Somatosensory System
 Introduction
IntroductionGeneral sensory mechanism
Introduction
Sensations
Sensory division of the human nervous system is concerned with collection of the information about outside world and changes occurring within the body itself. Sensation refers to conscious perception of sensory information reaching the brain. Sensations may be broadly classified into two groups.
1. Special senses. These include visual sensations, auditory sensations, gustatory (taste sensation) and olfactory (smell) sensations.
2. Somaesthetic senses. These, depending upon their point of origin, can be classified into three types.
A. Exteroceptive sensations also known as cutaneous sensations arise from the surface of the body and these include:
B. Visceral sensations arise from the viscera, i.e. internal organs and is called visceral sensation.
C. Proprioceptive and kinaesthetic sensations arise from the muscles, tendons and joints. These include the following.
(i) Proprioceptive sensations. These are concerned with the physical state of the body, i.e. the sense of position, tendon and muscle sensations, deep pressure and sense of equilibrium.
(ii) Kinaesthetic sensations or kinaesthesia. It is the conscious recognition of rate of movement of different parts of the body. Kinaesthetic sensations include both:
Components of sensory system
The sensory division of the human nervous system includes following components:
1. Sensory receptors. These are specialized cells that transduce stimulus energy into neural signals.
2. Afferent neurons. These carry sensory impulses to the sensory cortex and constitute the neural pathway.
3. Sensory cortex. It includes the sensory areas of cerebral cortex. The information received in the sensory cortex results in a conscious perception of the stimulus, i.e. a sensation
Receptors
Sensory receptors are specialized cells that receive stimuli from the external or internal environment and transduce these signals into neural signals. A stimulus is a change of environment of sufficient intensity to evoke a response in an organism. The external stimuli may be mechanical, chemical, thermal, auditory or visual.
Classification of receptors
Receptors for somatic sensations
A Receptors for cutaneous sensations include
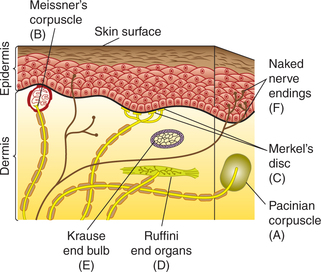
1. Mechanoreceptors or receptors of touch, pressure and vibration sensation provide information about these stimuli to skin. In general, these receptors consist of an unmyelinated axon surrounded by lamellated connective tissue corpuscles. These include (Fig. 10.3-1):
2. Thermoreceptors. Thermoreceptors are responsible for detecting temperature sensation. Free nerve endings of unmyelinated C fibres form the warm receptors and that of small myelinated Aδ fibres form the cold receptors.
3. Nociceptors. Nociceptors or pain receptors refer to special type of free nerve endings of:
B Receptors for proprioceptive and kinaesthetic sensations
The receptors concerned are called proprioceptors and include:
• Muscle spindle or stretch receptors (see page 520).
• Joint receptors located in the joint capsules and ligaments around the joints. Ruffini's end organs are the most important receptors for this function. A few pacinian corpuscles are also involved.
• Golgi tendon organ (see page 522).
• Vestibular receptor (see page 531).
Sensory transduction
Sensory transduction refers to the phenomenon of transduction of environmental signals into neural signals by the receptors. Steps of sensory transduction are:
Arrival of stimulus to receptor
The stimulus arriving at the given sensory receptor may be in the form of:
• Mechanical force causing depression of the skin which stimulates mechanoreceptors,
• Light or electromagnetic wave which stimulates photoreceptors of the retina,
• Chemical, e.g. a molecule of NaCl on the tongue which stimulates chemoreceptors,
• Cold or warm temperature stimulating thermoreceptors and
• Sound energy stimulating auditory receptors, and so on and so forth.
Production of receptor potential
When a stimulus excites the receptor, it changes the potential across the membrane of the receptors. This change in the potential is called receptor or generator potential.
Mechanism of development of receptor potential (Fig. 10.3-2)
The change in membrane potential in a receptor is caused by a change in permeability of membrane of the unmyelinated terminals to Na+. The resultant influx of Na+ causes development of generator or receptor potential.
