Chapter 5 Nervous system and special senses
The presence of a nervous system allows an animal to respond, in a coordinated manner, to both the demands of the external environment and the internal changes within its body. The functions of the nervous system are:
Although the nervous system works as a well-integrated composite unit within the body, for descriptive purposes, it can be divided into:

Nervous tissue
Neuron structure
The main cell of the nervous system is the neuron (see Ch. 2, Fig. 2.16). Neurons are responsible for the transmission of nerve impulses throughout the nervous tissue. Nerve impulses pass from one neuron to another by means of structures known as synapses. All nerve pathways are made up of neurons and synapses. Complex nervous tissue, such as that of the brain and spinal cord, is made of neurons supported by connective tissue and neuroglial cells, whose function is to supply nutrients to and carry waste materials away from the neurons.
Each neuron consists of the following parts:
Nerve impulses from the dendrons are directed across the cell body towards the axon hillock and continue down the axon, reaching their final destination very rapidly. The speed of transmission along the axon is increased by the presence of a myelin sheath. Myelin is a lipoprotein material made by Schwann cells surrounding the axon. Its whitish appearance contributes to the colour of the more visible nerves in the body and to the white matter of the CNS. The myelin sheath is interrupted at intervals of about 1 mm by spaces known as the nodes of Ranvier and it is through these that the axon tissue receives its nutrient and oxygen supply. Non-myelinated fibres are embedded within the Schwann cells (Fig. 5.1) and despite their name are actually covered in a single layer of myelin. Totally uncovered fibres are rare and may be found in areas such as the cornea of the eye.
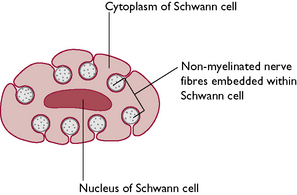
Fig. 5.1 Cross-section through a Schwann cell showing non-myelinated nerve fibres embedded within the cytoplasm.
Neurons vary in size – the diameter of the axons and dendrons may be a few micrometres and the length depends on the destination of the nerve fibre. This may be anything from a few millimetres to more than a metre long. Neurons also vary in shape (Fig. 5.2).
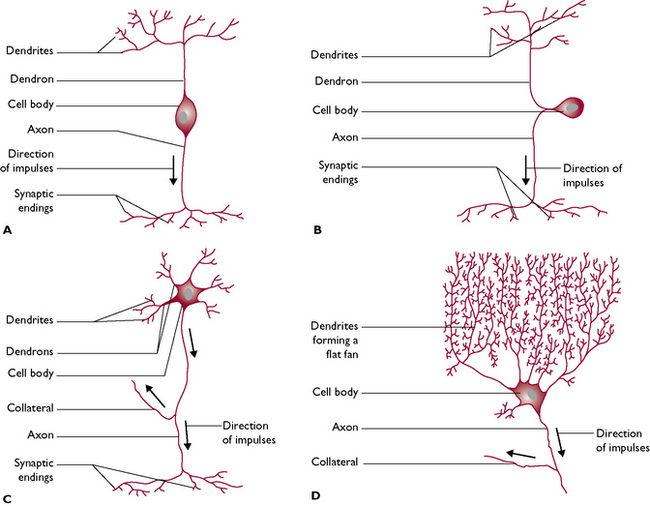
Fig. 5.2 The variation in shape of neurons. A Bipolar neuron. B Pseudo-unipolar neuron, e.g. a sensory neuron. C Multipolar neuron, e.g. a motor neuron. D Multipolar neuron, e.g. Purkinje cell from the cerebellum of the brain.
Synapses
Each axon terminates in a structure called a synapse (Fig. 5.3). Where an axon terminates on an individual muscle fibre, the synapse is referred to as a neuromuscular junction and the nerve impulse stimulates contraction of the muscle fibre. Within the button-like presynaptic ending are vesicles containing chemical transmitter substances. The most common chemical is acetylcholine but others found in the body include adrenaline (epinephrine), serotonin and dopamine.
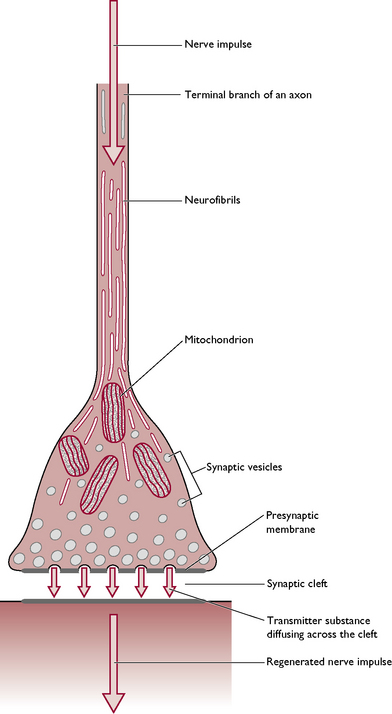
As the nerve impulse travels down the axon, the vesicles drift towards the presynaptic membrane (Fig. 5.3) and release the transmitter into the synaptic cleft. It diffuses rapidly across this gap and combines with the postsynaptic membrane, making the membrane more ‘excitable’ and allowing the transmission of the nerve impulse to continue on down the nerve fibre. The effect is stopped by the release of cholinesterases – enzymes that destroy any acetylcholine remaining in the synaptic cleft – and the synapse returns to its resting state ready for the next nerve impulse. Effective transmission of a nerve impulse across the synapse will only occur in the presence of calcium ions.
Generation of a nerve impulse
Nerve impulses transmitted along axons and dendrites can be considered to be electrical phenomena. An impulse changes the electrical charge of the neuron by altering the relationship between the negative charge of the cell contents and the positive charge of the cell membrane. This results from a change in the permeability of the cell membrane to sodium and potassium ions, causing an exchange of ions between the inside and the outside of the neuron (Fig. 5.4).
