Chapter 14 The Visual System
1. The eye’s anatomy is adapted to the eye’s role as a visual receptor organ.
2. Through the process of accommodation, the lens changes shape to focus images from various distances onto the retina.
3. The vertebrate retina consists of five major cell types.
4. The fovea solves a distortion problem found in other areas of the retina.
5. Pigment behind the retina either absorbs or reflects light, depending on the animal’s habits.
6. Photoreception and transduction of light occur in the rods and cones.
7. The response of the photoreceptor to light is synaptically transmitted to the ganglion cells by the bipolar cells.
8. The electroretinogram records the electrical response of the retina to a flashing light.
9. Ganglion cell axons transmit action potentials to the visual cortex by way of the lateral geniculate nucleus.
10. The diameter of the pupil is controlled by the autonomic nervous system.
11. The retina, optic nerve, and autonomic nerve supply controlling the pupil can be tested with a flashlight.
The eyes are complex sense organs that are basically an extension of the brain. They evolved from primitive light-sensing spots on the surface of invertebrates and in some species have developed many remarkable variations, providing special advantages in various ecological niches. Each eye has a layer of receptors, a lens system for focusing an image on these receptors, and a system of axons for transmitting action potentials to the brain. This chapter describes how these and other components of the eye work.
The Eye’s Anatomy Is Adapted to the Eye’s Role as a Visual Receptor Organ
Figure 14-1 shows the anatomy of the normal eye in the horizontal plane. The white, outer protective layer encasing most of the eyeball is called the sclera. It is modified anteriorly into a clear, stratified squamous epithelial layer called the cornea. In the posterior two thirds of the eye the sclera is lined with a vascular and pigmented layer called the choroid. Interior to the choroid is the retina, the layer containing the photoreceptors.
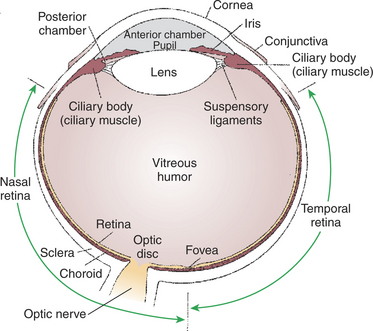
FIGURE 14-1 Schematic diagram of a horizontal section through the right eye as viewed from above.
(Redrawn from Walls GL: The vertebrate eye and its adaptive radiation, Cranbrook Institute of Science, Bulletin 19, 1942.)
As light passes through the transparent cornea, it undergoes some bending that will ultimately help to focus it on the retina. After passing through the cornea, the light enters a compartment called the anterior chamber. The anterior chamber and the posterior chamber are filled with a clear, waterlike fluid called aqueous humor that supplies important nutrients to the cornea (as well as the lens). Separating the anterior and posterior chambers is a diaphragm of varying size called the iris. The iris is a pigmented structure containing dilator and constrictor smooth muscle fibers arranged to vary the diameter of the pupil, the hole in the iris through which light passes on its way to the retina. The size of the pupil regulates the amount of light entering the eye. Behind the iris is the lens. The lens is suspended in the eye by the suspensory ligaments, which attach to the lens and to the ciliary body, a muscular structure at the base of the iris. The lens provides variable focusing power, in contrast to the fixed cornea.
Behind the lens is a chamber filled with a gelatinous fluid called the vitreous humor. Because of the viscosity of this fluid, the pressure generated by the aqueous humor, and the fairly inelastic nature of the sclera and cornea, the globe of the eye is basically spherical. The vitreous humor can also clear ocular debris potentially obstructing the light path. Behind the vitreous humor is the neural retinal layer where light is transduced into the electrical activity of neurons. The retina is interrupted at a point where axons of the retina’s ganglion cell layer, which travel across the inner surface of the retina, leave the eye on their way to the brain. This point, the optic disc, is a recognizable structure when the eye is examined with an ophthalmoscope (Figure 14-2). The interruption of the retina at the optic disc produces a “blind spot,” another name for the optic disc. The retinal ganglion cell axons leaving the eye at the optic disc give rise to the optic nerve (cranial nerve II), a cranial nerve so rich in axons that there are more axons in both optic nerves than in all the dorsal roots of the spinal cord.
