Chapter 8 Skeletal Muscle Receptor Organs
1. The muscle spindle stretch receptor is an encapsulated organ of specialized muscle fibers with separate motor and sensory innervations.
2. The muscle spindle conveys information about muscle length to the central nervous system.
3. Muscle stretch and action potentials along spindle sensory neurons lead to reflex contraction of the extrafusal muscle fibers.
4. The central nervous system can control spindle sensitivity directly through the gamma (γ) motor neurons.
5. The Golgi tendon organ lies in series between muscle and tendon and detects muscle tension.
Movement, characteristic of all animals, is the end product of skeletal muscle contraction. It is orchestrated by the central nervous system (CNS) through its control of the motor unit (see Chapter 6), which has been referred to as movement’s “final common path.” To control body movement appropriately, the CNS must (1) assess the effect of gravity on the many muscles of the body, (2) determine the initial position of the body parts to be moved, and (3) detect any discrepancy between the intended movement and the movement that actually occurs. Once such discrepancies are detected, appropriate adjustments can be made.
Two important receptor systems have evolved in the skeletal muscles of mammals to provide the CNS with the aforementioned information: the muscle spindle and the Golgi tendon organ (Figure 8-1). The muscle spindles, arranged in parallel to the contracting skeletal muscle fibers, provide information about muscle length. The Golgi tendon organ, arranged in series with the contracting skeletal muscle fibers, detects muscle tension. This chapter discusses the anatomy and physiology of these two receptor organs; Chapter 10 discusses how the CNS uses the information gathered from these organs to coordinate posture and locomotion. Some of this information is used in reflex arcs of the type described in Chapter 7.
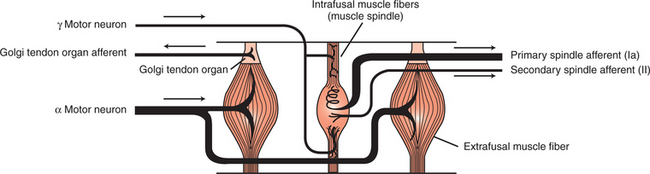
FIGURE 8-1 Skeletal muscles have two important receptors: the muscle spindle and the Golgi tendon organ. The intrafusal muscle fibers (muscle spindle) are arranged in parallel with the extrafusal muscle fibers; the Golgi tendon organ is in series with the extrafusal fibers. Arrows indicate direction of action potential flow along respective axons.
(Modified from Kandel ER, Schwartz JH: Principles of neural science, ed 2, New York, 1985, Elsevier Science Publishing.)
The Muscle Spindle Stretch Receptor Is an Encapsulated Organ of Specialized Muscle Fibers with Separate Motor and Sensory Innervations
The muscle spindle is an encapsulated group of about 3 to 12 small, slender, specialized skeletal muscle fibers (Figure 8-2). Because their capsule is spindle shaped, or fusiform, these muscle fibers are called intrafusal muscle fibers. The muscle fibers that cause physical shortening of the muscle (the majority of muscle fibers in a muscle belly), located outside of the capsule, are called extrafusal muscle fibers. Extrafusal muscle fibers often span the length of the gross muscle from origin to insertion tendon; intrafusal muscle fibers and their capsules are much shorter (about 4-10 mm long). In addition, the intrafusal muscle fiber endings are attached to the extracellular matrix of, and lie in parallel to, the extrafusal muscle fibers. Therefore, if the muscle is stretched, lengthening the extrafusal muscle fibers, the intrafusal fibers of the muscle spindle are also stretched.
