated from the lateral plantar vessels and nerves by a thin layer of fascia.
Flexor digitorum brevis is supplied by the lateral and medial plantar arteries, the plantar metatarsal arteries and the plantar digital arteries to the lateral four toes.
Flexor digitorum brevis flexes the lesser toes at the proximal interphalangeal joint, with equal effect in any position of the ankle joint.
To test the action of flexor digitorum brevis, the examiner passively extends the distal interphalangeal joint and asks the individual to flex the toes at the proximal interphalangeal joint. Contracture of the flexor tendon can lead to toe deformities, and release or lengthening procedures may be required. The muscle belly is sometimes excised as a flap to cover a soft tissue defect.
Abductor digiti minimi
Abductor digiti minimi arises from both processes of the calcaneal tuberosity, from the plantar surface of the bone between them, from the plantar aponeurosis and from the intermuscular septum between the muscle and flexor digitorum brevis. Its tendon glides in a smooth groove on the plantar surface of the base of the fifth metatarsal and is attached, with flexor digiti minimi brevis, to the lateral side of the base of the proximal phalanx of the fifth toe; hence it is more a flexor than an abductor. Some of the fibres arising from the lateral calcaneal process usually reach the tip of the tuberosity of the fifth metatarsal (Figs 84.8B; 84.11) and may form a separate muscle, abductor ossis metatarsi digiti quinti. An accessory slip from the base of the fifth metatarsal is not infrequent.
Abductor digiti minimi lies along the lateral border of the foot, and its medial margin is related to the lateral plantar vessels and nerve (Fig. 84.20).
Abductor digiti minimi is supplied by the medial and lateral plantar arteries (Fig. 84.20; see Fig. 84.24A), the plantar digital artery to the lateral side of minimus, branches from the plantar arch, the fourth plantar metatarsal artery and end twigs of the arcuate and lateral tarsal arteries (Fig. 84.9).
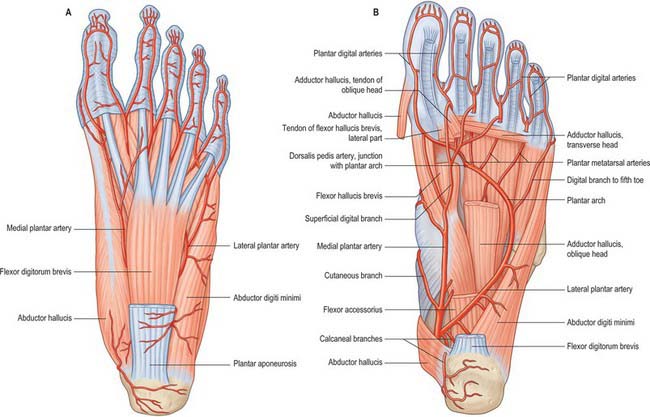
Plantar muscles of the foot: second layer
The second layer consists of flexor digitorum accessorius and four lumbrical muscles. The tendons of flexor hallucis longus and flexor digitorum longus run in the same plane as the muscles of the second layer (Fig. 84.19B): flexor hallucis longus and flexor digitorum longus are described in Chapter 83.
Flexor tendon sheaths
The terminations of the tendons of the long and short flexor muscles are contained in osseo-aponeurotic canals similar to those which occur in the fingers. These canals are bounded above by the phalanges, and below by fibrous bands, the digital fibrous sheaths, which arch across the tendons and attach on either side to the margins of the phalanges (Fig. 84.19B). Along the proximal and intermediate phalanges, the fibrous bands are strong, and the fibres are transverse (anular part); opposite the joints they are much thinner and the fibres decussate (cruciform part). Each osseo-aponeurotic canal has a synovial lining, which is reflected around its tendon; within this sheath, vincula tendinum are arranged as they are in the fingers.
Flexor digitorum accessorius
Flexor digitorum accessorius (Fig. 84.19B), also known as quadratus plantae, arises by two heads, with the long plantar ligament situated deeply in the interval between the two heads. The medial head is larger and more fleshy and is attached to the medial concave surface of the calcaneus, below the groove for the tendon of flexor hallucis longus. The lateral head is flat and tendinous and is attached to the calcaneus distal to the lateral process of the tuberosity, and to the long plantar ligament. The muscle belly inserts into the tendon of flexor digitorum longus at the point where it is bound by a fibrous slip to the tendon of flexor hallucis longus and where it divides into its four tendons.
The muscle is sometimes absent altogether. Its distal attachment to the tendons of flexor digitorum longus may vary, which means that the fourth and fifth long flexor tendons may, at times, fail to receive slips from the accessorius.
The medial plantar nerve passes medial to and the lateral plantar nerve passes superficial to flexor accessorius.
Lumbrical muscles
The lumbrical muscles are four small muscles (numbered from the medial side of the foot) which are accessory to the tendons of flexor digitorum longus (Fig. 84.19A,B). They arise from these tendons as far back as their angles of separation, each springing from the sides of two adjacent tendons, except for the first lumbrical, which arises only from the medial border of the first tendon. The muscles end in tendons that pass distally on the medial sides of the four lesser toes and are attached to the dorsal digital expansions on their proximal phalanges.
The lumbricals are intimately related to the tendons of flexor digitorum longus before the latter enter their corresponding fibrous flexor sheaths. The lumbricals remain outside the fibrous flexor sheaths and cross the plantar aspects of the deep transverse metatarsal ligaments before reaching the dorsal digital expansions.
The lumbricals are supplied by the lateral plantar artery and plantar arch, and by four plantar metatarsal arteries (four distal perforating arteries joined by three proximal perforating arteries). Their tendons are supplied by twigs from the dorsal digital arteries (and their parent dorsal metatarsal arteries) to the lateral four toes.
Plantar muscles of the foot: third layer
The third layer of the foot contains the shorter intrinsic muscles of the hallux and minimus, i.e. flexor hallucis brevis, adductor hallucis and flexor digiti minimi brevis (Fig. 84.21).
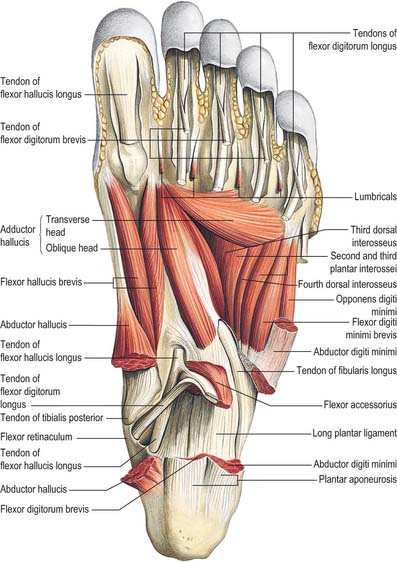
Flexor hallucis brevis
Flexor hallucis brevis has a bifurcate tendon of origin (Figs 84.19A,B; 84.21). The lateral limb arises from the medial part of the plantar surface of the cuboid, posterior to the groove for the tendon of fibularis longus, and from the adjacent part of the lateral cuneiform. The medial limb has a deep attachment directly continuous with the lateral division of the tendon of tibialis posterior, and a more superficial attachment to the middle band of the medial intermuscular septum. The belly of the muscle divides into medial and lateral parts, the twin tendons of which are attached to the sides of the base of the proximal phalanx of the hallux. The medial part blends with the tendon of abductor hallucis, and the lateral with that of adductor hallucis, as they reach their terminations.
