Chapter 7 The Concept of a Reflex
The reflex arc, the neural substrate of a reflex, is fundamental to the physiology of posture and locomotion, as well as to the clinical examination of the nervous system. A reflex can be defined as an involuntary, qualitatively unvarying response of the nervous system to a stimulus. Reflexes are the simplest behavioral example of the general function of the nervous system: collection of sensory input, integration, and motor output. Reflexes are often both critical for survival and components of more complex behaviors. The anatomy and function of a reflex arc are often fully developed at birth.
A Reflex Arc Contains Five Fundamental Components
All reflex arcs contain five basic components (Figure 7-1). If any one of these five components malfunctions, the reflex response is altered.
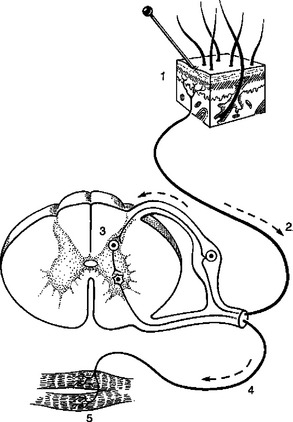
FIGURE 7-1 A reflex arc contains five fundamental components: 1, a receptor; 2, a sensory neuron; 3, one or more synapses in the central nervous system; 4, a motor neuron; and 5, a target organ, usually a muscle.
(From De Lahunta A, editor: Veterinary neuroanatomy and clinical neurology, Philadelphia, 1983, Saunders.)
1. All reflex arcs begin with a sensory receptor. Sensory receptors vary widely within the body, but all share a common function: they transduce a range of environmental energy, or the presence of an environmental chemical, into a cellular response that directly or indirectly produces action potentials along a sensory neuron. In other words, these receptors collect environmental signals and turn them into a format that can be understood by the nervous system. For example, receptors of the retina transduce light; those in the skin transduce heat, cold, pressure, and other cutaneous stimuli; muscle spindle receptors transduce stretch; and taste receptors transduce chemical stimuli from ingested material. A primary receptor is a neuron with a specialized region for stimulus transduction (e.g., retinal photoreceptors; see Figure 14-6). A secondary receptor is a nonneural cell specialized for stimulus transduction that in turn affects neural activity by releasing neurotransmitter onto a neuron (e.g., vestibular hair cells; see Figure 11-2). Action potentials resulting from stimulus transduction are generated along sensory neurons at a frequency proportional to the intensity of the transduced stimulus. This proportionality between the intensity with which the receptor is stimulated and the frequency of the resulting sensory neuron action potentials is called frequency coding; it is one major way the receptor communicates to the central nervous system (CNS) the intensity of light, heat, stretch, and so forth, that it has transduced. Stronger stimuli will also activate a larger number of sensory receptors, known as the population code of stimulus intensity.
2. The next component in a reflex arc, alluded to earlier, is a sensory neuron (CNS afferent). These neurons carry action potentials, resulting from receptor activation, to the CNS. Again, in some cases the receptor is just a specialized, usually peripheral, region of the sensory neuron (primary receptors). In other cases the receptor is physically separate from and synapses on the sensory neuron (secondary receptors). Sensory neurons enter the spinal cord by way of the dorsal roots or enter the brain through cranial nerves.
3. The third component of a reflex arc is a synapse in the CNS. Actually, for most reflex arcs, more than one synapse occurs in series (polysynaptic). However, some reflex arcs that originate from the muscle spindle are monosynaptic. In polysynaptic reflexes, where one or more neurons lie between the sensory neuron input to the CNS and the motor neuron output, these interposed neurons are called interneurons and can also be considered part of this third component of the reflex arc.
4. The fourth component is a motor neuron (CNS efferent), which carries action potentials from the CNS to the synapse with the target (effector) organ. Motor neurons leave the spinal cord through the ventral roots, and motor neurons leave the brain through the cranial nerves.
5. The last component is some target organ (effector organ) that causes the reflex response. This is usually a muscle, such as the skeletal muscle fibers of the quadriceps muscle of the leg, in the case of the “knee jerk” (muscle stretch) reflex, or the smooth muscle of the iris in the pupillary light reflex.
