Somatic Motor System
 Highest level of motor control
Highest level of motor controlIntroduction
The execution, planning, co-ordination and adjustments of the movements of the body are under the influ ence of different parts of the nervous system which together constitute the somatic motor system, which is organized as three-tier system consisting of highest level of motor control, middle level of motor control and lowest level of motor control (Fig.10.4-1).
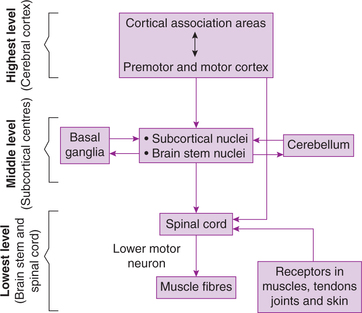
1. Highest level of motor control involves activi ties of various areas of cerebral cortex. It is mainly concerned with generation of the idea of voluntary movements (motor plan) and issuing the motor commands for their execution.
2. Middle level of motor control involves activities of various subcortical centres such as basal ganglia, some brain stem nuclei and cerebellum. The middle level of motor control is concerned with developing and perfecting each motor programme and subprogramme for bringing out a motor act. It also supervises the implementation of motor programme.
3. Lowest level of motor control is exerted by cranial nerve nuclei in brain stem and spinal cord. The spinal cord contains the final common pathway through which a movement is executed.
Feedback signals to CNS from the proprioceptors in muscles, joints, skin and other sensory receptors are used to adjust the motor commands during somatic motor activity.
In view of the above background, the somatic motor control system is discussed in detail under following headings:
Components of somatic motor control system
I Highest level of motor control
Motor cortex
Areas of motor cortex include (Fig.10.1-2):
• Primary motor cortex (Brodmann's area 4).
• Premotor cortex, which includes Brodmann's area 6, 8, 44 and 45.
Note. For details of motor cortex (see page 472).
Functional role of motor cortex in control of voluntary movements
Supplementary motor cortex is responsible for generating the idea for a movement. Then it plans the movements. Lateral cerebellum and basal ganglia are also involved in the planning and programming of movements.
Primary motor cortex is responsible for execution of movement.
Premotor cortex co-ordinates the voluntary activity.
Plasticity property of motor cortex. The motor system ‘learns by doing’, and performance improves with repetition. This involves synaptic plasticity. The motor cortex shows plasticity. This has been confirmed by PET and functional MRI (fMRI) in intact experimental animals and humans.
Descending motor pathways from motor cortex
A. Pyramidal tracts include the tracts which are constituted by the axons that transmit motor signals directly from the cortex to spinal cord (corticospinal tracts) and cranial nerve nucleus (corticobulbar tracts). Corticospinal fibres are divided into two tracts (Fig.10.1-3 see page 493).
Lateral corticospinal tract is constituted by 80% of fibres which cross the midline in the medul lary pyramids. These fibres are responsible for making skilled precision movements (e.g. the muscles that move the fingers and hands, and the muscles that produce speech) (lateral motor system).
Anterior corticospinal tract is formed by 20% uncrossed fibres which descend ipsilaterally in the ventral white column of the spinal cord. This pathway controls the axial (trunk muscles) and proximal limb muscles that are concerned with posture and equilibrium (medial motor system).
B. Extrapyramidal tracts. Extrapyramidal pathways are chiefly concerned with regulation of muscle tone, and posture and equilibrium.
II Middle level of motor control
The middle level of motor control specifies the postures and movements necessary to carry out the desired acts. It involves activities of basal ganglia, cerebellum and brain stem.
A. Basal ganglia
Physiologically basal ganglia include corpus striatum (caudate nucleus and lenticular nucleus having two parts, e.g. putamen and globus pallidus), subthalamic nucleus and substantia nigra (see page 476).
Role of basal ganglia in somatic motor activity
• Cognitive control of motor activity is executed by the basal ganglia.
• Timing and scaling of the intensity of the movements.
• Subconscious execution of some movements.
Note. For details of basal ganglia (see page 477).
B. Cerebellum
Functionally, the cerebellum has been divided into three divisions: vestibulocerebellum, spinocerebellum and corticocerebellum, which play important role in different motor activities.
1. Control of voluntary movements. Corticocerebellum, also called cerebral cerebellum, is intimately asso ciated with control of timing, rate, range (extent), duration, direction and strength of a movement. Corti co cerebellum performs this function by following actions:
(i) Comparator function. The cerebellum receives inputs from the command neurons about the sequential intended plan of movements for the next fraction of second. It also gets feedback (afferents) from the proprioceptive endings of muscles, tendons and joints about what actual movements result. All these information are integrated and the corrective signals are sent to the motor cortex. This happens through the closed feedback loop.
(ii) Damping action. Corticocerebellum sends impu lses to the cerebral cortex to discharge appropriate signals to the muscles so that any extra or exaggeration of muscular activity does not occur, and thus overshooting is prevented.
(iii) Timing and programming of the skilled movements is done by corticocerebellum feedback loop which modulates the motor command of pyramidal tracts through two-way communication.
(iv) Servomechanism. Cerebellum lets the cerebral cortex to discharge the signals which are already programmed and stored at sensory motor cortex, and does not influence much. However, if there is any disturbance or interference, the corticocerebellum immediately influences the cerebral cortex and corrects the movements. This action of corticocerebellum is known as servomechanism.
2. Control of body posture and equilibrium is done by vestibulocerebellum (see page 490).
3. Control of muscle tone and stretch reflex is the function of spinocerebellum (see page 490).
Note. For details about cerebellum (see page 486).
C. Brain stem
Reticular formation and vestibular nuclei are important components of the motor control system present in the brain stem.
Reticular formation. The motor control centres within the reticular formation are a relay station for all des cending motor commands, except those requiring the greatest precision, which are transferred directly from the cortex to spinal cord.
Vestibular nuclei. Vestibular nuclei, located within the brain stem and cerebellum, receive information from vestibular receptors via vestibular nerve fibres (8th cranial nerve). Vestibular system reflexes:
III Lowest level of motor control
The lowest level of motor control is exerted by motor nuclei of cranial nerves and spinal cord. The spinal cord contains the final common pathway through which a movement is executed.
