16 Streptococcus and enterococcus
Pharyngitis; scarlet fever; skin and soft tissue infections; streptococcal toxic shock syndrome; pneumonia; meningitis; urinary tract infections; rheumatic fever; post-streptococcal glomerulonephritis
Key points
• Str. pyogenes (group A streptococcus) is among the most prevalent of human bacterial pathogens.
• Str. pyogenes and group C and G streptococcal infections range from sore throat, scarlet fever and superficial skin infections to invasive soft tissue infections and septicaemia.
• Str. pyogenes and pyogenic group C and G streptococci produce several superantigenic extracellular toxins that are involved in the pathogenesis of the rash associated with scarlet fever and streptococcal toxic shock syndrome.
• Rheumatic fever and acute glomerulonephritis are potential immune-mediated sequelae of infections with Str. pyogenes.
• Str. agalactiae (group B streptococci) causes neonatal septicaemia and meningitis.
• Str. pneumoniae is the principal cause of pneumonia, middle ear infections and meningitis and is one of the four most frequent causes of fatal infections worldwide.
• Commensal streptococci of the oral cavity are among the most frequent causes of subacute bacterial endocarditis.
Streptococci is the general term for a diverse collection of Gram-positive cocci that typically grow as chains or pairs (Greek: streptos = pliant or chain; coccos = a grain or berry) (Fig. 16.1). Virtually all of the streptococci that are important in human medicine and dentistry fall into the genera Streptococcus and Enterococcus. Occasional opportunistic infections are associated with other genera of streptococci, such as Peptostreptococcus (see Ch. 36) and Abiotrophia (‘nutritionally variant streptococci’).
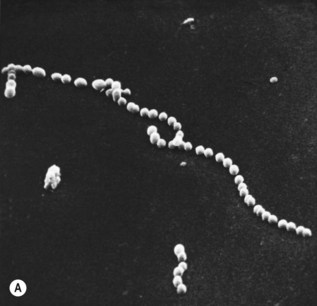
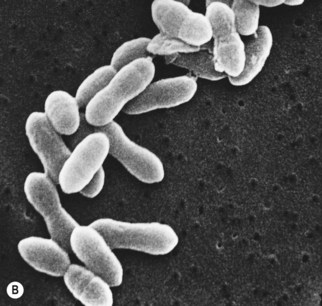
Fig. 16.1 Scanning electron micrograph of (A) Str. pyogenes showing typical chain formation (original magnification ×2000) and (B) Str. pneumoniae showing typical diplococcus formation (original magnification ×7000).
Courtesy of AP Shelton, University Hospital, Nottingham.
Streptococci are generally strong fermenters of carbohydrates, resulting in the production of lactic acid, a property responsible for the involvement of some oral streptococci in the decalcification of teeth, i.e. dental caries, and also used in the dairy industry. Most are facultative anaerobes, although peptostreptococci are obligate anaerobes. Streptococci do not produce spores and are non-motile. They are catalase negative.
Classification
The genus Streptococcus includes important pathogens and commensals of mucosal membranes of the upper respiratory tract and, for some species, the intestines. The genus Enterococcus, which is also an intestinal commensal, is related to the other streptococci, but is classified separately.
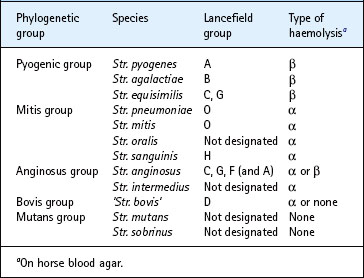
The genus Streptococcus includes more than 70 species. With few exceptions, the individual species are exclusively associated, as either pathogens or commensals, with man or a particular animal. The genus consists of five clusters of species (Table 16.1), each of which is characterized by distinct pathogenic potential and other properties:
• The pyogenic (pus generating) group includes most species that are overt human and animal pathogens.
• The mitis group includes commensals of the human oral cavity and pharynx, although one of the species, Streptococcus pneumoniae, is also one of the most important human pathogens.
• The anginosus and salivarius groups are part of the commensal microbiota of the oral cavity and pharynx.
• The bovis group belongs in the colon.
• The mutans group of streptococci colonizes exclusively the tooth surfaces of man and some animals; some species belonging to this cluster are involved in the development of dental caries.
Virtually all of the commensal species, including the enterococci, are opportunistic pathogens, primarily if they gain access to the bloodstream from the oral cavity or from the gut.
Haemolytic activity
Early attempts to distinguish between pathogenic and commensal streptococci recognized different types of reactions around colonies on blood agar plates. Colonies of streptococci belonging to the Pyogenic group are generally surrounded by a clear zone, usually several millimetres in diameter, caused by lysis of red blood cells in the agar medium induced by bacterial haemolysins. This is called β-haemolysis and constitutes the principal marker for potentially pathogenic streptococci in cultures of throat swabs or other clinical samples. Accordingly, the pyogenic streptococci are also referred to as haemolytic streptococci.
In contrast, most commensal streptococci give rise to a green discoloration around colonies on blood agar. This phenomenon is termed α-haemolysis, although not caused by haemolysis. The factor causing the green discoloration is hydrogen peroxide, which oxidizes haemoglobin to the green methaemoglobin.
Collectively, commensal streptococci are often called ‘viridans streptococci’, which refers to their α-haemolytic property (viridis = green). Not quite logically, this term also includes the few streptococci, such as those of the salivarius and mutans groups, that induce neither α- nor β-haemolysis. Moreover, in common usage, the term excludes Str. pneumoniae, although this species is also α-haemolytic.
Lancefield grouping
An important method of distinguishing between pyogenic streptococci is the serological classification pioneered by the American bacteriologist Rebecca Lancefield, who detected different versions of the major cell wall polysaccharide among the pyogenic streptococci.
This polysaccharide can be extracted from streptococci with hot hydrochloric acid and the different forms can be distinguished by precipitation with specific antibodies raised in rabbits. The polysaccharide is referred to as the group polysaccharide and identifies a number of different serological groups labelled by capital letters (Lancefield groups A, B, C, etc.). Among the pyogenic streptococci, some serological groups are identical to distinct species (Table 16.1).
Streptococcus pyogenes
This species, which consists of Lancefield group A streptococci, is among the most prevalent of human bacterial pathogens. It is associated exclusively with human infections. It causes a wide range of suppurative infections in the respiratory tract and skin, life-threatening soft tissue infections, and certain types of toxin-associated reactions. Some of these infections may, in addition, result in severe non-suppurative sequelae due to adverse immunological reactions induced by the infecting streptococci. A similar spectrum of infections may be caused by the closely related group C and group G streptococcis (Str. equisimilis, also known as Str. dysgalactiae subspecies equisimilis).