In reality, the final reflex response to a stimulus in mammals is rarely, if ever, the product of a monosynaptic reflex arc acting in isolation. Even if a sensory neuron participates in a monosynaptic reflex arc, it will often give off branches in the CNS that participate in polysynaptic reflex circuits. In addition, even the simplest mammalian reflex responses often involve both the excitation of a given muscle or muscles and the inhibition of another (usually antagonistic) muscle or muscles. The knee jerk reflex is a good illustration of these points (see Figure 8-3). With regard to the individual sensory neurons that underlie this reflex, some terminal branches make excitatory monosynaptic connections with motor neurons that activate the quadriceps muscle. Other terminal branches of that same sensory neuron participate in a disynaptic circuit that inhibits motor neurons innervating the antagonistic hamstring muscle.
Also, reflexes do not operate in isolation from the rest of the nervous system. Inputs from other regions of the nervous system, onto the CNS components of a reflex arc, can modulate the sensitivity of the reflex response. For example, if input from the brain to the spinal cord is damaged, reflexes in that region of the cord become exaggerated.
Reflex Arcs Can Be Segmental or Intersegmental
A segmental reflex is a reflex in which the reflex arc passes through only a small rostrocaudal portion of the CNS (Figure 7-2, A). In such cases the sensory neuron entrance to the CNS, the CNS circuitry, and the motor neuron exit all have a similar rostrocaudal location. The quadriceps stretch reflex (knee jerk reflex) and the pupillary light reflex are examples of segmental reflexes because they use only, respectively, a small number of spinal cord segments (e.g., L4-L6) or a small rostrocaudal region of the brainstem.
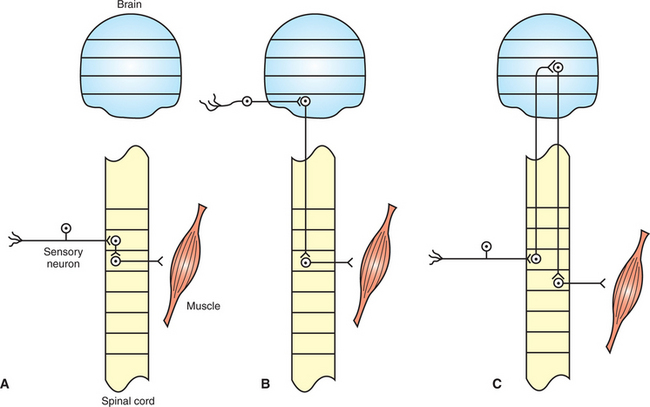
FIGURE 7-2 Segmental and intersegmental reflexes. A, In the segmental reflex the sensory neuron input, central nervous system (CNS) circuitry, and motor neuron output traverse only a small number of rostrocaudal segments of the CNS. B, Intersegmental reflex arcs traverse several CNS segments. In some intersegmental reflexes the sensory neuron input and motor neuron output are separated by several segments. C, The long-loop intersegmental reflex arc traverses several CNS segments, even though the sensory input and motor output are located in close rostrocaudal proximity. Horizontal lines delimit either spinal cord segments (e.g., L1, L2) or major brain divisions (e.g., medulla, pons).
In an intersegmental reflex the reflex arc traverses many segments of the spinal cord or several major brain divisions (e.g., medulla to midbrain). In one class of intersegmental reflex, the motor neuron exit is located, or extends, a considerable rostral or caudal distance from the location of the sensory neuron entrance to the CNS (Figure 7-2, B). Examples include the vestibulospinal reflexes that produce postural adjustments in response to acceleration or tilt of the head. The sensory input, originating in the vestibular apparatus of the inner ear, enters the CNS near the pontomedullary border of the brainstem, and the motor neurons exit from the spinal cord, over a large number of spinal cord segments.
For some intersegmental reflexes, the sensory neuron entrance and motor neuron exit are at a similar rostrocaudal location, but the CNS circuitry lying in between travels to and returns from a distant region of the CNS (Figure 7-2, C). These are sometimes referred to as long-loop intersegmental reflexes. The proprioceptive positioning reaction is often associated with this category. This reaction involves the animal promptly returning its paw to the normal pad-down position after the clinician flexes it to make the dorsal surface touch the floor or tabletop. The sensorimotor circuitry of this reaction courses from the limb’s peripheral nerves and associated spinal cord segments all the way through the cerebral cortex and back to the limb’s spinal cord segments and peripheral nerves.
Reflex Arcs Are Widespread in the Nervous System, and Reflexes Underlie a Major Portion of the Neurological Examination
Reflex arcs are ubiquitous in the nervous system and are the basis of much of an animal’s subconscious, involuntary response to its environment. Much of a veterinarian’s clinical examination of the nervous system involves evoking reflex responses, such as the pupillary light reflex, muscle stretch (knee jerk) reflex, and flexor reflex.