Spinal cord
Motor neurons
Motor neurons of spinal cord present in ventral horn are:
1. Alpha motor neurons. These are the largest neurons. These are responsible for contraction of muscles in upper limbs, trunk and lower part of the body.
2. Gamma motor neurons. These neurons innervate the intrafusal fibres of the muscle spindles and are responsible for maintenance of muscle tone.
3. Interneurons. The interneurons actually receive the bulk of synaptic input that reaches the spinal cord, either as incoming sensory information or signals descending from higher centres in the brain.
4. Renshaw cells are interneurons that receive input from collateral branches of the axons of α-motor neurons. Their axons carry the impulses back to the cell bodies of the same α-motor neurons. These are inhibitory neurons which play an important role in synaptic inhibition at the spinal cord.
Skeletal muscles: the effector organ of somatic motor system
The motor activity, be it in the form of walking, physical labour, skilled work like typing, performing surgical operations or even expression of thoughts and feelings through gesture or speech, is a result of highly co-ordinated movements produced by the skeletal muscles. The skeletal muscles, thus, form the effector organ of the somatic motor system. The physiology of skeletal muscle has been discussed in Chapter 2.3. However, certain aspects which need elaboration of skeletal muscle as effector organ and are relevant to complete the study of somatic motor system are:
Motor unit
The motor unit is the functional module used by the motor control system to carry out a movement. The movement produced by a skeletal muscle basically depends upon the pattern and ratio of discharge of motor neurons supplying the muscle. A motor unit consists of single motor neuron and the muscle fibres that it innervates (see page 54).
Muscle sensors
Muscle sensors refer to the proprioceptors present in the muscles, tendons of muscles, joints, ligaments and fasciae. Proprioceptors are the receptors which give information about change in position of different parts of the body in space, especially joints or tension of muscles at any given moment. The muscle sensors are:
In addition to the above proprioceptors, the laby rinth also contains proprioceptors.
Muscle tone
Definition. Muscle tone is defined as resistance offered to active or passive stretch. In other words, muscle tone refers to sustained partial state of contraction of the muscle under resting condition, i.e. a state of partial tetanus. The muscle tone is present in all the muscles, but is well pronounced in the extensor muscles, i.e. antigravity muscles.
Basis of muscle tone. The muscle tone is purely a function of myotactic reflex (stretch reflex) occurring due to low frequency and asynchronous discharge of γ-motor neurons (see page 521). The discharge is out of phase with each other, which ultimately merges to produce smooth muscle contraction.
Anomalies of muscle tone are hypotonia and hypertonia.
1. Hypotonia refers to decrease in muscle tone. The hypotonic or also called flaccid muscle offers little or no resistance to stretching. The muscles are generally hypotonic when the rate of γ-efferent discharge is low, i.e. when stretch reflex becomes hypoactive.
2. Hypertonia refers to increase in muscle tone. The hypertonic or spastic muscle offers high resistance to stretch. The muscles are generally hypertonic when the rate of γ-efferent discharge is high, i.e. when stretch reflex becomes hyperactive. Hypertonia is of two types:
• Spasticity refers to hypertonia which is confined to only one group of muscles. For example, lesions of internal capsule and upper motor neuron lesions produce spasticity.
• Rigidity refers to hypertonia which involves both groups of muscles, i.e. extensor as well as flexors equally. For example, lesions of basal ganglia produce rigidity.
Reflex activity
General considerations
A reflex is an involuntary response to a peripheral nervous stimulation. In other words, it is a mechanism by which sensory impulse is automatically converted into a motor effect through the involvement of CNS. It is a type of protective mechanism which tries to protect the body from irreparable damage.
Reflex arc
The pathway for a reflex activity is called reflex arc. It consists of (Fig.10.4-2):
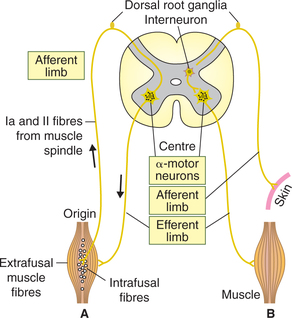
1. Afferent limb of each reflex arc consists of a receptor and an afferent or sensory nerve.
2. Centre. This is the part of CNS (spinal cord or brain) where afferent limb ends and either synapses directly with efferent motor neuron or establishes con nection with the efferent neuron via interneurons (internuncial or intercalated neurons).
3. Efferent limb of a reflex arc consists of an efferent or motor nerve and an effector organ.
Classification of reflexes
Reflexes can be classified in different ways.
I Depending upon the number of synapses (Fig.10.4-2)
1. Monosynaptic reflexes, e.g. stretch reflexes (biceps, triceps or knee jerk).
2. Disynaptic reflexes, i.e. one interneuron is placed between afferent and efferent neurons of the reflex arc, e.g. inverse stretch reflex.
3. Polysynaptic reflexes are characterized by more than one interneuron placed between afferent and efferent neurons of the reflex arc, e.g. withdrawal reflex, cross flexor reflex and cross extensor reflex.
Spinal cord reflexes
Spinal cord reflexes, according to the receptors from which they originate the spinal cord reflexes, can be categorized into muscle reflexes and cutaneous reflexes.
Muscle reflexes. Two important reflexes which ori ginate in muscles are:
Cutaneous reflexes. The most important of the cutaneous reflexes is:
1 Stretch reflex
Stretch reflex, also known as myotactic reflex, refers to reflex contraction of a muscle that is stretched. It is the best known monosynaptic reflex in the body. Stimulus that evokes the reflex response is ‘stretch’ to the muscle. Stretch reflex is well developed in antigravity muscles such as extensor group of muscles of legs and flexor groups of muscles of arm. Examples of stretch reflexes are knee jerk, ankle jerk, biceps jerk and triceps jerk (see page 524).
