12 The Neck, Back, and Vertebral Column of the Dog and Cat
The neck and back regions are of increasing clinical importance in the companion animals. This is due to the better recognition of veterinary physiotherapy as a profession and to mounting evidence that lameness in dogs is frequently referable to back problems.
CONFORMATION AND SURFACE ANATOMY
The length and proportions of the neck vary with the breed; its transverse section, generally circular in smaller dogs, is somewhat compressed from side to side in larger breeds but widens toward the trunk, with which it blends smoothly. Only a few breeds show a significant elevation at the withers. In most the back slopes slightly downward toward the tail. German Shepherd dogs show a much larger slope; these dogs walk with strongly flexed stifles and hocks. In some breeds the back is level, and in a few (including the Greyhound) it rises toward the loins after dipping over the thorax. The carriage of the tail is variable. Some conformations are characteristic of certain breeds (e.g., the tightly coiled tail of the Spitz breeds), while others express temporary mood, such as the stiffened tail that is held level or upright to denote aggressive intent and the tail depressed to cover the anus in the cringing submissive attitude. The back of a sitting dog is almost straight.
Surprisingly little of the vertebral column is palpable, even in moderately lean subjects. The external occipital protuberance is a distinct landmark at the cranial end of the neck, and behind this the wings of the atlas and the spinous process of the axis are easily distinguished, confirming the position of these two vertebrae close to the dorsal surface. The remaining cervical vertebrae are more deeply placed, and it is sometimes only with difficulty—if at all—that their transverse and spinous processes can be appreciated. Only the tips of the spinous processes can be palpated with certainty in the remainder of the column until the tail is reached. The dorsal parts of the scapulae and the iliac crests provide certain landmarks in the regions of the withers and hindquarters.
In cats the dorsal borders of the scapulae are very prominent and bound a hollow over the adjacent part of the vertebral column. The hollow deepens and the scapular ridges become very pronounced when the trunk is lowered between the forelimbs in the posture cats assume when stalking. Cats also vary in the conformation of the neck, trunk, and tail. Many shorthair cats can be described as cobby, an adjective suggesting a short, thick neck and a thick, deep and fairly short trunk that is carried rather close to the ground. Cats of Oriental breeds are more slender and have a longer and narrower trunk raised from the ground on limbs that are proportionally longer, especially behind. The slinky, svelte appearance is accentuated by the longer tail and smooth flat coat. When a cat sits, its back is arched. The neutral carriage of the tail is slightly drooping, but changes from this posture are frequent and revealing to observers of cat behavior. Domestic cats, uniquely and as a behavioral trait acquired in domestication, often carry the tail upright when they are apparently content and at ease, as when greeting an attentive owner or meeting a conspecific from which they anticipate a friendly reception. The tucked-under position of the tail of the fearful cat crouching in submission and the side-to-side lashing of the cat in a pugnacious mood or merely irritated by unwanted attention will be universally familiar.
THE VERTEBRAL COLUMN
The dog has 7 cervical, 13 thoracic, 7 lumbar, 3 sacral, and about 20 caudal vertebrae as a rule (Figure 12–1); the most common variation is the reduction to six of the lumbar vertebrae. The precaudal vertebrae formula is the same in cats, in which the individual bones are generally more slender and differ from those of the dog in subtle ways that are easy to recognize but difficult to define (Figure 12–2).

Figure 12–1 The skeleton of the dog. 1, Wing of atlas, first cervical vertebra (C1); 2, spine of axis (C2); 3, ligamentum nuchae; 4, scapula; 5, last cervical vertebra (C7); 6, cranial end (manubrium) of sternum; 7, humerus; 8, ulna; 8′, olecranon; 9, radius; 10, carpal bones; 11, metacarpal bones; 12, proximal, middle, and distal phalanges; 13, sacrum; 14, hip bone (os coxae); 15, femur; 16, patella; 17, fibula; 18, tibia; 19, tarsal bones; 19′, calcanean tuber; 20, metatarsal bones; T1, L1, and Cd1, first thoracic, lumbar, and caudal (tail) vertebrae.

Figure 12–2 The feline skeleton. 1, Axis (C2); 2, scapula; 3, manubrium of sternum; 4, clavicle; 5, humerus; 6, radius; 7, ulna; 7′, olecranon; 8, carpal bones; 9, sacrum; 10, hip bone (os coxae); 11, femur; 11′, patella; 12, fibula; 13, tibia; 14, tarsal bones; 14′, calcaneus; 15, metatarsal bones; C1, T1, L1, and Cd1, first cervical, thoracic, lumbar, and caudal (tail) vertebrae.
The intervertebral disks of both dog and cat are relatively thicker than in most species and contribute some 15% and 17% to 20%, respectively, of the total length of the column. Longitudinal growth of the column continues until approximately 12 months of age, when the epiphyses fuse with the bodies of the vertebrae—except in the sacral region where there is some delay. Table 12–1 records the ages at which the secondary ossification centers of the vertebrae appear and those at which they later fuse.