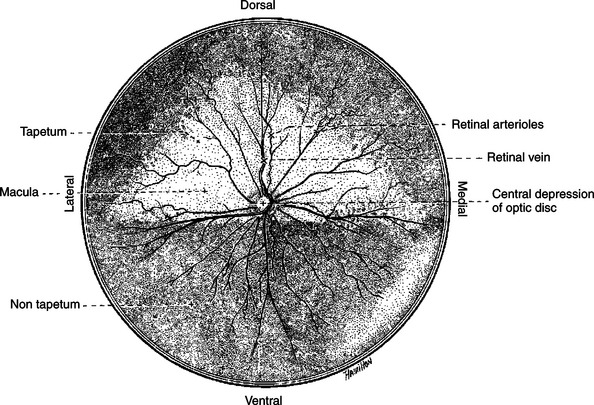
FIGURE 14-2 Ocular fundus of the right eye of a dog, as viewed with an ophthalmoscope, showing the optic disc and retinal blood vessels.
(From Evans HE, Christensen GC: Miller’s anatomy of the dog, ed 2, Philadelphia, 1979, Saunders.)
Also visible through the ophthalmoscope, on the surface of the retina, are the retinal blood vessels (see Figure 14-2). This network of arteries and veins enters the retina at the optic disc and provides much of the nutrition to the retina. Vessels of the choroid provide the remaining nutrition to the retina. Examination of retinal vessels often provides valuable clues to abnormalities elsewhere in the cardiovascular system.
The lacrimal gland, located near the lateral canthus of the eye, produces tears in response to parasympathetic nerve stimulation. Tears then flow over the cornea and are drained into the nose by the nasolacrimal duct. A regular flow of tears across the cornea is essential to the health of the cornea.
Through the Process of Accommodation, the Lens Changes Shape to Focus Images from Various Distances onto the Retina
When a camera focuses the images of objects at various distances from the film, the distance between the lens and the film is changed. The eye, however, focuses images by changing the shape of the lens, not by changing the distance between the lens and the retina.
Figure 14-3 shows the primary structures responsible for accommodation, the process by which the lens adds extra focusing power by changing its shape. The lens of the eye is made up of an elastic lens capsule containing a jellylike substance. If the eye’s lens were taken out of the eye, it would assume a spherical shape because of the elasticity of its capsule, much like a balloon filled with jelly. When suspended in the relaxed eye, however, the elasticity of the suspensory ligaments pulls on the equator of the lens, causing it to flatten in its anterior-posterior dimension. This flattened, less convex lens causes less refraction of light rays and allows the focus onto the retina of objects more than 20 feet away. To focus the image of objects closer to the eye, however, the lens must assume a more spherical, convex shape. This is accomplished by the contraction of the ciliary muscles of the ciliary body. This contraction of the ciliary muscle is much like the contraction of a sphincter and decreases the inner diameter of the ciliary body. In turn, this moves the attachments of the suspensory ligaments to a new position closer to the lens and decreases the pull on the equator of the lens. The result, given the inherent elasticity of the lens capsule, is a more spherical lens with more light refraction that focuses onto the retina the image of nearer objects. The more the ciliary muscle contracts, the more spherical the lens becomes.
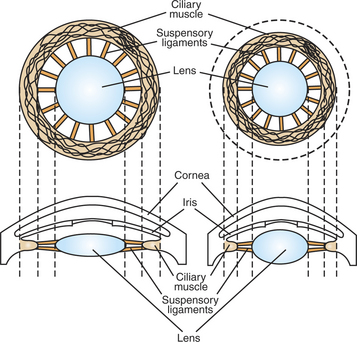
FIGURE 14-3 Primary ocular structures responsible for accommodation. The shape of the lens is shown when the ciliary muscle is relaxed (left) and contracted (right).