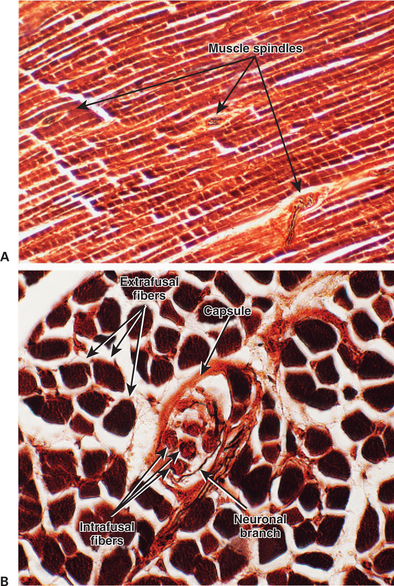
FIGURE 8-2 The muscle spindle receptor is an encapsulated group of specialized (intrafusal) skeletal muscle fibers supplied with both motor and sensory innervation. A, longitudinal section through a skeletal muscle showing that the encapsulated muscle spindles are oriented parallel to the more numerous extrafusal fibers of the muscle. The ends of the muscle spindle are attached to the extracellular matrix of the extrafusal fibers. B, Higher-magnification view of a transverse section through a muscle spindle. Intrafusal fibers can be seen within the spindle’s tissue capsule. These fibers are fewer, shorter, and more slender than the surrounding extrafusal fibers. A portion of the spindle’s innervation can also be seen.
(Images courtesy Dr. Tom Caceci, Department of Biomedical Sciences and Pathobiology, College of Veterinary Medicine, Virginia Tech.)
Unlike extrafusal muscle fibers, the contractile elements of intrafusal muscle fibers are restricted to their polar ends, with none in their middle (equatorial) region. Therefore, their polar ends can contract, but their equatorial region cannot. Such contraction does not directly contribute to the shortening of the gross muscle, but it can tighten the region of the intrafusal fiber that lies between the two poles. As discussed later, this can have a dramatic effect on the muscle spindle’s sensitivity for transducing muscle stretch.
Spindle sensory neurons arise from the equatorial region of the intrafusal muscle fibers and carry action potentials from the spindle to the CNS by way of the peripheral nerves. These CNS afferents enter the spinal cord through the dorsal roots (Figure 8-3). The contractile, polar regions of the intrafusal muscle fibers are innervated by motor neurons called gamma (γ) motor neurons. Extrafusal muscle fibers—the muscle fibers that cause the physical shortening of the muscle—are supplied by a different population of motor neurons (those that comprise the motor units) called alpha (α) motor neurons. Although γ motor neurons go to intrafusal muscle fibers and α motor neurons go to the extrafusal muscle fibers, these CNS efferents both have their cell bodies in the ventral horn of the spinal cord, and their axons leave through the ventral roots.
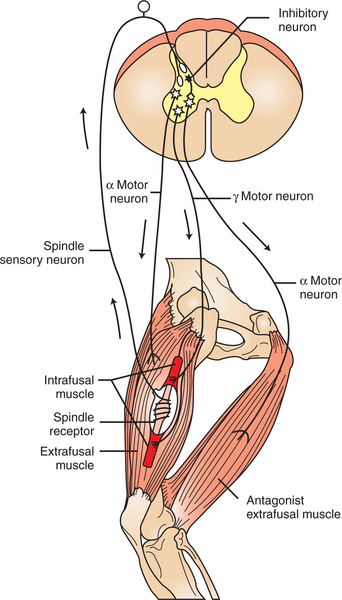
FIGURE 8-3 The muscle spindle stretch reflex (illustrated here as the “knee jerk reflex”) begins when the spindle receptor organ is stretched. This causes action potentials on the receptor’s sensory neurons, which in turn cause excitatory postsynaptic potentials on the α motor neurons returning to the extrafusal muscle fibers of that same muscle. Action potentials on the α motor neurons cause extrafusal muscle fibers to contract, and the knee extends (“jerks”). Through an inhibitory interneuron, the α motor neurons to the antagonist muscles are simultaneously inhibited.
The Muscle Spindle Conveys Information About Muscle Length to the Central Nervous System
As noted, stretching a muscle can stretch the intrafusal fibers of the muscle spindle that lie in parallel to the extrafusal muscle fibers. Stretching (lengthening) the equatorial segment of the intrafusal muscle fiber generates action potentials along the spindle sensory neurons. As the equatorial segment is lengthened, it is believed that stretch-sensitive ion channels open on the sensory neurons, leading to membrane depolarization and action potential generation. Action potentials are generated along the sensory neuron output of the muscle spindle in proportion to the amount of lengthening of the middle of the intrafusal muscle fibers. There are actually subclasses of intrafusal fibers (dynamic nuclear bag, static nuclear bag, nuclear chain) and spindle sensory neurons (Ia, II). This variety allows the sensory neuron output of the spindle to detect not only a change in length during the dynamic phase of muscle lengthening, but also the rate of lengthening, as well as the steady-state length of the muscle as the animal holds the joint still (Figure 8-4).