Table 12–1 Development and Maturation of the Canine* Vertebral Column
| Ossification Centers Present of Birth (after Birth) | Approximate Age at Growth Plate Closure Observed on Radiographs |
|---|---|
| Vertebrae, except C1 and C2 | |
| Cranial epiphysis (2–8 wk) | 7–14 mo† |
| Body | |
| Caudal epiphysis (2–8 wk) | 7–14 mo† |
| Two sides of arch | |
| Atlas | |
| Ventral arch | |
| Two sides of dorsal arch | 4 mo‡ |
| Axis | |
| Apex of dens (3–4 mo) | 3–4 mo‡ |
| Dens and cran. articular surface | 7–9 mo‡ |
| Intercentrum (3 wk) | 4 mo‡ |
| Body | |
| Caudal epiphysis (3 wk) | 7–9 mo‡ |
| Two sides of arch | 3 mo‡ |
* Similar information for the cat appears to be lacking.
† Based on Hare WCD: Zur Ossifikation und Vereinigung der Wirbelepiphysen beim Hund, Wien Tierärztl Monatsschr 48:210–215, 1961.
‡ Based on Hare WCD: Radiographic anatomy of the cervical region of the canine vertebral column, JAVMA 139:209–220, 1961.
From de Lahunta, A., and R.E. Habel, 1986.
The contours of the vertebral column do not reproduce the dorsal profile of the standing animal. The convex nape is followed by a relatively straight cervical section. A pronounced but concealed change in direction at the cervicothoracic junction redirects the column on an ascending course in relation to the contour of the back. The caudal thoracic and lumbar segments are fairly straight (depending on the breed), but over the pelvis the column curves ventrally into the tail.
The caudal end of the cervical segment is the most flexible part, and this enables the dog to reach almost every part of its trunk and limbs with its mouth. Ventral flexion to lower the head to the ground is mainly the result of movement in the cranial thoracic joints, and the cervical vertebrae are merely brought into line. Considerable mobility of the caudal thoracic and lumbar joints is necessary for the alternating sagittal flexion and extension of the back in the bounding gallop used by both cats and dogs when moving at speed. This enables the hindlimbs to be placed alongside (if not ahead of) the forelimbs, after which the hindlimb joints and those of the column extend to hurl the body forward. Lateral flexion of the joints of the thoracic and lumbar segments is surprisingly free and enables dogs to curl up when sleeping. The spine of the cat is even more supple.
At three locations in the vertebral column the dorsal parts of the vertebral arches are less closely connected and leave relatively wide interarcuate spaces: the atlantooccipital space between the occipital bone and the first vertebra, the atlantoaxial space between the first and second vertebrae, and the lumbosacral space between the last lumbar vertebra and the sacrum. These interarcuate spaces are of clinical importance because they can be used to allow entry to the vertebral canal for injections or for obtaining samples of cerebrospinal fluid. From the clinical point of view it is important to be familiar with the appearance of the vertebral column in radiographs of both juvenile and mature animals, especially at these three junctions (Figures 12–3, 12–5, and 12–6).

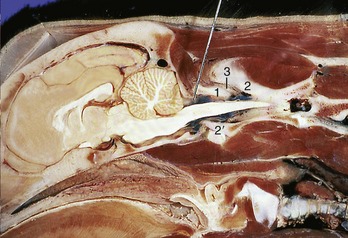
Figure 12–3 A, Lateral radiograph of the head–neck junction of an intubated dog. Note the needle in the atlantooccipital space for a cerebrospinal fluid tap. The dorsal contour of the skull is marked. B, Myelogram of an intubated dog. 1, Lateral vertebral foramen of atlas; 1′, wing of atlas; 2, axis; 3, cricoid cartilage; 4, angular process of mandible; 5, tympanic bulla; 6, soft palate; 7, spine of scapula; 8, spinous process of T1; 9, ventral tubercle of C6.
Because of the frequency with which spinal problems are encountered in clinical practice, it may be useful to recapitulate and amplify the descriptions given in Chapter 2.
ATLAS
The ventral arch of the atlas is considerably narrower (craniocaudally) than the dorsal one. The lateral vertebral foramen for the first cervical nerve is close to the cranial border of the dorsal arch; a notch in the cranial border of the wing replaces the alar foramen of other species and transmits the ventral branch of the same nerve. The wings, extended transverse processes, slope caudally and overlap the atlantoaxial junction. The base of the wing is perforated by the transverse foramen (see Figure 2–7, A).
The dorsal and ventral arches of the atlas participate in the deep cranial articular foveae, which receive the occipital condyles. The single joint cavity has a U shape with its dorsal parts widely spaced and only narrowly joined ventrally. The atlantooccipital membrane extends from the dorsal border of the foramen magnum to the dorsal arch of the atlas, and by attaching laterally to the joint capsules, it closes the atlantooccipital aperture. This membrane is punctured in the collection of cerebrospinal fluid and in the injection of radiopaque contrast agent into the subarchnoid space (Figure 12–3, A-B, and Figure 12–4).
AXIS
The axis is characterized by its length and its enormous spinous process, which overhangs both the dorsal arch of the atlas and the laminae of the third vertebra and carries the caudal articular processes. The cranial extent of the spinous process matches that of the dens, which rests on the dorsal surface of the ventral arch of the atlas (see Figure 2–7 and Figure 2–8). The dens, the displaced body of the atlas, is the pivot around which the atlas and thus the head rotates. The atlantoaxial joint is enclosed by a single joint capsule. The two bones are held in apposition by a thin median ligament (ligamentum apicis dentis), which connects the tip of the dens with the ventral border of the foramen magnum (see Figure 2–13), and paired (alar) ligaments, which pass obliquely from the dens to the ventrolateral borders of the foramen. The dens is further secured by a transverse ligament connecting the inner walls of the ventral arch of the atlas across its dorsal surface. This transverse ligament allows rotations but prevents impingement of the dens on the spinal cord. The dens plays an important role in stability of the atlantoaxial joint, and in cases of aberrant development of the growth plate (early fusion, partial fusion, or nonfusion) there is instability; this is most often seen in miniature breeds (Chihuahua, Toy Pomeranian, Pekingese, and Toy Poodle).