A sesamoid bone usually occurs in each tendon near its attachment. Clinical problems with flexor hallucis brevis are usually related to the associated sesamoid bones. However, excision of both sesamoids leads to disruption of both tendons and a subsequent extension deformity at the first metatarsophalangeal joint: such surgery is therefore not recommended.
Accessory slips may arise proximally from the calcaneus or long plantar ligament. A tendinous slip may extend to the proximal phalanx of the second toe.
Flexor hallucis brevis lies on the underside of the first metatarsal shaft; abductor hallucis lies medially. The medial digital nerve to the hallux and the tendon of flexor hallucis longus pass to the hallux on its plantar surface. The medial plantar nerve lies more superficially on its lateral side.
Adductor hallucis
Adductor hallucis arises by oblique and transverse heads (Figs 84.19B; 84.21). The oblique head springs from the bases of the second, third and fourth metatarsal bones, and from the fibrous sheath of the tendon of fibularis longus. The transverse head, a narrow, flat fasciculus, arises from the plantar metatarsophalangeal ligaments of the third, fourth and fifth toes (sometimes only from the third and fourth), and from the deep transverse metatarsal ligaments between them. The oblique head has medial and lateral parts. The medial part blends with the lateral part of flexor hallucis brevis and is attached to the lateral sesamoid bone of the hallux. The lateral part joins the transverse head and is also attached to the lateral sesamoid bone and directly to the base of the first phalanx of the hallux. There is no phalangeal attachment for the transverse part of the muscle; fibres that fail to reach the lateral sesamoid bone are attached with the oblique part.
The transverse part of adductor hallucis is sometimes absent; part of the muscle may be attached to the first metatarsal, constituting an opponens hallucis; a slip may also extend to the proximal phalanx of the second toe.
Adductor hallucis lies plantar to the metatarsal shafts and interossei and the long and short toe flexors. The medial and lateral plantar arteries and nerves are superficial, and flexor hallucis brevis is proximal and medial.
Adductor hallucis is supplied by branches of the medial plantar artery, the lateral plantar artery, the plantar arch and the first to fourth plantar metatarsal arteries.
Flexor digiti minimi brevis
Flexor digiti minimi brevis arises from the medial part of the plantar surface of the base of the fifth metatarsal bone, and from the sheath of fibularis longus (Figs 84.19B; 84.21). It has a distal tendon that inserts into the lateral side of the base of the proximal phalanx of the minimus; this tendon usually blends laterally with that of abductor digiti minimi. Occasionally, some of its deeper fibres extend to the lateral part of the distal half of the fifth metatarsal bone, constituting what may be described as a distinct muscle, opponens digiti minimi.
The fifth metatarsal shaft lies on the deep surface of flexor digiti minimi brevis, the interossei lie medially and abductor digiti minimi is lateral. The most lateral branch of the lateral plantar nerve lies superficially and just medial to flexor digiti minimi brevis.
Flexor digiti minimi brevis is supplied by end twigs of the arcuate and lateral tarsal arteries, and the lateral plantar artery and its digital (plantar) branch to the lateral side of minimus.
Plantar muscles of the foot: fourth layer
The fourth layer of muscles of the foot consists of the plantar and dorsal interossei and the tendons of tibialis posterior and fibularis longus (tibialis posterior and fibularis longus are described in Ch. 83). The interossei resemble their counterparts in the hand except that, when describing adduction and abduction of the toes, the axis of reference is a longitudinal axis corresponding to the shaft of the 2nd metatarsal (unlike the hand where reference is made to the long axis of the third metacarpal).
Dorsal interossei
The dorsal interossei (Fig. 84.22A) are situated between the metatarsal bones. They consist of four bipennate muscles, each arising by two heads from the sides of the adjacent metatarsal bones. Their tendons are attached to the bases of the proximal phalanges and to the dorsal digital expansions. The first inserts into the medial side of the second toe; the other three pass to the lateral sides of the second, third and fourth toes.
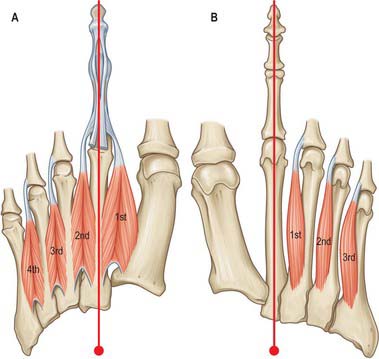
Between the heads of each of the three lateral muscles, there is an angular space through which a perforating artery passes to the dorsum of the foot. Between the heads of the first muscle, the corresponding space transmits the terminal part of the dorsalis pedis artery to the sole (see Fig. 84.1).
Dorsal interossei are supplied by the arcuate artery, lateral and medial tarsal arteries, the first to fourth plantar arteries and the first to fourth dorsal metatarsal arteries (receiving proximal and distal perforating arteries), and by the dorsal digital arteries of the lateral four toes.
Dorsal interossei are supplied by the deep branch of the lateral plantar nerve (S2 and S3), except that of the fourth interosseous space, which is supplied by the superficial branch of the lateral plantar nerve.
Plantar interossei
There are three plantar interossei (Fig. 84.22B). They lie below, rather than between, the metatarsal bones, and each is connected to one metatarsal bone only. They arise from the bases and medial sides of the third, fourth and fifth metatarsal bones, and insert into the medial sides of the bases of the proximal phalanges of the numerically corresponding toes, and into their dorsal digital expansions.
The plantar interossei lie plantar to the dorsal interossei and deep to the muscles of the third layer.
Plantar interossei are supplied by the lateral plantar artery, the plantar arch, the second to fourth plantar metatarsal arteries and the dorsal digital arteries of the lateral three toes.
Plantar interossei are supplied by the deep branch of the lateral plantar nerve (S2 and S3), except that of the fourth interosseous space, which is supplied by the superficial branch of the lateral plantar nerve.
Extensor muscles of the foot
Extensor digitorum brevis and extensor hallucis brevis
Extensor digitorum brevis (Fig. 84.2A) is a thin muscle which arises from the distal part of the superolateral surface of the calcaneus in front of the shallow lateral groove for fibularis brevis, from the interosseous talocalcaneal ligament, and from the deep surface of the stem of the inferior extensor retinaculum. It slants distally and medially across the dorsum of the foot, and ends in four tendons. The medial part of the muscle is usually a more or less distinct slip, ending in a tendon that crosses the dorsalis pedis artery superficially to insert into the dorsal aspect of the base of the proximal phalanx of the hallux; this slip is sometimes termed extensor hallucis brevis. The other three tendons attach to the lateral sides of the tendons of extensor digitorum longus for the second, third and fourth toes.