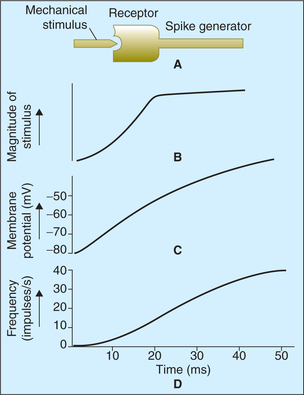
Fig. 10.3-2 Mechanism of development of generator potential and its relationship with intensity of stimulus. A, Stimulus to mechanoreceptor causes its deformation, which opens up channels which are permeable to Na+ causing membrane depolarization; B, the magnitude of stimulus intensity; C, the receptor potential (generator potential) follows the time course and D, the action potential. Note. The magnitude of generator potential and frequency of action potential are proportionate to the magnitude of the stimulus.
Usually the current is inward, which produces depolarization of the receptor. The exception is in the photoreceptors, where light causes hyperpolarization.
Production of action potential in a sensory nerve
The receptor potential developed in unmyelinated nerve ending (transducer region) depolarizes the sensory nerve at the first node of Ranvier (spike generator region) by electrotonic depolarization current sink action. When the receptor potential rises above the threshold level (i.e. above 10 mV), it brings the membrane potential of the first node of Ranvier to the firing level causing production of action potential, which is propagated in the nerve fibre (Fig. 10.3-2D). Thus, the first node of Ranvier (spike generator region) converts the graded response of the receptor into action potential. Greater the magnitude of receptor potential, greater is the rate of discharge of action potentials in the nerve fibre.
Properties of receptors
1. Specificity of response. Each receptor is easily stimulated (has low threshold) by only one type of appropriate (adequate) specific stimulus. This specificity of response by a particular receptor is also called law of adequate stimulus. Although each receptor is exquisitely sensitive to its adequate stimulus, receptors can respond to other forms of energy if the intensity is high enough. For example:
Adequate stimulus for rod and cones of retina is light. Pressure on the eyeball can also stimulate retinal receptors, but the threshold of these receptors to pressure is much higher than the threshold of the pressure receptors in skin.
2. Production of receptor potential on stimulation. As described in detail above.
3. Adaptation. When a receptor is continuously stimulated with the same strength of stimulus, the frequency of action potential in its sensory nerve decreases progressively. This property is called adaptation. Depending on the rate of adaptation the receptors are of two types.
Tonic receptors. These receptors keep on firing action potentials continuously during stimulus application. Such receptors are important for life as they keep the brain constantly apprised of status of the body and its relation to its surroundings. Examples of such receptors are:
Phasic receptors. These are rapidly adapting receptors. These transmit signals only when the stimulus strength is changed. Examples of phasic receptors are:
4. Effect of strength of stimulus. Receptor potential amplitude depends on the strength of stimulus. During the stimulation of a receptor, if the response given by the receptor is to be doubled, the strength of stimulus must be increased 10 times. This phenomenon is called Weber-Fechner law.
5. Effect of velocity of stimulus. The magnitude of the receptor potential rises with rate of change of stimulus application. It also applies to removal of stimulus, e.g. off response.
6. Projection. When any part of sensory path is stimulated, conscious sensation referred to the location of receptor is produced. This is called law of projection (for details see encoding of sensation).
Somatosensory system
Somatosensory system can be discussed in under following headings:
Somatic sensations
Touch, pressure and vibration sensations
Mechanoreceptors provide information about touch, pressure and vibration stimuli to skin (Fig. 10.3-1).
The salient features of main mechanoreceptors are summarized in Table 10.3-1.
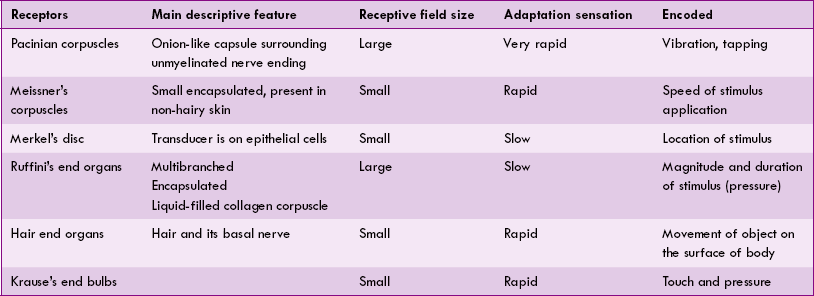
Functions of touch and pressure mechanoreceptors
1. Detection of touch, pressure and vibration sensations by mechanoreceptors. Touch, pressure and vibration are considered to be different form of the same sensation.