The atlantoaxial membrane closes the interarcuate space; its median part is thickened by elastic fibers that connect the cranial tip of the spine of the axis with the tubercle on the dorsal arch of the atlas (dorsal atlantoaxial ligament) (Figure 12–4/3).
THIRD TO SEVENTH CERVICAL VERTEBRAE
The spinous processes of the remaining cervical vertebrae increase in height and in cranial inclination. The ventral crests are most prominent at the caudal ends of the bodies, marking the positions of the intervertebral disks directly caudal to them. The transverse processes have distinct cranial and caudal extensions (ventral and dorsal tubercles). The ventral tubercle of the sixth vertebra is a nearly sagittal plate that projects considerably below the contour of the body (Figure 12–3/9). The transverse process of the seventh is a rodlike lateral projection that does not overlap the body ventrally. The caudal extremity of that body bears an articular fovea for the head of the first rib. The flat articular surfaces of the synovial joints are nearly horizontal. The cranial articular processes, which provide the ventral component of these joints, narrow the large intervertebral foramina from above.
The wide joint spaces of the atlantooccipital and the atlantoaxial joint support relatively free vertical and rotational movements. The nuchal ligament extends from the spinous process of the axis to the tip of the first thoracic spinous process; it is then continued by the supraspinous ligament until the third sacral vertebra. The nuchal ligament plays an important role in the support of the head of the dog and must be spared during surgery (see Figure 2–8/3). The ligament is not present in cats, but they do possess a supraspinous ligament.
Disorders of the cervical vertebral column, producing compression of the spinal cord, occur in large dogs, especially the Great Dane and the Doberman Pinscher. These disorders may involve deformation of the vertebral arch, malformation of articular facets, vertebral instability of C5–C6 or C6–C7, and dorsal displacement of the vertebral body.
THORACIC VERTEBRAE
The bodies of the thoracic vertebrae are relatively short but increase in length from the tenth caudally (Figure 12–5). The long spinous processes of the first half of the thoracic region are of about equal length. Those of the second half gradually decrease in height; their caudal inclination changes at the eleventh thoracic, the anticlinal vertebra. A more noteworthy change occurs in the orientation of the articular surfaces. On the first 10 (or so) thoracic vertebrae these surfaces lie roughly in a dorsal plane (like those of the cervical vertebrae); caudal to this they are nearly sagittal, and the cranial articular processes enclose the caudal ones (see Figure 2–10). The articular spaces of the former joints are best depicted in lateral radiographs (Figure 12–6), and those of the latter, in ventrodorsal radiographs. The more cranial thoracic vertebrae favor lateral movement of the column, whereas the more caudal bones favor sagittal flexion and extension. Other features of the canine and feline vertebrae are the presence of the mamillary and accessory processes. The mamillary processes are short dorsal projections of the transverse processes that first appear at the third thoracic vertebra and, from the eleventh, migrate dorsally to surmount the cranial articular processes. The accessory processes arise from the caudal border of the pedicle and are present from the midthoracic to midlumbar regions; they are confined to the last three thoracic vertebrae in cats (see Figure 2–11/1,2).

Figure 12–5 Lateral radiographs of canine (A) and feline (B) thoracic and lumbar vertebrae. Radiograph A was obtained after the injection of a contrast agent into the subarachnoid space. 1, Scapular spines; 2, spinous process of T5; 3, anticlinal vertebra (T11); 4, spinous process of L1; 5, rudimentary rib; 6, sublumbar muscles.

LUMBAR VERTEBRAE
The lumbar vertebrae continue several features of the thoracic vertebrae. Their bodies are about twice as long as those of the first thoracic vertebrae and are characterized by long transverse processes that sweep cranioventrally, overlapping the preceding vertebra (Figure 12–7/1). The ventral deflection of these processes is even more pronounced in the cat. The interarcuate spaces of both lumbar and thoracic segments are very small, which makes access with a needle to this part of the vertebral canal difficult. The space at the lumbosacral junction is much better suited for this purpose. It is about 1 cm in diameter (in medium-sized dogs) and lies in the transverse plane of the highest palpable points on the wings of the ilia but about 2 cm deeper. In the cat, the interarcuate space between the last two lumbar vertebrae is also wide enough to allow injection into the vertebral canal.

Figure 12–7 Lateral radiograph of the lumbar region of a cat. 1, Transverse processes of L4; 2, articular processes; 3, last lumbar vertebra (L7); 4, promontory (of sacrum); 5, shaft of ilium; 6, first tail vertebra (Cdl).
The mamillary processes are also fused with the cranial articular ones in the lumbar regions.
SACRAL VERTEBRAE
Fusion of the three segments that constitute the sacrum may not be completed until 18 months after birth; fusion involves both the vertebrae and the ossified intervertebral disks. The sacrum is deeply embedded between the wings of the ilia so that only the spinous processes (sacral crest) are palpable through the skin; however, its caudoventral part and the first few (or more) caudal vertebrae can be palpated digitally per rectum. The first sacral vertebra forms a rigid joint with the wing of the ilium.
