17 The Hindlimb of the Dog and Cat
THE CROUP, HIP, AND THIGH
(See also pp. 88, 91, and 93–97.)
The habitual stance varies among breeds. The major differences are well illustrated by the German Shepherd, which tends to crouch with the back and croup sloping down toward the tail (and the hip, stifle, and hock joints markedly flexed), and the Boxer, which favors a stiffer, more upright posture (with the major joints, particularly the hock, significantly straighter). The more upright limb appears to predispose to several common stifle disorders. In the Greyhound and other lean, short-coated dogs, the contours of the croup may reproduce the form of the underlying muscles, the superficial gluteal especially; however, such details are more often obscured by subcutaneous fat or a thick coat. The major skeletal landmarks are always palpable and reveal the small angle the ilium makes with the vertebral column.
The dorsal and ventral spines of the ilium are very prominent. The convex (iliac) crest joining these points can also be followed in its length and provides a convenient site for bone marrow biopsy in larger breeds; it is too thin to serve this purpose in smaller animals. A narrow strip of the pelvic floor bordering the ischial arch can usually be palpated between the salient tubers. In the dog the cordlike sacrotuberous ligaments, which are lacking in cats, can also be felt as they approach these projections from their origins on the sacrum. The greater trochanter of the femur is found cranial to the ischial tuber, and because its summit is very nearly level with the femoral head, it provides a good guide to the position of the joint, which is not itself palpable. Attention should be paid to the spacing of these features of the ilium, ischium, and femur because alteration may reveal luxation of the femur. This is a relatively frequent mishap; the femoral head is most often displaced dorsocranially (which widens the ischiofemoral gap) but may pass dorsocaudally or, though rarely, ventrocaudally when it may engage within the obturator foramen. Luxation may be confirmed by rotating the thigh outward while the thumb is pressed between the trochanter and the tuber; the movement normally forces the thumb from the recess, but a luxated femur is unable to exert the necessary leverage.
Although the hip joint is constructed according to the usual plan, it possesses greater range and versatility of movement in the dog and cat than in other domestic species. The enhanced potential for abduction is shown by the ease with which dogs cock their legs when urinating, while the general versatility, taken in combination with the suppleness of the trunk, enables both species to reach most parts of the head, neck, and thorax when scratching with the hindpaw. The articular surfaces reflect these abilities. The femoral head is an almost perfect hemisphere, marred only by the small central fovea where the intracapsular ligament (of the femoral head) inserts; it is deeply seated within the acetabular cup, which is only slightly extended by a labrum about its rim (see Figure 2–62). There are no peripheral ligaments to limit movement, although some capsule reinforcements can be identified. The intracapsular ligament, though variable in length and thickness, is generally lax enough to survive intact when the head is subluxated; this may be the case when there is preexisting dysplasia of the joint, as then the ligament is often hypertrophied. In normal hips, the ligament is thought to exert a braking function on movements that threaten the stability of the joint. The joint capsule also maintains the femoral head within the socket and prevents overextension and flexion. The fit of the femoral head within the acetabulum can be estimated from a ventrodorsal radiograph of the pelvis by measuring the “Norberg angle,” that is, the angle between the line connecting the centers of the femoral heads and that connecting the center of a femoral head with the cranial part of the related acetabular rim. An angle of less than 105° indicates displacement and suggests dysplasia.
The blood supply to the joint capsule, the femoral neck, and the proximal epiphysis arises from the lateral and medial circumflex femoral arteries, which, with some assistance from the caudal gluteal artery, form an extracapsular ring from which branches pierce the joint capsule. These ascend the femoral neck and provide the epiphyseal arteries of the head. Arteries demonstrable in the ligament of the femoral head are thought to be of little significance in the dog but make a major contribution to the supply of the femoral head of the kitten. Trauma to the femoral neck often leads to its absorption because of the limited blood supply.
The most convenient access to the joint, for puncture and in surgery, is from the craniolateral direction. An approach between the tensor and biceps muscles exposes the proximal part of the vastus lateralis (whose origin runs from just below the greater trochanter) and the gluteal muscles that clothe the joint directly. The important structures endangered are the sciatic nerve and the caudal gluteal vessels; however, because these cross the dorsocaudal aspect of the joint as they proceed into the thigh, the risk is relatively remote.