2. Two-point discrimination. It is the ability to distinguish two touch stimuli separately. The minimum distance by which two touch stimuli can be perceived as separate stimuli varies from a 2-3 mm on the lips and fingertips to over 60 mm on the back of the trunk. The difference in the distance between the two points seems to be related to the density of touch receptors in different parts of the body.
3. Stereognosis. Stereognosis refers to the ability to recognize familiar objects such as a key, coins, pen, pencil, spoons, etc. by merely handling them without looking at them.
Temperature sensation
Thermoreceptors
Location. Thermoreceptors are located in the skin of all parts of the body. However, density of thermoreceptors is greatest in the lips, moderate in the fingertips and least in the skin of trunk.
Stimulus. Thermoreceptor responds to the temperature of subcutaneous tissue surrounding them and not to the environmental temperature as such. Because of this reason cold metal objects feel colder than wooden objects of the same temperature. The metal being good conductor, conducts heat away from the skin more rapidly and cools the subcutaneous tissue to a greater degree than the wood.
The salient features of response exhibited by warm and cold receptors are given below.
They are activated when skin temperature is between 30°C and 43°C (Fig. 10.3-3). The firing rate of warm receptors reaches a peak at temperatures of approximately 42°C.
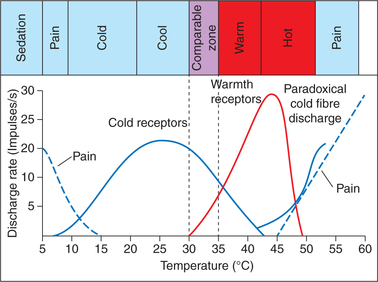
• They are activated when the skin temperature is between 10°C and 40°C (Fig. 10.3-3). The steady state firing rate of cold receptors reaches a peak at temperatures between 25°C and 30°C.
• Paradoxically, temperatures between 45°C and 50°C stimulate cold fibres as well as pain fibres producing a mixed sensations of cold and pain (Fig. 10.3-3).
• Cold temperature below 10°C stimulates only pain receptors (Fig. 10.3-3).
• Thus, between 30°C and 40°C both cold and warm receptors are stimulated, which help the person in fine gradation of temperature. Therefore, between 30°C and 40°C (neutral or comfort zone), complete perceptual adaptation occurs (i.e. awareness of temperature disappears).
Pain sensation
Definition and purpose
Pain refers to an unpleasant sensory and emotional experience associated with actual or potential tissue damage.
Pain sensation is different from other sensations because its purpose is to inform the brain that the stimulus is physically damaging. Therefore, though the pain sensation is unpleasant, it is useful in the following ways:
• It makes one aware of a harmful agent in close contact with the body and body gives preferential treatment to this information.
• It causes the individual to react to remove the pain stimulus to prevent further damage to the tissues.
• Pain receptors are non-adaptable receptors; therefore, they keep the person apprised of damaging stimulus as long as it persists. Thus pain sensation has a protective function.
Pain receptors, stimuli and chemical mediators of pain
Nociceptors refer to special type of free nerve endings of:
Types. Nociceptors broadly can be grouped as somatic nociceptors and visceral nociceptors.
1. Somatic nociceptors are free nerve endings of Aδ and C fibres as mentioned above.
2. Visceral nociceptors. Visceral pain is not produced by many tissue-damaging stimuli such as localized cutting, clamping or burning. However, widespread inflammation, ischaemia, mesenteric stretching, or spasm or dilatation of hollow viscera produce pain.
Pain receptors are activated by three types of noxious stimuli: mechanical, thermal and chemical.
Chemical stimuli. Damaged tissue releases certain chemicals which act on nociceptors and cause pain sensations. Chemical mediators of pain include:
• K+, ATP and ADP are released following cell death.
• Bradykinin is released by dying cells.