CAUDAL VERTEBRAE
Features of certain caudal vertebrae (usually the fourth to sixth) are the hemal arches, small V-shaped bones attached to the caudal ends of the ventral surfaces (see Figure 2–12/9). Short hemal processes are found in similar positions on several more segments. They offer protection to the vessels on the ventral surface of the vertebrae. Congenital anomalies of the cat tail include the distinctive Manx “bob” and the kinking formerly common in Siamese, though largely bred out of the modern breed.
INTERVERTEBRAL DISKS
Intervertebral disks are present in every intervertebral space except that between the first and second cervical vertebrae (p. 39). The functional importance of the disks lies in their contributions to the flexibility of the spine and to the distribution of pressure over the extremities of the vertebrae. Their clinical significance lies in the frequency with which degenerative changes occur; these may almost be regarded as part of the normal aging process when not too profound. The changes include the metaplastic changes of fibrous tissue, the calcification of the gelatinous nucleus, and, frequently, the separation and rupture of the fibrous lamellae of the anulus. The narrow dorsal part of the anulus is most vulnerable, and when degeneration is advanced, stretching or total rupture of this section allows disk material to protrude into the vertebral canal where it may press (through the meninges) on the spinal cord and nerves producing various and often severe dysfunctions.
The dorsal longitudinal ligament is well-developed in the cervical region, which prevents dorsal herniation of disk material into the vertebral canal. Instead, the disk material protrudes dorsolaterally toward the roots of the spinal nerves, which results in their compression. Approximately 15% of disk problems in dogs occur in the cervical region; the clinical signs are neck pain, spasms of shoulder muscles, and lameness due to pain referred to a forelimb. The presence of the intercapital ligaments (between the heads of a rib pair passing beneath the dorsal ligament) at the joints T1–T2 to T9–T10 offers almost complete protection against herniation to the greater part of the thoracic cord (see Figure 2–18). Thoracolumbar lesions account for the remaining 85% of intervertebral disk problems (T11/12 to L1–L2). In the caudal thoracic and lumbar regions of the spine where the dorsal longitudinal ligament is thinner, dorsal protrusions and consequent spinal cord compressions are more frequent.
Common radiographic findings in cases of herniation are narrowing or collapse of the intervertebral disk space, collapse of the synovial joints, narrowing of the intervertebral foramen, and calcified material within the vertebral canal. Misinterpretation of apparent narrowing of intervertebral disk spaces is easy if insufficient attention is paid to the geometry of image formation (p. 5). Furthermore, it should be emphasized that nuclear calcifications are often evident in radiographs obtained of dogs that present no signs of dysfunction or pain. The intervertebral disks of cats are not immune to degeneration, but for reasons that are obscure, affected animals very often fail to manifest any clinical signs.
There are both breed and regional differences in the incidence of disk pathology. Chondrodystrophic breeds, such as the Dachshund and Pekingese, in which the degenerative process is both precocious and accentuated, are particularly prone to protrusions at a relatively early age. In normal dogs, disk disease is characterized by slow fibroid degeneration, most evident between 8 to 10 years of age; mineralization of the disk is unusual. As a result of chronic degenerative disk disease (without clinical signs), spondylosis may develop. The sites most frequently involved are those undergoing the greatest mechanical stress. As a result of the stress, bony spurs are formed ventral and lateral to the intervertebral disk space, leading ultimately to complete fusion of vertebrae. On survey radiographs the presence of spondylosis is often regarded as an incidental finding.
THE VERTEBRAL CANAL
The diameter of the vertebral canal is greatest at the level of the first and second cervical vertebrae. It is reduced in width throughout the cervical spine, increases again in the cranial thoracic region, and becomes narrower in the caudal thoracic region. The diameter widens again in the lumbar region to accommodate the caudal intumescence of the cord, before gradually narrowing when followed into the tail.
The spinal cord and nerve roots are surrounded by three meningeal layers: the tough outer fibrous dura mater, the thin arachnoid membrane lining the inner surface of the dura, and the pia mater, which is attached to the spinal cord. The dura mater adheres to the periosteum of the first two cervical vertebrae but separates thereafter (Figure 12–8), leaving a relatively narrow epidural space (see Figure 12–4) that contains fat. The cerebrospinal fluid in the subarachnoid space and the epidural fat cushion the cord and allow displacement during normal movements of the spine.

Figure 12–8 Dorsal view of opened vertebral canal (cat). 1, Spinal nn. penetrating arachnoid and dura mater.
The cord is the thickest in the atlas, where it measures about 1 cm. Elsewhere, except for the cervical and lumbar enlargements, it is approximately half that diameter. The cervical enlargement involves cord segments C6–T1, from which the nerves forming the brachial plexus arise, while the lumbar enlargement with a similar relationship to the lumbosacral plexus involves cord segments L5–S1. The ascent of the cord (p. 307) within the vertebral column explains the topography of its segments (Figure 12–9). Most cervical spinal cord segments are positioned about half a vertebra, and most thoracic segments a whole vertebra, cranial to the vertebra of the same numerical designation, while the caudal thoracic and cranial lumbar segments occupy vertebrae of the same designation. From the midlumbar region onward, the cord segments are markedly shorter, and this generally places the end of the cord over the last interlumbar joint (Figure 12–9 and Figure 8–56, B). The cervical and lumbosacral enlargements lie in the sixth and seventh cervical and the fourth and fifth lumbar vertebrae, respectively. The ascent is less marked in small dogs, in which the cord may reach the sacrum; in large dogs it may end at L4. The sacral canal contains only spinal nerves and the dural sheath, which extends about 2 cm beyond the cord. The termination of the cord is said to be variable in cats: all levels from the caudal border of L7 to the caudal border of S3 have been given by different authors. Some of this uncertainty may be due to individual and breed variation, but probably the more cranial limit is likely to be nearer the mark in adults, the more caudal one in young kittens.

