2 A biological basis
Touching, stroking and rubbing the skin is an integral part of our lives. Through these actions, we communicate and transmit feelings and emotions to one another. Skin rubbing and caressing can become more formalised and take the form of a massage, to aid relaxation and promote a feeling of well being. For the massage to progress to a more specific therapeutic outcome it must be directed towards a more specific purpose and be aimed at promoting physiological and/or psychological change. It must therefore be based on sound theoretical principles. The structure, function and dysfunction of the body must be understood so that clinical decisions may be made and appropriate techniques selected to induce desirable change. This chapter discusses a range of anatomical, physiological and pathological issues, providing an essential basis for the therapeutic use of massage. The chapter covers topic areas specific to massage; extrinsic material can be found in greater detail in other texts.
Skin
The skin is the tough, waterproof, external surface of the body. It is also known as the integument, or cutaneous membrane. What makes the skin such a fascinating structure is its ability to combine a protective and insulating function of great physiological significance with an important role in communication. It conveys emotional responses through its vascular changes (flushing with pleasure or embarrassment, blanching with shock or fear, for example); it plays a part in expressing words or emotions through its transmission of the coordinated contractions of the underlying muscles of facial expression and is essential for tactile communication. By being sensitive to these reactions, the therapist may learn a considerable amount about the patient's psychological and emotional state and can modify massage treatment subtly and appropriately. By using her awareness of these properties, the therapist can communicate attitudes essential to obtaining trust and relaxation—respect, friendliness, approachability, concern and understanding—even before touching the skin, thereby enhancing her personal approach.
Skin varies in thickness between 0.5 and 2–3 mm. It is of vital importance for survival: it forms a protective layer for deeper structures and is a richly innervated sensory organ (the largest in the body) to feed information about the environment to the nervous system, thus acting as a warning system and protective mechanism. It regulates temperature due to its neurovascular mechanisms and insulating properties (fat, found in the hypodermis, conducts heat two-thirds less efficiently than other tissues); prevents fluid loss; allows excretion and absorption of substances; and acts as a chemical and bacterial barrier. The rate of blood flow into the vascular plexi associated with the skin varies from 0 to 30% of total cardiac output. It is controlled by the sympathetic response to core or environmental temperature changes, which alters the degree of vasoconstriction of the arterioles and arteriovenous anastomoses feeding into the venous plexi of the skin. This changing of blood flow affects core body temperature as heat is then lost from the body by radiation, a composite of conduction whereby heat is lost into anything the body touches (for example, a chair or bed), and is thus self-limiting, and convection whereby heat is removed as the surrounding air circulates. Heat is consequently lost through evaporation: 0.58 kilocalories of heat are lost for each gram of water that evaporates. It is therefore important to cover any parts of the body not directly involved in the massage or that are awaiting massage, to avoid excessive heat loss during a treatment session.
The first thing to be noticed by the massage student when touching the skin is that it is usually soft and, in most parts of the body, smooth. This top outer layer which can be touched is the epidermis, the epithelium of the skin. It consists of keratinised, stratified, squamous epithelium which is arranged in five laminae according to their cell type (Fig. 2.1). These five layers are arranged in two zones: the deeper is known as the zona germinativa, a single layer of columnar cells, and the more superficial zone is the stratum basale. Terminology, however, varies between anatomists and biologists (Thibodeau & Patton 2007). Cells are continuously being lost from the surface and replaced from the deeper layer.
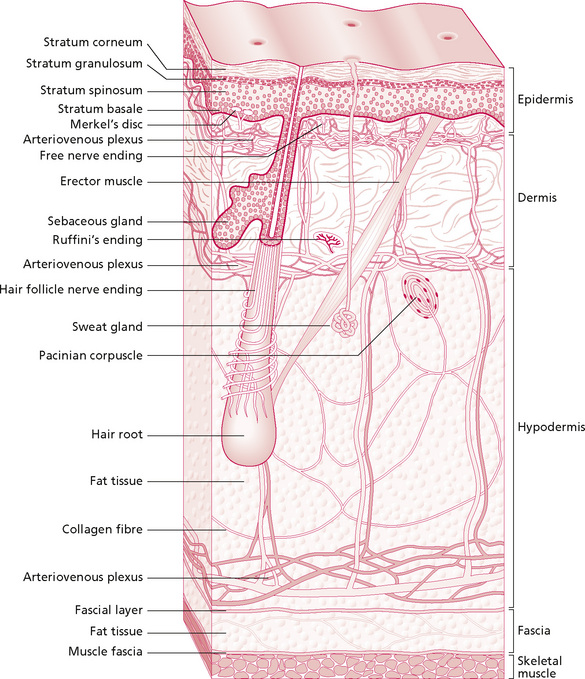
Figure 2.1 • Layers of the skin and circulatory plexi.
Reproduced, with permission, from Holey (1995). Originally published in Schuh I 1994 Bindegewebsmassage. Fischer-Verlag, Stuttgart.