• Leukotrienes are released from mast cells.
• Serotonin is released from platelets.
• Histamine is released from mast cells.
• Accumulation of lactic acid in tissues due to anaerobic mechanism during ischaemia.
• Substance P and calcitonin gene-related peptide (CGRP).
• Prostaglandins and substance P enhance the sensitivity of pain endings but do not directly excite them.
Qualitative types of pain sensations
Qualitatively pain sensations are of two types: fast pain and slow pain.
Fast pain is a sharp, well-localized, pricking sensation that results from activation of the nociceptors on the Aδ fibres. The fast pain sensations travel faster and thus appear within 0.1 ms after the application of stimulus.
Clinical types of pain
In clinical practice pain sensations can be classified as:
Somatic pain, as the name indicates, arises from the tissues of the body other than viscera. It is of two types:
Superficial somatic pain arises from the skin and superficial tissues. Its features are usually similar to the fast pain.
Deep somatic pain arises from the muscles, joints, bones and fascia. Usually its features are similar to that of slow pain.
Features of visceral pain are:
• Poorly localized because pain receptors in viscera are comparatively few.
• Unpleasant because of emotional perception.
• Autonomic symptoms in the form of nausea, vomiting, profuse sweating and lowering of blood pressure.
• Reflex contraction of skeletal muscle of abdominal wall, clinically known as guarding, is a common association especially when inflammation of viscera involves peritoneum. It is a protective reflex which helps to protect the underlying inflamed structures from unintentional injury.
• Radiates or is referred to other site (see referred pain).
Referred pain as the name indicates is that pain which originates due to irritation of a visceral organ and is felt not only in the organ but also in some other somatic structure (usually skin) supplied by the same neural segment.
Characteristic features of referred pain are:
1. Such a pain is said to be referred to the second structure. For example,
• In myocardial ischaemia, pain is referred to the left shoulder and arm.
• Pain due to stone in lower part of ureter is usually referred to the corresponding testis and inner thigh.
• Inflammation of diaphragm secondary to pleurisy or severe cholecystitis produces pain at the tip of shoulder.
2. Pain is usually referred to a structure with common embryonic origin, and hence are innervated by a common neural segment. This principle is called the dermatomal rule. For example, embryologically the heart and the left arm have the same segmental origin. Similarly, the testes and kidney develop from the same primitive urogenital ridge.
Theories of referred pain are discussed below.
1. Convergence theory: According to this theory when the first-order neurons carrying pain sensation from a somatic area and a visceral organ converge on a common second-order neuron (Fig. 10.3-4A) the brain is unable to identify the source of pain. Since somatic pain is far more common, the brain interprets all pain as somatic pain even when the source is actually visceral.
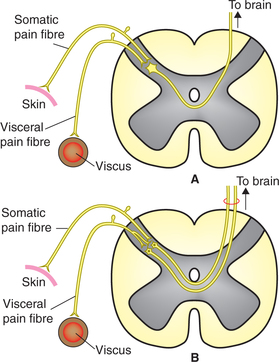
2. Facilitation theory: According to this theory the visceral irritation is inadequate for producing pain by itself.
However, it facilitates pain fibres from somatic structures (Fig. 10.3-4B) so that even minor somatic irritation produces perceptible pain.
Sometimes visceral pain is experienced both locally and also at distant point (referred pain). In fact pain seems to spread from the local area to the distant site. This is called radiating pain. Example of radiating pain are:
When the sensory fibres carrying pain sensations are stimulated anywhere in their course to the sensory cortex, the pain sensations evoked are projected to the area where receptors are located. This is called projected pain. It follows the law of projection (see page 514). Examples of projected pain are:
• After amputation of a limb, sometime patient complains of intense pain in the absent limb (phantom limb). The pain sensations are produced due to irritation of nociceptive fibres at the stump but are projected to the area where receptors used to be located.
• Striking the elbow causes pain to be projected to the hand.
Causalgia
Causalgia is a condition in which spontaneous burning pain sensation occurs after a long time in the area of even trivial injuries. It is also accompanied by hyperalgesia and reflex sympathetic dystrophy.