Reflex arc of stretch reflex (Fig.10.4-2)
1. Afferent limb. Afferent limb consists of receptor muscle spindle and afferent nerve.
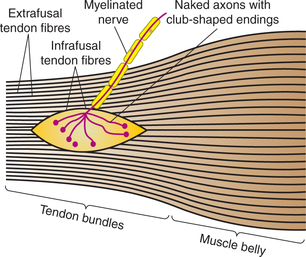
Structure of muscle spindle (Fig.10.4-3). Each muscle spindle consists of 3–10 small muscle fibres (called intrafusal muscle fibres) encapsulated in a thin connective tissue capsule containing fluid. The muscle spindles are present in between and parallel to the extrafusal fibres. Either end of the muscle spindle is attached to the endomysium of the extrafusal muscle fibres.
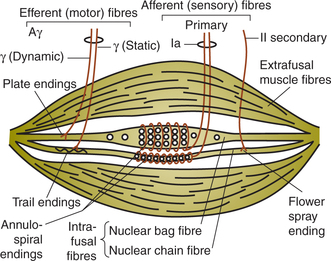
Intrafusal muscle fibres consist of a central non-contractile portion and is devoid of striations. The central part of each intrafusal fibre is sensory portion. Intrafusal fibres are of two types:
Nerve supply of the muscle spindle. The muscle spindle is innervated by both sensory and motor nerve fibres.
(i) Sensory nerve supply. There are two types of sensory fibres:
Group Ia fibres, also known as primary sensory endings, supplying central receptor portions of both nuclear bags as well as nuclear chain fibres. The primary endings are stimulated when the muscle spindle is stretched.
Type II fibres, also known as secondary sensory end ings, innervate the receptor portion of mainly nuclear chain fibres. These nerve endings respond mainly to sustained stretch, therefore measure the muscle length.
(ii) Motor supply. The efferent fibres to the muscle spindle are called gamma fibres because their axons belong to the Aγ group of fibres. There are two types of gamma fibres:
• Dynamic gamma fibres primarily innervate the striated poles of nuclear bag fibres. These fibres increase the sensitivity of the Ia afferent fibres to stretch.
• Static gamma fibres primarily innervate the striated poles of nuclear chain fibres. They increase the tonic activity in the Ia afferent fibres at any given muscle length.
• Afferent nerve. The afferent nerve fibres emerging from the muscle spindle travel along the spinal nerve, enter the spinal cord through the dorsal root and send branches to every α-motor neuron that goes to the muscle from which the Ia originated.
2. Centre. Centre for stretch reflex is the ventral grey horn area where the afferent nerve ends and syna pses directly with the α-motor neuron.
Reciprocal innervation in stretch reflex
The stretch reflex is characterized by reciprocal innervation, i.e. excitation of one group of muscles is associated with inhibition of the antagonistic group of muscles on the same side, allowing the agonistic muscles to contract without interference. Reciprocal innervation is one of the important features of both flexor and extensor reflexes.
Pathway of reciprocal innervation is biphasic. A collateral from each Ia fibre passes in the spinal cord to an inhibitory interneuron (Golgi bottle neuron) that synapses directly on one of the motor neurons supplying the antagonist muscles. Reciprocal innervation is very important in spinal reflexes, which are involved in locomotion.
Role of γ-motor neurons
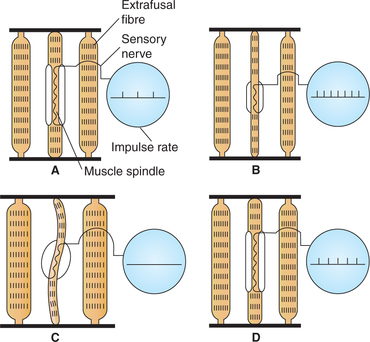
As discussed above, the firing rate of primary nerve endings (Ia fibres) increases when the muscle is stretched (Fig.10.4-4B), and causes reflex contraction of the muscle by increased α-motor neuron activity. Contraction of the extrafusal muscle fibres makes the muscle spindle slack and decrease the firing rate of Ia fibres (Fig.10.4-4C). The decreased rate of Ia afferent discharge that occurs during muscle contraction is called unloading of muscle spindle and is functionally disadvantageous because the CNS stops receiving information about the rate and extent of muscle shortening. However, by the activity of γ-motor neurons this unloading is prevented (Fig.10.4-4D). The γ-motor neurons cause the striated poles of intrafusal fibres of muscle spindle to shorten along with shortening of extrafusal fibres during muscle contraction. As a result of contraction of the striated polar regions of intrafusal fibres the central receptor region of the intrafusal fibres remains stretched during muscle contraction, and unloading does not occur. In this way the γ-motor neuron activity adjusts the sensitivity of the muscle spindle so that it will respond appropriately during muscle contraction as well. Further, the γ-motor neurons control both dynamic as well as static activity of muscle spindle.
Higher control of stretch reflex
Though the stretch reflex is a spinal reflex, the activity in the reflex arc can be modified (inhibited or facilitated) by higher centres through their influence on the nerve fibres involved in stretch reflex.
Some of the important brain areas that facilitate or inhibit the stretch reflex are (Fig.10.4.5):
• Facilitatory reticular formation is a large area in the brain stem which discharges spontaneously in response to afferent input. This increases discharge of γ-motor neurons and stretch reflex becomes hyperactive.
• Inhibitory reticular formation is a small area which does not discharge spontaneously. It acts by inhibiting γ-efferent neuron discharge, thereby decreasing the spindle sensitivity.
• Cerebral motor cortex and cerebellum reflexly inhibits the stretch reflex by stimulating the inhib itory reticular formation.
Other factors which influence γ-efferent discharge are:
• Anxiety causes an increased discharge, a fact that probably explains the hyperactive tendon reflexes sometimes seen in anxious patients.
• Stimulation of skin, especially by noxious agents, increases γ-efferent discharge. This fact is sometimes used as reinforcement to elicit deep tendon reflexes (such as knee jerk), which are not being elicited otherwise. For it, the individual is asked to pull the hands apart when the flexed fingers are hooked together; this facilitates the knee jerk (Jendrassik's manoeuvre). It is contributed to increased γ-efferent discharge initiated by afferent impulses from the hands.