In humans, as the lens ages, it becomes less elastic and tends to become less spherical, even when the ciliary muscles contract. Many people older than 40 years of age need reading glasses to help their less elastic lens focus on objects nearby.
The lens should be clear and free of opacities. In cataracts, however, the lens becomes more opaque, causing random refraction of light and blurring vision, often leading eventually to blindness.
The Vertebrate Retina Consists of Five Major Cell Types
The retina is derived from neuroectoderm, a specialized portion of the ectoderm giving rise to the brain. For this reason, the retina is a fairly complex extension of the brain capable of considerable processing of the visual image before the information is transmitted toward the cerebral cortex.
The vertebrate retina consists of five major cell types: photoreceptor cells, bipolar cells, horizontal cells, amacrine cells, and ganglion cells (Figure 14-4). As in the cerebral and cerebellar cortices, the arrangement of these component cells is fairly consistent across the retina, suggesting a basic underlying processing mechanism and giving the retina a layered histology. However, local variations in the density of some cell types and in synaptic architecture account for particular functional variations within the retina.
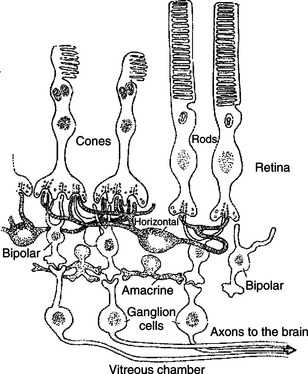
FIGURE 14-4 Vertebrate retina consists of five major cell types: photoreceptor cells (either rods or cones), bipolar cells, horizontal cells, amacrine cells, and ganglion cells.
(Redrawn from Kandel ER, Schwartz JH: Principles of neural science, ed 2, New York, 1985, Elsevier Science Publishing.)
There are two types of retinal photoreceptor cells: rods and cones (Figure 14-4). Both rods and cones make direct synaptic connection with the interneurons called bipolar cells, which connect the receptors with the ganglion cells. As previously noted, ganglion cell axons traverse the inner surface of the retina and converge at the optic disc to leave the eye and send action potentials to the brain through the optic nerves.
Modifying the flow of information at the synapses among the photoreceptors, bipolar cells, and ganglion cells are two interneuron cell types: the horizontal cells and the amacrine cells (Figure 14-4). The horizontal cells mediate lateral interactions among the photoreceptors and bipolar cells. The amacrine cells mediate lateral interactions among the bipolar cells and the ganglion cells.
The Fovea Solves a Distortion Problem Found in Other Areas of the Retina
The retinal ganglion cells are located in the inner retina (closer to the vitreous humor), whereas the photoreceptor cells (rods and cones) are located in the outer retina (closer to the choroid; see Figure 14-4). Therefore, throughout most of the retina, light rays travel through ganglion cells, bipolar cells, amacrine cells, and horizontal cells before reaching the photoreceptors. Although these inner neurons are unmyelinated (the ganglion cell axons become myelinated on leaving the eye) and therefore relatively transparent, they still cause some distortion of light rays.
The fovea is an area of the central retina designed especially to minimize this distortion. In the center of the fovea, in an area called the foveola, the inner ganglion and bipolar cells are pushed aside, allowing light rays unobstructed access to the photoreceptors (Figure 14-5). This is functionally significant because it allows light to have a less distorted path to the region of the retina associated with the highest visual acuity. The optic disc is just nasal to the fovea.
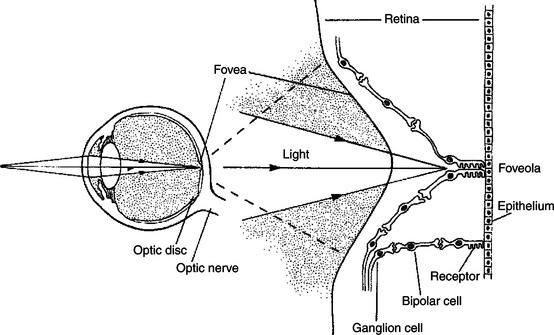
FIGURE 14-5 In most of the retina, light must first pass through overlying layers of neurons and their processes before it reaches the photoreceptors. in the center of the fovea, however, in the foveola, these neural elements are shifted aside; therefore, light has an unobstructed pathway to the photoreceptors in this region. An enlarged drawing of the foveal region is shown on the right.