Figure 12–9 Caudal end of the canine spinal cord in situ, dorsal view. 1, Third lumbar vertebra; 2, last lumbar vertebra (L7); 3, sacrum; 4, first caudal vertebra; 5, dura mater; 6, dorsal root ganglion; 7, approximate level of L7 cord segment; 8, end of spinal cord; 9, cauda equina.
The cauda equina is the bundle of spinal nerves remaining after the termination of the spinal cord. It consists of the spinal nerves L6–Cd5 and includes those that form the sciatic (L6–S1) and pudendal (S2–3) nerves, among others. Each nerve emerges from its respective intervertebral foramen. It may be trapped here by herniation of the adjacent disk. In the sacrum the ventral branches of the first two sacral nerves emerge through foramina in the floor of the sacral canal. The “cauda equina syndrome” is an important cause of neurological dysfunction and pain and is caused by entrapment of one or more of the nerves of the cauda equina. The symptoms may include lower back pain, atrophy of muscles innervated by the sciatic nerve, paresis, tail weakness, incontinence of bladder and bowel, and paresthesias (sensory disturbance possibly provoking self-mutilation). Entrapment may also be caused by bone pathology (e.g., osteochondrosis of the first sacral vertebra) or by secondary hypertrophy of the ligaments.
Puncture of the subarachnoid space is performed for the collection of cerebrospinal fluid and for the injection of contrast media for myelography. Myelograms outline the subarachnoid space and may thus reveal cord lesions that are not apparent in survey radiographs; they are especially useful in revealing multiple lesions or when survey radiographs do not match the clinical signs. Recommended sites for puncture are the atlantooccipital space and the lumbar spine at levels L4–L5 and L5–L6. Atlantooccipital punctures enter the cerebromedullary cistern of the subarachnoid space (see Figure 12–4). In this procedure, the head is strongly flexed on the neck, and an entry is made midway between the external occipital protuberance and the cranial tip of the spinous process of the axis. These landmarks are more easily identified before flexion (Figure 12–3, A). The lumbar puncture (see Figure 12–7) is more difficult and can result in penetration of the spinal cord; it is reported that the clinical effect is minimal, although it provokes a histological reaction.
Epidural anesthetics are administered at the lumbosacral or the sacrocaudal space. The lumbosacral space is located a little in front of the cranial dorsal iliac spines (see Figure 12–7). A misleading impression of its size may be obtained by failure to appreciate that a last lumbar spinous process is relatively short and fails to approach as closely to the skin as that of the preceding vertebra. The sacrocaudal space is smaller, and the defining features of the vertebrae less salient, but compensation is found in the closeness of this space to the skin.
As in other species, the internal vertebral venous plexus consists of two longitudinal valveless veins on the floor of the vertebral canal, where they are embedded in epidural fat (Figure 12–10/6′; p. 314). The left and right veins anastomose frequently at different levels; some connections pass below the dorsal longitudinal ligament while others pass through the vertebral bodies. The plexus receives blood from the spinal cord and vertebrae; it is linked to extensive but less regular external networks and to adjacent great veins (caudal vena cava, azygous vein) by intervertebral veins. These veins, which may be double and triple, cushion the spinal nerves where they leave the vertebral canal.

Figure 12–10 Transverse section of the back of a dog at the level of the first lumbar vertebra. 1, Multifidus and spinalis; 2, longissimus and iliocostalis; 3, last rib; 4, aorta; 5, right crus of diaphragm; 6, first lumbar vertebra; 6′, internal vertebral venous plexus.
At the foramen magnum the veins of the internal plexus are continuous with the right and left basilar sinuses that lead from the system of venous sinuses on the floor of the cranial cavity.
Like many other mammals, the dog makes use of is tail to maintain balance when executing various energetic maneuvers, but the tail is also used as means of communication. Sometimes it is necessary to amputate part of the tail after an injury. A feature relevant to the operation includes the presence of the median caudal artery running below the vertebral bodies. The artery obtains no special protection where it follows the roof of the pelvis, but caudal to this it is partly shielded by processes of bone. The protection takes the form of separate V-shaped hemal bones located below the fourth to sixth tail vertebrae and hemal processes projecting from the ventral aspect of the more distal vertebral bodies. Obviously, amputation is simplest at the level of an intervertebral disk. It may be mentioned that these disks are not immune from the degenerative processes described earlier. In many European countries tail docking for cosmetic reasons is illegal.
THE MUSCLES ASSOCIATED WITH THE VERTEBRAL COLUMN
The muscles directly associated with the neck and back mainly extend between points on the vertebrae (and ribs), but some also attach to the skull, the ilium, and, where the psoas group is concerned, the femur.