Pain suppression systems in CNS
The degree of reaction to painful stimuli varies from individual to individual, mainly because of existence of pain suppression systems in the central nervous system. The pain suppression consists of two major components:
A Spinal pain suppression system
There exists a pain inhibitory complex in dorsal horn of spinal cord which blocks the pain signals at the initial entry point to the spinal cord.
Gate control hypothesis has been put forward by (Metzak and Wall in 1965). According to this hypothesis, the dorsal grey horn acts as gate for transmission of pain sensation and this gate can be partly or completely closed by:
1. Segmental suppression. It has been observed that activation of large myelinated touch fibres (Aβ) reduces pain. Aβ fibres give collaterals which cause presynaptic inhibition (primary afferent depolarization) of pain carrying (both type C and Aδ) fibres, where they synapse in the dorsal horn (Fig. 10.3-5). Although poorly understood, such circuitry probably explains the relief of pain achieved by following manoeuvres:
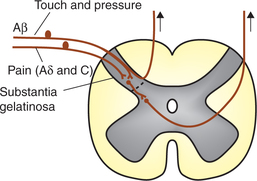
Fig. 10.3-5 Spinal pain suppression system. Note the collateral from Aβ fibres from touch receptors cause presynaptic inhibition of pain afferent Aδ and C fibres.
• Rubbing or massage or pressure in the vicinity of painful area.
• Local application of warmth or cold.
• Local application of counterirritants, i.e. stimulation of skin.
• Transcutaneous electric nerve stimulation (TENS) in which pain site or the nerves leading from it are stimulated by electrodes placed on the surface of skin.
2. Supraspinal suppression is caused by the supraspinal suppression system described below.
B Supraspinal pain suppression system
There exist three different supraspinal descending pain modulation pathways:
• Descending serotonergic and opioid inhibitory system,
1. Descending serotonergic and opioid inhibitory system.
Components of this system (Fig. 10.3-6) are:
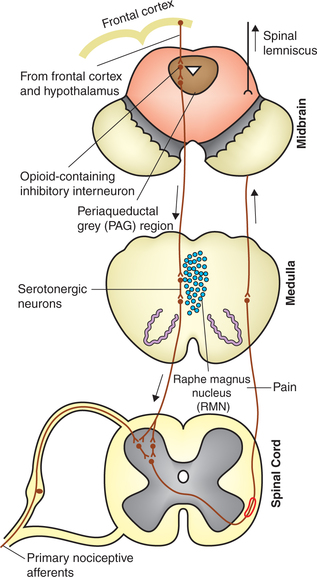
(i) Raphe magnus nucleus (RMN). The serotonergic neurons of the RMN project down the dorsolateral column to influence the neurons in dorsal horn of spinal cord, which are excited by primary nociceptive afferents. The serotonergic fibres are believed to exert their effect by postsynaptic inhibition.
(ii) Periaqueductal grey (PAG) area in the midbrain. It inhibits pain by stimulating the RMN. Neurons of PAG have opioid receptors on their surface membranes. When opioid receptors are stimulated by exogenously administered opioid compounds (analgesics) or by endogenous opioid neurotransmitters (endorphins and enkephalins) found in the brain, the pain suppression circuitry is activated and this leads to reduced pain perception.
Conditions under which descending serotonergic and opioid pain inhibitory system is stimulated are:
(i) When limbic system is stimulated. Limbic system is the seat of emotions. Fibres from limbic system supply the PAG. This explains why a soldier wounded in the battlefield may feel no pain during the heat of battle.
(ii) Autofeedback. When the spinothalamic tract (STT) is stimulated, the collaterals from STT stimulate the descending inhibitory pathway.
2. Descending purinergic inhibitory system, comprising specifically of adenosine. Adenosine exhibits both pre and postsynaptic actions, and produces antinociception by indirect interaction with excitatory amino acid release.
3. Descending noradrenergic inhibitory system. Fibres of this system originate from the locus coeruleus and medullary reticular formation, and descend in dorsolateral fasciculus. Environmental factors such as stress may activate this descending inhibitory mechanism.
Other sensations
This group includes other sensations except somatic sensations (touch, pressure, pain and temperature) like:
Itch. It is an irritative skin condition which occurs due to mild stimulation (especially when something moves across the skin).