(From Kandel ER, Schwartz JH: Principles of neural science, ed 2, New York, 1985, Elsevier Science Publishing.)
Pigment Behind the Retina Either Absorbs or Reflects Light, Depending on the Animal’s Habits
In animals that rely heavily on acute, daylight vision, there is a dark pigment in the epithelial layer between the photoreceptors and the choroid. This pigment absorbs light that has passed by the photoreceptors without stimulating them. If such light were reflected back into the retina, the sharpness of the visual image would be blurred. In nocturnal animals, however, these pigmented layers contain a reflecting pigment and are called the tapetum. This allows the retina to make optimal use of what light it receives, but at the expense of visual acuity. Reflection of light off the tapetum causes the familiar “night shine” from nocturnal animals’ eyes.
Photoreception and Transduction of Light Occur in the Rods and Cones
The anatomical structures of the rod and cone photoreceptors are similar, but there are some important differences. Because they are neurons, the rods and cones are primary receptors. Both cell types are divided into three parts: a synaptic terminal, an inner segment, and an outer segment (Figure 14-6). The photoreceptor synaptic terminal synapses with the bipolar cells. The inner segment includes the nucleus, mitochondria, and other cytoplasmic structures. The inner and outer segments are connected by a microtubule-containing cilium. The outer portions are specialized for photoreception. They contain an elaborate array of stacked membranous discs whose membranes contain visual photopigments. Visual photopigment is a light-sensitive molecular complex that initiates a biochemical chain of events, transducing light into neural electrical signals.
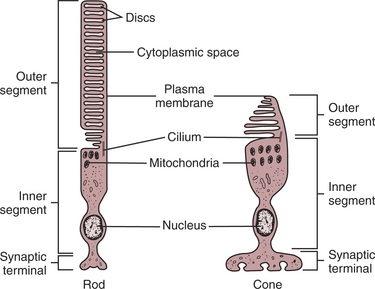
FIGURE 14-6 The two types of photoreceptors, rods and cones, have characteristic structures. Both rod cells and cone cells are differentiated into inner and outer segments connected by a cilium. The inner segments of both cell types contain the nucleus and most of the cell’s biosynthetic machinery and are continuous with the synaptic terminals. The membranous discs in the outer segment contain the light-transducing photopigment. The discs in the outer segments of rod cells are separated from the plasma membrane, whereas the discs of cone cells are not.
(Modified from O’Brien DF: The chemistry of vision, Science 218:961, 1982; from Kandel ER, Schwartz JH: Principles of neural science, ed 2, New York, 1985, Elsevier Science Publishing.)
The discs in the outer segments of the photoreceptors are regularly being formed near the cilium and phagocytized by the pigmented epithelium. Loss of this normal turnover in the outer segment may be important in several retinal diseases (e.g., retinitis pigmentosa). Photopigments of the discs are made up of proteins, called opsins, and retinal, an aldehyde of vitamin A. The light-sensitive retinal molecule is bound to the opsin, which is a member of the G-protein–coupled membrane receptor family (see Chapter 1). When light is absorbed by a rod or cone, the retinal is transformed in a way that activates the opsin, producing a change in the concentration of an intracellular second messenger, which leads to a change in the membrane potential of the photoreceptor cell. The configurational change in retinal is the only light-sensitive event in vision.
Unlike most sensory receptor cell membranes that depolarize with stimulation, photoreceptors hyperpolarize when struck by light. In rods the visual photopigment is called rhodopsin. In the dark, many gated sodium ion (Na+) channels remain open, allowing leakage of Na+ ions into the rod, which keeps the membrane in a depolarized state. When photons of light strike rhodopsin, the resulting change in second-messenger concentration produces a closing of many of the gated Na+ channels. The result is a hyperpolarization of the receptor cell membrane and a decrease in transmitter released at the synapse with the bipolar cell. Photoreception in cones works in a similar manner except that the photopigment opsin is different from that of rhodopsin. Depending on the species, different populations of cones may have different opsins. Because different opsins differ in their ability to absorb particular wavelengths of light, the number of different cone populations in a species is related to the ability of that species to discriminate different colors.