This natural process occurs constantly, but certain events will speed it up. Friction on the skin, for example, caused by the clothes when dressing or during massage, results in desquamation. Massage will often remove the top layer (stratum corneum), which is readily replaced. This occurs when skin may be seen to ‘rub off’ or may be left on the treatment plinth following treatment. This is a normal and painless process. It tends to be excessive if the skin is very dry or visibly flaky (following removal of a plaster cast, for example) and can be reduced by the application of an oil-based lubricant. In severe cases, a soap and oil solution is particularly beneficial. The epidermal cells become gradually flatter and more keratinised as they move to the surface—more than 90% of epidermal cells are keratinocytes (Thibodeau & Patton 2007). Keratin, a protein, is important for hydration of the skin. Dry skin has reduced water content and the keratin allows swelling when the skin is wet. This is useful information to have when selecting the appropriate media for massage: the choice of oils, creams or talcs should be based not only on the type and purpose of massage but also on the individual skin quality. Keratin provides protection, and skin that has been soaked and has a whitened, wrinkled appearance should not be massaged as the effects of treatment on the superficial layer will be difficult to predict and monitor.
The dermis lies beneath the epidermis and forms most of the skin thickness. Whereas the epidermis is composed mostly of cells, the dermis contains collagen and elastin fibres, which give the skin its mechanical properties. The dermis is flexible and varies in thickness from the dense layer on the soles of the feet to the thin layer of the eyelids. It is composed of connective tissue (see below) arranged in two layers: the papillary layer is superficial to the deeper reticular layer.
The more superficial papillary layer of the dermis connects the epidermis with the dermis. Tiny conical projections, the papillae, project into the undersurface of the epidermis. These are sensitive and vascular, and range from being sparse to lying in dense lines which are seen as ridges on the surface of the skin, for example on the pads of the fingers and toes. They create friction for gripping and are used for fingerprint identification.
The reticular layer contains mainly thick collagen fibres (for strength) interspersed with reticular and elastin fibres (the latter for stretch and pliability) which form a tough interwoven layer. The directional lay of the skin in different parts of the body results from some of these fibres lying parallel to the skin surface. Skin is always under tension and the lines along which this tension lies are known as Langer's or Kraissl's lines (Fig. 2.2). They are important because they dictate the natural variation in tension when the skin resists movement and when it heals. If a cut in the skin follows the tension lines, scarring will be minimal as there will be minimal stress on the wound—an important principle utilised by surgeons. The fibroblast cells (which secrete the precursor for collagen) and phagocytes, important in immune defence mechanisms, are found in this layer.
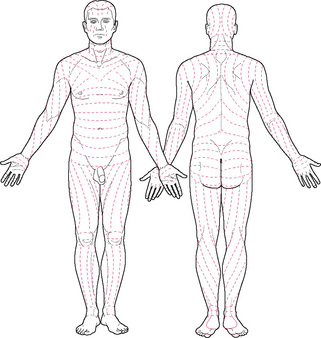
Figure 2.2 • Tension lines of the skin.
Reprinted from Anatomy: regional and applied, Last (1984), with permission from Elsevier.
The wrinkling in the skin around the nipple and scrotum is caused by the presence of smooth muscle fibres in the dermis. Stretch marks (striae gravidarum) follow partial rupture of the fibres of the reticular layer. It is commonly thought that this follows stretching of the skin in pregnancy, or fat deposition. This does not explain the common occurrence of these marks in thin people or the nulliparous, often on or above the sacrum, which could indicate weakening of the dermis due to the action of hormones or disease. As these marks show some fragility of the tissues, care should be taken when handling them. In particular, this tissue should not be overstretched by manipulation, to avoid possible further rupture of fibres and more extensive marking.
Circulation in the skin
Blood supply to glabrous (non-hairy) skin is maintained by arteriovenous anastomoses, found in the deeper layers of the dermis. They are surrounded by smooth muscle, the glomera, which maintain blood supply to the skin, despite variations in blood flow due to vascular responses aimed at maintaining body temperature.
Blood vessels within skin are found lying in and running between three flat horizontal plexi, as shown in Figure 2.1. Here, it can be seen that small arteries pierce the superficial fascia and form a horizontal plexus known as the rete cutaneum at the interface between the dermis and superficial fascia. It gives off vessels to supply the adipose tissue, glands and follicles. Some vessels reach the junction between the reticular and papillary layers of the dermis where they form another flat plexus, the rete subpapillare or superficial plexus. From here, capillaries supply the dermal papillae before travelling back to the venous plexus immediately below the superficial plexus, draining into the flat intermediate plexus in the middle of the reticular layer of the dermis which connects to the deep laminar venous plexus at the dermis–superficial fascia junction.
As the vessels tend to lie in plexi at the different interfaces of skin, movement in the form of manipulation between layers will influence the circulation. Capillaries running through the layers will be similarly influenced by gross movements of the tissue as a whole. This is discussed in more detail in Chapter 3.