The muscle is subject to much variation, e.g. accessory slips from the talus and navicular, an extra tendon to the fifth digit, an absence of one or more tendons. It may be connected to the adjacent dorsal interosseous muscles.
The most medial tendon, that of extensor hallucis brevis, courses dorsomedially and passes superficial to the dorsalis pedis artery and the deep fibular nerve. The remaining three tendons pass obliquely deep to the corresponding tendons of extensor digitorum longus.
Extensor digitorum brevis is supplied by the anterior perforating branch of the fibular artery, the anterior lateral malleolar artery, lateral tarsal arteries, dorsalis pedis arteries, arcuate artery, the first, second and third dorsal metatarsal arteries, proximal and distal perforating arteries, and the dorsal digital arteries to the medial four toes (including the hallux).
Extensor digitorum brevis is supplied by the lateral terminal branch of the deep fibular nerve, L5, S1.
The muscle assists in extending the phalanges of the middle three toes via the tendons of extensor digitorum longus; in the hallux, it assists in extension of the metatarsophalangeal joint.
Laceration of extensor digitorum brevis leads to little in the way of functional impairment, because the long extensors can compensate for the loss of the muscle. The proximal part of the muscle can be used as interposition material to prevent bone fusion after resection of a calcaneonavicular bar (a common tarsal coalition). The muscle belly and tendon of extensor hallucis brevis serve as guides to the location of the dorsalis pedis artery and deep fibular nerve. The tendon of extensor hallucis brevis can be used as a local graft.
VASCULAR SUPPLY
ARTERIES
Dorsalis pedis artery
The dorsalis pedis artery is the dorsal artery of the foot (Figs 84.1; 84.23), and is the continuation of the anterior tibial artery distal to the ankle. It passes to the proximal end of the first intermetatarsal space, where it turns into the sole between the heads of the first dorsal interosseous to complete the plantar arch, and provides the first plantar metatarsal artery.
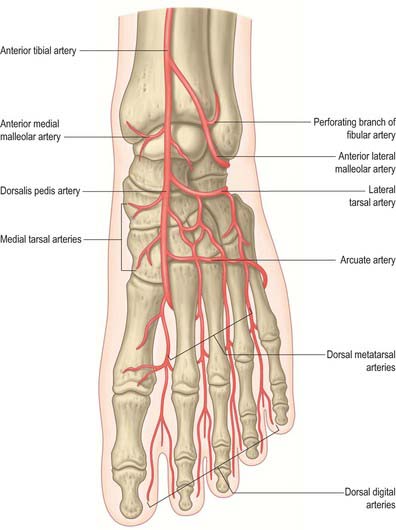
The artery may be larger than normal, to compensate for a small lateral plantar artery. It may be absent, in which event it is replaced by a large perforating branch of the fibular artery. It often diverges laterally from its usual route.
Relations
The dorsalis pedis artery crosses, successively, the talocrural articular capsule, talus, navicular and intermediate cuneiform and their ligaments; superficial to it are the skin, fasciae, inferior extensor retinaculum and, near its termination, extensor hallucis brevis. Medial to it is the tendon of extensor hallucis longus and lateral to it are the medial tendon of extensor digitorum longus and medial terminal branch of the deep fibular nerve. The tendons are useful landmarks in the planning of safe anatomical approaches in surgery of the ankle and foot.
Surface anatomy
The pulsation of the dorsal artery of the foot is palpable from the midpoint between the malleoli to the proximal end of the first intermetatarsal space.
Branches
The dorsalis pedis artery gives off the tarsal, arcuate and first dorsal metatarsal arteries.
There are two tarsal arteries, lateral and medial (Fig. 84.23). They arise as the dorsalis pedis artery crosses the navicular. The lateral runs laterally under extensor digitorum brevis; it supplies this muscle and the tarsal articulations, and anastomoses with branches of the arcuate, anterior lateral malleolar and lateral plantar arteries, and the perforating branch of the fibular artery. Two or three medial tarsal arteries ramify on the medial border of the foot and join the medial malleolar network.
The arcuate artery (Fig. 84.23) arises near the medial cuneiform, passes laterally over the metatarsal bases, deep to the tendons of the digital extensors, and anastomoses with the lateral tarsal and plantar arteries. It supplies the second to fourth dorsal metatarsal arteries, running distally superficial to the corresponding dorsal interossei, and divides into two dorsal digital branches for the adjoining toes in the interdigital clefts. Proximally, in the interosseous spaces, these branches receive proximal perforating branches from the plantar arch. Distally, they are joined by distal perforating branches from the plantar metatarsal arteries. The fourth dorsal metatarsal artery sends a branch to the lateral side of the fifth toe.
First dorsal metatarsal artery
The first dorsal metatarsal artery (Fig. 84.23) arises just before the dorsalis pedis artery enters the sole. It runs distally on the first dorsal interosseous and divides at the cleft between the first and second toes. One branch passes under the tendon of extensor hallucis longus and supplies the medial side of the hallux; the other bifurcates to supply the adjoining sides of the hallux and the second toe.
Cutaneous vessels from the dorsalis pedis artery
The dorsalis pedis artery and its first dorsal metatarsal branch give rise to small direct cutaneous branches that supply the dorsal foot skin between the extensor retinaculum and the first web space. This vessel provides the basis for a fasciocutaneous flap raised from this region, and which may be used to cover superficial defects elsewhere.
Plantar arch
The plantar arch (Fig. 84.24) is deeply situated, extending from the fifth metatarsal base to the proximal end of the first interosseous space. Convex distally, it is plantar to the bases of the second to fourth metatarsal bones and corresponding interossei, but dorsal to the oblique part of adductor hallucis.
Branches
The plantar arch gives off three perforating and four plantar metatarsal branches, and numerous branches that supply the skin, fasciae and muscles in the sole. Three perforating branches ascend through the proximal ends of the second to fourth intermetatarsal spaces, between the heads of dorsal interossei, and anastomose with the dorsal metatarsal arteries. Four plantar metatarsal arteries extend distally between the metatarsal bones in contact with the interossei (Fig. 84.24). Each divides into two plantar digital arteries, supplying the adjacent digital aspects. Near its division, each plantar metatarsal artery sends a distal perforating branch dorsally to join a dorsal metatarsal artery. The first plantar metatarsal artery springs from the junction between the lateral plantar and dorsalis pedis arteries, and sends a digital branch to the medial side of the hallux. The lateral digital branch for the fifth toe arises directly from the lateral plantar artery near the fifth metatarsal base. Haemorrhage from the plantar arch is difficult to stem, because of the depth of the vessel and its important close relations.