Some of the infections caused by Str. pyogenes resemble those caused by Staphylococcus aureus, but the clinical characteristics associated with these two groups of pyogenic cocci are often distinct. Similarities and differences can be explained by the virulence factors expressed by the two species.
Pathogenesis
Virulence factors
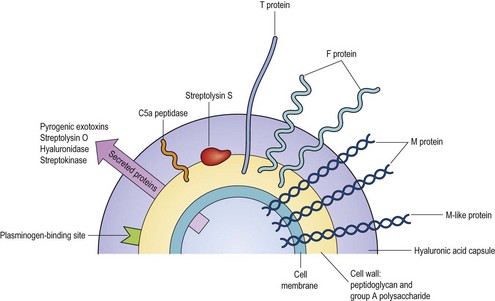
Strains of Str. pyogenes express a large arsenal of virulence factors involved in adherence, evasion of host immunity, and tissue damage (Fig. 16.2). Although some factors are expressed by all clinical isolates, others are variably present among Str. pyogenes strains. This variation is due to the horizontal transfer of virulence genes among strains, primarily by bacteriophages (transduction, see p. 75), and probably explains the temporal variations in the prevalence of severe infections and sequelae. It furthermore explains differences in the virulence of individual strains and, to some extent, the different clinical pictures that may be associated with infections due to Str. pyogenes. Many of the virulence factors of Str. pyogenes are also expressed by some of the other species of pyogenic streptococci. In some species pathogenic for animals the corresponding virulence factors are expressed in a form specifically adapted to interact with a particular host.
Adhesion
Interaction with host fibronectin, a matrix protein on eukaryotic cells, is considered the principal mechanism by which Str. pyogenes binds to epithelial cells of the pharynx and skin. The structure that recognizes host fibronectin is located on the F protein, which is one of the many proteins expressed on the surface of Str. pyogenes (Fig. 16.2). The interaction between the streptococcal F protein and host cell fibronectin also mediates internalization of the bacteria into host cells.
In addition to the F protein, surface-exposed lipoteichoic acid and M proteins appear to be involved in adherence to mucosal and skin epithelial cells.
M proteins
The ability of Str. pyogenes to resist phagocytosis by polymorphonuclear leucocytes is largely due to the cell-surface exposed M protein. The M protein is anchored in the cytoplasmic membrane, spans the entire cell wall, and protrudes from the cell surface as fibrils (Fig. 16.2). Some strains produce two different M proteins with antiphagocytic activity, and some an additional structurally related M-like protein. All of these proteins can bind various plasma proteins of the host, including fibrinogen, plasminogen, albumin, immunoglobulin (Ig) G, IgA, the proteinase inhibitor α2-macroglobulin and some regulatory factors from the complement system (factor H and C4b-binding protein). As well as masking the bacterial surface with host proteins, some of these affinities are responsible for the ability of M proteins to resist phagocytosis. Thus, factor H can destabilize the important opsonin C3b when deposited on the bacterial surface. Likewise, the C4b-binding protein inhibits surface complement deposition by stimulating degradation of both C4b and C3b.
Str. pyogenes can release some of the M proteins with its own cysteine protease. If shed into the circulation the M protein forms complexes with fibrinogen, which by indirect pathways induces the release of inflammatory mediators from neutrophils. This process plays an important role in the leakage of plasma into the tissues and lungs, and the ensuing low blood pressure seen in some invasive infections with Str. pyogenes.
Acquired resistance to infection by Str. pyogenes is the result of antibodies to the M protein molecule in secretions and plasma. However, as a result of genetic polymorphism in the gene encoding the M protein, the most distal part of the protein shows extensive variability among strains. As a consequence, individuals may suffer from recurrent Str. pyogenes infections with strains expressing different versions of the M protein. More than 150 different types of M protein have been identified by serological means and gene sequencing.
Capsule
Some strains of Str. pyogenes form a capsule composed of hyaluronic acid. Such strains grow as mucoid colonies on blood agar and are highly virulent in animal models. Although capsule production is rare among isolates from uncomplicated pharyngitis, a significant proportion of isolates from severe infections have a capsule. Like other bacterial capsules, the capsule has an antiphagocytic effect. The relative significance of the M protein and the capsule as antiphagocytic factors differs among strains.
The capsule is identical to the hyaluronic acid of the connective tissue of the host and is not immunogenic. In this way the bacteria can disguise themselves with an immunological ‘self’ substance.
C5a peptidase
The C5a peptidase, which is also found in most human pathogenic strains of Str. agalactiae, is present on the surface of all strains of Str. pyogenes. It specifically cleaves, and thereby inactivates, human C5a, one of the principal chemo-attractants of phagocytic cells.
Streptolysins
Str. pyogenes produces two distinct haemolysins, termed streptolysins O (oxygen labile) and S (serum soluble), both of which lyse erythrocytes, polymorphonuclear leucocytes and platelets by forming pores in their cell membrane.
Streptolysin O belongs to a family of haemolysins found in many pathogenic bacteria. Intravenous injection into experimental animals causes death within seconds, as the result of an acute toxic action on the heart. Serum antibodies can be demonstrated after streptococcal infection, particularly after severe infections.
Streptolysin S is responsible for the β-haemolysis around colonies on blood agar plates. It can also induce the release of lysosomal contents with subsequent cell death after engulfment by phagocytes. In contrast to streptolysin O, it is not immunogenic.
Pyrogenic exotoxins
Most strains of Str. pyogenes produce one or more toxins that are called pyrogenic (fever generating) exotoxins because of their ability to induce fever. Three, designated SPE-A, SPE-B and SPE-C, have been characterized extensively, but there are several others. Purified SPE-A causes death when injected into rabbits and is the most toxic of the three, but SPE-B also causes myocardial necrosis and death in experimental animals.
The genes for SPE-A and SPE-C are transmitted between strains by bacteriophage, and stable production depends on lysogenic conversion in a manner analogous to toxin production by Corynebacterium diphtheriae (see p. 75). Even among strains that possess the genes, the quantity of toxin secreted varies dramatically.
SPE-A and SPE-C are also called erythrogenic toxins, as they are responsible for the rash observed in patients with scarlatina. They are genetically related to Staph. aureus enterotoxins and, like these, have superantigen activity. By cross-linking MHC class II molecules on antigen-presenting cells and the Vβ domain of the antigen receptor on a subset of T lymphocytes, these toxins cause comprehensive (antigen independent) activation of regulatory T lymphocytes. The result is massive release of pro-inflammatory cytokines such as interleukin (IL)-1 and IL-2, tumour necrosis factor (TNF)-α and interferon-γ. These cytokines may cause a variety of clinical signs, including inflammation, hypotensive shock and organ failure seen in some patients with severe streptococcal disease. The different clinical outcome of infections with the same Str. pyogenes strain appears to be due to the fact that the toxins bind preferentially to certain major histocompatibility complex (MHC) class II tissue types.