Subcutaneous tissue
The layer of connective tissue under the dermis is adipose (fat) tissue and may be termed the hypodermis or subcutis. It is a layer of superficial fascia in which fibres from the reticular layer of the dermis extend to lie in bundles between fat lobules, giving it its sometimes characteristic appearance. It varies in density and thickness in individuals; for example, it is generally thicker in women than in men, and varies in different parts of the body. This layer is important in that, apart from its insulating properties, it gives mobility to the skin. One of the effects of massage is that it maintains or restores this mobility, particularly at the tissue interfaces. The sliding of the layers on each other can easily be felt.
Innervation of the skin
Skin has an important sensory function, which gives it a role in communication, reproduction, protection and coordinated movement. Responsibility for this is held by the many nerve endings responsive to sensory information. This is transmitted along nerves of the peripheral nervous system to the central nervous system, where it is interpreted and a response is made. The area of innervation of any one nerve fibre varies considerably, with overlap between the receptive fields of two adjacent nerve fibres. In the fingers, for example, each single fibre supplies a small area of skin. This is accompanied by a low sensitivity threshold and a high degree of spatial localisation. This means that the nerve is easily triggered by sensory stimuli and the brain can pinpoint the location of the stimulus very precisely. This high degree of sensitivity is particularly marked on the lips and external genitalia. On an area such as the back, however, a single nerve fibre will supply a larger area of skin, the nerve has a higher sensitivity threshold and the brain is less precise in its spatial localisation.
These nerves penetrate the superficial fascia and ramify through the dermis. They lie in plexi in the papillary layer of the dermis and around the hair follicles. The nerve endings are predominantly myelinated and non-myelinated ‘free’ nerve endings found in the dermis and lower parts of the epidermis. They monitor temperature and some pain. In addition there are specialised end-organs which include:
• Merkel discs which respond to shear forces and vertical pressure: this stimuli is magnified by hair follicles and structural ridges grouped into Iggo dome receptors, on the underside of the epithelium;
• Meissner's corpuscles, found in the dermal papillae: these are rapidly adapting mechanoreceptors which are responsive to mechanical deformation;
• Pacinian corpuscles in the deeper layers of the skin and superficial and deep fascia: they are innervated by a single axon only, and respond to a narrow range of vibration frequencies and rapid changes within the tissues; and
• Ruffini's corpuscles in the deeper dermis and hypodermis respond to stretch: they have an important tactile function.
The endings show specificity as a result of their differential sensitivities, and interpretation of the modality of sensation is due to the location of their termination in the brain. The endings listed above are those which have most relevance to massage. They are stimulated by mechanical deformation which stretches the membrane, thus opening the channels through which ions pass to depolarise the nerve fibre. The Meissner's corpuscle consists of a central nerve fibre surrounded by terminal nerve filaments within an elongated capsule. The construction of the Pacinian corpuscle consists of a central nerve fibre surrounded by capsular layers (Fig. 2.3). The fibre is distorted in various ways by compression of any part of the capsule. The fluid in the corpuscle immediately redistributes so that the deformation is no longer transmitted to the central fibre until the force is removed. Thus, repetitive forces rather than continuous ones will affect the nervous system more strongly; this demonstrates the importance of the continuous movement in massage.
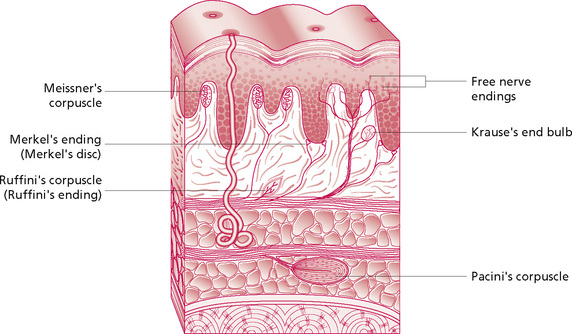
Figure 2.3 • Somatic sensory receptors - exteroceptors.
Reprinted from Anatomy and Physiology 6e, Thibodeau & Patton (2007) with permission from Elsevier.
Sensory stimuli are carried in nerve fibre types II (type A β and γ), III (type A δ) and IV (type C), which are the smaller, slower conducting types. The stimulus enters the spinal cord via the dorsal horn and diverges or converges via neuronal pools. It passes to the brain either (1) through the dorsal column, crossing to the opposite side in the medulla, to the medial lemniscus and the thalamus, or (2) by crossing to the opposite side of the spinal cord, passing through the anterior and lateral white columns to the brainstem and thalamus. The former is for rapid, sensitive transmission of sensation and the latter for a slightly cruder response. The thalamus relays the sensory information to the cerebral cortex for localisation, interpretation and controlled response.
The epidermis and dermis filter mechanical and thermal stimuli, which attenuate in different ways as they are transmitted through the layers. This is probably because different stimuli need to be strongest at specific depths in order to have maximum impact on sensory end-organs.