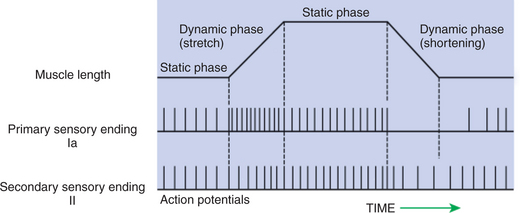
FIGURE 8-4 The muscle spindle can signal the steady-state length of the muscle, as well as the onset and velocity of stretch. When the muscle is stretched, both type Ia and type II spindle sensory neurons have a higher action-potential firing rate at the muscle’s new static length. During the dynamic stretching phase, the action-potential firing rate of the type Ia spindle sensory neuron increases rapidly and in proportion to the velocity of stretch. Spindle sensory neurons can also register a decreased steady-state length of the muscle, but type Ia and type II neurons display differential sensitivity to the dynamic phase of shortening
(From Brodal P: The central nervous system: structure and function, ed 2, New York, 1998, Oxford University Press.)
When a muscle is held at a constant length, such as when a joint is held still, there is usually enough stretch of the muscle’s spindles to produce a steady frequency of action potential discharge in the sensory neuron output of the muscle spindle. This steady-state discharge gives the muscle spindle organ the ability to inform the CNS not only about a subsequent lengthening of the muscle, which would produce a proportional increase in action potential discharge, but about a shortening of the muscle as well, which would produce a proportional decrease in action potential discharge from the steady state (see Figure 8-4).
Muscle Stretch and Action Potentials Along Spindle Sensory Neurons Lead to Reflex Contraction of the Extrafusal Muscle Fibers
The sensory output neurons of the muscle spindle enter the CNS, where they make excitatory, monosynaptic connections with α motor neurons that return to the extrafusal fibers of the same muscle (see Figure 8-3). Therefore, stretching a given muscle can lead to a rapid, reflex contraction of that same muscle, bringing it back to its original length. Stretching the muscle lengthens the intrafusal muscle fibers of the spindle, increasing the frequency of action potential discharge along the sensory output neurons of the spindle. This leads to an increase in action potential frequency in the α motor neurons on which the spindle sensory neurons synapse. This produces contraction of the extrafusal fibers innervated by those α motor neurons, which results in contraction (shortening) of the muscle. Contraction of the muscle results in a shortening of the muscle spindle’s equatorial region. This eventually reduces the frequency of action potentials occurring on the spindle sensory neurons to the prestretch level, terminating the response. (The cycle is a classic negative-feedback system.)
The reflex just described can be elicited by striking the patellar tendon (insertion tendon of quadriceps muscle) with a blunt object. Because this tendon goes over a “pulley” (the patella), hitting this tendon results in a longitudinal stretch of the whole quadriceps muscle, thus also stretching the muscle spindles. Action potentials from spindle sensory neurons go to the lumbar spinal cord, by way of the dorsal roots, and cause excitatory postsynaptic potentials (EPSPs) on the α motor neurons of the motor units that return to the quadriceps muscle. This causes contraction of the quadriceps muscle and an extension of the knee joint and is an example of the muscle stretch reflex, or myotatic reflex. When it is applied to the quadriceps muscle, it is called the knee jerk reflex, but the mechanisms are present in almost all muscles. However, this is the muscle from which it is easiest to evoke the stretch reflex, because it is one of the few whose tendon goes over a sesamoid pulley before inserting on the next bone. Because of the pulley under the tendon, a lateral deflection of the tendon, as from a reflex hammer, results in a longitudinal stretch of the muscle and thus the reflex. Hitting other tendons only moves the muscle belly laterally and does not easily result in the stretch reflex. Therefore, in the clinical neurological examination of most animals, the knee jerk reflex is the most commonly evoked muscle stretch reflex.