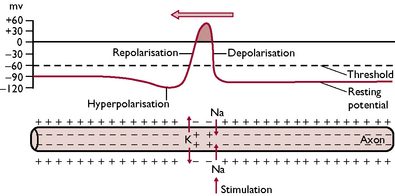
Fig. 5.4 The alterations in electrical charge that occur in an axon as a nerve impulse travels along it from right to left. The charge differences are brought about by movement of sodium and potassium ions across the axon membrane.
For a short time after the impulse has passed, the nerve fibre becomes refractive, i.e. it cannot be reactivated by another impulse until the permeability of the cell membrane returns to normal. This ensures that the flow of impulses travels only in one direction.
A nerve impulse is an all-or-nothing phenomenon – the nerve is either stimulated or it is not; there is no gradation of impulse. A particular neuron will always transmit with the same power or the same speed. The different effects of the nervous system depend on:
Classification of nerves
Peripheral nerves can be classified according to their function and to the structures they supply.
An intercalated neuron is one that lies between a sensory neuron and a motor neuron. Such neurons may not always be present within a nerve pathway.
Afferent and efferent are also terms used within other systems, e.g. the blood vascular system, to describe blood and lymphatic capillaries going to and leaving organs.
Nerve fibres may be further classified according to the organ with which they are associated.
The central nervous system
During embryonic development of the CNS, a hollow neural tube forms from the ectodermal layer of the inner cell mass of the embryo. This neural tube runs along the dorsal surface of the embryo and, as it develops, nerve fibres grow out laterally extending to all parts of the body and eventually forming the peripheral nervous system. The anterior end of the tube becomes the brain and the remaining tube becomes the spinal cord. The brain and spinal cord are hollow and are filled with cerebrospinal fluid (CSF).
The brain
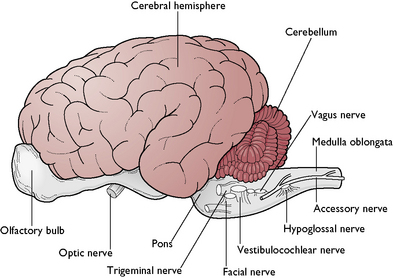
The function of the brain is to control and coordinate all the activities of the normal body. The brain is a hollow, swollen structure lying within the cranial cavity of the skull, which protects it from mechanical damage. In many species, three distinct areas can be identified – these are the forebrain, midbrain and hindbrain. In mammals, this arrangement may be difficult to recognise because the right and left cerebral hemispheres of the forebrain are enlarged and overlie the midbrain (Fig. 5.5).
Forebrain
This consists of the cerebrum, thalamus and hypothalamus.
The cerebrum – or right and left cerebral hemispheres – takes up the greater part of the forebrain and contains up to 90% of all the neurons in the entire nervous system (Fig. 5.5). The right and left sides are linked by the corpus callosum. This consists of a tract of white matter whose nerve fibres run across the brain between the right and left hemispheres. It is found in the roof of the third ventricle, which is part of the hollow ventricular system inside the brain and contains CSF.
The surface of the hemispheres is deeply folded, which enables a large surface area to be enclosed within the small cranial cavity and allows nutrients in the CSF to reach the cell bodies of neurons lying deep inside the brain tissue. The folds are known as gyri (upfolds), sulci (shallow depressions) and fissures (deep crevices). The two hemispheres are divided by the longitudinal fissure. Each cerebral hemisphere consists of four lobes each of which contains specific centres or nuclei with specific functions related to conscious thought and action.
The tissue of the cerebral hemispheres consists of:
The thalamus is found deep in the tissue of the posterior part of the forebrain. Its function is to process information from the sense organs and relay it to the cerebral cortex.
The hypothalamus lies ventral to the thalamus and has several functions:
On the ventral surface of the forebrain (Fig. 5.6) the following structures can be seen:
Midbrain
This is a short length of brain lying between the forebrain and the hindbrain. It is overhung by the cerebral hemispheres and is not easy to see in the gross specimen. It acts as a pathway for fibres running from the hindbrain to the forebrain carrying the senses of hearing and sight.
Hindbrain
This consists of the cerebellum, pons and medulla oblongata.
The cerebellum lies on the dorsal surface of the hindbrain (Fig. 5.5). Its name means ‘little brain’ as it was originally thought to be a smaller version of the cerebrum. It has a globular appearance and is covered in deep fissures. In cross-section, the tissue is very obviously divided into an outer cortex of grey matter and an inner layer of white matter. The grey matter consists of numerous Purkinje cells (Fig. 5.2), each of which forms thousands of synapses with other neurons.
The cerebellum controls balance and coordination. It receives information from the semicircular canals of the inner ear and muscle spindles within skeletal muscles. Voluntary movements are initiated by the cerebrum and fine adjustments are made and coordinated by the cerebellum.
The pons lies ventral to the cerebellum and forms a bridge of nerve fibres between the cerebellar hemispheres. It contains centres that control respiration.
The medulla oblongata extends from the pons and merges into the spinal cord (Fig. 5.5). It contains centres responsible for the control of respiration and blood pressure.
Injury or malformation of the cerebellum results in incoordination and spasticity. Kittens whose dam was infected with the feline enteritis virus during pregnancy may be born with cerebellar hypoplasia (underdevelopment) and will never be able to walk in a normal coordinated fashion.
The medulla oblongata and the pons form part of the brain stem that is responsible for the vital control of respiration and blood pressure. If the brain stem is damaged, e.g. by brain stem herniation through the foramen magnum, which may occur if the brain swells as a result of inflammation or traumatic damage, then respiration and blood pressure will be affected and the animal will die – often described as being ‘brain stem dead’.