Differences in the functional properties of rods and cones, differences in their retinal distribution, and differences in the synaptic organization of other retinal neurons to which they pass their information result in a “rod retinal system” and a “cone retinal system” that have different functional attributes. Because individual rods are more sensitive to light than individual cones, and because several rods feed their synaptic input to a single bipolar cell (convergence), the rod system is the night-vision system concerned with detecting the presence of light. Because of the convergence of many rods on a bipolar cell, however, the rod system is not good for discerning the detail of a visual image (poor visual acuity). Also, since the rods are more highly concentrated in the peripheral retina than the cones, this region is better at detecting the presence of light than discerning its details. Finally, because all rods contain the same visual photopigment, the rod system cannot discriminate different colors.
As noted, a cone is less sensitive to light than a rod. Also, one or only a few cones feed their synaptic input to a single bipolar cell. Therefore the cone system needs higher levels of illumination to function compared with the rod system. In fact, the cone system actually represents the day-vision system because the rod system does not function well in high levels of illumination. However, due to a lack of convergence onto bipolar cells, the cone system is good at discerning image details. Because the cones have a particularly high density in the region of the fovea (the central retina), the fovea is the part of the retina with the highest visual acuity. This makes sense considering the fovea is where light falls when an animal looks directly at an object of interest. Again, different populations of cones can contain different photopigments, each differentially sensitive to a range of light frequencies. Thus the cone system is also the color-vision system. Generally, the more cone photopigments present in the retina, each within a different population of cones, the better is the ability of the species to discriminate different wavelengths of light, or to “see” color. In primates there are three different popu-lations of cones, each with a different photopigment. According to the Young-Helmholtz theory, the brain assigns color to a given wavelength of light by comparing the relative activation of the different cone populations. Humans who lack one or more of these cone types, as a result of faulty genetic transmission, cannot see colors within particular ranges of the visible spectrum of light. The most common types of such “color blindness” are linked to the X chromosome.
The extent to which various veterinary species perceive color is still controversial. It is believed that mammalian ancestors may have had four different types of cones, whereas early nocturnal mammals may have had only two types of cones, having traded some of their color discrimination ability for rod light sensitivity. Most current mammals, including dogs, have only two types of cones. As noted, primates have three types of cones. It is thought that their successful exploitation of color-rich arboreal environments may have induced selection pressure for additional color discrimination ability, above that imparted by two populations of cones. Modern birds appear to have four populations of cones, one of which is sensitive to light in the ultraviolet (UV) range of the spectrum. Behavioral experiments that show their ability to discriminate UV light suggest that birds have a richer color perception than primates. Lizards, turtles, and some fish also possess UV-sensitive cones. Only primates are known to have the color vision with which humans are familiar.
The Response of the Photoreceptor to Light Is Synaptically Transmitted to the Ganglion Cells by the Bipolar Cells
The hyperpolarizing response of the rods and cones to light influences bipolar cells by a chemical synapse. In turn, the bipolar cell influences action potential frequencies in the ganglion cell axons on their way to the brain. Considerable processing of the visual image occurs in the retina, such as the initial stages of color discrimination, contrast enhancement, and directional sensitivity, in addition to light-dark discrimination and detection of large changes in luminance. The synaptic interactions among photoreceptors, bipolar cells, and horizontal cells (see Figure 14-4) play a significant role in contrast enhancement and directional sensitivity. The interactions of amacrine cells with bipolar cells and ganglion cells appear to play a role in processes such as the ability to detect changes in the rate of alternation between light and dark. A more detailed description of the synaptic and membrane changes in the chain of transmission within the retina, accounting for these types of retinal image processing, is beyond the scope of this book. To learn more about the many interesting and unusual phenomena occurring in the retina, the reader should refer to the Bibliography.