Connective tissue
Skin is a type of connective tissue. The connective tissue that lies under the skin is important as a base for providing attachment to skin, providing both anchorage and mobility. These tissues are all formed from the embryonic mesoderm (middle layer) and all are basically composed of the same constituents. The presence of specialised cells, or an alteration in the predominance of any constituent in a particular tissue, ensures that the different connective tissues are adapted and well suited to their purpose. Thus, the appearance and properties of the different connective tissues vary significantly. They range from fascia, ligament and tendon to specialised types including cartilage and bone.
The cells of connective tissue lie within a matrix formed by fibres lying in ground substance. The fibres are collagen, reticulin and elastin fibres, the first offering resistance to stress while the last allows for stretch. There are more than ten types of collagen.
Fibrillar collagen types I and III are the most relevant here, being found in tendon, skin, ligament and skin vessels. These types have the greatest stiffness (see below). The ground substance is composed of water and organic molecules, predominantly glycosaminoglycans (GAGs). These molecules have water-binding properties and are responsible for the amount of water present in the tissue. This gives them a role in the diffusion of molecules through the tissue, notably incoming nutrients and outgoing metabolites. They are also important for communication between cells and for their adhesion to collagen. Their structure makes them springy and so able to absorb shock. This mechanism is enhanced by their being held under tension by the surrounding collagen fibres. In a specialised connective tissue such as cartilage, this springiness allows compression when weight-bearing and restoration of shape when the weight is removed. In the superficial tissues, these molecules allow take-up or removal of liquid, which appears to be partly under hormonal control. Skin, therefore, can be ‘dry’ or well hydrated, terms that refer to the skin itself rather than its surface. This can be palpated as a change in consistency and loss of compliancy of skin and is different from the presence of fluid in the tissue spaces, which gives the skin a ‘spongy’ feel. Hyaluranon has a role in hydration and enables gliding movement within the tissue (Culav et al 1999).
The cells of connective tissue are:
• Fibroblasts: synthesise collagen and reticulin, important in repair;
• Plasma cells: produce antibodies, present in large numbers in pathological states;
• Mast cells: in loose connective tissue, release inflammatory agents of histamine, serotonin, heparin;
• Fat cells: mobilisation of the contained fat is under nervous, hormonal and chemical control; and
• Pigment cells (chromatophores): in the corium, contain melanin.
Biomechanics of connective tissue
The main constituent of connective tissue is the protein collagen, the function of which is to resist axial tension and which has been shown to exhibit standard stress–strain behaviour. Collagen is derived from its precursor procollagen, synthesised by the fibroblast. Procollagen becomes tropocollagen, which in turn forms microfibrils. Under the electron microscope, collagen fibres are seen to be arranged in bundles and have a crimped appearance, thought to be due to molecular attachment. There are several types of collagen. Type I collagen (in dermis and fascia) consists of three polypeptide chains, coiled in a left-handed helix; these helical chains are coiled together in a right-handed helix. This type gives resistance to tension and provides filtration and support. The basal lamina of epithelia contains type IV collagen, which synthesises epithelial cells. Hydrogen cross-links are formed between chains, and also between molecules, giving stability at fibril level and between fibrils, assisting in the formation of collagen fibres. They allow the tissue to function under mechanical stress.
The orientation of fibres depends on the stresses to which the fibre is subjected. Connective tissue needs to be pliable yet very strong. The stress–strain behaviour of biological materials may be demonstrated by a stress–strain curve (Fig. 2.4). These have been plotted for various types and constituents of connective tissue, the exact behaviour differing slightly in each:
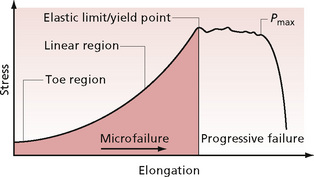
Figure 2.4 • Typical stress–strain curve for connective tissue.
Stress = force applied per unit area; strain = proportional elongation; Pmax = maximum load point.
When longitudinal stress is applied, the tissue responds firstly by elongation. This occurs in the toe region of the curve and is thought to be due to a straightening out of the crimping in the fibres. It may also be due to some interfibrillar sliding and shear of ground substance which flows between the fibres. Stress and strain are linearly proportional up to the proportionality limit. Elongation that occurs in this region will not be permanent. The elastic limit is the point to which maximum stress may be applied without permanent deformation. As the loading continues, a greater force is required to increase elongation. This is due to the fact that the tissue becomes stiffer.
Schliep, Klingler and Lehman-Horn (2005) have found that, during an initial isometric stretch, fluid is extruded which returns when the stretch is removed. This will affect the apparent stiffness of the tissue and also indicates that tissue fluid is refreshed and renewed during massage.
The next part of the curve is the linear region. Microfailure begins to occur in some of the fibre bundles, but the tissue retains its external appearance of continuity. The end of the linear region is known as the yield point of the tissue. Here, elongation (or yielding) can occur without a corresponding increase in load. Once beyond this point, major failure of fibre bundles occurs. The external appearance changes, and the smooth outline is lost; this is termed ‘necking’. The same effect is seen, for example, when an elastic band is overstretched or becomes overused and its width changes and appears uneven. It loses strength but still has gross continuity. When the maximum load point is reached at Pmax, complete rupture occurs. Its position on the curve designates the rupture strength.