Protection of the brain
Normal brain function is essential for the maintenance of an animal’s life. If it is damaged mechanically or chemically, function will be impaired and the animal may die. Several structures have evolved within the brain to protect it from harm.
Cranium
This bony structure forms a tough outer shell to protect the soft brain tissue from physical damage.
Ventricular system
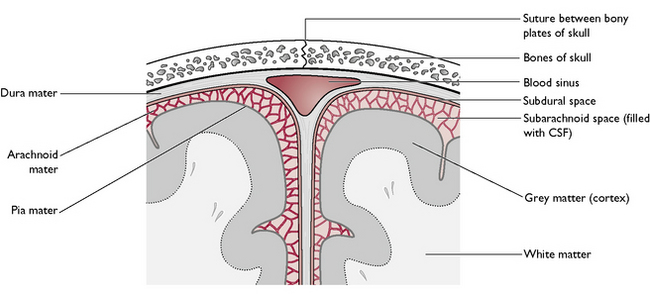
The ventricular system (Fig. 5.7) is derived from the hollow neural tube of the embryo. The lumen of this tube develops into a series of interconnecting canals and cavities or ventricles found inside the brain and spinal cord. The ventricles and the central canal are filled with cerebrospinal fluid (CSF). This also surrounds the outside of the brain, lying in the subarachnoid space (Fig. 5.8). CSF is secreted by networks of blood capillaries known as choroid plexuses lying in the roofs of the ventricles. It is a clear fluid, resembling plasma, but has no protein in it – it is an example of a transcellular fluid. The function of CSF is to protect the CNS from damage by sudden movement or knocks and to provide nutrients to the nervous tissue of the brain and spinal cord.
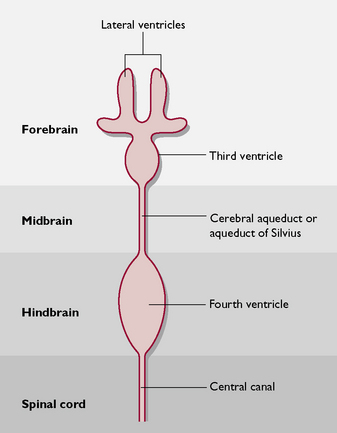
Fig. 5.7 Dorsal view of the ventricular system of the brain and its relationship to the areas of the brain (not to scale).
Samples of CSF can be collected from the cisterna magna, lying between the cerebellum and the medulla oblongata. The patient is anaesthetised and restrained in sternal or lateral recumbency, with its chin touching its chest. A spinal needle is slowly advanced into the space until CSF begins to drip from the hub.
Meninges
The meninges (Fig. 5.8) consist of three protective layers around the brain and spinal cord. From the outermost layer inwards, they are:
Blood–brain barrier
This is a modification of the neuroglial tissue that connects and supports all the neurons within the nervous tissue. Different types of neuroglial cells surround the blood capillaries, creating an almost impermeable layer, to protect the brain from substances that are harmful to or not needed by the brain. These include urea, certain proteins and antibiotics. Other materials such as oxygen, sodium and potassium ions and glucose can pass rapidly through the barrier to be used for brain metabolism.
The action of general anaesthetic agents relies on their ability to pass through the blood–brain barrier and affect the neurons in the brain. Drugs that affect the brain must be in a lipophilic form, i.e. soluble in lipids, so that they are able to pass through the phospholipid cell membranes of the neuroglial cells.
The spinal cord
The spinal cord is a glistening white tube running from the medulla oblongata of the brain to the level of the sixth or seventh lumbar vertebra, where it breaks up into several terminal spinal nerves forming a structure known as the cauda equina (Fig. 5.9). It leaves the skull through the largest foramen in the body, the foramen magnum, and runs through the vertebral canal formed by the interlinking vertebrae that protect it. The cord decreases in size from its cranial to caudal end, but in the caudal cervical and midlumbar regions it may be thicker as these correspond to areas where collections or plexuses of nerves leave the spinal cord to supply the fore and hindlimbs.
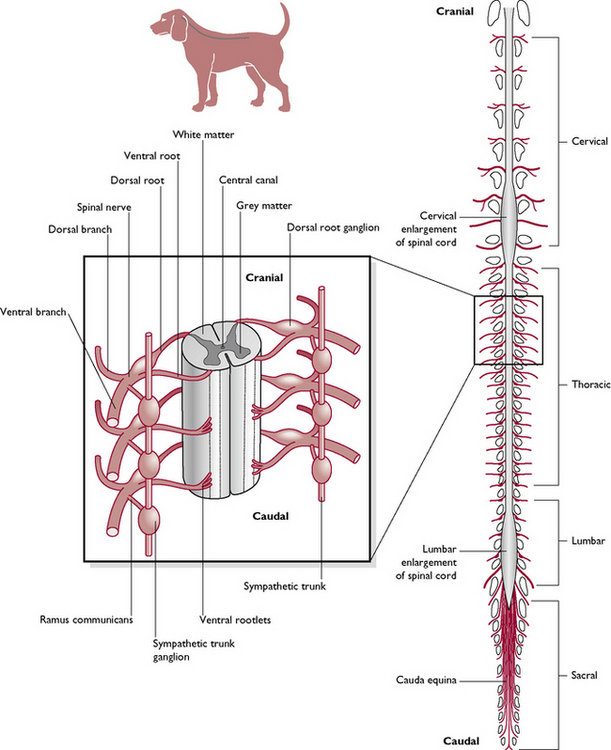
The spinal cord develops segmentally in the embryo and, although this is difficult to see in the adult, each segment corresponds to a different vertebra and gives off a pair of spinal nerves – one to the right and one to the left. The spinal nerves leave the vertebral canal through the appropriate intervertebral foramen.