Only a superficial acquaintance with these muscles is required in order to appreciate their functional importance and the suitability of the expaxial division for receiving intramuscular injections. A much more detailed knowledge of the locations, construction, and attachments of the individual units is required for those who may comtemplate surgery upon the vertebral column.
The descriptions that follow supply the basic information which is most conveniently arranged under the following heads: the expaxial division; the hypaxial division, consisting of distinct cervical and lumbar groupings; and those muscles whose actions are confined to the movements of the head.
EPAXIAL MUSCLES
The epaxial muscles (see Figure 2–22, B) are used for intramuscular injections. Less commonly, they must be separated and detached when access to the vertebral column is necessary. The epaxial muscles comprise three longitudinal systems: iliocostalis, longissimus, and, transversospinalis. The hypaxial muscles comprise the longus colli and longus capitis muscles in the cervical and cranial thoracic regions, and the psoas muscle in the lumbar region.
The splenius muscle is a strong muscle on the dorsolateral aspect of the neck, extending from the withers to the occiput (see Figure 2–23, A/4). It covers the longissimus capitis muscle, the semispinalis capitis muscle, and parts of the spinalis et semispinalis cervicis et thoracis muscle. It originates from the spinocostotransverse fascia, the spinous processes of the first three thoracic vertebrae, and the nuchal ligament and inserts on the nuchal crest and the mastoid process.
The iliocostalis muscle is relatively thin (see Figure 2–23, B/17) and has only lumbar and thoracic parts. Its bundles span several vertebral segments and, in general, run from caudomedial and dorsal to craniolateral and ventral. The muscle is easily identified over the ribs by the glistening tendons. It arises caudally from the wing of the ilium and also by lumbar fascia from the spinous processes of the lumbar vertebrae. The lumbar portion reduces in size cranially and inserts on the last three to four ribs. The thoracic portion arises lateral to the lumbar part but without any sharp demarcation and extends from the twelfth rib to the transverse process of the last cervical vertebrae.
The iliocostalis is lateral to the longissimus system and is covered by the dorsal serratus and the origins of the latissimus and abdominal oblique muscles. The lumbar part of the iliocostalis muscle of the cat is hardly separate from the longissimus.
The longissimus muscle is much thicker than the preceding muscle (see Figure 2–23, B). Its bundles are similarly oriented but are largely fused, giving a uniform appearance to the lumbar and thoracic regions. The thoracolumbar part (longissimus dorsi) is credited with the powerful extension of the vertebral column during the propulsive phase of the gallop. It is related medially to the multifidus, and, over the thoracic vertebrae, it is covered dorsally by the spinalis et semispinalis (Figure 12–10/1,2), although it is separated from both of these by a fibrous septum, which serves as the origin of the last-named muscle. The ventral edge of this septum ends near the transverse processes of the vertebrae and is a landmark in the surgical approach to the intervertebral disks.
The lumbar part arises from the wing of the ilium and the lumbar spinous processes, against which it lies. Along its length it detaches several bundles, arranged in a lateral and a medial row, which cover the bases of the lumbar transverse processes before ending on accessory processes of the cranial six lumbar vertebrae. The caudal narrow part, not covered by the middle gluteus, inserts dorsally mainly on the arch of the last lumbar vertebra and the last intervertebral disk, with more limited insertion on the sixth and fifth lumbar vertebrae. The longissimus lumborum is covered by a dense aponeurosis, separated from the thoracolumbar fascia by fat.
The thoracic part (see Figure 2–23, B/16″) inserts by medial tendons on the transverse or accessory processes of the thoracic vertebrae and by lateral tendons on the necks of the last seven ribs. The dorsal branches of the thoracic nerves pass between the medial and lateral tendon.
The cervical part (see Figure 2–23, B/16′) of the longissimus muscle has a triangular form, filling up the angle between the cervical and thoracic vertebrae, and comprises four incompletely separable bundles, which arise from the transverse and articular processes of the first thoracic vertebrae and insert on the transverse processes of the sixth to third cervical vertebrae.
The longissimus capitis muscle, strong and flat, lies medial to the longissimus cervicis and the splenius muscle (see Figure 2–23/16′). It originates from the transverse processes of the first three thoracic vertebrae and from the caudal articular processes of the last three to four cervical vertebrae. It runs over the dorsal surface of the atlas and inserts on the mastoid process, fused at the level of the atlas with that of the splenius muscle.
The longissimus atlantis muscle, present in only 20% of dogs, arises from the articular processes of the last three cervical vertebrae and ends on the wing of the atlas.
In the cat there is a longissimus capitis but not an atlantis. Furthermore, it is not possible to separate the cervical and thoracic longissimus muscles; a shallow longitudinal groove appears to separate the lumbar portion into lateral and medial parts.
The more complex transversospinalis system is more intimately related to the vertebrae. Some fascicles connect one vertebra to the next, while others span several vertebrae; most are oriented from caudoventral and lateral to craniodorsal and medial, in contrast to the direction taken by the preceding muscles. The transversospinalis system comprises spinalis et semispinalis thoracis et cervicis, semispinalis capitis, and several less important, more obviously segmental muscles (multifidi, intertransversarii, interspinales and rotatores) that lie directly on the vertebrae (see Figure 2–23, B/15).