Most changes that occur as a result of massage are in the toe region of the curve. The therapist should take care not to overstretch the tissues, thereby causing damage to the internal structure of connective tissue fibres. Each massage student should learn to be sensitive to the ‘end-feel’ of the tissues (see Chapter 5). Any permanent deformation is termed plastic and occurs with microfailure—breakage of cross-bridges. These are molecular bonds which bind adjacent fibres together. When lengthening occurs, the bonds are broken; they then re-form further along the fibres, enabling the fibres to reposition themselves in relation to each other, resulting in increased length of the tissue. The undamaged fibres absorb a greater proportion of the load as a new length is established, which reflects the balance between elastic recoil and the resistance of the water and GAGs to compression.
Biological materials also have viscosity, a property of fluids of resistance to flow, and elasticity, a property of solids. They are therefore said to be visco-elastic. The response is dependent on how quickly the load is applied or removed (regardless of the size of the strain). The quicker the repetitive loading and unloading occurs, the stiffer the material becomes, as there is less chance for the ground substance to flow between fibres. This continuous loading and unloading causes friction and a rise in temperature, as energy is dissipated as heat when the tissue is returned to its original length. This quality is known as hysteresis. Consideration should be given to the speed at which massage is undertaken and the therapist should be sensitive to how stiff the tissues feel. If an increase in tissue mobility is aimed at, the progression of treatment should be partly dictated by this stiffness and the rate at which any lengthening is felt to occur.
If a low constant or repetitive load is maintained over a long period, the elongation is by creep, whereas if the length is held constant, elongation occurs by relaxation. Excess fluid in the tissues which remains for a long period of time, for example in chronic oedema or lymphoedema, will stretch and produce a creep effect on the surrounding tissues and they will not be restored to their original shape following removal of the fluid. This is important as connective tissue helps to maintain the fluid pressure in the tissue spaces. If the pressure reduces, due to stretching of connective tissue as a result of creep, the pressure within the vessels may be comparatively greater and external pressure must be applied to prevent swelling. For example, pressure stockings or sleeves may be worn to maintain the beneficial effects of oedema massage.
Massage produces low repetitive load over the medium term and may cause a non-permanent creep response. Biomechanical study of connective tissue indicates that the quicker tissue is loaded, the stiffer it becomes owing to its visco-elastic properties. Dry tissue produces more friction, and is known to lose compliance, elongating less readily than when the tissue has good water content. Thus, the rate of massage should be suitable for the type and condition of the tissue.
It is known that connective tissue initially responds to stretch with an increased stiffness (Yahia et al 1993) thought to be due to the presence of contractile cells, but this response has not been fully understood until recently. Schleip et al (2005) found this to be due to changes in matrix hydration. Myofibroblasts have recently been found in fascia (Schliep et al 2006). These are contractile and it has been demonstrated experimentally that they respond to pharmacological agents, notably mepyramine (a antihistamine), whose effects can last for up to two hours. Histamine and oxytocin have a lesser effect in that the contractile response is shorter lasting. A nitric oxide donator triggered brief relaxation in some tissue samples. The same research team found that approximately 50% of the tissue bundle samples exhibited a stronger resistance force in response to 5% isometric strain and this lasted for up to 45 minutes (Schliep et al 2005, 2006). These findings are of considerable interest in explaining some of the phenomena felt by the therapist when handling tissues.
Goldfarb et al (2001) compared the effects of low- and high-force rehabilitation on collagen synthesis and extracellular matrix maturation in sutured intrasynovial flexor tendons in dogs. They found that higher force rehabilitation did not accelerate healing, as no changes were seen in the biomechanical composition of the healing tendon over a period of 6 weeks. This suggests that there is no advantage in applying heavy stresses to the tissues in relation to healing speed.
The implications of tissue biomechanics in the different phases of healing are discussed later.
Fluid system dynamics
The therapist must be able to recognise subtle changes in the fluid content of the tissues. One of the first things she notices when performing any form of deep massage in the tissues is that the fluid balance between the circulation and the tissue spaces changes, sometimes rapidly. To understand how the squeezing, pulling and stretching manipulations affect the tissue fluid, it is necessary first to examine tissue fluid dynamics.
The fluid of the body serves to transmit nutrients and to bathe structures, ensuring that the correct chemical and electrolyte balance is maintained around the cells. In some instances this is crucial—around the heart, for example, which cannot function correctly without its muscle cells being bathed in a perfect balance of electrolytes. It may appear, simply, that newly oxygenated blood is pumped from the heart and carried in the arteries, arterioles and capillaries (in order of descending size) and, when deoxygenated, returns for replenishment via the capillaries, venules and veins. However, the mechanism by which specific components of the blood flow into and out of the tissues, and maintenance of the delicate balance of fluid inside and outside the tissue spaces, warrant closer examination.