Unlike SPE-A and SPE-C, all strains of Str. pyogenes produce SPE-B, which is a potent cysteine proteinase capable of cleaving many host proteins.
None of the pyrogenic exotoxins is associated unambiguously with any of the clinical syndromes caused by Str. pyogenes. However, most isolates from episodes of severe invasive disease and toxic shock-like syndrome produce SPE-A, and the protease SPE-B appears to be responsible for the extensive tissue destruction observed in many patients with severe invasive infections, including necrotizing fasciitis.
Hyaluronidase
Str. pyogenes and several other pyogenic streptococci use a secreted hyaluronidase to degrade hyaluronic acid, the ground substance of host connective tissue. This property may facilitate the spread of infection along fascial planes. During infections, particularly those involving the skin, serum antibody titres to hyaluronidase show a significant rise.
Streptokinase
Streptokinase, also known as fibrinolysin, is another spreading factor. It is expressed by all strains of Str. pyogenes and cooperates with a surface-expressed plasminogen-binding site on the bacteria. Once host plasminogen is bound to the bacterial surface, it is activated to plasmin by streptokinase. Thus, in contrast to Staph. aureus, which aims at hiding behind a wall of coagulated plasma (fibrin), Str. pyogenes employs host plasmin to hinder the build-up of fibrin barriers. As a result, soft tissue infections due to Str. pyogenes are more diffuse, and often rapidly spreading, than the well localized abscesses that typify staphylococcal infections.
Clinical features
Although a general decrease in the prevalence of serious infections with Str. pyogenes has occurred since the mid nineteenth century, there has been a resurgence in severe streptococcal infections and increased mortality due to streptococcal sepsis since the early 1980s.
The most common route of entry of Str. pyogenes is the upper respiratory tract, which is usually the primary site of infection and also serves as a focus for other types of infection. Spread from person to person is by respiratory droplets or by direct contact with infected wounds or sores on the skin. Not all individuals colonized by Str. pyogenes in the upper respiratory tract develop clinical signs of infection.
After an acute upper respiratory tract infection, the convalescent patient may carry the infecting streptococci for some weeks. Only a few healthy adults carry Str. pyogenes in the respiratory tract, but the carriage rate in young school children is just over 10%. It may be considerably higher before or during an epidemic.
Non-invasive streptococcal disease
The most common infections caused by Str. pyogenes are relatively mild and non-invasive infections of the upper respiratory tract (pharyngitis) and skin (impetigo). In the USA more than 10 million cases of non-invasive Str. pyogenes infection are estimated to occur annually.
Pharyngitis
This is the most common infection caused by Str. pyogenes. Clinical signs such as abrupt onset of sore throat, fever, malaise and headache generally develop 2–4 days after exposure to the pathogen. The posterior pharynx is usually diffusely reddened, with enlarged tonsils that may show patches of grey–white exudate on their surface and, sometimes, accumulations of pus in the crypts. The local inflammation results in swelling of cervical lymph nodes. Occasionally, tonsillar abscesses develop; this is a very painful condition and potentially dangerous as the pathogen may spread to neighbouring regions and to the bloodstream.
Despite the significant symptoms and clinical signs, differentiating streptococcal pharyngitis (‘strep throat’) from viral pharyngitis is impossible without microbiological or serological examination. Culture studies show that 20–30% of cases of pharyngitis are associated with Str. pyogenes and Str. equisimilis.
Scarlet fever
Pharyngitis caused by certain pyrogenic exotoxin-producing strains of Str. pyogenes may be associated with a diffuse erythematous rash of the skin and mucous membranes (Fig. 16.3). The condition is known as scarlet fever or scarlatina. The rash develops within 1–2 days after the first symptoms of pharyngitis and initially appears on the upper chest, then spreading to the extremities. After an initial phase with a yellowish-white coating, the tongue becomes red and denuded (’strawberry tongue’).
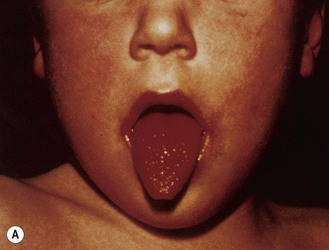
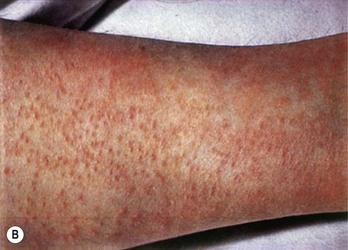
Fig. 16.3 The characteristic erythematous rash on (A) tongue and (B) skin associated with scarlet fever.
Between 1860 and 1870 the mean annual death rate from scarlet fever in England and Wales was close to 2500 per million of the population. Since then a steady decline in incidence has been observed. By the end of the twentieth century the number of scarlet fever cases in England and Wales had fallen below 25 per million, and death is now extremely rare.
Skin infections
Str. pyogenes may cause several types of skin infection, sometimes in association with Staph. aureus. The superficial and localized skin infection, known as impetigo or pyoderma, occurs mainly in children (Fig. 16.4). It primarily affects exposed areas on the face, arms or legs. The skin becomes colonized after contact with an infected person and the bacteria enter the skin through small defects. Initially, clear vesicles develop, which within a few days become pus filled. Secondary spread is often seen as a result of scratching.

Potentially more severe is the acute skin infection erysipelas (erythros = red; pella = skin), which occurs in the superficial layers of the skin (cellulitis) and involves the lymphatics. The infection is characterized by diffuse redness of the skin, and patients experience local pain, enlargement of regional lymph nodes and fever. Untreated, the infection may spread to the bloodstream, and was often fatal before antibiotics became available.
Invasive soft tissue infections
Severe, sometimes life-threatening, Str. pyogenes infections may occur when the bacteria get into normally sterile parts of the body. The most severe forms of invasive infections are necrotizing fasciitis, streptococcal toxic shock syndrome and puerperal fever, all of which are associated with bacteraemia.
Worldwide, rates of invasive infection increased from the mid 1980s to the early 1990s. In 2002, around 9000 cases of invasive Str. pyogenes infection occurred in the USA. Although these infections may occur in previously healthy individuals, patients with chronic illnesses such as cancer and diabetes, and those on kidney dialysis or receiving steroids, have a higher risk. Even with antibiotic treatment death occurs in 10–13% of all invasive cases: 45% of patients with toxic shock syndrome and 25% of patients with necrotizing fasciitis.
Necrotizing fasciitis
This infection progresses very rapidly, destroying fat and fascia. Although Str. pyogenes gains entry to these tissues through the skin after trauma, often of a minor nature, the skin itself may show only minimal signs of infection and may indeed be spared (Fig. 16.5). Systemic shock and general deterioration occur very quickly. The disease affects the fit young person with no obvious underlying pathology, as well as the immunocompromised.