The spinalis et semispinalis thoracis et cervicis muscles extend from the midlumbar region to the spine of the axis and lie against the lateral surface of the spinous processes (see Figure 2–24, A/2″-2″′) dorsomedial to the longissimus thoracis. Their fascicles connect spinous and mamillary processes with more cranial spinous processes. It is a powerful muscle incompletely divided into a lateral part, the spinalis et semispinalis thoracis, and a medial part, the spinalis cervicis.
The spinalis et semispinalis thoracis (lateral part) arises from the aponeurosis of the thoracic part of the longissimus muscle and from the midlumbar spinous processes. It inserts on the spinous processes of the sixth thoracic forward to the sixth cervical vertebrae. The tendons that are attached to the last two cervical spines are particularly powerful and form a plate that is fused with the tendon of the medial part of the muscle. In the cat, the spinalis et semispinalis thoracis arises only from the tenth or eleventh thoracic spinous process forward.
The spinalis cervicis (medial part) lies dorsomedial to the lateral part. It arises from the spinous processes of the sixth to first thoracic vertebrae and continues as a flat muscle with four tendinous inscriptions in its belly on the spinous processes of the fifth until second cervical vertebrae.
The semispinalis capitis is a more independent neck muscle and lies between the splenius and the cervical components of the preceding muscle (see Figure 2–23, B/15). It is clearly divided into the biventer cervicis and the complexus, which both contact their fellows and the ligamentum nuchae in the median plane. The biventer is the more dorsal and more caudal of the two. It arises from and around the transverse processes of the first few thoracic vertebrae, medial to the longissimus cervicis and capitis, and ends on the occipital bone ventral to the external occipital protuberance. It can be identified by several tendinous inscriptions. The complexus arises from the articular processes of the caudal four cervical vertebrae and first thoracic vertebra and ends on the nuchal crest; it is not segmented.
In the cat, the biventer is relatively poorly developed, and it presents only two to three tendinous inscriptions. The complexus, even less developed, is divided by a distinct strip of tendon that runs horizontally through its middle.
The multifidus is more distinctly segmented in the dog than in the cat, especially in the cervical region. The lumbar part in the dog consists of 10 to 11 bundles that arise from the mamillary process of the first caudal vertebra, the rudimentary articular processes of the sacrum, and the mamillary processes of the lumbar vertebrae and last two thoracic vertebrae (see Figure 2–24, B/2′). As a rule, two segments are passed by each bundle: thus the insertions are to the spinous processes of the sixth lumbar to the tenth thoracic vertebrae. The thoracic part comprises nine distinct bundles that arise from the mamillary and transverse processes of most thoracic vertebrae and, after passing two segments, insert on the spinous processes of the first eight thoracic and last cervical vertebrae.
The cervical part, completely covered by the semispinalis capitis, consists of six individual parts that divide into large lateral and smaller medial bundles.
The intertransversarii muscles, divisible into lumbar, thoracic, and cervical units, are sometimes regarded as being split from the longissimus system. The lumbar units are especially well developed in cats, whereas in dogs the muscle is composed of thin bundles. In both species they unite the mamillary and accessory processes of the lumbar vertebrae and last four thoracic vertebrae with the transverse processes of the twelfth to sixth thoracic vertebrae, never passing more than three segments.
The cervical intertransversarii are much stronger and are arranged in dorsal, middle, and ventral layers. The dorsal layer is located between the insertions of the longissimus cervicis and capitis muscles and comprises five bundles only partly separable. The middle layer consists of five to six thin, separate parts, of which the deeper fibers run from segment to segment and the superficial ones always pass over one segment. The ventral layer lies dorsal to the longus capitis. It arises from the ventral border, the transverse process of the sixth cervical vertebra, and runs forward, as three digitations, to those of the fourth to second vertebrae.
The interspinales muscles connect the spinous processes of the vertebrae in the lumbar, thoracic, and cervical regions; the lumbar portions are completely covered by the multifidus. The thoracic parts are broader.
Deep rotator muscles lie medial to the multifidi in the cranial thoracic region. The eight long rotators extend between the transverse and spinous, spanning two joints; the nine short ones pass between adjacent vertebrae. It is not possible to separate the rotator muscles from the multifidus in the cat.
CERVICAL HYPAXIAL MUSCLES
The longus colli is located on the ventral aspect of the cervical and first few thoracic vertebrae. The thoracic part originates from the ventral surface of the first six thoracic vertebrae and inserts on the platelike process of the sixth cervical vertebra. The cervical part originates by separate cervical vertebrae and inserts on the ventral parts of the bodies of more cranial cervical vertebrae near the midline (see Figure 2–24, B/9).
The longus capitis is a long flat muscle that lies on the lateral and ventral sides of the cervical vertebrae lateral to the longus colli. It arises from the transverse processes of the middle five cervical vertebrae and inserts on the occipital bone between the tympanic bullae.
LUMBAR HYPAXIAL MUSCLES
The psoas minor muscle can be found between the iliac fascia and peritoneum ventrally and the iliopsoas and quadratus lumborum muscles dorsally. It originates from the bodies of the last thoracic and the first four to five lumbar vertebrae. The strong flat tendon has a shiny appearance and inserts on the iliopubic eminence at the pelvic inlet. It stabilizes and flexes the lumbar part of the vertebral column.