The Electroretinogram Records the Electrical Response of the Retina to a Flashing Light
The electroretinogram (ERG) is a clinical electrophysiological recording from the cornea and skin near the eye. It records the electrical response of the retina to a light flashed into the eye. It has three waves: the A wave, corresponding primarily to the activation of visual pigment and photoreceptors; the B wave, caused primarily by the response of retinal bipolar cells; and a slower C wave, thought to originate in the pigment epithelium. The ERG is a fairly simple, general electrodiagnostic tool for assessing some forms of retinal dysfunction or degeneration.
Ganglion Cell Axons Transmit Action Potentials to the Visual Cortex by Way of the Lateral Geniculate Nucleus
Sets of retinal ganglion cells leaving the eye participate in three important visual pathways: the retino-geniculo-striate pathway, the retino-tectal pathway, and the retino-hypothalamic pathway. The retino-geniculo-striate pathway is principally involved in conscious visual perception of form, color, motion, orientation, and depth. The retino-tectal pathway plays an important role in pupillary reflexes and reflex orientation of the eye to visual targets. The retino-hypothalamic pathway plays a role in the regulation of physiological rhythms by light-dark cycles (e.g., seasonal changes in day length).
In order for the retinal image, originating from light in the visual field, to reach consciousness, the information must be transferred to the visual cortex. Figure 14-7 shows the retino-geniculo-striate pathway by which the axons of the retinal ganglion cells project to the lateral geniculate nucleus of the thalamus and by which cells in the lateral geniculate nucleus project their axons to the primary visual cortex of the occipital lobe. Note that ganglion cell axons from the temporal retina (closest to the ear; see Figure 14-1) travel along the optic nerve to the optic chiasm and then project ipsilaterally to the lateral geniculate nucleus on the same side of the brain. Ganglion cell axons from the nasal retina (closest to the nose) come to the optic chiasm and cross to the contralateral lateral geniculate nucleus. Retinal ganglion cell axons between the region of the optic chiasm and the lateral geniculate nuclei are referred to as the optic tracts. Cells in each lateral geniculate nucleus then send axons to the ipsilateral primary visual cortex, in the occipital lobe, by way of the optic radiations. This arrangement results in the right half of each retina sending information to the right primary visual cortex and the left half of each retina sending information to the left primary visual cortex. Light originating in the left half of the environmental visual field generally falls on the right half of the retina of each eye, whereas light originating in the right half of the environmental visual field falls on the left half of the retina of each eye. Given the anatomical map of the retino-geniculo-striate pathway in Figure 14-7, information from an image arising in the left field of vision would be received in the right primary visual cortex, and image information arising from the right field of vision would be received in the left primary visual cortex.
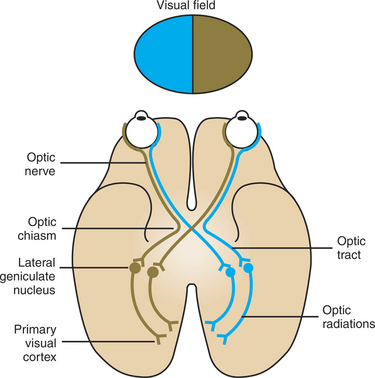
FIGURE 14-7 Retino-geniculo-striate pathway to cerebral cortex. Axons of the retinal ganglion cells project to the lateral geniculate nucleus of the thalamus, and axons from cells of the lateral geniculate nucleus project to the ipsilateral primary visual cortex of the occipital lobe. The right environmental visual field projects to the left half of each retina, and the left environmental visual field projects to the right half of each retina. The right half of each retina sends information to the right primary visual cortex, and the left half of each retina sends information to the left primary visual cortex. Therefore, information from the left visual field arrives in the right primary visual cortex, and information from the right visual field arrives in the left primary visual cortex. The labeled components of each pathway are present on both sides of the brain.