If any of the five components of the reflex arc malfunctions, the expected reflex response is altered or does not occur. It is important to know the general anatomy, physiology, and expected normal clinical response of the common reflexes in order to perform a neurological examination so that lesions can be localized. For example, loss of pupillary constriction to light in conjunction with normal responses to other visual stimuli, such as avoiding a rapidly oncoming object, suggests that the problem is probably not with the sensory receptor or sensory neuron components of the reflex arc. Several such reflexes are discussed in detail in subsequent chapters.
CLINICAL CORRELATIONS
Trauma in a Foal
History.
Distraught owners call you about their 4-month-old Tennessee Walking Horse colt. He appeared normal this morning when they let him out to pasture with his mother, but later this afternoon, the mare and the foal did not come in to be fed. The owners went out to the pasture and found the mare with the foal, who would not get up. He was lying on his side and seemed unable to position himself sternal. When the owners tried to reposition him, the foal thrashed, trying to get away. You tell the owners not to move the foal and that you will be there soon.
Clinical Examination.
The foal appears to be responsive but in great pain and unable to rise. Temperature, pulse, and respirations are all mildly increased. There appears to be a swelling along the cervical (neck) area in the region of C1-C3. The swelling is hard (bone) and has some fluid (inflammation) as well. There is some crepitus in the area of the swelling (possible fracture site). The foal displays no other areas of swelling or trauma. Neurological examination reveals normal cranial nerves. In the front limbs the biceps and triceps reflexes seem increased on both sides. Deep pain is present, and cutaneous sensation is increased bilaterally. In the hind limbs the femoral, sciatic, and tibial responses are increased.
Comment.
Although it is difficult to localize a fracture definitively, based on history and physical examination a fracture seems likely. The fracture appears to be in the region of C1-C3. Radiographs would be ideal to make a definitive diagnosis. On neurological testing of the biceps, triceps, sciatic, femoral, and cranial tibial responses, all assess segmental reflex arcs. Because of a high cervical fracture, the descending motor tracts that supply both the thoracic and the pelvic limbs are affected. From point of initiation, the segmental reflexes tested involve the muscle spindles, which detect stretch, followed by sensory fibers in the peripheral nerves, the dorsal root and its ganglion, and the central branches of the sensory fibers projecting onto the ventral horn cell of the same spinal segment. The efferent pathway involves the ventral horn cell (lower motor neuron), followed by the ventral root, motor fibers in the peripheral nerve, neuromuscular junction, and then the myofibers being tested. The presence but exaggeration of the segmental reflexes reflects a change in the modulation of these reflexes by the damaged descending motor tracts that pass through the area of the lesion.
Treatment.
The prognosis for this foal is poor. Based on the physical examination and clinical signs, a fracture is likely, and there is little hope for recovery. The complications associated with trying to manage a foal as the fracture heals are enormous. The fracture may not heal, and the foal could have severe residual neurological deficits. In most cases, these foals are euthanized fairly quickly because of the poor prognosis.
Bailey JG. Muscle physiology. Reece WO, ed. Duke’s physiology of domestic animals, ed 12, Ithaca, NY: Comstock Cornell University Press, 2004.
Binder MD. Peripheral motor control: spinal reflex actions of muscle, joint and cutaneous receptors. Patton HD, Fuchs AF, Hille B, et al. Textbook of physiology, ed 21, Philadelphia: Saunders, 1989.
Boron WF, Boulpaep EL. Medical physiology: a cellular and molecular approach, updated edition. Philadelphia: Saunders, 2005.
Divers TJ, Smith MO. Spinal fractures and luxations and spinal cord trauma. Smith B, ed. Large animal internal medicine, ed 3, St Louis: Mosby, 2002.
Guyton AC, Hall JE. Textbook of medical physiology, ed 11. Philadelphia: Saunders, 2006.
Kandel ER, Schwartz JH, Jessell TM. Principles of neural science, ed 4, New York: McGraw-Hill, 2000.
Lorenz MD, Kornegay J. Oliver and Lorenz’s handbook of veterinary neurology, ed 4. Philadelphia: Saunders, 2004.
PRACTICE QUESTIONS
1. Which of the following is not always a component of a reflex arc?
2. Which of the following regarding sensory receptors is false?
3. When the intensity with which a receptor is stimulated is increased, what happens to the frequency of action potentials along the sensory neuron from that receptor?
4. Which of the following is not an example of a segmental reflex?