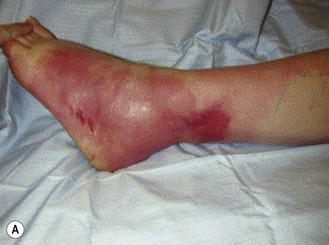
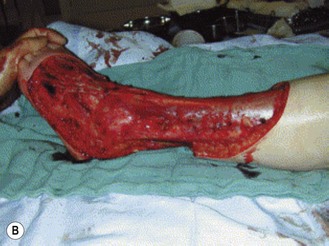
Fig. 16.5 Necrotizing fasciitis (A) before and (B) after surgical exploration and debridement.
Courtesy of Dr M Llewelyn, reproduced with permission from Elsevier.
The clinical diagnosis may be difficult because Staph. aureus and anaerobes such as Clostridium perfringens can produce a similar clinical picture. Streptococci can be isolated from the blood, blister fluid and cultures of the infected area.
Streptococcal toxic shock syndrome
Patients with invasive and bacteraemic Str. pyogenes infections, and in particular necrotizing fasciitis, may develop streptococcal toxic shock syndrome. The disease, which was first described in the late 1980s, is the result of the release of streptococcal toxins to the bloodstream. A striking feature of this acute fulminating disease is severe pain at the site of initial infection, usually the soft tissues. The additional clinical signs resemble those of staphylococcal toxic shock syndrome (see p. 178) and include fever, malaise, nausea, vomiting and diarrhoea, dizziness, confusion and a flat rash over large parts of the body. Without treatment, the disease progresses to shock and general organ failure.
Other suppurative infections
Historically, Str. pyogenes has been an important cause of puerperal sepsis. However, since the introduction of antibiotic therapy, this and other suppurative infections such as lymphangitis, pneumonia and meningitis are relatively rare.
Bacteraemia
Str. pyogenes is the second most common (after Str. agalactiae) of the pyogenic streptococci isolated from blood cultures. Bacteraemia is seen regularly in patients with necrotizing fasciitis and toxic shock syndrome, but rarely as a complication to pharyngitis and local skin infections. Once in the blood, Str. pyogenes multiplies with incredible speed (doubling time 18 min), and the mortality rate approaches 40%. The potential complications include acute endocarditis leading to heart failure.
Non-suppurative sequelae
Two serious diseases may develop as sequelae to Str. pyogenes infections:
1. Rheumatic fever, a potential sequela to pharyngitis (including scarlet fever).
2. Acute glomerulonephritis, which is primarily, but not exclusively, associated with skin infections.
Both are caused by immune reactions induced by the streptococcal infection. The first clinical signs appear 1–5 weeks after the infection and at a time when the bacteria may have been eradicated by the immune system or as a result of antibiotic therapy.
Clinical correlations suggest that certain forms of psoriasis may also be triggered by Str. pyogenes throat infection. Preliminary evidence supports the hypothesis that some streptococcal superantigens cause disruption of immunological tolerance of a CD8+ T cell subset that recognizes cross-reactive epitopes on M proteins and skin keratin.
Rheumatic fever
This manifests as an inflammation of the joints (arthritis), heart (carditis), central nervous system (chorea), skin (erythema marginatum) and/or subcutaneous nodules. Polyarticular arthritis is the most common manifestation, whereas carditis is the most serious as it leads to permanent damage, particularly of the heart valves.
Rheumatic fever is a major cause of acquired heart disease in young people throughout the world. The incidence of rheumatic heart disease worldwide ranges from 0.5 to 11 per 1000 of the population. New cases are relatively rare in most of Europe, but increased incidences have been observed among the aboriginal populations of Australia and New Zealand, and in Hawaii and Sri Lanka. Outbreaks of rheumatic fever have also been seen in the USA.
The disease is autoimmune in nature and is believed to result from the production of autoreactive (and polyspecific) antibodies and T lymphocytes induced by cross-reactive components of the bacteria and host tissues. Therefore, repeated episodes of Str. pyogenes infection increase the severity of the disease. The major antigens involved are myosin, tropomyosin, laminin and keratin in the human tissues, and the group A antigen (a polymer of N-acetylglucosamine) in the Str. pyogenes cell wall in addition to epitopes on some variants of surface M proteins.
Acute post-streptococcal glomerulonephritis
The clinical manifestations include:
Unlike rheumatic disease, outbreaks of post-streptococcal acute glomerulonephritis have continued to decline in most parts of the world. Regions that still exhibit a high incidence of this disease include Africa, the Caribbean, South America, New Zealand and Kuwait.
Post-streptococcal glomerulonephritis is usually referred to as an immune complex-mediated disease. However, the exact pathogenesis is not clear. Several mechanisms have been proposed, including:
• immune complex deposition in the glomeruli
• reaction of antibodies cross-reactive with streptococcal and glomerular antigens
• alterations of glomerular tissues by streptococcal products such as streptokinase
• direct complement activation by streptococcal components that have a direct affinity for glomerular tissues.
Disease is associated with a limited number of M types of Str. pyogenes, and there is evidence that particular variants of streptokinase are crucial nephritogenic factors.
Unlike rheumatic fever, there is a general absence of individual recurrences, suggesting that antibodies to nephritogenic factors protect against disease rather than the opposite.
Streptococcus agalactiae
Str. agalactiae is equivalent to Lancefield group B streptococci. Its primary human habitat is the colon. It may be carried in the throat and, importantly, 10–40% of women intermittently carry Str. agalactiae in the vagina.
Previously, Str. agalactiae was recognized primarily as a cause of bovine mastitis (agalactia, want of milk). However, since 1960 it has become the leading cause of neonatal infections in industrialized countries and is also an important cause of morbidity among peripartum women and non-pregnant adults with chronic medical conditions. Among β-haemolytic streptococci, Str. agalactiae is the most frequent isolate from blood cultures.
Pathogenesis
Virulence factors
Str. agalactiae produces several virulence factors, including haemolysins, capsule polysaccharide, C5a peptidase (only human pathogenic strains), hyaluronidase (not all strains), and various surface proteins that bind human IgA and serve as adhesins.
Ten different types of the capsular polysaccharide have been identified (Ia, Ib and II–IX). The serotype most frequently associated with neonatal infections is type III, whereas infections in adults are more evenly distributed over the different serotypes.
Among the haemolysins produced by Str. agalactiae, one, known as the CAMP factor (so-called because it was originally described by Christie, Atkins and Munch-Petersen), plays an important role in the recognition of this species in the laboratory. The CAMP factor lyses sheep or bovine red blood cells pretreated with the β-toxin of Staph. aureus (Fig. 16.6). Purified CAMP factor protein is fatal to rabbits when injected intravenously.