Posterior tibial artery
Before the posterior tibial artery divides into its two main terminal branches, it gives off a communicating branch which runs posteriorly across the tibia approximately 5 cm above its distal end, deep to flexor hallucis longus, and joins a communicating branch of the fibular artery; calcaneal branches, which arise just proximal to the termination of the posterior tibial artery, pierce the flexor retinaculum and supply the skin and fat behind the calcaneal tendon; the artery of the tarsal canal. The terminal branches of the posterior tibial artery are the medial and lateral plantar arteries.
Branches
The medial plantar artery is the smaller terminal branch of the posterior tibial artery (Fig. 84.24). It arises midway between the medial malleolus and the medial calcaneal tubercle and passes distally along the medial side of the foot, with the medial plantar nerve lateral to it. At first deep to abductor hallucis, it runs distally between abductor hallucis and flexor digitorum brevis, supplying both. Near the first metatarsal base, when its calibre is already diminished as a result of supplying numerous muscular branches, it is further diminished by a superficial stem. It passes to the medial border of the hallux, where it anastomoses with a branch of the first plantar metatarsal artery. Its superficial stem then trifurcates and supplies three superficial digital branches that accompany the digital branches of the medial plantar nerve and join the first to third plantar metatarsal arteries.
The lateral plantar artery is the larger terminal branch of the posterior tibial artery (Fig. 84.24). It passes distally and laterally to the fifth metatarsal base; the lateral plantar nerve is medial. The plantar nerves lie between the plantar arteries. Turning medially, with the deep branch of the nerve, it gains the interval between the first and second metatarsal bases, and unites with the dorsalis pedis artery to complete the plantar arch. As it passes laterally, it is first between the calcaneus and abductor hallucis, then between flexor digitorum brevis and flexor accessorius. Running distally to the fifth metatarsal base, it passes between flexor digitorum brevis and abductor digiti minimi and is covered by the plantar aponeurosis, superficial fascia and skin.
Muscular branches supply the adjoining muscles. Superficial branches emerge along the intermuscular septum to supply the skin and subcutaneous tissue over the lateral part of the sole. Anastomotic branches run to the lateral border, and join branches of the lateral tarsal and arcuate arteries. Sometimes, a calcaneal branch pierces abductor hallucis to supply the skin of the heel.
Perforator flaps in the ankle and foot region
The arteries around the ankle and in the foot are the medial and lateral calcaneal artery, medial and lateral plantar artery and the dorsalis pedis artery. In the foot the two plantar arteries with communicating arteries from dorsalis pedis give out multiple small perforators. In the sole of the foot the perforators emerge on either side of the plantar aponeurosis, and also pass through it, to supply the skin. ‘Island flaps’ from these perforators may be advanced to reconstruct small defects in the weight bearing area. The skin of the sole of the foot is highly specialized and therefore ideally defects in this region should be reconstructed using local skin.
DEEP AND SUPERFICIAL VENOUS SYSTEMS IN THE FOOT
Plantar digital veins arise from plexuses in the plantar regions of the toes. They connect with dorsal digital veins to form four plantar metatarsal veins which run proximally in the intermetatarsal spaces and connect via perforating veins with dorsal veins, then continue to form the deep plantar venous arch which is situated alongside the plantar arterial arch. From this venous arch, medial and lateral plantar veins run near the corresponding arteries and, after communicating with the great and small saphenous veins, form the posterior tibial veins behind the medial malleolus.
The principal named superficial veins are the great and small saphenous. Their numerous tributaries are mostly (but not wholly) unnamed; named vessels will be noted (see Fig. 79.9A,B). As in the upper limb, the vessels will be described centripetally from peripheral to major drainage channels.
Dorsal digital veins receive rami from the plantar digital veins in the clefts between the toes and then join to form dorsal metatarsal veins, which are united across the proximal parts of the metatarsal bones in a dorsal venous arch. Proximal to this arch, an irregular dorsal venous network receives tributaries from deep veins and is continuous proximally with a venous network in the leg. At each side of the foot, this network connects with medial and lateral marginal veins, which are both formed mainly by veins from more superficial parts of the sole. In the sole, superficial veins form a plantar cutaneous arch across the roots of the toes and also drain into the medial and lateral marginal veins. Proximal to the plantar arch there is a plantar cutaneous venous plexus, especially dense in the fat of the heel. It connects with the plantar cutaneous venous arch and other deep veins, but drains mainly into the marginal veins. The veins of the sole are an important part of the lower limb ‘venous pump’ system aiding propulsion of blood up the limb. Intermittent foot compression devices are available to enhance this flow and so reduce the risk of deep vein thrombosis during periods of increased risk, e.g. after surgery.
INNERVATION OF THE FOOT
Tibial nerve
The branches of the tibial nerve that innervate structures in the ankle and foot are articular, muscular, sural, medial calcaneal and medial and lateral plantar nerves. The course and distribution of the tibial nerve in the calf are described in Chapter 83.
Medial calcaneal nerve
The medial calcaneal nerve arises from the tibial nerve and perforates the flexor retinaculum to supply the skin of the heel and medial side of the sole.
Medial plantar nerve
The medial plantar nerve is the larger terminal division of the tibial nerve, and lies lateral to the medial plantar artery. From its origin under the flexor retinaculum, it passes deep to abductor hallucis, then appears between it and flexor digitorum brevis, gives off a medial proper digital nerve to the hallux, and divides near the metatarsal bases into three common plantar digital nerves.
Cutaneous branches pierce the plantar aponeurosis between abductor hallucis and flexor digitorum brevis to supply the skin of the sole of the foot. Muscular branches supply abductor hallucis, flexor digitorum brevis, flexor hallucis brevis and the first lumbrical. The first two arise near the origin of the nerve and enter the deep surfaces of the muscles. The branch to flexor hallucis brevis is from the hallucal medial digital nerve, and that to the first lumbrical from the first common plantar digital nerve. Articular branches supply the joints of the tarsus and metatarsus.
Three common plantar digital nerves pass between the slips of the plantar aponeurosis, each dividing into two proper digital branches. The first supplies adjacent sides of the hallux and second toe and the second supplies adjacent sides of the second and third toes; the third supplies adjacent sides of the third and fourth toes, and also connects with the lateral plantar nerve. The first gives a branch to the first lumbrical. Each proper digital nerve has cutaneous and articular branches: near the distal phalanges a dorsal branch supplies structures around the nail, and the termination of each nerve supplies the ball of the toe.
The common digital branches of the medial plantar nerve are distributed in a manner similar to those of the median nerve, as are the motor branches of the two nerves. In the hand, the median nerve supplies abductor and flexor pollicis brevis, opponens pollicis and the first and second lumbricals. An opponens is absent in the foot, but abductor hallucis, flexor hallucis brevis and the first lumbrical are all supplied by the medial plantar nerve. Flexor digitorum brevis and flexor digitorum superficialis (innervated by the median nerve) correspond, but the innervation of the second lumbrical of the foot does not mirror the arrangement in the hand.