In cross-section (Fig. 5.10) the spinal cord consists of:
The peripheral nervous system
The peripheral nervous system consists of all the nerves given off from the central nervous system. These are:
Cranial nerves
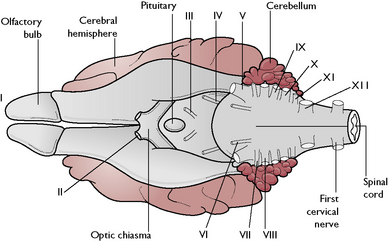
The 12 pairs of cranial nerves arise from the brain (Fig. 5.6) and leave the cranial cavity by various foramina. The majority of these nerves supply structures around the head and are relatively short, but some supply structures at some distance from the point at which they leave the brain, e.g. the vagus nerve (cranial nerve X), which is the longest nerve in the body.
Cranial nerves are referred to by their name and by a Roman numeral (Table 5.1). They may carry:
| Cranial nerve | Type of nerve fibre | Function |
|---|---|---|
| I Olfactory | Sensory | Carries the sense of smell or olfaction from the olfactory bulbs to the brain |
| II Optic | Sensory | Carries information about sight from the eyes to the brain; some nerve fibres from each eye cross over, via the optic chiasma, to the opposite side of the brain – in this way each side of the brain receives information from both eyes |
| III Oculomotor | Motor | Supplies the extrinsic muscles of the eye, enabling the eye to make minute, delicate movements; accurate movement of the eye is essential for survival |
| IV Trochlear | Motor | Supplies the extrinsic muscles of the eye |
| V Trigeminal | Mixed | Sensory fibres are carried from the skin around the eye and the face; motor fibres supplied to the muscles of mastication – mainly the temporal and masseter |
| VI Abducens | Motor | Supplies the extrinsic muscles of the eye |
| VII Facial | Motor | Supplies the muscles of facial expression including those associated with the movement of the lips, ears and skin around the eyes |
| VIII Vestibulocochlear (auditory) | Sensory | Vestibular branch carries the sensation of balance from the semicircular canals of the inner ear; cochlear branch carries the sensation of hearing from the cochlea of the inner ear |
| IX Glossopharyngeal | Mixed | Carries the sensation of taste from the taste buds on the tongue and pharynx; supplies motor fibres to the muscles of the pharynx |
| X Vagus | Mixed | Carries sensory fibres from the pharynx and larynx; supplies motor fibres to the muscles of the larynx; parasympathetic visceral motor fibres to the heart and various thoracic and abdominal organs including the gastrointestinal tract down to the descending colon |
| XI Accessory (spinal accessory) | Motor | Supplies the muscles of the neck and shoulder |
| XII Hypoglossal | Motor | Supplies the muscles of the tongue |
Spinal nerves
As the spinal cord passes down the vertebral canal each segment, corresponding to a vertebra, gives off a pair of spinal nerves – one to the right side and one to the left (Fig. 5.9). These nerves are numbered according to the number of the spinal vertebra in front. However, there are eight cervical nerves but only seven cervical vertebrae – the first cervical nerve (Ce1) leaves the spinal cord in front of C1; Ce7 leaves in front of C7 and Ce 8 leaves behind it. From then on, each nerve leaves from behind the appropriate vertebra.
Each spinal nerve consists of two components (Fig. 5.10):
In some nerve pathways, there may be one or more intercalated neurons lying in the grey matter between the sensory and motor neurons. The sensory and motor nerve fibres of each spinal nerve come together in the same myelin sheath and leave the vertebral column via the relevant intervertebral foramen as a mixed nerve (Fig. 5.9).
Examples of major peripheral nerve pathways within the limbs
Pectoral limb
Most structures within the pectoral limb are supplied by the brachial plexus, which lies above the area of the axilla and just below the spinal cord. It is formed by the spinal nerves from C6, C7, C8, T1 and T2, which combine to form various named nerves within the limb.
If an animal damages the radial nerve in its forelimb it will knuckle over on its forepaw leading to excessive wear of the skin on the dorsal surface of the paw – the ventral surface is protected from normal wear by the pads. This occurs because the animal is unable to use the extensor muscles that bring the limb into its normal position. The damaged nerve may recover but if it has been severed the only treatment may be to amputate the forelimb.
Pelvic limb
Structures within the hind limb are supplied by nerves from the lumbosacral plexus formed by the spinal nerves from L3, L4, L5, L6, L7, S1 and S2. These combine to form the various named nerves of the hind limb.
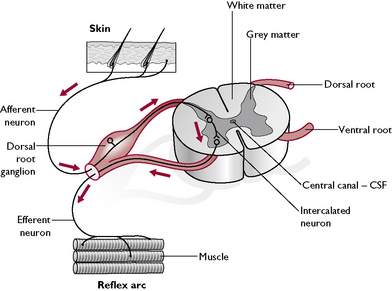
Reflex arcs
A reflex arc is a fixed involuntary response to certain stimuli. The response is always the same. It is rapid and automatic and involves only nerve pathways in the spinal cord, using the appropriate spinal nerve. Reflex arcs are a means of protection and produce a prompt response to potentially damaging phenomena such as heat or sharp objects.
Common reflexes observed in the dog and cat are:
Using the patellar reflex as an example, we can follow the pathway taken by the nerve impulse (Fig. 5.10). The tendon of the quadriceps femoris muscle passes over the patella as the straight patellar ligament and inserts on the tibial tuberosity of the proximal tibia. Within the muscle fibres are stretch receptors known as muscle spindles, whose function is to monitor muscle tone. The nerve pathway is as follows:
This is an example of a monosynaptic reflex as there is only one set of synapses. A polysynaptic reflex, e.g. the withdrawal reflex, involves one or more intercalated neurons and several synapses within the grey matter.