2 Golgi tendon reflex (disynaptic reflex)
The Golgi tendon reflex, also called ‘inverse stretch reflex’, is a disynaptic reflex. The receptors involved are the Golgi tendon organs.
Golgi tendon organs
Golgi tendon organs (Fig.10.4-5) are high-threshold stretch receptors located in the tendons and musculoaponeurotic junction. They are placed in series between the muscle fibres and the tendon, and are thus stretched whenever the muscle contracts.
The Golgi tendon organs are supplied by Ib type sensory nerve fibres.
Pathway and activity of reflex (Fig.10.4-6)
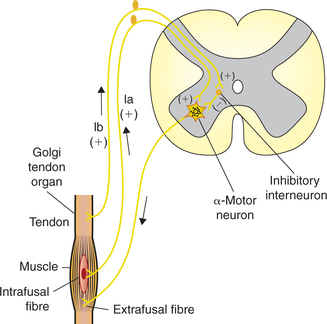
• When a muscle contracts, the muscle tension increases. The Golgi tendon organ detects the muscle tension and sends impulses through afferent (group Ib) fibres which enter the spinal cord through dorsal root.
• In the spinal cord, the group Ib afferents stimulate the inhibitory interneurons.
• The inhibitory interneurons in turn release inhib itory mediator glycine, which inhibits α-motor neurons and causes relaxation of the muscle that was originally contracted.
• At the same time, due to reciprocal innervation, the antagonistic muscles are excited.
Physiological role or the functions of Golgi tendon reflex has been described as a protective reflex in which a strong and potentially damaging muscle force reflex ively inhibits the muscle, causing the muscle to lengthen instead of trying to maintain the force and risking damage.
Clasp–knife reflex refers to an exaggerated form of the Golgi tendon reflex, which can occur with disease of the corticospinal tracts (hypertonicity or spasticity). For example, when the arm is hypertonic, the increa sed sensitivity of the muscle spindles in the extensor muscles (triceps) causes resistance to flexion of the arm. Eventually, tension in the triceps increases to the point at which it activates Golgi tendon reflex, causing triceps to relax and the arm to flex closed like a jack knife, hence the name clasp–knife reflex. The physiological name for it is lengthening reaction because it is the response of a spastic muscle to lengthening.
3 Withdrawal reflex (polysynaptic reflex)
Definition and receptors
Definition. Withdrawal reflex, also known as flexor reflex, is a cutaneous reflex which occurs in response to nociceptive (pain) stimuli and is characterized by removal of a body part from painful stimulus.
Receptors for withdrawal reflex are nociceptors located in free nerve endings of Aδ and C fibres.
Pathway (reflex arc) of withdrawal reflex (Fig.10.4-7)
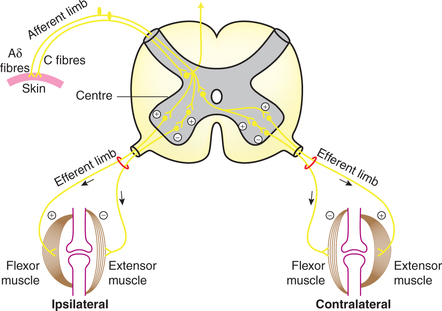
• The pain fibres carrying impulses, upon entering the spinal cord, synapse on many interneurons. Some of these also convey information to CNS. Others form several reflex pathways.
– Some of the interneurons project onto α-motor neurons on the ipsilateral side and stimulate the flexors which withdraw the limbs.
– Some of the interneurons form inhibitory pathway and terminate on α-motor neurons supplying the extensor muscles on the ipsilateral side producing their relaxation.
– Some of the interneurons cross to the opposite side of spinal cord and end on the α-motor neurons supplying the extensors on contralateral side. In case of need, this pathway produces extension of the opposite limbs (crossed extensor reflex).
Effector organs
The effector organs of the withdrawal reflex are the skeletal muscles that cause withdrawal of the limb.
Response in withdrawal reflex
The reflex response to a painful stimulus varies from just withdrawal of the affected part to withdrawal of the whole body depending upon the strength of painful stimulus and location of the stimulus.
Crossed extensor reflex response (two-limb response). When a strong stimulus is applied to a limb, the response includes not only flexion and withdrawal of the limb but also extension of the opposite limb. This crossed extensor response is produced by the interneuronal pathway which crosses to the opp osite side of spinal cord. In lower limbs crossed extensor reflex allows one limb to support the body while other is raised off the ground.
Clinical reflexes
Clinically the reflexes can be grouped as:
I Physiological reflexes
1. Superficial reflexes. These reflexes are initiated in response to stimulation of receptors on skin (cutaneous reflexes, e.g. plantar, abdominal, cremas te ric, bulbocavernous) or mucous membranes (muc ous membrane reflexes), e.g. corneal, conjunctival and palatal reflex. The superficial reflexes are summarized in Table 10.4-1.
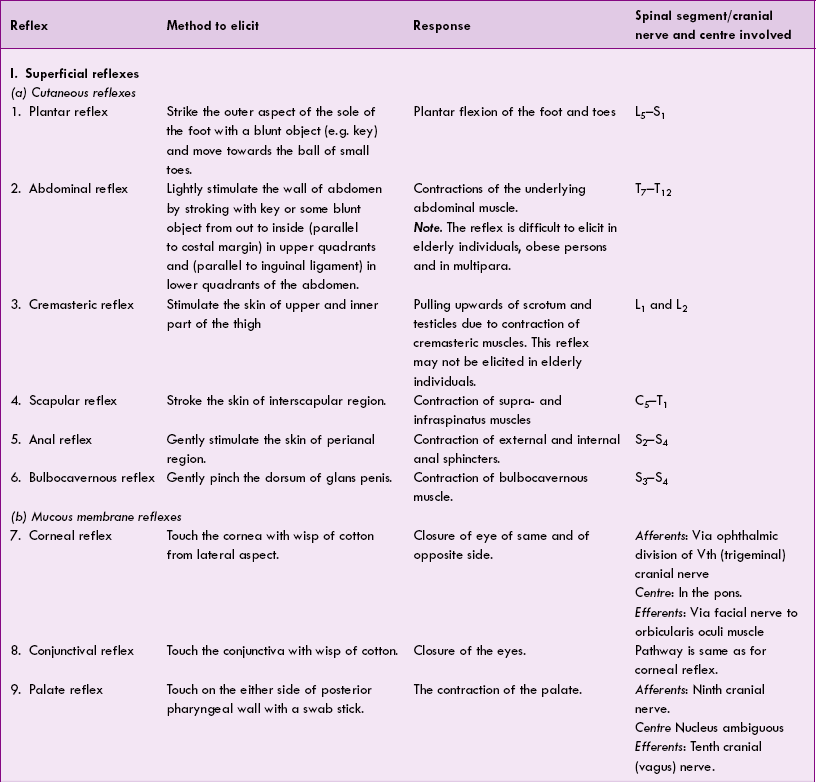


2. Deep reflexes. These reflexes are basically stretch reflexes and are elicited on stroking a tendon, so are called tendon reflexes (e.g. knee jerk, ankle jerk); the stretch reflex has been described in detail on page 520; however, the various clinically known stretch reflexes are summarized in Table 10.4-1.