Clinical features
Infection in the neonate
Two different entities are recognized:
1. Early-onset disease, most cases of which present at or within 12 h of birth.
2. Late-onset disease, presenting more than 7 days and up to 3 months after birth.
Early-onset disease
This results from ascending spread of Str. agalactiae from the vagina into the amniotic fluid, which is then aspirated by the infant and results in septicaemia in the infant or the mother, or both. Infants borne by mothers carrying Str. agalactiae may also become colonized during passage through the vagina.
Depending on the site of initial contamination, neonates may be ill at birth or develop acute and fulminating illness a few hours, or a day or two, later. The clinical symptoms include lethargy, cyanosis and apnoea; when septicaemia progresses, shock ensues and death will occur if treatment is not instituted quickly. Meningitis and pulmonary infection may be associated.
As a result of improved recognition and prompt treatment of babies with symptoms, the fatality rate has been reduced to less than 10%. However, considerable morbidity persists among some survivors, especially those with meningitis.
Risk factors for neonatal colonization and infection are:
The immune status of the mother, and hence the level of maternal IgG antibodies in the infant, appears to be more important than the degree of colonization of the mother’s genital tract by Str. agalactiae.
It is possible that Str. agalactiae itself may cause premature rupture of membranes as a result of secretion of proteases and activation of local inflammation.
Late-onset disease
Purulent meningitis is the most common manifestation, but septic arthritis, osteomyelitis, conjunctivitis, sinusitis, otitis media, endocarditis and peritonitis also occur. The incidence of invasive infection is higher among pre-term infants than among those born at term.
The pathogenesis is distinct from that of early-onset disease. There is usually no history of obstetric complications and the disease is unrelated to vaginal colonization in the mother. Many cases are acquired in hospital. Ward staff can be carriers of Str. agalactiae, and contamination of the baby may occur during nursing procedures, with subsequent baby-to-baby spread. Mastitis in the mother has also been described as a source of infection.
Infections in the adult
Ascending spread of Str. agalactiae leading to amniotic infection may result in abortion, chorio-amnionitis, post-partum sepsis (endometritis) and other infections (e.g. pneumonia) in the postpartum period in young, previously healthy women.
Str. agalactiae is also a frequent cause of infection in certain risk groups of non-pregnant adults. Disease may manifest as sepsis, pneumonia, soft tissue infections such as cellulitis and arthritis, and urinary tract infections complicated by bacteraemia. The risk factors in these patients are diabetes mellitus, liver cirrhosis, renal failure, stroke and cancer. Older age, independent of underlying medical conditions, increases the risk of invasive Str. agalactiae infection.
Other pyogenic streptococci
Str. suis serotype 2 (group R streptococci) cause septicaemia and meningitis in pigs. They belong to a phylogenetic lineage separate from the other pyogenic streptococci. They occasionally infect people in contact with contaminated pork or infected pigs, and may cause septicaemia, meningitis and respiratory tract infections. Abattoir workers, butchers and, to a lesser extent, those involved in domestic food preparation are at risk.
Streptococcus pneumoniae
Str. pneumoniae, commonly called the pneumococcus, is a member of the oropharyngeal microbiota of 5–70% of the population, with the highest isolation rate in children during the winter months. In contrast to other streptococci, Str. pneumoniae generally occurs as characteristic diplococci (see Fig. 16.1). Although closely related genetically to the commensal Str. mitis and Str. oralis, Str. pneumoniae is one of the four most frequent causes of fatal infections worldwide. It primarily causes disease of the airways and associated tissues (middle ear, paranasal sinuses, mastoids and lung parenchyma), but may spread to other sites, such as the meninges, joints, peritoneum, and endocardium.
Str. pneumoniae is genetically very flexible because of frequent recombination between individual strains. Gene transfer is by transformation and may result in the expression of a different capsular serotype. Experimental transfer of capsule genes in pneumococci was the basis of the original demonstration that DNA contains the genetic information in cells.
Pathogenesis
Virulence factors
Capsule
The capsular polysaccharide is a crucial virulence factor. The capsule is antiphagocytic, inhibiting complement deposition and phagocytosis where type-specific opsonic antibody is absent. A total of 91 different capsular serotypes have been identified.
The serotypes are designated by numbers, and those that are structurally related are grouped together (1, 2, 3, 4, 5, 6A, 6B, etc.). The different serotypes differ in virulence. Thus, about 90% of cases of bacteraemic pneumococcal pneumonia and meningitis are caused by some 23 serotypes.
IgA1 protease
Like the two other principal causes of bacterial meningitis (Neisseria meningitidis and Haemophilus influenzae), pneumococci produce an extracellular protease that specifically cleaves human IgA1 in the hinge region. This protease enables these pathogens to evade the protective functions of the principal immunoglobulin isotype of the upper respiratory tract.
Pneumolysin
Pneumococci produce an intracellular membrane-damaging toxin known as pneumolysin, which is released by autolysis. Pneumolysin inhibits:
In experimental models it induces the features of lobar pneumonia and contributes to the mortality associated with this disease.
Autolysin
When activated, the pneumococcal autolysin breaks the peptide cross-linking of the cell wall peptidoglycan, leading to lysis of the bacteria. Autolysis enables the release of pneumolysin and, in addition, large amounts of cell wall fragments. The massive inflammatory response to these peptidoglycan fragments is an important component of the pathogenesis of pneumococcal pneumonia and meningitis.
Clinical features
Predisposing factors
Most Str. pneumoniae infections are associated with various predisposing conditions. Although occasional clusterings of pneumococcal infections are recognized, person-to-person spread is uncommon.
Pneumonia results from aspiration of pneumococci contained in upper airway secretions into the lower respiratory tract; for example when the normal mechanisms of mucus entrapment and expulsion by an intact glottic reflex and mucociliary escalator are impaired. This situation may arise in:
• disturbed consciousness in association with general anaesthesia, convulsions, alcoholism, epilepsy or head trauma
• respiratory viral infections, such as influenza
• chronic bronchitis and other forms of chronic bronchial sepsis.
Other predisposing disease states in which pneumococcal pneumonia may be the terminal event include:
Immune deficiencies that predispose to pneumococcal infection include:
In these conditions there is either a relative or absolute deficiency of opsonic antibody activity or an inability to induce a sufficient type-specific antibody response. Tuftsin, a naturally occurring tetrapeptide secreted by the spleen, also plays a role in combating pneumococcal sepsis; particularly at risk are those deficient in splenic activity:
Human immunodeficiency virus (HIV) infection carries an increased risk of bacterial infections, including those caused by the pneumococcus, particularly in children.