The muscle spindle organ and stretch reflex allow the CNS to make automatic, usually unconscious adjustments to muscle stretch imposed by small changes in body position or the weighting of a muscle. Such adjustments can return the muscle to its original length, often returning a joint to its original position. Clinical examination of the stretch reflex provides clues about the peripheral or CNS integrity of its sensory and motor components.
When the stretch reflex acts to return a joint to its original position, the antagonist of the stretched muscle must relax in order for the joint to be able to move. Therefore, in the stretch reflex, some terminal branches of individual spindle sensory neurons do not synapse directly on the α motor neurons of the stretched muscle, but rather synapse on inhibitory spinal interneurons (see Figure 8-3). These neurons, which lie completely within the spinal cord, are also activated by the stretch of the muscle. When they fire action potentials, however, they cause inhibitory postsynaptic potentials (IPSPs) on α motor neurons that innervate the antagonist of the stretched muscle. This prevents the contraction of the antagonistic muscle.
The Central Nervous System Can Control Spindle Sensitivity Directly Through the Gamma (γ) Motor Neurons
Contraction of the extrafusal muscle fibers is controlled by the larger α motor neurons; intrafusal muscle fibers of the spindle are controlled by the smaller γ motor neurons. The γ motor neurons innervate the intrafusal muscle fibers at their polar ends (see Figure 8-1 and Figure 8-3), the regions containing contractile protein. Action potentials on the γ motor neurons cause shortening of the polar regions of the intrafusal muscle fibers, stretching the equatorial portion.
An important function for this motor innervation of a receptor organ is to regulate the sensitivity of the muscle spindle. Shortening of a gross muscle resulting from initiation of extrafusal muscle fiber contraction has the potential to slacken the intrafusal muscle fibers given their parallel relationship to the extrafusal fibers. This would severely limit the ability of the muscle spindle to transduce stretch. However, this does not normally occur because contraction of the polar regions of intrafusal fibers resulting from γ motor neuron activation is initiated concurrently with shortening of extrafusal fibers caused by α motor neuron activation. This allows the spindle receptor organ to remain taut and sensitive to sudden stretches of the gross muscle over the entire range of its length. This γ motor neuron control mechanism can also function to differentially regulate the sensitivity of the muscle spindle, depending on the type of movement to be made (e.g., novel and unpredictable vs. stereotypical). Chapter 10 describes how co-activation of both the α and γ motor neurons allows the brain to test whether the amount of contraction intended by the brain was what actually occurred.
The Golgi Tendon Organ Lies in Series Between Muscle and Tendon and Detects Muscle Tension
Each Golgi tendon organ is a slender capsule, at the junction between muscle and tendon, in series with 15 to 20 extrafusal skeletal muscle fibers (Figure 8-5). The capsule of each tendon organ contains a complement of braided collagen fascicles, among which the branches of a single sensory neuron are intertwined. This sensory neuron, as with those of the muscle spindle, carries action potentials to the CNS by way of peripheral nerve and dorsal root. The Golgi tendon organ has no motor innervation.
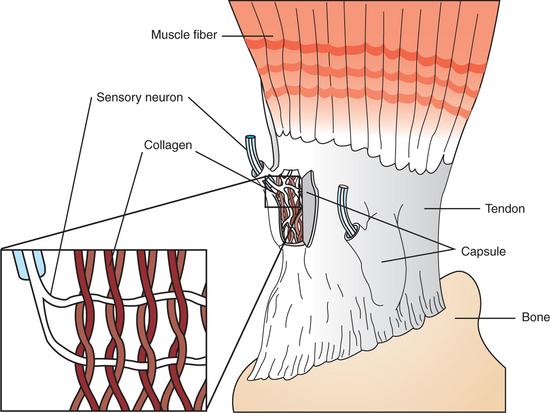
FIGURE 8-5 The Golgi tendon organ is located in the tendons of skeletal muscle, in series with the extrafusal fibers. It detects tension in the tendon, produced by muscle contraction, and sends information about this tension to the central nervous system. Sensory neuron branches of the organ are intertwined among braided collagen fibrils (inset), which fold up and pinch the neural branches when tension develops in the tendon.