3. Visceral reflexes are elicited from the visceral organ or at least one part of the reflex arc is formed by autonomic nerve, e.g. carotid sinus reflex (see page 183), micturition reflex (see page 301). A few of clinically known visceral reflexes are summarized in Table 10.4-1.
Regulation of posture
Physiologically posture refers to subconscious adjustment of tone in different muscles so as to maintain balance during displacement of the body caused by gravity or acceleration.
For the purpose of understanding, the regulation of posture can be discussed under two main headings:
• Mechanisms involved in maintenance of posture and
• Role of different regions of nervous system in maintenance of posture.
Mechanisms involved in maintenance of posture
The two main mechanisms involved in maintenance of posture are:
Role of tone in antigravity muscles in maintenance of posture
Largely, the posture is maintained through reflex adjustments of tone in the antigravity muscles. The basic postural reflex involved in the control of muscle tone is stretch reflex, which is described in detail.
Posture control is required not only for hold ing the body in erect position but also for fixation of the body parts over adjoining body segments. The centre of grav ity of head passes in front of the atlanto-occipital joint. Thus head has always got a tendency to roll forwards. To hold the head in erect position cervico-occipital muscles are to be maintained in a state of constant tension.
In the upright position, gravity tends to displace the body downward, stretching quadriceps muscles as the legs flex at the knees. The muscle stretch evokes discharge from the muscle spindles of the quadriceps leading to its reflex contraction. This ensures that the knee joints, i.e. the main weight-bearing joints, do not give way under the effect of gravity. This maintains the leg as a pillar of support and thus counteracts the gravitational displacement of the body.
In general, the antigravity muscles of the body are endowed with a somewhat higher muscle tone than the other muscles of the body.
In human beings, flexors of upper extremity and extensors of lower extremity are the main antigravity muscles. Retractors of neck, the elevators of joint, supraspinatus, the extensors of back, rectal muscles of abdominal wall, extensors of knee and ankle are the muscles which exhibit greatest degree of tone. When these muscles completely relax (as in unconscious person), the body collapses.
Various postural reflexes (described below) influence the medial motor system and the motor neurons of antigravity muscles.
Maintenance of muscle tone
Stretch reflex, as mentioned earlier, plays the main role in maintenance of muscle tone (page 522).
The centre for muscle tone lies in anterior motor neurons (α-motor neurons) of spinal cord, which are stimulated through a constant γ-motor neuron discharge. Activity of γ- and α-motor neurons is modified by both extrapyramidal and pyramidal fibres which terminate on them (directly or through interneurons). Mainly extrapyramidal system is responsible for maintaining tone (Fig.10.4-8).
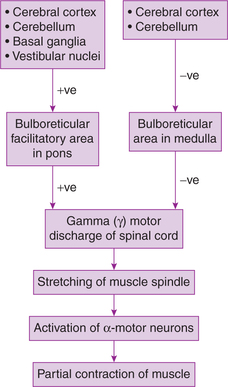
Postural reflexes
The postural reflexes help to maintain the body in upright and balanced position. They also provide adjustments necessary to maintain a stable posture during voluntary activity.
Reflex arc of postural reflexes is as follows (Fig.10.4-9):
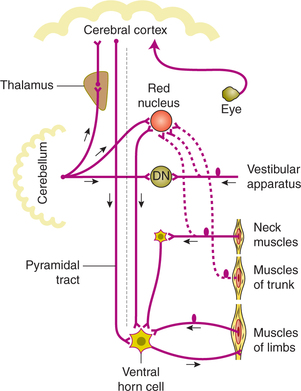
• Afferent pathways of reflex arc come from the eyes, the vestibular apparatus and the proprioceptors.
• Integrating centres are formed by neuronal networks in the brain stem and spinal cord.
• Efferent pathways consist of α-motor neurons supplying the various skeletal muscles which form the effector organs.
Types of postural reflexes. Broadly, postural reflexes are of two types.
• Static reflexes. These are elicited by gravitational pull and involve sustained contraction of muscles.
• Statokinetic reflexes. These reflexes, also called phasic reflexes, are elicited by acceleratory displacement of the body. They maintain a stable postural background for voluntary activity.
• Both these types of postural reflexes are integrated at various levels in the CNS from the spinal cord to cerebral cortex and are affected largely by pyramidal pathways.
The various postural reflexes are summarized in Table 10.4-2.