Acute infections of the middle ear and paranasal sinuses occur in otherwise healthy children, but are usually preceded by a viral infection of the upper respiratory tract leading to local inflammation and swelling, and obstruction of the flow from these sites.
Pneumonia
Str. pneumoniae is the most frequent cause of pneumonia. The estimated annual incidence is 1–3 per 1000 of the population, with a 5% case fatality rate. Pneumococcal pneumonia follows aspiration with subsequent migration through the bronchial mucosa to involve the peribronchial lymphatics. The inflammatory reaction is focused primarily within the alveolus of a single lobule or lobe, although multilobar disease can also occur. Contiguous spread commonly results in inflammatory involvement of the pleura; this may progress to empyema.
Pericarditis is an uncommon but well recognized complication. Occasionally, lung necrosis and intrapulmonary abscess formation occur with the more virulent pneumococcal serotypes. Bacteraemia may complicate pneumococcal pneumonia in up to 15% of patients. This can result in metastatic involvement of the meninges, joints and, rarely, the endocardium.
The mortality rate from pneumococcal pneumonia in those admitted to hospital in the UK is approximately 15%. It is increased by age, underlying disease, bloodstream involvement, metastatic infection and certain types of pneumococci with large capsules (e.g. serotype 3).
Otitis media and sinusitis
Middle ear infections (otitis media) affect approximately half of all children between the ages of 6 months and 3 years; approximately one-third of cases are caused by Str. pneumoniae. Disease occurs after acquisition of a new strain to which there is no pre-existing immunity. The prevalence is highest among children attending kinder garten or primary school, where there is a constant exchange of pneumococcal strains.
Meningitis
Str. pneumoniae is among the three leading causes of bacterial meningitis. It is assumed that invasion arises from the pharynx to the meninges via the bloodstream, as bacteraemia usually coexists. Meningitis may occasionally complicate pneumococcal infection at other sites, such as the lung and middle ear.
The incidence of pneumococcal meningitis is bimodal and affects children less than 3 years of age and adults of 45 years and above. The fatality rates are 20% and 30%, respectively, considerably higher than those associated with other types of bacterial meningitis.
Commensal streptococci
Viridans streptococci
The viridans streptococci, and in particular the species of the mitis and salivarius groups, are dominant members of the resident microbiota of the oral cavity and pharynx in all age groups. They play an important role by inhibiting the colonization of many pathogens, including pyogenic streptococci. This is achieved by two different mechanisms:
1. production of bacteriocins (see p. 35)
2. production of hydrogen peroxide (also responsible for α-haemolysis).
Most strains secrete bacteriocins. Experimental implantation of strains of Str. salivarius with strong bacteriocin activity prevents colonization with Str. pyogenes in humans.
Mitis group
Str. mitis, Str. oralis and Str. sanguinis, among other viridans streptococci, colonize tooth surfaces as well as mucosal membranes. Because of their presence in the bacterial biofilms (dental plaque) on tooth surfaces, these species may enter the bloodstream during dental procedures such as tooth extraction or vigorous tooth cleaning, particularly when the gingival tissue is inflamed.
In healthy individuals, bacteria of such low virulence are cleared from the circulation within 1 h. However, in patients with various predisposing conditions (Table 16.2), in particular heart valve damage due to post-streptococcal rheumatic fever, the circulating streptococci may settle in a niche protected from phagocytic cells. Local growth on the surface of heart valves eventually causes scarring and functional deficiency. As the disease progresses over several months, it is referred to as subacute bacterial endocarditis. It is usually accompanied by intermittent fever. Disruption of bacteria from the cardiac vegetations may cause embolic abscesses in various organs, including the brain. Until endocarditis due to skin staphylococci became more prevalent as a result of intravenous drug abuse, viridans streptococci were the most frequent causes of infective endocarditis.
Table 16.2 Factors predisposing to infective endocarditis
| Cardiac factors | Non-cardiac factors |
|---|---|
a Procedures in which bleeding occurs.
Str. mitis and Str. oralis are increasingly recognized as causes of often fatal septicaemias in immunocompromised patients.
Mutans group
Str. mutans and Str. sobrinus exclusively colonize tooth enamel and do not occur until tooth eruption. Their proportions in the biofilm forming on tooth surfaces (dental plaque) are closely related to sugar consumption, and they are a major cause of dental caries because of their ability to produce large amounts of lactic acid even at pH values below 5.0. Like most other plaque streptococci they may cause subacute bacterial endocarditis.
Enterococcus species
As indicated by the name, members of the genus Enterococcus have their natural habitat in the human intestines. The species most commonly associated with human disease are E. faecalis and E. faecium. The diseases with which they are associated are:
E. faecalis and E. faecium are important causes of wound and urinary tract infection in hospital patients and may cause sporadic outbreaks. Bacteraemia carries a poor prognosis as it often occurs in patients with major underlying pathology and in those who are immunocompromised.
Laboratory diagnosis
Collection of specimens
The diagnosis of streptococcal infections is established by demonstrating the presence of the pathogen in throat or skin swabs, pus, blood cultures, cerebrospinal fluid (CSF), expectorates or urine according to the site of infection. In pneumococcal meningitis the CSF is often macroscopically cloudy. The cell count is usually increased markedly and shows a predominance of polymorphonuclear leucocytes. Typical Gram-positive diplococci can commonly be demonstrated, sometimes in enormous numbers, by Gram-stain examination of a CSF deposit. The appearance is often typical, and a presumptive diagnosis can be made to allow appropriate therapy to be started before the identity of the organism is confirmed by culture.
Blood cultures are of value in patients with invasive streptococcal infections. This is also the case in patients with suspected pneumococcal pneumonia, particularly when this is severe, as up to 15% of patients are bacteraemic. Detection of bacteria by direct plating or microscopy of blood is not feasible owing to their low density.
Other body sites that may merit investigation according to the clinical presentation include joint and peritoneal fluids. Tympanocentesis provides the possibility of establishing the microbial cause of otitis media, but as most of these infections settle spontaneously, or with the assistance of a few days’ antibiotic treatment, tympanocentesis is not usually necessary.
Cultivation and identification
Unlike staphylococci, streptococci lack the enzyme catalase, which releases oxygen from hydrogen peroxide. Catalase-negative Gram-positive cocci are therefore likely to be streptococci. The appearance of cocci in obvious chains (see Fig. 16.1) is another useful criterion, but the length of the chains varies with the species and conditions of growth. Optimal chain formation is seen in broth cultures. There may be marked variation in size and shape, particularly in older cultures or in direct smears from purulent exudates.
The primary cultivation medium for streptococci is blood agar, supplemented, whenever enterococci are suspected, with an agar medium (e.g. MacConkey’s medium) selective for enterobacteria. Streptococci of aetiological significance usually predominate in the culture even when the sample is taken from a site with a resident microbiota.