Lateral plantar nerve
The lateral plantar nerve supplies the skin of the fifth toe, the lateral half of the fourth toe, and most of the deep muscles of the foot (Fig. 84.20). Its distribution therefore closely resembles that of the ulnar nerve in the hand. It passes laterally forwards medial to the lateral plantar artery, towards the tubercle of the fifth metatarsal. Next, it passes between flexores digitorum brevis and accessorius, and ends between flexor digiti minimi brevis and abductor digiti minimi by dividing into superficial and deep branches. Before division, it supplies flexor digitorum accessorius and abductor digiti minimi and gives rise to small branches that pierce the plantar fascia to supply the skin of the lateral part of the sole (Fig. 84.25). The superficial branch splits into two common plantar digital nerves: the lateral supplies the lateral side of the fifth toe, flexor digiti minimi brevis and the two interossei in the fourth intermetatarsal space; the medial connects with the third common plantar digital branch of the medial plantar nerve and divides into two to supply the adjoining sides of the fourth and fifth toes. The deep branch accompanies the lateral plantar artery deep to the flexor tendons and adductor hallucis and supplies the second to fourth lumbricals, adductor hallucis and all the interossei (except those of the fourth intermetatarsal space). Branches to the second and third lumbricals pass distally deep to the transverse head of adductor hallucis, and curl round its distal border to reach them.
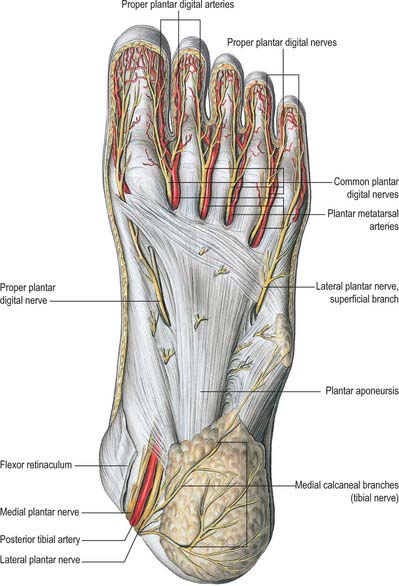
Nerve entrapment syndromes in the foot
All nerves of the foot can be affected by entrapment, leading classically to a burning sensation in the distribution of that nerve. Tarsal tunnel syndrome is much less common than carpal tunnel syndrome. The flexor retinaculum may compress the tibial nerve or either of its branches (medial and lateral plantar nerves). However entrapment at this level is most commonly due to a space-occupying lesion, e.g. ganglion, or compression by a leash of vessels or the deep fascia associated with abductor hallucis. Compression of the first branch of the lateral plantar nerve by the deep fascia of abductor hallucis can lead to heel pain. The medial plantar nerve can be compressed at the ‘knot of Henry’: this is usually related to jogging. The ‘knot of Henry’ is the anatomical landmark where the tendon of flexor hallucis longus crosses deep to the tendon of flexor digitorum longus, to reach its medial side in the sole of the foot. The superficial fibular nerve can be damaged in severe inversion injuries of the ankle, and the deep fibular nerve is sometimes compressed by osteophytes in the region of the second tarsometatarsal joint. Sural nerve entrapment is usually not due to compression by fascial elements; rather, it is secondary to distal fibular trauma and subsequent scar formation around the nerve. Entrapment of the common digital nerve as it passes under the intermetatarsal ligament of the third (or less commonly the second) webspace can result in a Morton’s neuroma, which is probably the most common form of nerve entrapment in the foot.
BIOMECHANICS OF STANDING, WALKING AND RUNNING
PLANES OF MOTION
Much confusion surrounds the descriptive terms for movement in the foot and ankle. Plantar flexion and dorsiflexion refer to movement in the sagittal plane and occur principally, but not exclusively, at the ankle, metatarsophalangeal and interphalangeal joints. Inversion is tilting of the plantar surface of the foot towards the midline, and eversion is tilting away from the midline. This is motion in the coronal plane and takes place principally in the talocalcaneal and transverse tarsal joints. Adduction is movement of the foot towards the midline in the transverse plane; abduction is movement away from the midline. This movement occurs at the transverse tarsal joints and, to a limited degree, the first tarsometatarsal joint and the metatarsophalangeal joints.
Supination describes a three-dimensional movement and is a combination of adduction, inversion and plantar flexion. Pronation is the opposite motion, i.e. a combination of abduction, eversion and dorsiflexion. Pronation and supination are usually better terms than eversion and inversion, as the latter rarely occur in isolation and the former describe the ‘compound’ motion that usually occurs.
Active movements occur at the ankle, talocalcaneonavicular and subtalar joints. Movements at the ankle joint are almost entirely restricted to dorsi- and plantar flexion, but slight rotation may occur in plantar flexion. The ranges of movement at the talocalcaneonavicular and subtalar joint are greater: inversion and eversion mainly occur here.
STANDING
Humans are bipedal: we stand and walk with an erect trunk and knees which are almost straight. Moreover, we are plantigrade, i.e. we set the whole length of the foot down on the ground, whereas most medium to large mammals are digitigrade, i.e. they stand and walk on their toes, and ungulates stand on hooves on the tips of their toes.
When we stand motionless, the ground exerts a force on the feet called the ‘ground reaction force’, which is equal and opposite to body weight. The latter acts at the ‘centre of gravity’, a point a little above the hip joints and anterior to the vertebral column. The force on the ground is centred vertically below it, at the centre of pressure. When we stand comfortably, the centre of pressure is usually midway between the insteps of our two feet. We do not remain absolutely still, but sway slightly. As we age, our postural control deteriorates, which is a factor in the development of unsteadiness and risk of falling.
When considering the distribution of load in the lower limb, a knowledge of limb alignment is important. The ‘anatomic axis’ is produced by lines passing along the shafts of the femur and tibia, and is therefore angled at the knee by 5° to 7°. The ‘mechanical axis’ is a straight line connecting the centres of the femoral head and ankle. Exactly where the line passes through the knee has yet to be defined: for convenience it is usually taken to pass through the centre, but may well lie more medially. For the tibia, the lines of the mechanical and anatomic axes coincide. Whilst the weight of the body acts in a vertical line from the centre of gravity to the ground, the line of the mechanical axis of the lower limbs is inclined 3° to this in normal stance, with the ankles closer than hips.
In the sagittal plane, body weight acts along a line which passes a few centimetres anterior to the tibiotalar joint, exerting a moment that must be balanced by the plantar flexor muscles. Some electromyographic investigations of standing have found activity mainly in soleus, but others have found moderate activity in gastrocnemius as well. In either case it seems likely that the activity is mainly in type I (slow oxidative) fibres, since these can maintain tension more economically than faster fibres could do under the same conditions. Soleus contains mostly type I fibres, and gastrocnemius contains approximately 50%.