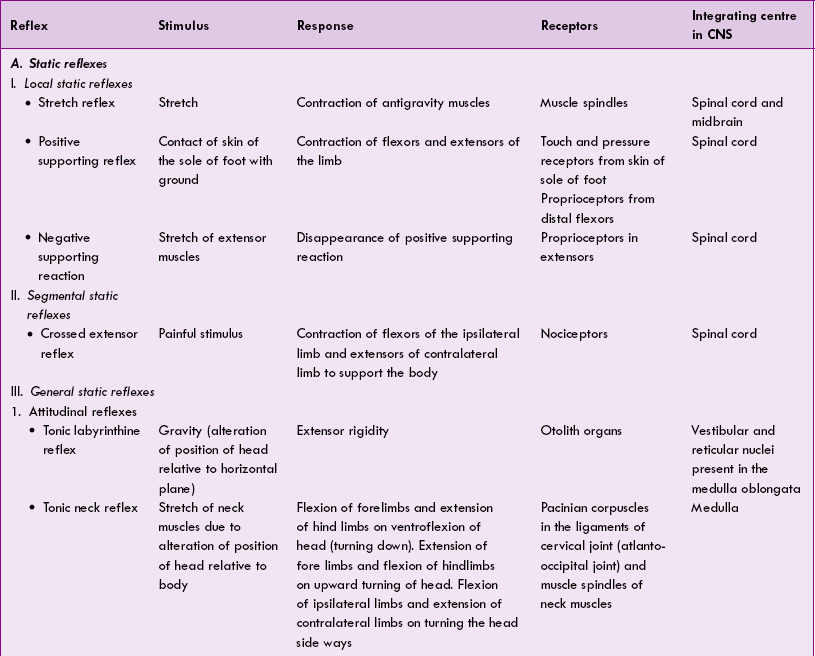

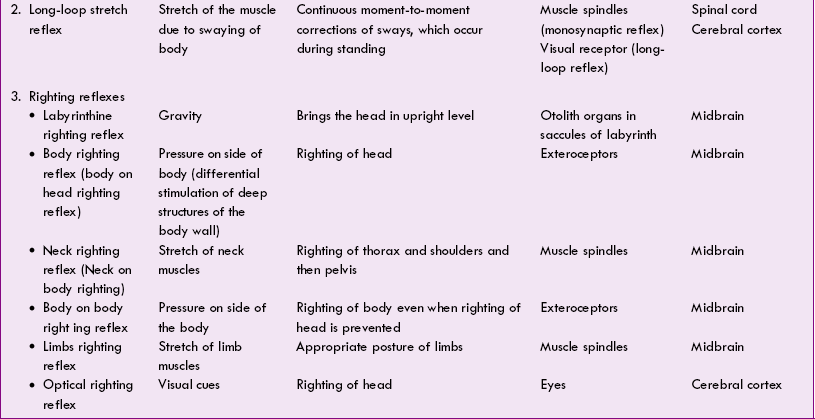
Role of different regions of nervous system for maintenance of posture
The role of different regions of the nervous system for the maintenance of posture can be experimentally investigated (usually in a cat) by producing transection in the neuraxis at various levels.
Role of spinal cord: spinal animal
Spinal animal
The role of spinal cord in the maintenance of posture can be studied in a spinal animal. The spinal animal can be produced by a transection in the spinal cord at cervical region and respiration is maintained artificially by respiratory pump. In this way, most of the cord functions can be studied in a spinal animal. If spinal cord is transected below the origin of phrenic nerve in the mid-thoracic region then diaphragmatic respiration continues and so the artificial respiration is not required.
Effects of spinal cord transection. As described earlier, the effects produced by complete spinal cord transection occur in three stages. For details (see page 494).
Posture in spinal animal during stage of reflex activity
Except the basic stretch reflex and supporting reflexes which are integrated in spinal cord (Table 10.4-2), all other postural reflexes are absent, as they require the integrity of upper motor neurons coming from various levels of neuraxis.
• Though stretch reflex and supporting reactions are present, but they are very weak and cannot support the weight of the animal. Therefore, the animal cannot stand on its legs.
• Muscle tone returns first in flexor muscles, therefore, flexors become less hypotonic than extensors producing paraplegia in flexion (both lower limbs are in state of flexion).
Role of brain stem: bulbospinal animal or decerebrate animal
Decerebrate animal
Decerebrate animal is one in whom the brain stem is transected at intercollicular level (between superior and inferior colliculi).
Characteristic features of a decerebrate animal are given below.
1. Decerebrate rigidity, i.e. spasticity in all the antigravity muscles occurs immediately after decerebration. Characteristic features of decerebrate rigidity (Fig.10.4-10) are:
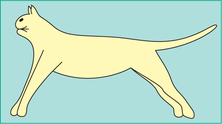
• Hyperextension of all the four limbs,
• Dorsiflexion (hyperextension) of tail and head and
• Extreme hyperextension of the spine (opisthoto nus) produces concave configuration of the back.
2. Postural reflexes present in decerebrate animal are those which have their integration centre in the spinal cord or medulla or pons. These include:
3. Righting reflexes are absent, therefore, decerebrate animal can stand on its four legs but slight displacement causes the decerebrate animal to topple over.
Role of midbrain: mesencephalic animal or high-decerebrate animal
Mesencephalic or high-decerebrate animal is one in whom the brain stem is transected at the rostral border of midbrain.
Characteristic features of a mesencephalic animal are given below.
1. Decerebrate rigidity, similar to that of bulbospinal animal, is present but it disappears when the limb is performing a reflex activity.
3. Animal cannot only stand but also typical quadrupedal walking movements can be reflexly performed.
4. Righting reflexes having integration centre in the midbrain are present. These include:
By means of the righting reflexes, the midbrain animal can bring its head right way up and get the body into the erect position under all circumstances.
Role of cerebellum
Spinocerebellum regulates the postural reflexes by modifying muscle tone. The γ-motor neurons reflexly modify the activity of α-motor neurons and thus regulate the muscle tone. Thus, cerebellum forms an important site of linkage of α–γ systems responsible for muscle tone.
Role of basal ganglia: decorticate animal
Decorticate animal is one in whom the whole cerebral cortex is removed but the basal ganglia and brain stem are left intact.
Postural characteristics of a decorticate animal
Moderate rigidity is present due to loss of the cortical area that inhibits spinal γ-motor neurons discharge via reticular formation. It is seen only when the animal is at rest. This is because the basal ganglia, which are intact in decorticate animal, activate the descending inhibitory reticular formation and thereby prevent hypertonia.
Typical posture in decorticate man consists of full extension of legs, arms lying across the chest, with semi flexion at elbow, slight pronation of forearm, and flexion of wrist and fingers (Fig.10.4-11).