Fluid movement occurs either by diffusion or osmosis along concentration gradients or by flow along pressure gradients. In diffusion or flow, water flows down the gradient, from an area of high concentration or pressure to one of lower concentration or pressure. However, in osmosis, water flows up the gradient, towards the side of the most concentrated solute. This physiological concept of functional gradient is an attempt by the body to achieve balance and uniformity. So, when two non-uniform areas exist, in either concentration or pressure, fluid will move to try to level them out.
After fluid has been filtered by the capillaries, it returns to the circulation by the venous end of the capillary loop. A small proportion is returned via the lymphatics. There are two key areas involved in maintenance of tissue fluid balance: the capillary loop and its surrounding environment, and the lymphatic vessel. The capillary loop has an arteriolar end and a venule end. This demonstrates that both sections of the blood vessel network are, in fact, continuous and the moment at which blood coming from the heart becomes blood going to the heart depends on a subtle pressure change.
The important mechanism by which equilibrium of fluid within the blood vessels and tissue spaces is maintained was first described by Starling. It is the mechanism by which the inherent leakiness of the capillaries through their semipermeable membranes is counteracted to maintain the volume of circulating plasma.
To understand these pressure changes, it is necessary to examine the blood, which is composed of plasma (clear liquid containing substances in solution or suspension), red and white cells, and plasma proteins (such as prothrombin and fibrinogen). Nutrients, for example oxygen, enter the tissue spaces from the bloodstream by diffusion, until concentration gradients are equal on both sides of the capillary wall. Plasma proteins are too large to leak out of the tiny pores in the capillary vessel walls and therefore create a pressure (known as osmotic pressure) inside the vessel. If you imagine liquid flowing into an empty vessel, its ease of passage will depend on the relationship between the quantity of liquid and the size of the vessel. If the vessel contains large particles, this effectively reduces the space available for the liquid to occupy and increases the pressure inside the vessel. This is exactly the effect of the plasma proteins. In some situations, they are able to leak out into the tissue spaces when the vessels dilate, which causes the pores in the vessel walls to increase in size. Adipose tissue contains type I capillaries which have uninterrupted membranes with ‘pores’ in them. The pressure from the fluid itself is known as the hydrostatic pressure. It is slight outside the vessel, but greater inside owing to the pumping of the heart. This pressure is opposed by the osmotic (or colloid osmotic, or oncotic) pressure, which is a reabsorption pressure resulting from the fluid itself which forces fluid back in the vessel. If the osmotic pressure inside the vessel and the hydrostatic pressure outside the vessel are equal, then fluid flow will not occur. In reality, at the arterial end of the capillary loop, the net result of these different pressures tends to produce an outward force, moving fluid into the tissue spaces. However, at the venous end of the capillary the net pressure tends to move fluid into the capillary, thus maintaining normal balance (Fig. 2.5). The development of oedema and its implications will be discussed subsequently. These pressures can be influenced by compression or movement of the tissue, as in massage, and natural pumping mechanisms can be enhanced mechanically in this way.
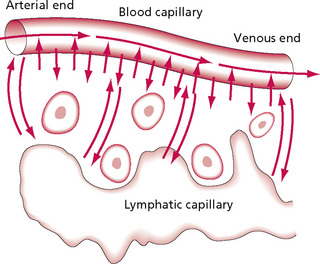
Figure 2.5 • Diffusion of fluid and dissolved substances between the capillary and interstitial fluid spaces.
Reprinted from Textbook of Medical Physiology 8e, Guyton (1991) with permission from Elsevier.
The lymphatics
The pressure in loose subcutaneous tissue is negative, maintained at −3 mmHg as a result of the pumping action of the lymphatics. One-tenth of fluid is removed via the lymphatics rather than the venous end of the capillary. Proteins and other substances of higher molecular size that leak into the interstitial spaces cannot return into the capillaries because of the adverse concentration gradient, and must return via a different route—the lymphatic system. If massage is to enhance this effect and improve this system, it needs to be employed in the correct way; thus an understanding of the lymphatics is necessary for successful massage.
The lymph system mirrors the vascular system in structure. Tiny lymph capillaries have blind endings in the tissue spaces which drain into veins. They form a mesh-like system in the tissue spaces, and are slightly larger than vascular capillaries. They are composed of a single layer of endothelial cells, the basement membrane often being absent. The capillaries unite to form larger vessels and have a coating of connective tissue outside the endothelium. The larger collecting trunks resemble small veins structurally, with smooth muscle present in their walls, which is important for the transmission of fluid. Unidirectional flow is maintained by the presence of semilunar, paired, one-way valves, which are found in higher numbers here than in veins (Fig. 2.6).
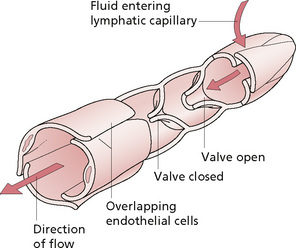
Figure 2.6 • Structure of a typical lymphatic capillary.
Reprinted from Anatomy and Physiology 6e, Thibodeau & Patton (2007) with permission from Elsevier.