In normal standing, the weight of the body above the knee joint acts slightly anteriorly to the axes of the knees: it exerts a moment that is balanced passively by tension in the ligaments and posterior capsule which prevent hyperextension. If the knees are kept straight, there is little or no electrical activity in the hamstrings and only moderate levels in the quadriceps. Generally, the combined weight of the trunk, arms and head seems to act slightly behind the hip joints, exerting a small moment that is balanced largely by iliopsoas. Thus a relaxed stance, with trunk erect and knees straight, requires very little activity in thigh muscles. This demands low energy consumption and permits long periods of standing. In single leg stance the weight of the upper parts of the body exerts a moment about the supporting hip which must be balanced by the abductor muscles, especially tensor fasciae latae and glutei medius and minimus. Damage to the glutei or their nerve supply, e.g. during hip surgery, is a potent cause of a limp.
PROPULSION
The contraction of tibialis posterior, gastrocnemius and soleus is the chief factor responsible for propulsion in walking, running and jumping. The propulsive action of these calf muscles is enhanced by arching of the foot and flexion of the toes. In walking, the weight of the foot is taken successively on the heel, lateral border and the ball of the foot. The last part of the foot to leave the ground is the anterior pillar of the medial longitudinal arch and the medial three toes. In the act of running, the heel does not touch the ground, but the point of take-off is still the anterior pillar of the medial longitudinal arch. As the heel leaves the ground, the toes gradually extend. Extension, of the hallux particularly, tightens the plantar aponeurosis and thus heightens the arch. At the same time, flexor hallucis longus and flexor digitorum longus elongate, which increases their subsequent contraction. The long and short toe flexors increase the force of take-off by exerting pressure on the ground. The most important muscle in this respect is flexor hallucis longus, which is strongly assisted by the short toe flexors. The lumbricals provide a balancing action to the long toe flexors and prevent buckling of the toes during the toe-off phase of gait.
Walking
In walking, each foot is on the ground (stance phase) for approximately 60% of the stride, and off the ground (swing phase) for approximately 40% (Fig. 84.26). Thus single-support phases (one foot on the ground) alternate with double-support phases (two feet on the ground). The knee is straight at heel strike and remains nearly straight (10–30°) for most of the stance phase of that leg, bending more only immediately before toe-off. During the swing phase the knee flexes to a maximum of 60° at mid-swing.
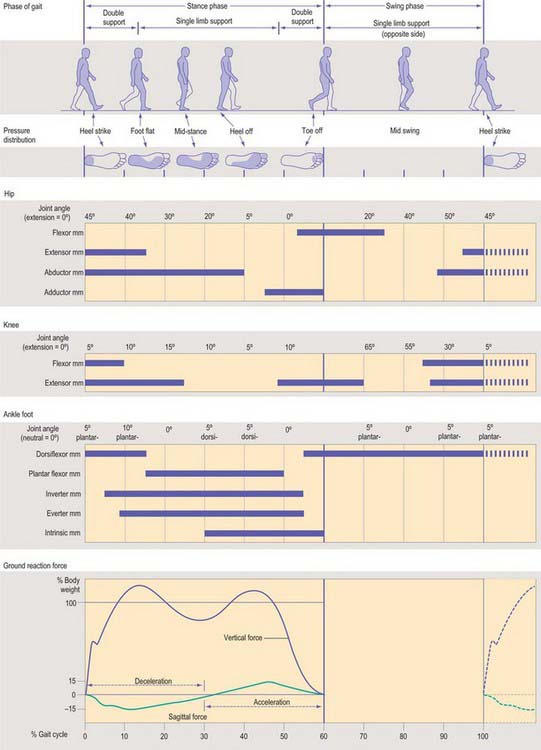
Fig. 84.26 The events that occur during the different phases of a normal gait cycle. Depicted are: distribution of pressure on the plantar surface of the foot; changes in the angles of hip, knee and ankle joints, together with activity in the corresponding muscle groups; and vertical and horizontal (sagittal plane) components of the ground reaction force during stance phase. (Chart collated from various sources by Michael Gunther, Department of Human Anatomy and Cell Biology, University of Liverpool.)
Stance phase starts with ‘heel strike’. With the foot still planted in front of the body, ‘foot flat’ is reached, and becomes ‘mid-stance’ when the body comes to be directly above the planted foot. The heel then rises as the contralateral foot makes contact with the ground (the ‘double stance’ phase). The last event of stance is ‘toe off ’ when the ‘swing phase’ starts. Early in the stance phase, while it is ‘foot flat’ in front of the trunk, the foot pushes downwards and forwards on the ground, decelerating the body as well as supporting it. Later, when the foot is behind the trunk, it pushes downwards and backwards, re-accelerating the body (Fig. 84.26). The speed, and therefore the kinetic energy, of the body passes through a maximum in each double-support phase, and a minimum in each single-support phase.
The height of the centre of gravity, and therefore the potential energy of the body, also fluctuates. This is inevitable if the knee is kept nearly straight, making the hip move in a near-circular arc about the ankle of the supporting foot. The vertical component of the total force exerted by both feet on the ground in the double-support phase is greater than body weight, giving the body an upward acceleration. The vertical component of the ground force during the single-support phase is less than body weight, giving the body a downward acceleration. The force fluctuations have to be larger at higher speeds, to give the same vertical movement in less time.
During walking, the potential energy of the body is high when its kinetic energy is low, and vice versa. Energy can thus be swapped back and forth between the two forms, as in a swinging pendulum. A frictionless pendulum would continue swinging forever with no fresh input of energy. In walking, the exchange of potential and kinetic energy is less perfect, and the leg muscles have to do work to replace the energy which is lost at each impact of a foot with the ground. Even so, much less work is needed because of the pendulum principle. A second pendulum effect can be seen in the forward swing of the legs. If the leg were a rigid rod swinging from a fixed pivot, it would swing forward too slowly for normal walking: because it is free to bend at the knee, and because the hip from which it swings rises and falls, the leg swings forward faster with very little need for muscular work.
These pendulum effects ensure that relatively little work is needed to drive the fluctuations of kinetic and potential energy which occur in walking. The additional work needed to overcome air resistance and joint friction is very small, because we travel slowly and have well-lubricated joints. Human walking is economical of energy. Measurements of oxygen consumption show that 70-kg men use only 230 joules of metabolic energy for every metre they walk at their most economical speed (approximately 1.3 metres per second), which is 140 J/m more than they would use if they stood still. Running uses 260 J/m more than standing. Walking is economical only near the optimum speed, and the energy cost of fast walking overtakes that of running at 2 m/s, at which speed adults usually break into a run. Children, with their shorter legs, start running at lower speeds: the theoretical expectation is that the critical speed should be proportional to the square root of leg length.
The function of arm swinging in walking is not fully understood but it aids postural stability. For further discussion of standing and walking see Alexander (1992).