Information reaching primary visual cortex, or V1 (also called striate cortex), is passed to other occipital lobe areas of visual cortex (e.g., V2, V3) for more complex processing, then to visual association areas of the parietal and temporal lobes for the highest forms of visual information processing and integration with other sensory modalities. There appears to be parallel processing of retinal information within the retino-geniculo-striate pathway and its higher-order processing regions. One major stream of information in this system apparently is concerned with conscious object identification (e.g., form, color), whereas a parallel stream of information relates to conscious awareness of where an object is located (e.g., motion, position). The three major projection pathways of retinal ganglion cell axons—retino-geniculo-striate, retino-tectal, and retino-hypothalamic—represent another example of the parallel processing of retinal information.
The Diameter of the Pupil Is Controlled by the Autonomic Nervous System
The iris of the eye contains two sets of smooth muscle fibers. One set, arranged in a circular pattern around the pupil, causes the pupil to become smaller (constrict) when the fibers contract. These constrictor fibers are innervated by postganglionic parasympathetic neurons whose cell bodies lie in the ciliary ganglion, just behind the eye, and which secrete acetylcholine as the neurotransmitter to the muscle. These parasympathetic postganglionic neurons are activated by parasympathetic preganglionic neurons whose axons travel in the oculomotor cranial nerve (cranial nerve III) and whose cell bodies reside in the Edinger-Westphal nucleus of the midbrain.
The other smooth muscle fibers of the iris are arranged radially from the pupil, resembling spokes of a wheel. When these radial smooth muscle fibers contract, they cause the pupil to become larger (dilate). These dilator fibers are innervated by the sympathetic nervous system. Sympathetic preganglionic neurons begin in the first two thoracic segments and course cranially in the vagosympathetic trunk to synapse in the superior cervical ganglion of the neck. The sympathetic postganglionic axons originate in cells of this ganglion and course to the region of the eye, where they innervate the dilator fibers of the iris, in addition to the muscle that helps lift the upper eyelid and the muscle that helps keep the “third eyelid” in place at the medial canthus of the eye. The sympathetic postganglionic axons also innervate sweat glands and vascular smooth muscle to the face.
The Retina, Optic Nerve, and Autonomic Nerve Supply Controlling the Pupil Can Be Tested with a Flashlight
When a light is shone into the eye, the pupil of that eye constricts. This action is called the direct pupillary light reflex. The light triggers the photoreception mechanism, leading to ganglion cell action potentials transmitted along the optic nerve. Some of the ganglion cell axons of the retino-tectal pathway synapse in the pretectal region of the brain. The pretectal neurons then synapse on the parasympathetic preganglionic neurons of the Edinger-Westphal nucleus, whose axons travel in the oculomotor nerve to synapse on parasympathetic postganglionic neurons in the ciliary ganglion. Stimulation of these postganglionic neurons causes constriction of the pupil by stimulating the constrictor smooth muscle fibers of the iris. A normal direct pupillary light reflex tests the integrity of the retina, the ipsilateral second and third cranial nerves, a limited region of the brainstem, and the iris. The axons of pretectal neurons also cross the midline, so that when a light is shone into one eye, not only does the pupil on the same side constrict (direct pupillary light reflex), but the contralateral pupil also constricts. This action is called the indirect or consensual pupillary light reflex. It also requires the integrity of the contralateral oculomotor (third) cranial nerve.
Aqueous Humor Determines Intraocular Pressure
Aqueous humor is a clear liquid found in the anterior and posterior chambers of the eye. Its rate of production and absorption is sufficiently high to replace the entire chamber’s volume several times a day.
Aqueous humor is secreted by the ciliary process, a system of fingerlike projections on the ciliary body of the posterior chamber. Aqueous humor is thought to be formed by the active transport of sodium, chloride, and possibly bicarbonate ions into the posterior chamber. This establishes an osmotic gradient, causing water to flow passively into the posterior chamber. Aqueous humor flows from the posterior to the anterior chamber through the pupil. Flow is caused by a pressure gradient established by the active process of formation in the posterior chamber.