The movement of lymph occurs via the following chain of events. Filtration pressure in the tissue spaces is caused by filtration of fluid from the vascular capillaries. The surrounding muscle pump effect compresses lymph vessels and lymph moves as directed by the valves. Flow is enhanced by respiratory movements, particularly inspiration, creating a negative pressure in the brachiocephalic veins, the pumping effect of muscle contraction and possibly arterial pulsing. Sympathetic stimulation causes contraction of the smooth muscle in the vessel walls, particularly near the valves. The endothelial cells of the lymphatic capillaries are attached, by anchoring filaments, to the surrounding connective tissue. When fluid enters the tissues they swell and the anchoring filaments pull the capillary open, allowing fluid to flow between the cells. This effect can also result from the mechanical deformation that occurs during massage. The overlapping cells produce a valve-like effect by which fluid can flow in, but not out, as the back-pressure closes the flap. The rate of flow is determined by the interstitial fluid pressure and lymphatic pump activity.
The factors increasing lymphatic flow are:
• Increased capillary pressure;
• Reduced plasma osmotic pressure;
When interstitial fluid pressure reaches a little above atmospheric pressure (0 mmHg), lymph flow fails to continue rising, probably due to fluid compressing the outer surfaces of the larger lymph vessels, impeding flow (this is why massage should be directed proximally). The lymphatic capillaries empty into collecting lymphatics. The vessel stretches with increased volume of fluid, and the smooth muscle in the walls of the vessels contracts (Starling's law). This effect is particularly marked immediately proximal to the valves, and so pumps fluid through the proximal valve. Thus, pumping is increased by the compression of the vessel (Guyton 1991). A pumping action is also exerted by actomyosin in the capillary end cells.
The lymph nodes are small oval structures composed of a collagenous capsule and an interior trabecula. Within this framework is a fine reticulum within which many lymphocytes are embedded, with fewer macrophages and reticular cells (littoral cells). Blood vessels and the efferent lymph vessel enter and leave the node at the hilum, while afferent lymph vessels enter it around the periphery. Points at which the reticulum is loose, relatively cell free, and where lymph flows freely are termed lymph sinuses. Within the cortex, collections of cells are termed lymphatic follicles, or nodules, and contain germinal centres.
Lymphatic fluid is transported from the tissue spaces to the nodes where it passes in close proximity to a range of phagocytic cells. The large surface area of the node's interior ensures that numerous cells are available. Micro-organisms and foreign bodies are removed from the bloodstream here, to be processed by the lymph glands, and certain cells and proteins are returned via the lymphatics. The lymphatics assist in reducing the threat of toxic spread throughout the bloodstream; however, they also provide a channel for the spread of infection or malignancy.
Fluid eventually returns to the bloodstream via the thoracic duct, which extends from the twelfth thoracic vertebra to the root of the neck. It passes through the diaphragm and travels on the posterior mediastinum to the neck, where it drains into the left subclavian and internal jugular veins. At the base of this duct is the cysterna chyli, which extends down to the first lumbar vertebra. It receives the right and left lumbar and intestinal lymphatic trunks, which in turn drain the lower limbs, pelvis and abdomen, respectively. In addition, drainage from the intercostal trunks, the left jugular trunk draining head and face, and the left subclavian trunk draining the upper limb, join the thoracic duct. The right lymphatic duct receives drainage from the right jugular trunk, the right subclavian trunk and the right bronchomediastinal trunk. Most drainage from the head and neck enters the deep cervical (superior and inferior) group of nodes, which in turn drain into the jugular trunk. There are also minor groups draining particular areas. Figure 2.7 gives a summary of drainage and shows the position of the nodes.
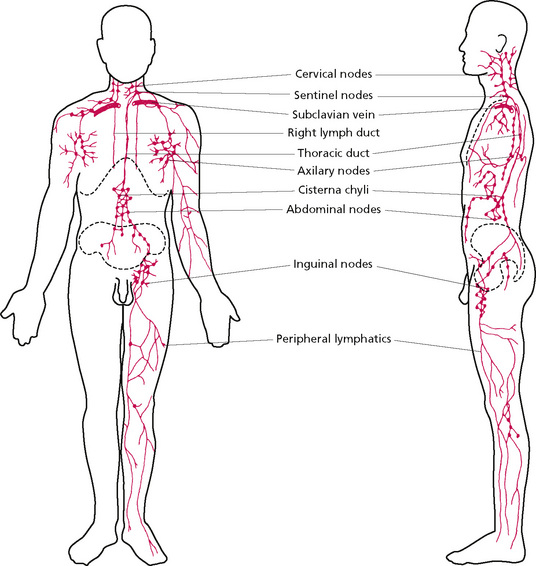
Figure 2.7 • Main lymphatic channels and glands.
Reprinted from Textbook of Medical Physiology 8e, Guyton (1991) with permission from Elsevier.
Lymph drainage from the skin occurs by a system of overlapping skin areas. The tiniest vessels are known as precollectors; they drain from several skin areas into a collector in the hypodermis. The skin area of one collector is a strip of skin—a ‘skin zone’. The zones of a single lymph vessel bundle have been termed a territory (Kubik & Manneston 1984). The borders of these territories are known as lymphatic watersheds as there is normally no functional connection across watersheds, although they anastomose with each other.