1. It occurs only in the skin, eyes and certain mucous membranes but not in the deep tissue and viscera.
2. It originates due to stimulation of itch receptors which are naked nerve endings of unmyelinated C fibres. The receptors are stimulated by two ways:
• By repeated local mechanical stimulation of skin.
• By certain chemical agents (e.g. bile salts, histamine and kinins).
3. Scratching relieves the itching. The mechanism is same as that of gate control hypothesis in pain sensation.
Tickle. Tickle is regarded as a pleasurable feeling as compared to itching (which gives annoying feeling) and pain (is an unpleasant feeling).
Synthetic sense. The combinations of various cutaneous sensations produce different experiences which are entirely different from primary sensation. Therefore the new experience is regarded as synthetic sense.
Pathways in somatosensory system (transmission of sensations)
Neurons of sensory pathway
Pathways in somatosensory system are formed by a chain of three neurons which ultimately reach the sensory cortex.
First-order neurons
These are the primary afferent neurons that receive the transduced signals from the sensory receptors and carry them to the spinal cord or brain stem. The cell bodies of the primary afferent neurons are located in dorsal root ganglia. These are T-shaped unipolar neurons with peripheral and central processes. The peripheral processes reach the sensory receptors and form the sensory part of spinal nerves (which are mixed nerves). Central processes constitute the dorsal nerve root of the spinal nerve. The fibres of first-order neurons in the spinal nerves and dorsal nerve root comprise Aα-, Aβ-, Aδ- and C-type fibres and are often referred to group I, II, III and IV, respectively by the sensory physiologists.
Sensory nerves and dermatomes
Sensory nerves
All sensory fibres reach the CNS through their cranial equivalents.
Dorsal nerve roots in spinal cord. Each dorsal nerve root is attached to the spinal cord through various rootlets. Each rootlet just before entering the spinal cord divides into medial and lateral divisions.
Medial divisions of each rootlet consists of myelinated group I and II fibres which include:
Dermatomes
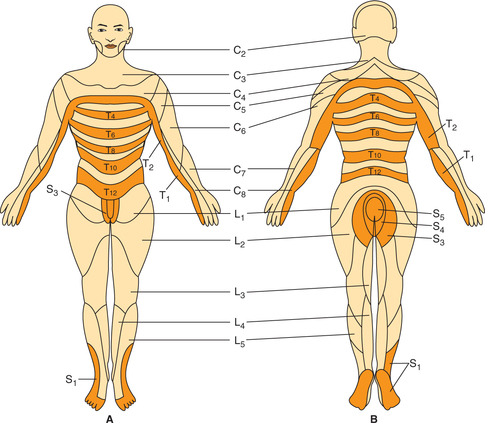
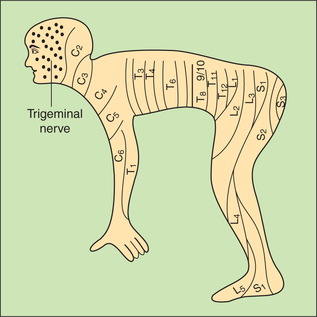
Dermatome refers to the area of skin supplied by one dorsal root (spinal cord segment). It is important to note that dermatomes are quite different from the peripheral nerve fields because fibres from one dermatome may be present in different peripheral nerves.
During embryo stage body is divided into orderly meta-meres. In the postnatal life, owing to excessive growth of limbs, the metamere arrangement, excepting in the trunk, is no longer present. Therefore, dermatomes are remnants of orderly metameric arrangement which has survived only in the trunk, where the dermatomes consist of a series of 12 narrow overlapping bands running from the vertebral column to the midventral line (Fig. 10.3-7). However, this apparent complexity of the dermatomes in man is simplified if the man is visualized as a quadruped animal (like monkey, the ancestor of man) (Fig. 10.3-8). The knowledge of dermatomes is utilized to know the level of spinal cord injury by mapping the area of altered sensation produced.
Ascending sensory tracts
The major ascending sensory tracts in the spinal cord have been grouped as:
Dorsal column sensory pathway
Dorsal column sensory pathway (Fig. 10.3-9) in man is well developed and completely myelinated.