Aqueous humor is then absorbed into the venous system at the angle between the cornea and the iris. This absorption is driven by a pressure gradient and is assisted, in many species, by a system of trabeculae and canals. If this absorption into the venous system is obstructed, intraocular pressure increases because the production of aqueous humor continues. This pathologic increase in intraocular pressure is called glaucoma. As intraocular pressure exceeds intravascular pressure in the blood supply to the retina, blindness results.
CLINICAL CORRELATIONS
Homonymous Hemianopia
History.
You examine a 10-year-old male German shepherd whose owner reports that the dog has recently begun to bump into objects with the left side of his face and has had two seizures. The seizures were characterized by turning of the head to the left and stiffening of the left front leg.
Clinical Examination.
Physical examination abnormalities are limited to the nervous system. When presented with a maze of unfamiliar objects in the examination room, the dog collides with objects as if he does not see from the left side. He seems somewhat weak in the left front leg. The dog is otherwise bright, alert, and responsive. Cranial nerve and spinal segmental reflexes are within normal limits, as are intersegmental, proprioceptive placing responses for the right front and right rear legs. However, the proprioceptive placing responses for the left front and left rear legs are quite prolonged.
Comment.
This dog’s history and neurological examination abnormalities are common in dogs with brain tumors. This dog has a tumor (neoplasm) arising from the meninges over the right posterior cerebral cortex. It is in this posterior (occipital) cortex that the visual image is interpreted from the visual field of the left side (see Figure 14-7). Lesion-induced functional damage to the right occipital cortex results in loss of vision from the left visual field (homonymous hemianopsia). Also in the right cerebral cortex, the conscious proprioception response for the left legs is interpreted. This dog’s seizures feature turning the head to the left and displaying transient rigidity of the left front leg, because the seizure activity arose from the cerebral cortex at the site of the tumor and spread to the right motor cortex but remained limited to the cerebral cortex on the right side. Because the pyramidal system’s corticospinal tract controlling the muscles of the left neck and left front leg arise in the right motor cortex (see Chapter 10), seizure activity causes the transient head turning and left leg stiffness.
Bear MF, Connors BW, Paradiso MA. Neuroscience: exploring the brain, ed 3. Philadelphia: Lippincott Williams & Wilkins, 2007.
Boron WF, Boulpaep EL. Medical physiology: a cellular and molecular approach, updated-edition. Philadelphia: Saunders, 2005.
Goldsmith TH. What birds see. Sci Am. 2006;295(1):68.
Goldstein BE. Sensation and perception, ed 6. Pacific Grove, Calif: Wadsworth, 2002.
Guyton AC, Hall JE. Textbook of medical physiology, ed 11. Philadelphia: Saunders, 2006.
Haines DE. Fundamental neuroscience for basic and clinical applications. Philadelphia: Churchill Livingstone, 2006.
Kandel ER, Schwartz JH, Jessell TM. Principles of neural science, ed 4, New York: McGraw-Hill, 2000.
Nichols JG, Martin AR, Wallace BG, Fuchs PA. From neuron to brain, ed 4. Sunderland, Mass: Sinauer Associates, 2001.
Purves D, Augustine G, Fitzpatrick D, et al. Neuroscience, ed 3. Sunderland, Mass: Sinauer Associates, 2004.
PRACTICE QUESTIONS
1. A patient whose left pupil diameter is smaller than normal, whose left upper eyelid droops, and whose left “third eyelid” is abnormally displaced likely has a lesion of which of the following structures?
2. Which of the following is not characteristic of the cone system?
3. Your friend, a member of the soccer team, is trying without much success to explain the cause for the team’s recent losing trend. A variety of implausible explanations are proposed until she mentions that she is progressively losing vision, but only in her left visual field, and has frequent headaches. You recommend that she see a neurologist because she likely has a lesion in her:
4. Which of the following regarding the lens is false?
5. You examine a patient’s pupillary light reflexes. Shining a light into the left eye produces both a positive direct and a positive consensual pupillary response. However, shining the light into the right eye produces neither a direct nor an indirect pupillary response. This patient’s pathological lesion is located in which of the following structures?