Development of walking
The average child sits at 6 months, crawls at 9 months, walks with support at 12 months, and without support at 18 months. The characteristic early gait matures rapidly and is similar to that of the adult by 3 years. Some minor changes occur up to 7 years which are largely a reflection of neurological development, but are also related to stature. Early gait is jerky, unsteady, and wide-based. The arms are held abducted at the shoulder and initially flexed at the elbows. Initial ground contact varies, and heel-toe, whole foot, and toe-heel are all possible. Generally a plantarflexed posture is adopted which contrasts with the adult pattern. In adults, heel strike is accompanied by a straight knee which then flexes. A child strikes the ground with a flexed knee which is then extended in response to weight-bearing, and a short time is spent in single leg stance (Fig. 84.27). Maturation is associated with diminution of base width, and increase in step length and velocity. The earliest changes are development of heel-strike, knee flexion during stance, and reciprocal upper limb swing. The period of single leg stance increases.
Fig. 84.27 Development of a mature gait. Top: 1-year-old. Note the flexed elbows and lack of arm swing. The foot is plantar flexed at contact. Middle: 3-year-old. Arm swing is now present as is heel strike. Bottom: 6-year-old. There is now an adult-type gait.
(By permission from Benson MKD, Fixsen JA, MacNicol MF (eds) 2001 Development of a mature gait. In: Children’s Orthopaedics and Fractures, 2nd edn. Edinburgh: Churchill Livingstone.)
Running
Walking involves dual-support phases, but in running each foot is on the ground for 40% (jogging) to 27% (sprinting) of the stride, so there is an aerial phase, the ‘double float’ phase, when neither foot is on the ground. The faster the subject runs, the shorter the stance phase: world-class sprinters spend approximately 22% of the gait cycle in stance. During each aerial phase the body rises and then falls under gravity, which means that its height and potential energy are maximal in the middle of this phase and minimal at mid-stance, when, in marked contrast to walking, the knee of the supporting leg bends. Walking has been described as ‘controlled falling’, as the centre of gravity falls from a peak in mid-stance to a low during double support. The analogy that is used for running is an individual on a ‘pogo stick’. The runner propels himself from a low point in mid-stance to a peak during double float. During walking, efficiency is maintained by the interchange of potential and kinetic energy. During running, when potential and kinetic energies are in phase, efficiency is maintained in two ways: storage and release of potential energy in the elastic structure of tendons, and transfer of energy between limb segments via muscles which cross two joints (e.g. rectus femoris and hamstrings). The changes in muscle belly lengths are relatively slight during running. The muscles are acting as tensioners of the tendons, indeed, most of the change of length is produced by the stretch and recoil of the tendons. It has been estimated that, of the kinetic and potential energy lost and regained in each stance phase, 35% is stored temporarily as elastic strain energy in the calcaneal tendon, and 17% in the ligaments of the arch of the foot. Together these springs approximately halve the work required from the muscles.
A tendon is capable of withstanding stresses (force per unit cross-sectional area) of at least 100 N/mm2 (10 kg force/mm2). It stretches elastically by approximately 8% before breaking, and its recoil returns 93% of the energy used to stretch it. This high energy return is important, not only because it reduces the work required from the muscles, but also because the lost mechanical energy becomes heat: leg tendons with poor energy return would overheat in running, and be damaged. The calcaneal tendon is the most important ‘spring’ in the leg. Most runners strike the ground first with the heel, and the centre of pressure moves rapidly forward to the distal heads of the metatarsals, where it remains for most of the stance phase. A large force is then required in the calcaneal tendon to balance the moments about the tibiotalar joint. For a 70-kg man running at middle-distance speed, the peak force in the tendon is 5000 N (0.5 tonne force), which is enough to stretch it by approximately 6%. The parts of the tendon that run alongside the muscle bellies of gastrocnemius and soleus must be stretched, as well as the free distal part.
The mean vertical force on the ground during a complete stride cycle of any gait must equal body weight. If the foot is on the ground for only a small fraction of the stride, the forces must then be high. Peak vertical forces on one foot are approximately 1.0 times body weight in walking, 2.5 times in jogging, and 3.5 times in sprinting. Leg muscles have to exert much larger forces in running than in walking because the ground reaction forces are higher and also because, for a given ground force, flexion of the knee increases the moments about it. The main sources of power are the gastrocnemius–soleus complex at push-off; quadriceps femoris after ‘stance phase reversal’ (when the lower limb muscles change from a decelerating role to one of generating power, the generation phase); the hip extensors during late swing and early stance; the hip abductors just after stance phase reversal.
As in walking, the ground force acts more or less in line with the leg, so the body is decelerated and reaccelerated during each stance phase. The stance phase starts with deceleration and absorption of energy. Power is generated after stance phase reversal as the limb pushes up with the knee extending and foot plantarflexed, and this continues in the swing phase as the limb is accelerated forward. Once the limb is ahead of the trunk, the final phase of swing phase absorption is initiated, during which the limb is decelerated.
The ground force acts upward on the metatarsal heads, and the calcaneal tendon pulls upwards on the calcaneus. The necessary balancing reaction occurs at the ankle, where the tibia presses downwards on the talus. Together these three forces flatten the longitudinal arch of the foot, forcing the ankle 10 mm nearer to the ground than it would be if the foot was rigid. Mechanical tests on amputated feet have shown that the foot is a reasonably good spring, giving an energy return of nearly 80%. The plantar aponeurosis, long and short plantar ligaments and the plantar calcaneonavicular ligament are all involved in the spring action: they are predominantly collagenous, and presumably have elastic properties similar to those of tendon.
A runner’s foot is still moving, typically at about 1.5 m/s, when the heel hits the ground. The impact is cushioned by the subcalcaneal foot pad, supplemented (when shoes are worn) by the compliance of the heel of the shoe. On contact the foot is supinated, which ‘locks’ the foot as a rigid structure, and is ideal for shock absorption. Immediately after heel strike the trunk progresses over the foot: passive pronation is initiated which makes the foot more flexible and therefore able to adapt to the running surface. As the foot approaches push-off, it becomes supinated again and as a consequence stiffens, which provides an excellent lever. These changes in foot posture cause rotational changes in the leg, and these in turn impact on the knee.
Eighty per cent of runners heel strike, and 20% have a mid foot strike. In sprinting the forefoot makes contact first to provide the greatest leverage. The knees never fully extend during running: at first contact, the knee is 25° flexed, increasing to a maximum during stance of 45° at stance phase reversal, and decreasing to 20° at toe-off. In running, the swing phase knee flexion increases 90° at the end of swing phase generation (up to 125° in sprinting), then decreases until contact with the ground again. (All values are approximate.)