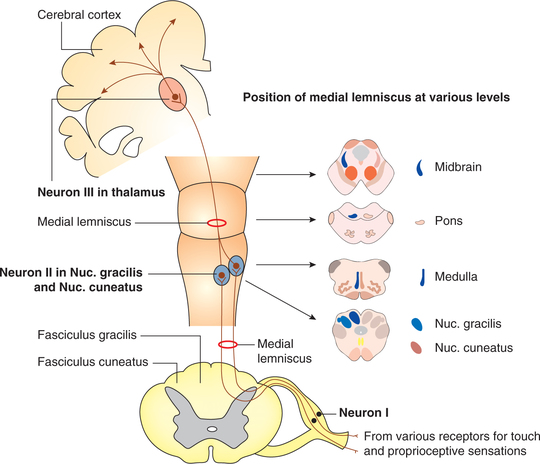
Fig. 10.3-9 Dorsal column sensory pathways. Fibres carrying touch, pressure and proprioceptive sensations are arranged as fasciculus gracilis and fasciculus cuneatus in dorsal column of spinal cord.
First-order neurons. These carry sensations of fine touch, tactile localization, two-point discrimination, vibration, pressure with intensity discrimination, and sense of position and proprioception. These fibres run in the spinal cord through dorsal column as fasciculus gracilis and fasciculus cuneatus (Fig. 10.3-9) to end in the nucleus gracilis and nucleus cuneatus, respectively. Many fibres of the dorsal columns, on their way up, terminate or relay in the dorsal grey horn.
Second-order neurons arise from the nucleus gracilis and nucleus cuneatus located in medulla oblongata. These decussate in the medulla as internal arcuate fibres and ascend up in the brain stem as medial lemniscus. The fibres of medial lemnisci terminate in the ventroposterior lateral (VPL) nucleus of thalamus.
Third-order neurons are from the VPL nucleus of thalamus. These ascend to the sensory cortex through the thalamic radiations.
Anterolateral sensory pathway
First-order neurons of anterolateral pathway carries sensations of pain, temperature and crude touch, and finally terminate on the nucleus proprius of the spinal cord.
Second-order neurons arise from the nucleus proprius and cross in the anterior commissure of the spinal cord to the opposite side in anterior and lateral white column in which they run upward to the brain as anterior and lateral spinothalamic tracts, respectively.
Anterior spinothalamic tract mainly carries sensation of crude touch, tickle and itch, etc. Higher up in the brain stem, the anterior spinothalamic tract joins the medial lemniscus of thalamus.
Lateral spinothalamic tract consists of the fibres carrying pain and temperature sensation, and higher up in the brain stem it is called the spinal lemniscus (Fig. 10.3-10).
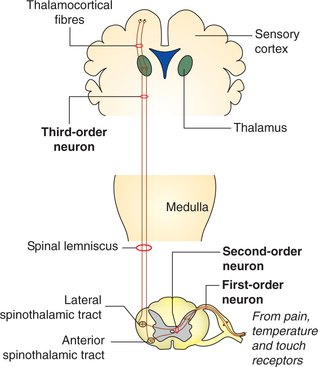
Third-order neurons arise from thalamus and carry tactile signals to the somatic sensory area of the cortex along with the fibres of dorsal column.
Dorsolateral column sensory pathway
Dorsolateral column pathways carry proprioceptive impulses arising from the muscles and joint receptors of the lower part of the body to the cerebellum. Recent investigations have shown that touch may also reach the cerebellum through these pathways.
First-order neurons receive impulses from muscle spindles, Golgi tendon organs and other proprioceptive receptors. Some fibres are related to end organs concerned with exteroceptive sensations (touch and pressure).
Second-order neurons are located in the junctional area between the ventral and dorsal grey column (laminae V, VI and VII) in the lumbar and sacral segments of spinal cord.
Pathway of sensations from face and oral cavity
The sensations of touch, pain and temperature from the face and oral cavity including teeth and proprioceptive information from the jaw muscles are carried by trigeminal nerve.