As lymph drains into a single duct in a proximal direction, it is important that any attempt to increase this drainage mechanically occurs in the same proximal direction. A lymphatic system that is blocked with fluid anywhere on its course will necessitate that extra mechanical drainage occurs in the trunk, to clear an area into which the more distal fluid can drain. This is discussed in more depth in Chapter 11.
Muscle tone
Massage can also affect skeletal muscle. A healthy muscle is always in a certain amount of resting tone, which is a response of the muscle fibres to nervous activity; this maintains the muscle in a state of slight, normally imperceptible, contraction. Tone can be increased, especially in postural muscles, by factors such as stress or cold. This is probably, in part, due to the fact that muscle spindles have a sympathetic innervation (Barker & Saito 1981). Tone is dependent on interaction between the muscle spindle (sensitive to length and rate of change in length of muscle) and the central nervous system. Stretching a muscle will stimulate the spindle and cause reflex muscle contraction, while reflex inhibition of the antagonist occurs. Massage can add an external stimulus to sensory organs and either increase tone by stimulation or reduce it, probably by facilitating an accommodation of the spindle, causing it to ‘reset’ at a lowered threshold of excitability. The sympathetic supply to muscle spindles means that any influence on the autonomic nervous system will affect muscle responses. Massage techniques which have a general relaxation effect, as well as local massage, will therefore change muscle tone.
Conclusion
It is important for the massage therapist to understand the effects of massage on the different body systems. The structure and function of the skin, how it is nourished with blood and the way in which fluid and fibres behave must be understood before the effects of massage can be explored. The mechanisms by which muscle tone is maintained and influenced are pertinent, as are the transmission and interpretation of touch and other types of sensation. Only by a full understanding of these factors can the therapist work autonomously and make effective and informed clinical decisions.
Key points
• Horizontal arteriovenous plexi lie at the skin interfaces and can be influenced by massage.
• The dermis gives a mobility to the skin which can be maintained or restored by massage.
• Fibres in the tissues can be overstretched beyond their elastic limit, causing damage.
• Failure of fibre bundles occurs beyond their yield point. This can be prevented by not stretching tissue beyond its end-feel.
• Massage principally works within the elastic limit of the stress–strain curve.
• Massage provides a low repetitive load over the medium term and may create a non-permanent creep response by which the tissue is temporarily elongated.
• Massage should not be done too quickly, to avoid a stiffening of connective tissue, as a result of its visco-elastic properties.
• ‘Dry’ tissue should be massaged more slowly as more friction will be produced between collagen fibres.
• The effectiveness of the venous system can be enhanced, to aid removal of excess tissue fluid, by increasing its pumping effects with massage.
• Lymphatic drainage can be increased by a pumping effect and a pulling on the filaments, which open the gaps between cells in the walls of lymphatic vessels.
• The muscle spindle can be stimulated or caused to accommodate by massage, producing a change in muscle tone.
Barker D., Saito M. Autonomic innervation of receptors and muscle fibres in cat skeletal muscle. Proc. R. Soc. Lond. B Biol Sci.. 1981;212:317-332.
Culav E.M., Clark C.H., Merrilees M.J. Connective tissues: matrix composition and its relevance to physical therapy. Phys. Ther.. 1999;79(3):308-319.
Goldfarb C.A., Harwood F., Silva M.J., et al. The effects of variations in applied rehabilitation force on collagen concentration and maturation at the intrasynovial flexor tendon repair site. J. Hand Surg. [Am.]. 2001;26(5):841-846.
Guyton A.C. Textbook of medical physiology. Philadelphia, PA: W B Saunders, 1991.
Holey L.A. Connective tissue manipulation: towards a scientific rationale. Physiotherapy. 1995;81(12):730-739.
Kubik S.T., Manneston M. Anatomie der Lymphkapillaren und Präkollektoren der Haut. In: Bonniger A., Partsch H., editors. Initiale Lymphstrombahn, Internat Symp. Zurich: G Thieme; 1984:62-69.
Last R.J. Anatomy, regional and applied. Edinburgh: Churchill Livingstone, 1984.
Schleip R., Klingler W., Lehmann-Horn F. Active fascial contractility: fascia may be able to contract in a smooth muscle-like manner and thereby influence musculoskeletal dynamics. Med. Hypotheses. 2005;65:273-277.
Schleip R., Naylor I.L., Ursu D., et al. Passive muscle stiffness may be influenced by active contractility of intramuscular connective tissue. Med. Hypotheses. 2006;66:66-71.
Thibodeau G.A., Patton K.T. Anatomy and physiology, sixth ed. St Louis: Mosby Elsevier, 2007.
Yahia L.H., Pigeon P., DesRosiers E.A. Viscoelastic properties of the human lumbodorsal fascia. J. Biomed. Eng.. 1993;15(5):425-429.