A reflex is unconscious and will still occur even if the spinal cord has been severed. However, if the web of the dog’s foot is pinched and it withdraws its foot and then yelps or bites you, this indicates that it has felt pain. There is conscious perception, involving the transmission of impulses up the tracts of white matter of the spinal cord to the brain and back to the muscles of the jaw, as well as along the pathway of the reflex arc. A conscious response indicates that the spinal cord is intact.
Reflexes may be described as being:
Conditioned reflexes are the basis of toilet training and of obedience training using ‘clickers’.
Autonomic nervous system
This can be considered to consist almost entirely of visceral motor nerves supplying cardiac muscle, smooth muscle and glandular tissue (including the liver and pancreas) of all the internal organs and of blood vessels. Control of the system is unconscious – an animal does not have to push food along its intestine or constrict its blood vessels by conscious thought (Fig. 5.11).
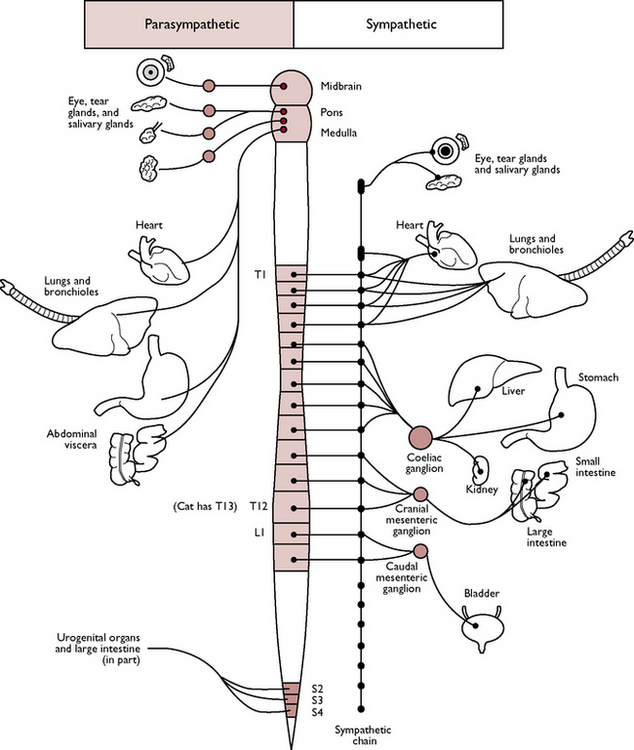
Fig. 5.11 Outline of the autonomic nervous system.
(With permission from Colville T, Bassett JM 2001 Clinical anatomy and physiology for veterinary technicians. Mosby, St Louis, MO, p 156.)
The autonomic nervous system can be divided into two parts: the sympathetic nervous system and the parasympathetic nervous system. The division is based on the anatomical derivation of the spinal nerves and on their differing effects. Most organs are supplied by both types and control involves a balance between the two (Table 5.2).
Table 5.2 The autonomic nervous system
| Sympathetic system | Parasympathetic system | |
|---|---|---|
| Origin of nerve fibres | Vertebrae T1 to L4 or L5 | Cranial nerves III, VII, IX, X; vertebrae S1, S2 |
| Preganglionic nerve fibres | Short; each nerve leads to a ganglion containing cell bodies and lying close under the vertebral column; there is a chain of ganglia – the sympathetic chain – one on each side of the vertebral column | Long; ganglia lie close to the organ they supply; there is no chain of ganglia |
| Postganglionic nerve fibres | Long; lead away from the sympathetic chain and travel towards the organ it supplies – usually follow the path of blood vessels | Short; fibres run a short distance from the ganglion to the organ |
| Areas supplied | Viscera in thorax, abdomen and pelvis; also supply sweat glands, blood vessels and piloerector muscles associated with hair follicles. Most ganglia are paired, but three are unpaired: | Structures in the head including the eye and salivary glands; vagus (X) supplies the heart, lungs, stomach, small intestine, pancreas and large intestine |
| Transmitter substances: | ||
| A. Within the system, i.e. between cell body and dendron | Acetylcholine | Acetylcholine |
| B. At the terminal synapses, i.e. between the axon and the effector organ | Noradrenaline (norepinephrine) | Acetylcholine |
| General effect | Prepares the body for ‘fear, flight, fight’; heart and respiratory rates are increased, blood vessels to skeletal muscle are dilated, blood glucose levels rise, piloerector muscles to the hairs contract so hackles are raised, gastrointestinal tract activity decreases | Animal is more relaxed; heart rate is slowed, respiratory bronchioles constrict, gastrointestinal tract activity increases – digestive juices and salivary secretion increases, peristalsis increases |
Special senses
Specialised receptor cells are adapted to respond to a particular stimulus and convey the information to the CNS. Information is processed within the CNS and an appropriate response is initiated. Mammals have evolved specialised organs in which to house the receptor cells – these are the special sense organs.
Taste buds – taste
The sensation of taste is known as gustation and the organs used to detect it are the taste buds (Fig. 5.12) found on the dorsal surface of the tongue, the epiglottis and the soft palate and covered in moist mucous membrane. Taste buds are chemoreceptors stimulated by the chemicals responsible for smell dissolved in the mucus around the oral cavity.