First-order neurons are located in the trigeminal ganglion, which is equivalent to dorsal nerve root ganglia in the spinal cord. The peripheral processes of these neurons form three divisions of the trigeminal nerve: ophthalmic, maxillary and mandibular which innervate different areas of the facial skin (Fig. 10.3-11). The central processes of these neurons of trigeminal ganglia terminate in different components of trigeminal sensory nucleus as (Fig. 10.3-12).
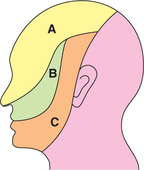
Fig. 10.3-11 The areas of face innervated by three divisions of the trigeminal nerve: A, ophthalmic; B, maxillary and C, mandibular.
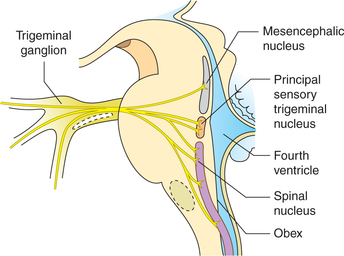
Fig. 10.3-12 Termination of central processes of trigeminal ganglion in three components of sensory nucleus of trigeminal nerve.
Principal sensory trigeminal nucleus, located in the pons, receives fibres carrying tactile sensations.
Spinal nucleus is elongated and extends down to the upper spinal cord. It receives fibres carrying pain and temperature sensations.
Mesencephalic nucleus, which extends from the pons into midbrain, receives fibres carrying proprioceptive information.
Second-order neurons. Axons of these neurons cross to the opposite side and ascend as trigeminal lemniscus to the ventroposterior medial (VPM) nucleus of thalamus.
Third-order neurons are located in the VPM nucleus of thalamus. All the sensations reaching this nucleus are carried primarily to sensory area of cerebral cortex by fibres passing through the posterior limb of internal capsule (superior thalamic radiations).
Role of thalamus in somatosensory system
Ventral posterior nucleus of thalamus is concerned with somatosensory system. It has two divisions:
For details, see page 482.
Somatic sensory cortex
Somatic sensory cortex is described under following headings:
For details, see page 474.
Encoding: recognition of type of sensation
As discussed above, the sensory receptors transduce all forms of sensory stimuli into a common type of neural signal, i.e. action potentials which are carried by the peripheral nerves and sensory tracts in the spinal cord and brain stem to the sensory cortex. The question arises how does brain differentiate between the action potentials generated from a touch receptor and a pain receptor, and interpret the sensation accordingly. It is believed that the sensory receptors themselves act as peripheral analysers. The intensity, location and quality of a stimulus are encoded as:
1 Encoding of stimulus intensity
The brain interprets different intensities of sensation (i.e. whether the touch is light or heavy; pain is mild, moderate or severe) by following two mechanisms:
A By frequency of action potential generated in sensory nerve fibres
The encoding of intensity of stimulus is related to the rate of impulse discharge in the sensory nerve fibres. The ’Weber-Fechner law’ states that the magnitude of sensation felt is directly proportional to the log of intensity of the stimulus.
2 Encoding of stimulus location
The stimulus location is recognized accurately due to point-to-point representation of the body in the somatosensory cortex. Therefore, when the sensory fibres are experimentally stimulated anywhere in their course to the cortex, the conscious sensation produced is referred to the location of the receptor. This principle is called the law of projection. Because of this reason, after amputation of a limb, sometime patient complains of intense pain in the absent limb (phantom limb). These sensations are produced due to irritation of the damaged nociceptive and proprioceptive afferents at the stump of amputated limb.
3 Encoding of stimulus quality
In general, action potentials are similar in all nerves, then why stimulation of a touch receptor causes sensation of touch and not of warmth. Similarly, stimulation of photo-receptors causes sensation of light and not of hearing. Stimulus quality is encoded by following mechanisms:
(i) Labelled line mechanism. It is the mechanism in which the stimulus quality is encoded by the particular neural pathway that is stimulated.
The specificity of sensory pathway from the receptors to the sensory cortex has been called the Muller's doctrine of specific nerve energies.
(ii) Pattern of activity within the neural pathway that is carrying information to the brain is used to encode stimulus quality.
(iii) Feature detectors are used in the most sophisticated mechanism of sensory coding. Feature detectors are neurons within the brain that integrate information from a variety of sensory fibres and fire to indicate the presence of a complex stimulus.