The pyogenic streptococci are detected initially by their β-haemolytic activity. The colonies are about 1 mm in diameter and, in contrast to those of staphylococci, lack pigment. Colonies of pneumococci are α-haemolytic, smooth, and may vary in size according to the amount of capsular polysaccharide produced; those of serotypes 3 and 37 are usually larger than the rest and have a watery or mucoid appearance. During prolonged incubation, autolysis of bacteria within the flat pneumococcal colonies results in a typical subsidence of the centre (‘draughtsman colonies’).
Species identification of pyogenic streptococci is based largely on serological detection of group antigens by immune precipitation or co-agglutination techniques. An additional test that is helpful in the presumptive identification of Str. pyogenes is the bacitracin sensitivity test. In contrast to most other streptococci, Str. pyogenes is uniformly sensitive and large inhibition zones are formed round bacitracin discs on blood agar. Likewise, Str. agalactiae can be identified presumptively by the CAMP reaction (see Fig. 16.6).
Pneumococci are distinguished from other α-haemolytic streptococci by their characteristic sensitivity to optochin (ethylhydrocupreine). Growth of pneumococci is inhibited around an optochin disc applied to an inoculated blood agar plate. With few exceptions, other α-haemolytic streptococci are not inhibited. In doubtful cases the identity of pneumococci is confirmed by demonstrating bile solubility, autolysis or reactivity to a poly-specific antiserum (‘omniserum’) against capsular polysaccharides.
Other streptococci are identified by biochemical characteristics, such as the ability to ferment various carbohydrates and hydrolyse amino acids. However, some of the viridans streptococci are notoriously difficult to identify.
Enterococci are unique among the streptococci in their ability to grow on bile-containing media.
Antigen detection
Numerous commercial kits are available for the detection of Str. pyogenes directly in throat swabs without cultivation. These diagnostic kits use specific antibodies to detect the group A antigen in the material on the swab. They allow practitioners to test whether a throat infection is caused by Str. pyogenes, but the sensitivity and specificity of individual tests vary.
Antibody detection
Detection of antibodies against antigens of Str. pyogenes is an important means of establishing the diagnosis of poststreptococcal rheumatic fever and glomerulonephritis. In many cases the initiating infection in the throat or on the skin is no longer present.
Immune responses vary depending on whether the original focus is the throat or skin. Antibodies against streptolysin O (the ASO test) are used to document antecedent streptococcal infection in the throat of patients with clinical signs of rheumatic fever. A significant increase in antibody titre appears 3–4 weeks after initial exposure to the micro-organism. Detection of increased levels of serum antibodies to streptococcal hyaluronidase and DNAase B is also of diagnostic importance.
ASO estimation is unreliable in pyoderma-associated acute glomerulonephritis. A raised ASO titre is not observed in these patients, perhaps because lipids present in the skin inactivate the streptolysin O. Detection of antibodies against streptococcal DNAase B is recommended as a diagnostic tool in these patients.
Typing of streptococci
Strains of Str. pyogenes can be subdivided into serological types. The most comprehensive typing scheme is based on structural differences in the highly variable surface M protein. More than 150 different M types may be distinguished with type-specific antisera or by sequence differences in the gene (emm) encoding the M protein. An alternative typing scheme is based on the surface protein known as the T antigen.
Apart from serving epidemiological purposes, particular M types are associated with particular types of infection. Thus, certain M types are more commonly associated with skin infections than mucosal infections. Recent increases in the rate and severity of invasive Str. pyogenes infections (toxic shock syndrome and necrotizing fasciitis) have been associated primarily with serotypes M-1 and M-3. Rheumatic fever is often, but not exclusively, associated with M serotypes 1, 3, 5, 6 and 18.
Pneumococci are typed on the basis of the differences in capsular polysaccharides, of which 91 have been described. The addition of India ink to a suspension of pneumococci shows the presence of the capsule as a clear halo around the organisms. Mixing a suspension of pneumococci with type-specific antisera increases the visibility of the capsule in the microscope, and is the basis of the quellung reaction or capsular swelling test. Serotyping of pneumococci is carried out mainly in reference laboratories.
Multilocus sequence typing (MLST) schemes based on allelic sequence profiles of seven housekeeping genes are available for several Streptococcus species and allow mapping of the global distribution of sequence types.
Treatment
Penicillin resistance has never been detected in Str. pyogenes. As a result, benzylpenicillin (penicillin G) or oral phenoxymethylpenicillin (penicillin V) are the drugs of choice for treatment of infections with Str. pyogenes. Antibiotic sensitivity tests are currently unnecessary when that species is identified as the infecting organism. In cases of hypersensitivity to penicillin, erythromycin is usually the second choice, but resistance occurs and is common in some countries.
Treatment for 3–5 days limits the effect of severe attacks of streptococcal infection and prevents suppurative complications such as otitis media, although the streptococci are eliminated from the infected area only if treatment is continued for 10 days.
Surgery is essential to remove damaged tissue in case of necrotizing fasciitis, as antibiotic penetration of the infected area is poor. Clindamycin is preferred to penicillin because it inhibits protein synthesis, including production of exotoxin.
Most strains of Str. agalactiae are susceptible to penicillins, macrolides and glycopeptides.
Although streptococci are intrinsically resistant to aminoglycosides, these agents interact synergically with penicillins and the combination is often used in the treatment of streptococcal and enterococcal endocarditis.
Pneumococci and viridans streptococci are often resistant to penicillins owing to mutations in the target penicillin-binding proteins. These mutations have accumulated in strains of Str. mitis and Str. oralis, and the altered genes have subsequently been transferred by genetic transformation to Str. pneumoniae. High-level penicillin resistance (minimum inhibitory concentration above 2 mg/L) was first recognized in 1977 in South Africa, where it was responsible for an epidemic of pneumococcal meningitis unresponsive to penicillin. The incidence of penicillin resistance is quite variable geographically and reflects the local level of antibiotic usage.
Most pneumococcal infections with strains exhibiting intermediate-level resistance to penicillin (minimum inhibitory concentration 0.1–1 mg/L) respond to highdose therapy; an exception is meningitis, because of problems of penetration into the CSF. Penicillin resistance in pneumococci and other viridans streptococci is often linked to resistance to several other antibiotics. Resistance to erythromycin, tetracycline and chloramphenicol is not uncommon, and tolerance even to vancomycin has been reported.
The dose of penicillin necessary to treat susceptible pneumococcal infection is determined largely by pharmacological factors at the site of infection. For example, pneumococcal pneumonia responds to doses of penicillin as low as 0.3 g (0.5 mega-units) twice daily, whereas pneumococcal meningitis requires much higher doses. In patients unable to tolerate penicillin, erythromycin is the most widely used alternative agent for respiratory pneumococcal infections.