Because the Golgi tendon organ is in series with a group of extrafusal fibers and the tendon, when the extrafusal fibers shorten during contraction, tension is applied to the tendon organ. This causes the braided collagen fibrils of the organ to tighten and squeeze the endings of the sensory neuron. Action potentials are therefore generated and sent to the CNS along the sensory neuron at a frequency proportional to the tension developed by the muscle. In contrast, the muscle spindle is arranged in parallel with the extrafusal muscle fibers, and when they contract, the spindle reduces its action potential frequency.
When action potentials from spindle sensory neurons reach the CNS, as mentioned earlier, they monosynaptically produce EPSPs in the α motor neurons returning to the same muscle. Action potentials along sensory neurons from Golgi tendon organs have the opposite effect: they activate inhibitory interneurons, polysynaptically producing IPSPs on α motor neurons to the same muscle. This leads to a reduced extrafusal muscle fiber contraction.
The two skeletal muscle receptors discussed in this chapter provide the CNS with vital information about muscle length (the spindle) and muscle tension (the Golgi tendon organ), and through their reflex circuitry, they help to keep these parameters within optimal ranges. The information these receptors provide, along with information from receptors of skin and joints, is essential if the CNS is to coordinate posture and locomotion.
CLINICAL CORRELATIONS
Femoral Nerve Mononeuropathy
History.
You examine an 8-year-old male golden retriever. The owner complains that the dog cannot bear weight on the right rear leg.
Clinical Examination.
Physical examination deficits are limited to the right rear leg, where you find that the quadriceps femoris muscles are much smaller than those of the left rear leg. The dog cannot bear weight on the right rear leg because the right quadriceps femoris muscles are paralyzed. When you tap on the left patellar tendon with a reflex hammer, the knee briskly extends (the knee jerk or muscle stretch reflex). However, when you tap on the right patellar tendon, no movement occurs.
Comment.
The quadriceps femoris muscle group is one of the major antigravity muscle groups of the leg causing the stifle joint (knee joint) to extend. The paralysis in this animal’s quadriceps muscle is the reason why he cannot bear weight on the leg. The small size of the right quadriceps muscle is caused by atrophy, or muscle wasting, which in turn is caused by the loss of α motor neurons to the extrafusal muscle fibers in the quadriceps muscle belly (see Chapter 9). This would also cause a loss of the muscle stretch reflex, because even though the spindle detected the stretch of the muscle belly caused by either gravity or the reflex hammer, the α motor neurons returning to the quadriceps muscle are unable to signal the muscle to contract, thus completing the reflex arc. This syndrome could occur if the femoral nerve is damaged by a tumor or trauma. If the pathological lesion were in the peripheral nerve rather than only in the ventral roots, there would likely be some sensory loss in addition to the motor deficits.
Acknowledgment
The authors thank Dr. Tom Caceci for persevering in the search through his histological specimens in order to capture the beautiful muscle spindle images of Figure 8-2.
Binder MD. Peripheral motor control: spinal reflex actions of muscle, joint and cutaneous receptors. Patton HD, Fuchs AF, Hille B, et al. Textbook of physiology, ed 21, Philadelphia: Saunders, 1989.
Brodal P. The central nervous system: structure and function, ed 3. New York: Oxford University Press, 2004.
Guyton AC, Hall JE. Textbook of medical physiology, ed 11. Philadelphia: Saunders, 2006.
Haines DE. Fundamental neuroscience. New York: Churchill Livingstone, 1997.
Kandel ER, Schwartz JH, Jessell TM. Principles of neural science, ed 4, New York: McGraw-Hill, 2000.
Purves D, Augustine G, Fitzpatrick D, et al. Neuroscience, ed 3. Sunderland, Mass: Sinauer Associates, 2004.
PRACTICE QUESTIONS
1. If the distance between the origin and insertion tendons is increased (the muscle is stretched), what happens to the frequency of action potentials along the sensory axons leaving the muscle spindles in that muscle?
2. Activation of the Golgi tendon organ of a given muscle:
3. Which of the following is not characteristic of the muscle spindle?