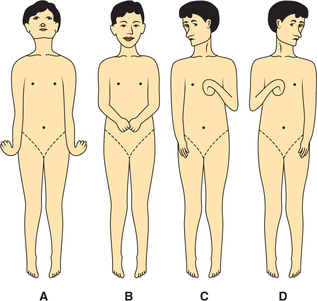
Postural reflexes. In decorticate man or animal following reflexes can be elicited.
Mechanism of standing in man
As mentioned earlier, the tall human being has to stand over a narrow base of feet; therefore, maintenance of erect posture is more difficult than the quadruped animals. Mechanisms which play important role in erect standing posture are discussed below.
Reflex adjustment in muscle tone of antigravity muscles undoubtedly plays most important role in making the man stand erect. A continued contraction of most of the trunk and leg muscles keeps the posture upright.
The effect of gravity has to be opposed by reflex contraction of some of the antigravity muscles all the time, otherwise a standing man may fall in any direction (forwards, backwards or sideways). The different antigravity muscles which contract sufficiently to restore the balance to oppose the fall under various circumstances are:
• Extensors of the trunk and flexors of the legs when the body sways forward.
• Recti abdominis and leg extensors when the body sways backward.
• Contralateral external oblique abdominal muscles maintain the balance when the body leans sideways.
• Head has a tendency to sway more than the trunk. Therefore, to hold the head in erect position the cervicooccipital muscles are to be maintained in a state of constant tension.
Reflex changes in antigravity muscles described above are induced by:
• Stretch receptors in the trunk and leg muscles.
• Visual afferents also play important role in reflex of upright posture in man. This is why, when the eyes are closed, the upright posture is less steady and there occurs more swaying (bending) of the trunk.
• Vestibular afferents help in maintaining the erect position of head.
Vestibular apparatus and equilibrium
Functional anatomy
The internal ear or labyrinth is situated in the petrouspart of the temporal bone. It consists of bony labyrinth andmembranous labyrinth (Fig.10.4-12).
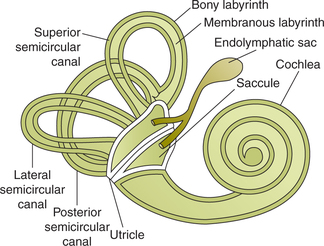
Bony labyrinth consists of vestibule, semicircular canals and the cochlea.
Membranous labyrinth is lodged within the bony labyrinth. It consists of:
• Utricle and saccule, which are lodged in the bony vestibule, and are collectively called otolith organs,
• Three semicircular ducts, which lie within the body of semicircular canals, and
Vestibular apparatus
The semicircular canals and the utricle and saccule collectively form the vestibular apparatus. The vesti bular apparatus plays important role in maintaining posture and equilibrium.
Semicircular canals. The three semicircular canals are arranged at right angles to each other so that all the three planes are represented as (Fig.10.4-13) follows.
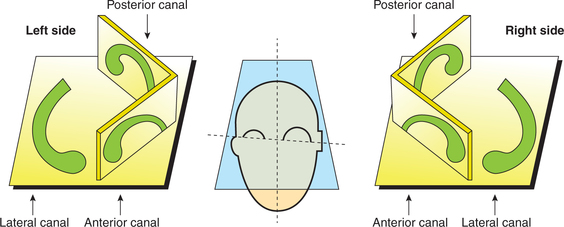
• Anterior semicircular canal is vertical and placed at right angles to the long axis of the petrous bone.
• Posterior semicircular canal is also vertical but is placed parallel to the long axis of the petrous bone.
• Lateral semicircular canal is set in a horizontal position making an angle of about 30° with the horizontal plane.
• It is important to note that:
– The right anterior and left posterior canals lie in the one plane, while the left anterior and right posterior canals lie in the other plane.
– One end of each semicircular canal is dilated and is called ampulla. The ampulla contains the receptor organ known as crista ampullaris (Fig.10.4-14).
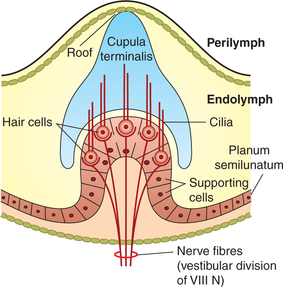
– The semicircular canals open into the utricle by means of five orifices. The ampullary end of each canal and narrow end of horizontal canal open inde pendently, while narrow ends of anterior and posterior canals open jointly by a common orifice.
Otolith organ refers combined to the two vestibular sacs called the utricle and saccule.
Utricle is the larger of the two vestibular sacs in which open the three semicircular canals. It is indirectly connected to the saccule and ductus endolymphaticus. The ductus endolymphaticus ends in a small bag-like structure called endolymphatic sac (Fig.10.4-12).
Saccule is a globular sac which is connected to utricle indirectly through the ductus utriculosaccularis, and to the cochlea via the ductus reunion (Fig.10.4-12).
Vestibular receptors
The receptor cells of the vestibular system are called hair cells which are slowly adapting mechanoreceptors.
Hair cells
The vestibular hair cells are of two types:
Cilia of hair cells (Fig.10.4-15). From the apex of each hair cell arise about 40–60 cilia. These cilia are called stereocilia which are motile. A large, non-motile cilium located at one end of the cell is called kinocilium.
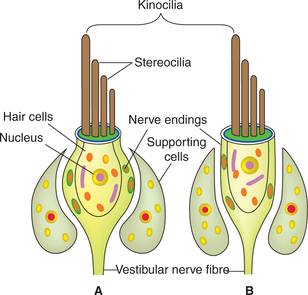
Activity of hair cells. The hair cells are polarized cells. When the stereocilia are bent towards the kinocilium, the cell depolarizes and when the stereocilia are bent away from the kinocilium, the cell hyperpolarizes. The changes in the activity of hair cells are conveyed to CNS by affe rent fibres which form the vestibular part of 8th cranial nerve.
Receptors in semicircular canals
The receptors, i.e. the hair cells of the semicircular canals, are located on a raised mass of tissue in the ampulla called the crista ampullaris. The crista ampul laris consists of following structures (Fig.10.4-14):
• Neuroepithelium is formed by the hair cells (described above) which are innervated by the primary afferent fibres of vestibular nerve.