Unlike other streptococci, enterococci are intrinsically resistant to cephalosporins. Sensitivity to penicillins and other antibiotics varies widely, and clinical isolates must be tested for their susceptibility. Vancomycin resistance has been observed in enterococci and is a problem in high-dependency areas of some hospitals.
Prevention and control
Hygienic measures
Skin infections with Str. pyogenes are usually associated with poor hygiene, and can to a large extent be prevented by standard hygienic measures. Late-onset neonatal infections with Str. agalactiae may also be prevented or significantly reduced by standard aseptic nursing procedures.
Likewise, hygiene is the most important preventive measure in relation to dental caries, which can be prevented largely by regular tooth-brushing with a fluoride-containing dentifrice combined with restricted consumption of fermentable carbohydrates, in particular sucrose.
Chemoprophylaxis
Prophylactic use of antibiotics is relevant in some streptococcal infections. As the primary attack of rheumatic fever usually occurs during childhood, long-term penicillin prophylaxis until adulthood is recommended to reduce the risk of further attacks and further heart injury. This is not the case in patients with acute glomerulonephritis, because of the lack of recurrences.
Two different approaches are used to prevent early-onset neonatal Str. agalactiae infections:
1. A risk-based strategy in which women of unknown colonization status receive intrapartum antibiotic prophylaxis in case of: threatened delivery at less than 37 weeks’ gestation; premature rupture of the membranes; intrapartum fever; or previous delivery of a child who developed neonatal infection
2. A screening-based approach in which all pregnant women at 35–37 weeks’ gestation are screened for Str. agalactiae colonization in vaginal and rectal specimens. All identified carriers are offered intrapartum chemoprophylaxis. This screening-based prophylaxis is used in the USA and in many European countries and has led to a significant decline in the incidence of early onset neonatal infections due to Str. agalactiae. Intravenous or intramuscular penicillin is the agent of choice because its antimicrobial spectrum, narrower than that of ampicillin, reduces the likelihood of resistance developing in other bacteria and selection of other potential pathogens. Antimicrobial susceptibility testing of GBS isolates is crucial for appropriate antibiotic prophylaxis selection for penicillin-allergic women because resistance to clindamycin, the most common agent used in this population, is increasing among GBS isolates.
Patients at risk of developing infective endocarditis (see Table 16.2) should be given prophylactic antibiotics in association with dental procedures that lead to bleeding. The current international recommendations are amoxicillin 1 h before dental treatment or, in case of penicillin allergy, clindamycin. If the patient has been on long-term penicillin prophylaxis, the oral streptococci are likely to have reduced susceptibility to penicillins, and clindamycin or vancomycin is recommended as the alternative. It is imperative that patients at risk maintain healthy periodontal conditions and that the amount of dental plaque is kept to a minimum.
Vaccines
Pyogenic streptococci
Attempts to develop a vaccine against Str. pyogenes infections have been hampered by two problems:
1. The considerable antigenic diversity of the M protein and other vaccine candidate antigens.
2. The potential immunological cross-reactivity of many of the antigens with host tissue components.
Several strategies are currently being tested, including oral vaccination.
A vaccine against neonatal Str. agalactiae infection based on protein-conjugated type III capsular polysaccharide is being tested for use primarily in women of reproductive age. However, additional serotypes are increasingly prevalent.
Pneumococci
Before the widespread availability of effective antimicrobial drugs the treatment of pneumococcal infections was based on the use of type-specific antiserum. This reduced the mortality rate associated with bacteraemic pneumococcal pneumonia, but not to the same extent that penicillin was subsequently shown to achieve. However, it indicated that type-specific antibody had a role in the control of pneumococcal disease and led to a variety of prototype vaccines. The vaccine that has been in use for many years contains a mixture of 23 polysaccharide serotypes chosen according to the prevalence of serotypes responsible for bacteraemic pneumococcal infection. It offers protection against 90% of isolates.
Like other vaccines based on pure polysaccharides, the immunogenicity of the multivalent vaccine is inadequate in those below 2 years of age and in those immunosuppressed as a result of malignancy, steroid therapy or other chronic disease. To overcome this problem, pneumococcal vaccines containing capsular polysaccharide coupled to a carrier protein are now available. These vaccines increase the immunogenicity of the polysaccharide by rendering the response dependent on T lymphocyte help. The current conjugate vaccines include 11–13 of the capsular polysaccharides, and are now part of the childhood vaccination programme in many countries.
Immunization is recommended for various groups at risk of pneumococcal disease, particularly those with congenital or surgical asplenia and those with hereditary haemoglobinopathies such as sickle cell disease, as pneumococcal infection can be fulminant in these patients. Vaccine efficacy is not complete and many clinicians also prescribe oral phenoxymethylpenicillin as long-term chemoprophylaxis in this high-risk group. In some countries, including the USA and the UK, the vaccine is recommended for those over 65 years of age, with or without previous ill health, although there have been difficulties in establishing scientifically the efficacy in this group.
Carapetis JR, Steer AC, Mulholland EK, Weber M. The global burden of group A streptococcal diseases. Lancet Infectious Diseases. 2005;5:685–694.
Cunningham MW. Pathogenesis of group A streptococcal infections. Clinical Microbiology Reviews. 2000;13:470–511.
Henriques-Normark B, Blomberg C, Dagerhamn J, et al. The rise and fall of bacterial clones: Streptococcus pneumoniae. Nature Reviews Microbiology. 2008;6:827–837.
Kilian M. Streptococcus and Lactobacillus, ed 9. Borriello P, Murray PR, Funke G, eds. Topley and Wilson’s Microbiology and Microbial Infections, Vol. 2. London: Arnold. 2005:833–881.
Llewelyn M, Cohen J. Superantigens: microbial agents that corrupt immunity. The Lancet Infectious Diseases. 2002;2:156–162.
Marsh PD, Martin MW. Oral Microbiology, ed 5. Oxford: Elsevier; 2009.
Mitchell TJ. The pathogenesis of streptococcal infections: from tooth decay to meningitis. Nature Reviews. 2003;1:219–230.
Reinert RR, Reinert S, van der Linden M, et al. Antimicrobial susceptibility of Streptococcus pneumoniae in eight European countries from 2001 to 2003. Antimicrobial Agents and Chemotherapy. 2005;49:2903–2913.
Verani JR, McGee L, Schrag SJ. Prevention of perinatal group B streptococcal disease. Revised guidelines from CDC. Morbidity and Mortality Weekly Report. 2010;59:1–32.
Centers for Disease Control and Prevention. Disease listing. http://www.cdc.gov/ncidod/dbmd/diseaseinfo/.
Multi Locus Sequence Typing. Schemes for streptococci and several other pathogenic bacteria. http://www.mlst.net/.