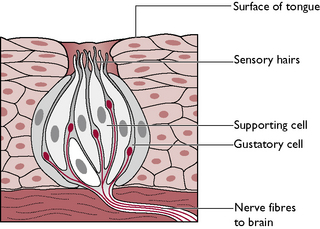
Each taste bud consists of receptor or gustatory cells surrounded by sustentacular or supporting cells. Projecting from the upper surface of each taste bud are hair-like processes. Fine nerve fibres which are branches of the facial (VII), glossopharyngeal (IX) and vagus (X) nerves conduct impulses away from the gustatory cells towards the brain, where they are interpreted as taste. The sensation of taste in animals is difficult to quantify. It is thought to be directed more towards selecting tastes that are harmless or harmful to them rather than to enjoyment or dislike, as in humans.
Nasal mucosa – smell
The sensation of smell is known as olfaction. The receptor cells are also chemoreceptors and the sensation is closely allied to gustation – the two sensations often work together. The receptor cells are rod-shaped bipolar neurons which are distributed throughout the mucosa covering the caudal part of the nasal cavities and the turbinate bones. The axon leaving each receptor cell combines with other axons to form the olfactory nerve fibres (I). Chemicals responsible for smell dissolve in the mucus of the nasal cavity and stimulate the production of nerve impulses. These are transmitted along the olfactory nerve fibres, through the cribriform plate of the ethmoid bone (dividing the nasal and cranial cavities) and into the olfactory bulbs of the forebrain, where they are interpreted as smell.
The eye and sight
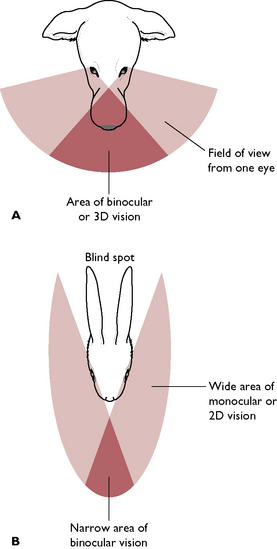
The eye is the organ of sight. Photoreceptor cells adapted to respond to the stimulus of light are found in the innermost layer of the eye – the retina. All mammals have a pair of eyes, each of which lies within a deep bony cavity of the skull known as the orbit. Each eye lies rostral to the cranial cavity, lateral to the nasal cavities and dorsal to the mouth. The dog and the cat are predatory species and their eyes point forwards (Fig. 5.13). This provides a wide area of binocular or 3D vision, enabling them to pinpoint the position of their prey accurately. Prey species such as the rabbit or the mouse have prominent eyes set on the sides of the head. These provide a wide area of monocular or 2D vision, which enables the animal to see the predator but not to fix its position – this does not matter, the important factor is that the predator is nearby and that the prey animal runs away.
Each eye consists of three main parts: the eyeball, the extrinsic muscles and the eyelids.
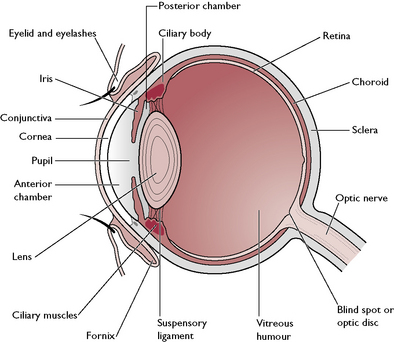
The eyeball
The eyeball (Fig. 5.14) is a globe-shaped structure made of three layers:
Sclera
The sclera forms the fibrous outer covering of the eye, in conjunction with the cornea.
The junction between the cornea and sclera is known as the limbus. This is the drainage point for the aqueous humour of the anterior chamber of the eye.
Uvea
The uvea, a vascular pigmented layer, is firmly attached to the sclera at the exit of the optic nerve but is less well attached in other areas (Fig. 5.14). It is made up of the following parts:
Retina
The retina is the innermost layer of the eye. Light is focused onto the photoreceptor or light sensitive cells of the retina by the lens and information is transmitted to the brain via the optic nerve (II). The retina is made of several layers (Fig. 5.16). Light travels through the outer layers before it stimulates the deeper photoreceptor cells.
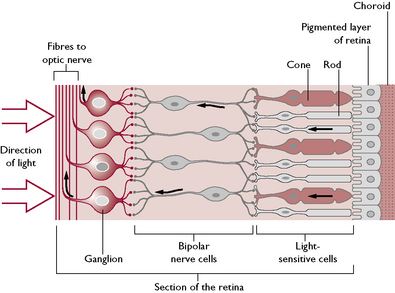
From the layer closest to the choroid the retina layers are as follows:
Within the centre of the eye is the lens – a transparent biconvex disc surrounded by the suspensory ligament and the ciliary body (Fig. 5.14). The tissue of the lens is crystalline and elastic, and its shape is altered by the contraction of the ciliary muscles around it. This enables the lens to focus rays of light on to the retina.
The iris divides the anterior part of the eye into two chambers:
The posterior part of the eye lying between the lens and the retina is known as the vitreous chamber. This occupies 80% of the volume of the eye and contains a clear, jelly-like substance known as vitreous humour.
The function of both the aqueous and vitreous humours is to provide nutrients for the structures within the eye and to maintain the shape of the eye.
The extrinsic muscles
The eyeball is well supplied with extrinsic muscles (see Ch. 4, Fig. 4.5). These originate on the connective tissue of the sclera and insert on the periosteum of the bones forming the orbit of the skull. They are essential for the fine movements of the eyeballs, which must be coordinated so that the eyes work together to achieve accurate binocular or 3D vision. The nerve supply to the extrinsic muscles is via the oculomotor (III), trochlear (IV) and abducens (VI) nerves.
In some medical conditions, e.g. diabetes mellitus, or as a result of old age or genetic factors, the lens may develop a cataract. The lens becomes opaque, which interferes with the passage of light rays to the retina, and the animal becomes blind. In some cases, the condition may be able to be treated by removal of the lens but the animal will then only have distance vision.