The iliopsoas muscle consists of the psoas major and the iliacus. It lies ventral to the quadratus lumborum and dorsal to the psoas minor. The psoas major arises from the bodies of the lumbar vertebrae and passes caudally, medial to the wings of the ilium, where it fuses with the iliacus to form the iliopsoas (see Figure 2–24, B/11). The iliacus arises from the wing and shaft of the ilium. Both muscles have a common insertion on the lesser trochanter of the femur. The combined muscle flexes the lumbar vertebral column and plays a role in protraction of the hindlimb.
The quadratus lumborum lies directly ventral to the bodies of the last three thoracic vertebrae and the bodies and transverse processes of all the lumbar vertebrae and ends on the medial surface of the wing of the ilium (see Figure 2–24, B/3).
MUSCLES CONTROLLING MOVEMENTS OF THE HEAD
The four straight and two oblique muscles associated with the atlantooccipital and atlantoaxial joints form a group of their own.
The rectus capitis dorsalis major (Figure 12–11/2) arises from the spine of the axis, just cranial to the attachment of the nuchal ligament, and inserts on the nuchal surface of the skull, ventral to the insertion of the semispinalis capitis, by which it is covered.
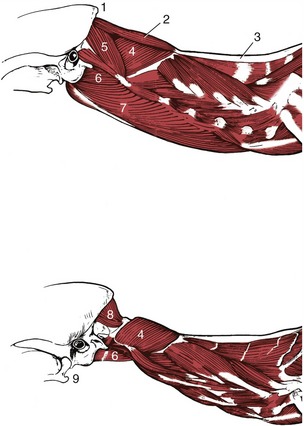
Figure 12–11 Muscles associated with the canine atlantooccipital and atlantoaxial joints, lateral view. 1, External occipital protuberance; 2, rectus capitis dorsalis major; 3, nuchal ligament; 4, obliquus capitis caudalis; 5, obliquus capitis cranialis; 6, rectus capitis ventralis; 7, longus capitis; 8, rectus capitis dorsalis minor; 9, angular process of mandible.
The rectus capitis dorsalis minor (Figure 12–11/8), deep to the preceding muscle, is a short flat muscle; it arises from the dorsal arch of the atlas and inserts on the skull above the foramen magnum.
The rectus capitis ventralis comes from the ventral arch of the atlas and goes to the ventral surface of the occipital bone. It lies dorsal to the much larger longus capitis, which inserts close by.
The rectus capitis lateralis passes between the ventral arch of the atlas and the paracondylar process of the occipital bone. The rectus muscles move the head both up and down and sideways.
The obliquus capitis cranialis arises from the cranial surface of the wing of the atlas and inserts on the nuchal surface of the skull.
The larger obliquus capitis caudalis arises from the lateral surface of the spine of the axis and inserts on the caudal surface of the wing of the atlas. The obliquus muscles are responsible for rotation of the head at the atlantoaxial joint.
CLINICAL CONSIDERATIONS
Muscles of the epaxial and hypaxial divisions must often be separated and detached when access to the vertebral column is necessary. In the neck the ventral approach is most often chosen, although a dorsal approach is also possible. In the lumbar region the dorsal approach is preferred.
The ventral approach to the cervical vertebrae (Figure 12–12) is indicated for disk fenestration in cases of herniation or for the treatment of atlantoaxial instability. The trachea is exposed through a ventral midline incision, midway between the sternomastoid and sternohyoid muscles. Reflection of the trachea to the left protects the esophagus and exposes the paired longus colli muscles, which can be separated longitudinally.
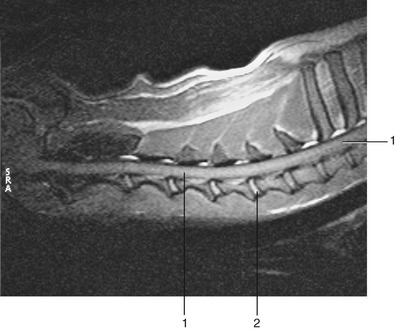
Figure 12–12 Midsagittal section of cervical region of dog, T2 weighted. 1, Spinal cord; 2, nucleus pulposus.
The dorsal approach to the cervical vertebrae is indicated for vertebral fractures; in this approach the biventer cervicis and the rectus capitis dorsalis major are exposed cranially, and the nuchal ligament, the spinalis et semispinalis cervicis, and multifidus cervicis muscles are exposed more caudally. The vertebral artery lies in the rectus capitis dorsalis major, ventrolateral to the synovial joint C1/C2 and must be avoided as the dissection is continued laterally.
The dorsal approach to the caudal cervical and cranial thoracic vertebrae for dorsal laminectomy (removal of part of the vertebral arch) and fracture repair first exposes the aponeuroses of the trapezius cranially and the rhomboid caudally. Then the subscapulares, splenius, and serratus dorsalis are exposed by lateral retraction of the trapezius and rhomboid muscles and the scapula. Finally, the semispinalis capitis, longissimus cervicis, the nuchal ligament, and the dorsal spines of the vertebrae are exposed by lateral retraction of the splenius and serratus dorsalis. The deep cervical artery passes through the semispinalis capitis.
A dorsal approach to the thoracolumbar vertebrae is indicated for dorsal laminectomy and thoracolumbar fractures. Lateral retraction of the lumbar fascia exposes the longissimus lumborum and multifidi caudally and the spinalis et semispinalis thoracis cranially. The multifidus, interspinalis, and rotatores longi are elevated from the spinous processes and vertebral arches. The dorsal branch of each spinal nerve emerges just cranial and ventral to the insertions of the longissimus on the accessory processes.