• Secretory epithelial cells surround the hair cells.
• Cupula is dome-shaped large mass of gelatinous material in which are embedded the cilia arising from the hair cells.
Stimulation of receptors in semicircular canals. The movements produced in the endolymph by the angular movements of head pushes the cupula backwards, causing the cilia of hair cells to bend. Depending upon whether the stereocilia are pushed towards or away from the kinocilium, the hair cell depolarizes or hyperpolarizes.
It is important to note that cupula is unaffected by linear acceleration force, as it has the same specific gravity as the endolymph.
Receptors in otolith organs
The receptors (hair cells) of the otolith organs (utricle and saccule) are located in a raised mass of tissue called macula. The macula consists of (Fig.10.4-16):
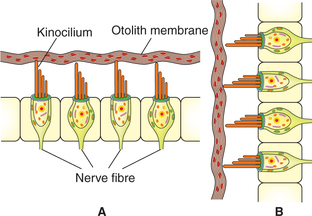
Fig. 10.4-16 Structure of macula: A, utricle (horizontally placed) and B, saccule (vertically placed).
• Neuroepithelium of macula like that of crista ampullaris is formed by hair cells (both type I and II).
• Otolith membrane. It is a flat gelatinous membrane covering the hair cells. This contains crystals of calcium carbonate called otoliths or otogonia which increase its specific gravity as compared to endolymph. The cilia of hair cells project in the gelatinous membrane.
Stimulation of receptors in otolith organs. The movements produced in the otolith membrane by linear acceleration of the head cause the cilia of hair cells to bend. This leads to excitation of vestibular afferents supplying these cells.
Orientation of the macula is (Fig.10.4-16):
• Macula of utricle is directed horizontally so its cilia are in vertical plane, which are stimulated by horizontally directed linear acceleration, e.g. moving in a car.
• Macula of saccule is directed vertically, so its cilia are in a horizontal plane and are stimulated by vertically directed linear acceleration, e.g. moving in a lift.
Mechanism of functioning of vestibular apparatus
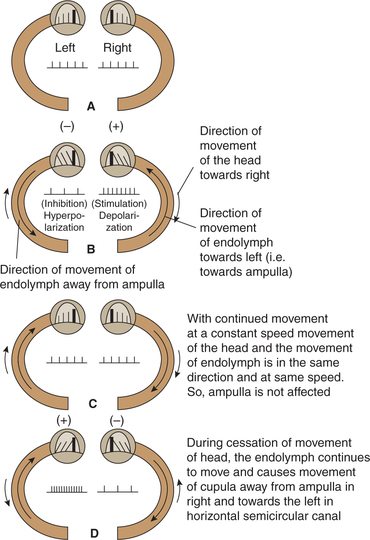
Salient features of functioning of semicircular canals
• Receptors of semicircular canals are stimulated by rotatory movements or angular acceleration of the head.
• Semicircular canals are oriented in three different planes, so movement of the head in any direction generates a unique pattern of activity.
• Receptors of horizontal canals are stimulated during rotation of head in vertical axis, while receptors of vertical canals are stimulated during rotation of head in anteroposterior or transverse axis.
• Receptors of semicircular canals are stimulated only at the beginning and at the stoppage of rotatory movements (Fig.10.4-17).
Salient features of functioning of utricle and saccule
• These provide information about linear acceleration and change in head position relative to the force of gravity.
• The effective stimulus being the pull of gravity on the otolith membrane. These receptors discharge tonically even in the absence of head movement.
• During linear acceleration of the head, the otolith membrane having more specific gravity lags behind due to inertia. This causes cilia of hair cells embedded in otolith membrane to bend. This leads to excitation of vestibular afferents supplying these cells.
• The hair cells of utricle are stimula ted during dorsiflexion or ventroflexion of the head, i.e. by nodding the head up and down (as in signifying yes).
• The hair cells of saccule are stimulated when the head is tilted sideways.
Vestibular pathways
First-order neurons. The afferent fibres carrying impulses from the hair cells are dendrites of the bipolar cells having their cell bodies in the vestibular or Scarpa's ganglion situated in the internal auditory meatus. Axons of these cells form the vestibular division of vestibulocochlear (8th cranial) nerve, which enters the medulla. These axons end in the vestibular nuclei of the same side (Fig.10.4-18).
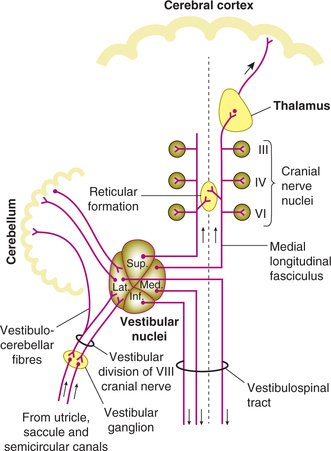
Vestibular nuclei. The vestibular nuclei contain cell bodies of the second-order neurons of the vestibular pathway.
Afferent connections. In addition to the main afferents from vestibular apparatus, the vestibular nuclei also receive inhibitory fibres from the cerebrum and cerebellum.
Vestibular reflexes
1 Vestibulo-ocular reflex
The vestibulo-ocular reflex maintains visual fixation during movements of the head by producing reflex nystagmus.
Nystagmus. For example, when the head is rotated to the left, the eyes move slowly towards the right in order to keep the image on the fovea. When the eyes have rotated as far as they can, they are rapidly returned to the centre of the socket. These reflex movements of the eyes are called nystagmus.
Functions of vestibular apparatus
Applied aspects
The motion sickness is a symptom complex occurring due to excessive and repeated stimulation of vestibular apparatus while travelling in automobile, ship, aircraft or spacecraft.
Characteristic features of motion sickness are unpleasant sensation of rotation accompanied by nausea, vomiting, sweating, pallor, salivation, headache, disorientation and even diarrhoea.
Prevention. Motion sickness can be prevented by taking antiemetic drugs such as Avomine, and by avoiding greasy and bulky food before travelling.