Glaucoma, or an increase in intraocular pressure, results if there is overproduction of aqueous humour or blockage of the fluid drainage point at the limbus of the eye. It may also be a symptom of hypertension, as seen in cats with chronic renal failure. Intraocular pressure is measured using a tonometer.
The eyelids
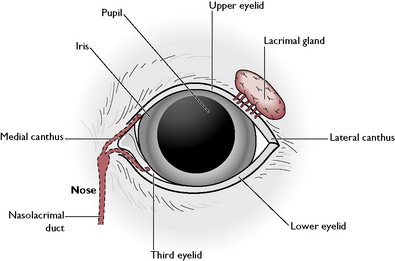
The bony orbit of the skull protects the majority of the eyeball but the anterior third is soft, relatively protuberant and at risk from mechanical damage. It is protected by a set of eyelids and associated glands. Each eye has an upper and lower eyelid joined at the medial canthus adjacent to the nose and at the lateral canthus (Fig. 5.17). The upper eyelid is more mobile than the lower one. Each eyelid is covered by hairy skin on the outer surface and lined by a layer of mucous membrane on the inner surface. This layer is continuous with the conjunctiva covering the cornea. The point where it reflects back is known as the fornix (Fig. 5.14). The inner surface of the eyelids is protected and lubricated by the Meibomian glands. Growing from the outer edge of the upper eyelids are well-defined cilia, resembling human eyelashes. These may be present but less obvious on the lower lids. Associated with these are sebaceous and rudimentary sweat glands.
In the dog, the following conditions associated with the eyelashes may be inherited characteristics:
Within the tissue of the eyelids are the palpebral muscles responsible for eyelid movement. The palpebral or blink reflex protects the eye from damage and is a useful means of assessing the depth of anaesthesia – it disappears as the level of anaesthesia deepens. Deep to the upper and lower eyelids, and lying in the medial canthus of each eye, is the third eyelid or nictitating membrane. In the dog, it is a T-shaped piece of hyaline cartilage and smooth muscle; in the cat it is made of elastic cartilage and smooth muscle. The third eyelid is covered on both sides by mucous membrane, which is continuous with the lining of the eyelids and the cornea. It is supplied with glandular and lymphoid tissue to protect the eye and keep it moist. Nerve supply to the third eye is via the sympathetic system.
The surface of the cornea is kept moist and protected from damage by the lacrimal gland, a modified cutaneous gland lying on the dorsolateral surface of eyeball beneath the upper eyelid (Fig. 5.17). This produces secretions known as tears which flow over the anterior surface of the eye and drain into two small openings in the area of the medial canthus. These openings lead into the nasolacrimal duct, which leaves the bony orbit and runs through the nasal cavity to open into the nostril.
The Harderian glands lie underneath the third eyelid and contribute to the lubricating and protective function of tears. In some dogs, particularly in brachycephalic breeds such as the bulldog, the Harderian glands may become enlarged and protrude from under the third eyelid. This is a condition often referred to as ‘cherry eye’ because the inflamed gland resembles a cherry. The treatment may be to attempt to push the gland back behind the eyelid or to remove it completely.
Formation of an image
The function of the eye is to conduct rays of light from an object and form an image on the retina (Fig. 5.18). This occurs in the following stages:
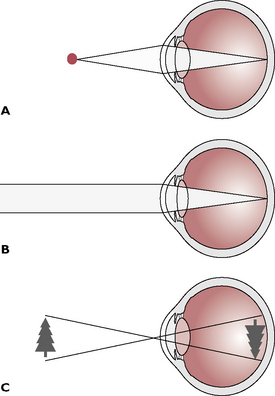
Fig. 5.18 Formation of the visual image. A Light rays from close objects are diverging (spreading out) as they reach the eye. The light is focussed by contraction of the ciliary muscles, which causes the lens to shorten and flatten, bending the light on to the retina. B Light rays from distant objects are effectively parallel as they reach the eye. These rays are focussed by relaxation of the ciliary muscles, which causes the lens to elongate and thin. C The image on the retina is upside down when it forms; the brain inverts this, allowing normal perception of the image the correct way up.
The ability to accommodate, or change the shape of the lens to focus on near and far objects, develops soon after birth. As the lens ages and becomes less flexible, the time taken for accommodation increases – in man this can be corrected by the use of glasses but animals must learn to adapt their behaviour to cope with this ‘old age’ change.
The ear – hearing and balance
The ear is the organ of hearing and balance. Receptor cells adapted to respond to sound waves and to the movement of the body are located within the inner ear. All mammals have a pair of ears located on the dorsal part of the head. Each ear can be divided into three parts:
The external ear
This consists of the following structures:
 To guide sound waves picked up from the external environment down the external auditory meatus to the tympanic membrane (ear drum) To act as a means of facial expression and communication between animals of the same species. Pinnae are used in conjunction with other parts of the body, e.g. the eyes and tail, to communicate dominance, fear, acceptance, etc.
To guide sound waves picked up from the external environment down the external auditory meatus to the tympanic membrane (ear drum) To act as a means of facial expression and communication between animals of the same species. Pinnae are used in conjunction with other parts of the body, e.g. the eyes and tail, to communicate dominance, fear, acceptance, etc.
The pinna varies in size and shape between breeds and has been altered by generations of selective breeding, leading to the development of characteristics many of which predispose to ear disease. The original wolf-like dog, from which all breeds are thought to be derived, has upright V-shaped ears – this is still seen in German Shepherds, Chows and Huskies. All cats have upright pointed ears that, until the recent introduction of the Scottish Fold with its twisted, flattened ear pinnae, have been left untouched by selection.