Movements of the foot
With the foot on the ground, body weight causes some supination and flattening of the longitudinal arches; about one-third of the weight borne by the forefoot is taken by the head of the first metatarsal. When a resting position becomes active, as occurs on starting to walk, the foot is pronated by muscular effort, and the first metatarsal is depressed (the second less so), which accentuates the longitudinal arch to its maximum height (Hicks 1953). Similar changes can be imposed on a weight-bearing foot by active lateral femoral rotation, which is transmitted through the tibia to the talus and entails passive supination of the foot. Medial femoral rotation has an opposite effect. When the foot is grounded and immobile, muscles which move it when it is freely suspended may exert effects on the leg, e.g. the dorsiflexors can then pull the leg forwards at the ankle joint.
The foot has two major functions: to support the body in standing and progression, and to lever it forwards and absorb shock in walking, running and jumping. To fulfil the first function, the pedal platform must be able to spread the stresses of standing and moving and be pliable enough to accommodate walking or running over uneven and sloping surfaces. To fulfil the second function, the foot must be transformable into a strong adjustable lever in order to resist inertia and powerful thrust: a segmented lever can best meet such stresses if it is arched.
In infants and young children, fatty, connective tissue on the plantar aspect may give the foot a flat appearance, indeed, soft tissues modify its appearance to varying degrees at all ages. However, the skeleton of the human foot is normally arched, and the sole of the foot is usually visibly concave. This curvature is customarily analysed into longitudinal and transverse arches. These arches vary individually in height, especially the longitudinal in its medial part. Since they are dynamic, their heights also differ in different phases of activity.
The medial longitudinal arch contains the calcaneus, talus, navicular, cuneiform and medial three metatarsal bones. Its summit, at the superior talar articular surface, takes the full thrust from the tibia and passes it backwards to the calcaneus, and forwards through the navicular and cuneiforms to the metatarsals. When the foot is grounded, these forces are transmitted through the three metatarsal heads and calcaneus (especially its tuberosity). The medial arch is higher, and more mobile and resilient than the lateral arch; its flattening progressively tightens the plantar calcaneonavicular ligament and plantar fascia. The lateral arch is adapted to transmit weight and thrust rather than to absorb such forces: the long plantar and plantar calcaneocuboid ligaments tighten as it flattens.
The lateral arch makes contact with the ground more extensively than the medial arch. As the foot flattens, an increasing fraction of load traverses soft tissues which are inferior to the entire arch. The whole lateral border usually touches the ground, whereas the medial border does not. However the medial border is visibly concave, usually even in standing, which explains the familiar outline of human footprints (though this varies with the position of the feet (Fig. 84.28), the development of associated soft tissues, and the nature of the surface). In any activity, as soon as the heel rises, the toes are extended and muscular structures (including the plantar aponeurosis) tighten up in the sole, accentuating the longitudinal arches. It has been suggested that tension diminishes in the deeper plantar ligaments in this phase.
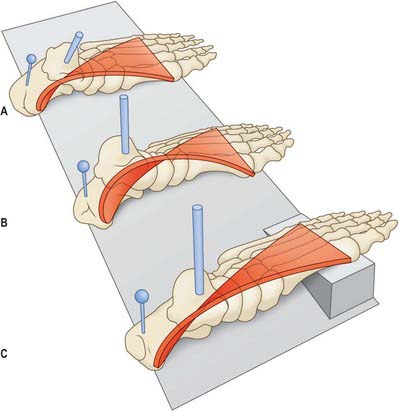
Fig. 84.28 The concept of the foot skeleton as a twisted plate which may be untwisted (supination) or further twisted (pronation) during the maintenance of a plantigrade stance in various positions of the foot. (Based upon MacConaill 1945, 1950.) A, The foot skeleton in supination, as in standing with the feet widely separated. Note the marked medial tilting of the talus and, to a lesser degree, of the calcaneus and the depression of the medial longitudinal arch. B, Relative pronation of the foot, as in standing with the feet close together. C, Supination of the foot when standing on an inclined surface; if the position of the wedge had been reversed, the foot skeleton would, of course, approach maximal pronation.
The sole is transversely concave, both in skeletal form and usually in external appearance, and serial transverse arches are most developed inferior to the metatarsus and adjoining tarsus. Transmission of force occurs at the metatarsal heads, to some degree along the lateral border of the foot, and through subjacent soft tissues.
In standing, with only body weight to support, both the intrinsic and extrinsic muscles appear to relax. If the longitudinal arches are allowed to sink as a result of muscular relaxation, the plantar ligaments tie the bones into an arched form. The medial arch is more elevated when the feet are together than when they are apart, i.e. inversion with supination increases as the feet are separated. This medial sag can be countered by voluntary contraction of muscles such as tibialis anterior. Pronation and supination ensure that in standing, whatever the position of the feet, a maximal weight-bearing area is grounded, from the metatarsal heads along the lateral border of the foot to the calcaneus. The twist imparted by pronation (which is partly undone in supination) prompts the likening of the foot to a twisted but resilient plate (Fig. 84.28), where adequate ground contact was ensured whatever the angle between the foot and leg, and adaptable resilience was imparted in standing and progression.
Alexander R McN. The Human Machine. London: Natural History Museum Publications, 1992.
Barclay-Smith E. The astragalo-calcaneo-navicular joint. J Anat Physiol. 1896;30:390-412.
This paper gave the first description of the astragalo-calcaneo-navicular joint and explained the importance of the joint complex in terms of hindfoot motion..
Coleman S, Chesnut W. A simple test for hindfoot flexibility in the cavovarus foot. Clin Orthop. 1977;123:60-62.
Cummins JE, Anson JB, Carr WB, Wright RR, Hauser DWE. The structure of the calcaneal tendon (of Achilles) in relation to orthopedic surgery with additional observations on the plantaris muscle. Surg Gynecol Obstet. 1946;83:107-116.
Gardner E, Gray DJ. The innervation of the joints of the foot. Anat Rec. 1968;161:141-148.
Harris RI, Beath T. Etiology of peroneal spastic flat foot. J Bone Joint Surg [Br]. 1948;30:624-634.
Hicks JH. The mechanics of the foot II. The plantar aponeurosis and the arch. J Anat. 1953;88:25-30.
Jones FW. Structure and Function as Seen in the Foot, 2nd edn., London: Baillière, Tindall and Cox; 1949:63-65.
One of the classic texts on the foot..
Kapandji IA. The Physiology of the Joints. Annotated Diagrams of the Mechanics of the Human Joints, 2nd edn (3 vols translated by LH Honoré: Vol 1 Upper Limb. Vol 2 Lower Limb. Vol 3 The Trunk and the Vertebral Column.). Edinburgh: Churchill Livingstone, 1970–1974.
Sarrafian SK. Anatomy of the Foot and Ankle, Descriptive, Topographic, Functional, 2nd edn. Philadelphia: Lippincott, 1993.
Taylor GI, Razaboni RM. Arteries of the Muscles of the Extremities and the Trunk. St Louis: Quality Medical Publishing Inc, 1994.
Wildenauer F. Die Blutversorgung des Talus. Z Anat Entwicklungsgesch. 1950;115:32-36.
The first comprehensive account of talar blood supply and the identification of the important artery of the tarsal canal..