Each pinna is formed from a sheet of elastic cartilage known as the auricular cartilage. This continues as an incomplete tube known as the external auditory meatus linked at the base to the annular cartilage. This in turn articulates with the external acoustic process of the tympanic bulla, forming a series of interlinking cartilages that enables the pinna to move freely to pick up sound waves.
Both sides of the auricular cartilage are covered in hairy skin – in most breeds the outer side is more hairy than the inner side. The covering of skin continues into the external auditory meatus where there are fewer hair follicles; the Poodle is particularly noted for its hairy ear canals. The skin is well supplied with ceruminous glands – modified sebaceous glands that secrete wax and protect the ear canal from damage and infection.
Otitis externa, or infection of the ear canal, is a common condition in dogs and cats. It may be caused by a variety of microorganisms, such as ear mites (Otodectes cynotis), bacteria and fungi, such as Malassezia pachydermatis. The shape of the pinna and the presence of hair may predispose to infection by creating a hot airless environment within the ear canal. Infection results in pain, aural discharge, self-trauma and, in some cases, aural haematoma.
The middle ear
The middle ear (Fig. 5.19) consists of:
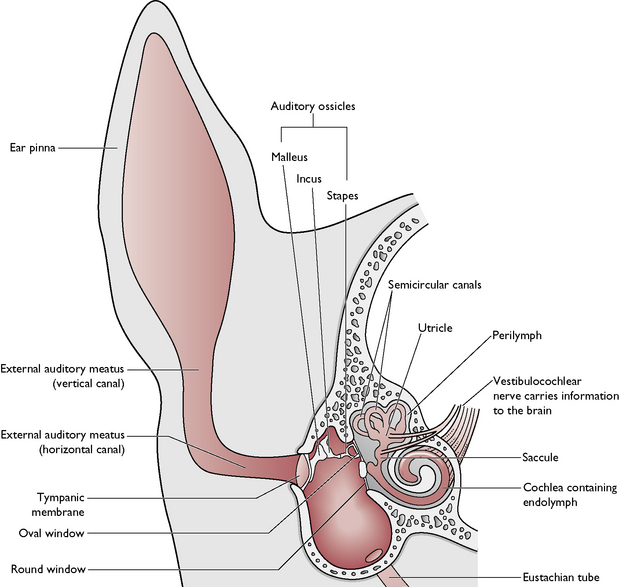
The function of the ossicles is to transmit the vibrations caused by sound waves across the middle ear to the inner ear.
The inner ear
The inner ear (Fig. 5.19) consists of:
Membranous cochlea – this has a similar shape to a snail’s shell. It forms a blind-ending ventral spiral and is filled with endolymph. Within the cochlea are a group of sensory receptor cells forming the spiral organ of Corti, whose function is to detect sound. Projecting from each receptor cell is a sensory hair and at the base is a nerve fibre that is part of the cochlear branch of the vestibulocochlear nerve (VII).
Perception of sound. Sound waves, picked up by the pinna and transmitted across the ear by the tympanic membrane and auditory ossicles to the oval window, set up vibrations or ripples first in the perilymph and then the endolymph. These ripples move the sensory hairs of the receptor cells in the organ of Corti, and this sends nerve impulses along their associated nerve fibres. The impulses are carried by the cochlear branch of the vestibulocochlear nerve (VIII) to the auditory cortex of the cerebral hemispheres, where they are interpreted as sound.
Membranous vestibule – this is made of two sac-like structures, the utricle (Fig. 5.20) and saccule, which connect with the cochlea and the semicircular canals and are filled with endolymph. Both these structures contain areas of sensory hair cells known as maculae, surrounded by jelly-like material containing calcium carbonate particles or otoliths. The function of these structures is to maintain balance when the animal is standing still.
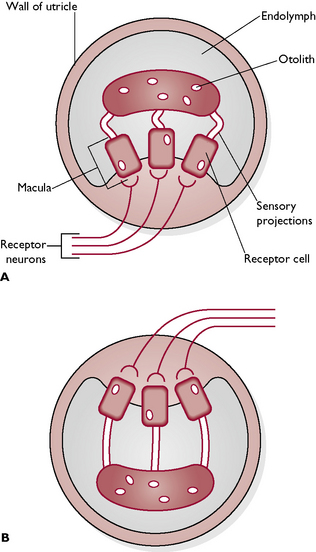
Fig. 5.20 Simplified section through a utricle. A With the head upright, there is no pull on the sensory projections of the utricle. B When the head is tilted or upside down, the otoliths fall away from the macula. The sensory projections become stretched and this causes impulses to be set up in the receptor cells. These are transmitted to the brain in the receptor neurons, giving information about the orientation of the head.
Static balance. When an animal is standing still, the head still makes minute movements. The pull of gravity moves the jelly-like material within the maculae of the utricle and saccule. This moves the sensory hairs, which transmit nerve impulses to the brain via the vestibular branch of the vestibulocochlear nerve (VIII). The brain interprets this information in terms of the position of the body in relation to the space around it.
Membranous semicircular canals – these are three canals, filled with endolymph, each of which describes two-thirds of a circle. The plane of each circle is approximately at right angles to the other two – in this way, the three dimensions of movement are monitored. Each canal is connected to the utricle by a swollen area known as an ampulla. Inside each ampulla is a cone-shaped projection or crista containing sensory hair cells embedded in a jelly-like cupula. The function of these structures is to maintain balance during movement.
Dynamic balance. As the animal moves, the endolymph in the semicircular canals moves and stimulates the cristae within the ampullae. Nerve impulses are transmitted by the hair cells to the brain via the vestibular branch of the vestibulocochlear nerve (VIII). Within the brain, they are passed to the cerebellum, where the information is coordinated.