The radiological anatomy is very relevant to the diagnosis of the two conditions that commonly affect the joint: luxation and dysplasia. For the standard ventrodorsal radiograph (Figure 17–1, A) the supine animal must be placed with its hindlimbs drawn uniformly backward to ensure symmetrical depiction of bilateral structures. Although most features of the pelvis are too obvious to require comment, attention may be drawn to the slight lateral bowing of the canine ilia (in contrast to their parallel course in the cat). The relationship between the rim of the acetabulum and the femoral head on which it is superimposed is of the greatest importance in determining the integrity of the joint (Figure 17–1/3). Attention is also directed to the relative radiolucency of the region (corresponding to the trochanteric fossa) between the greater and lesser trochanters of the femur, as it is sometimes misinterpreted. The less useful lateral view reveals the position of the hip joints below the first two caudal vertebrae (Figure 17–1, D).
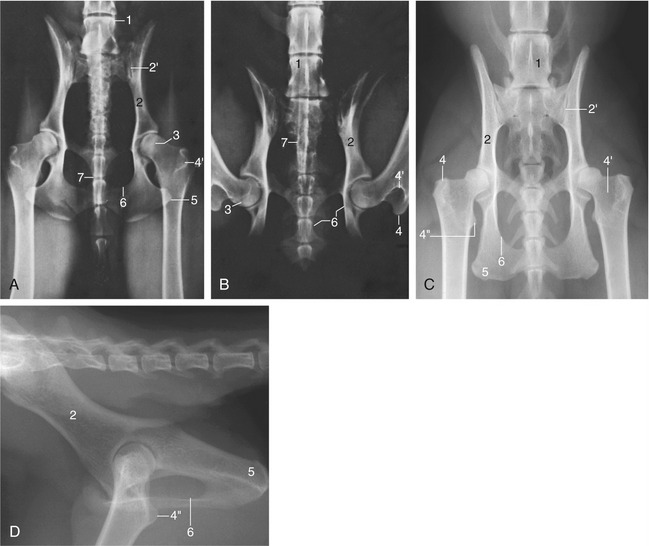
Figure 17–1 Ventrodorsal radiographic views of the canine pelvis with extended (A) and flexed (B) hip joints. C and D, Radiographs of the feline pelvis in ventrodorsal and lateral views; D is taken of a specimen. 1, Last lumbar vertebra (L7); 2, shaft of ilium; 2′, sacroiliac joint; 3, dorsal border of acetabulum superimposed on the femoral head; 4, greater trochanter; 4′, trochanteric fossa; 4″, lesser trochanter; 5, ischial tuber; 6, obturator foramen; 7, os penis superimposed on vertebrae.
The maturation of the skeleton can be followed in radiographs obtained from young animals. In puppies there are primary ossification centers for the bodies of the ilium, ischium, and pubis and for the acetabular bone and secondary centers for the iliac crest, ischial tuber, and the border of the ischial arch. The acetabular bone is the first to lose its independence, but this is followed by the merger of the other primary centers at a comparatively early age (4 to 6 months); the secondary centers remain distinct until much later (15 months to 5 years for the iliac crest and 8 to 14 months for the ischial tuber). Fusion at the proximal extremity of the femur is completed between the 6th and 12th month (Table 17–1 and Figure 5–74).
Table 17–1 Development and Maturation of the Hindlimb Skeleton
| Ossification Centers Present at Birth (After Birth) | Approximate Age at Growth Plate Closure Observed on Radiographs | |
|---|---|---|
| Dog | Cat3 | |
| Os coxae (hip bone) | ||
| Ilium | 4–6 mo1,2,6 | |
| Ischium | 4–6 mo1,2,6 | |
| Pubis | 4–6 mo1,2,6 | |
| Acetabular bone (7 wk) | 4–6 mo1,2,6 | |
| Iliac crest (4 mo) | 15 mo–5.5 yr2 | |
| Ischial tuber, caudal border of ischium (3 mo) | 8–14 mo2,6 | |
| Caudal pelvic symphysis, interischiadic bone (7 mo) | 15 mo–5 yr2,6 | |
| Pelvic symphysis closure (cranial to caudal) | 2.5–6.0 yr2 | |
| Femur | ||
| Lesser trochanter (8 wk) | 8–13 mo1,2,6 | 8–11 mo |
| Greater trochanter (8 wk) | 6–9 mo2,5 | 7–10 mo |
| Head (2 wk) | 6–9 mo2,5 | 7–10 mo |
| Diaphysis | ||
| Distal epiphysis (3 wk) | 6–12 mo2-5 | 13–19 mo |
| Trochlea (3 wk) | 3 mo6 | |
| Patella (9 wk) | ||
| Tibia | ||
| Tibial tuberosity (8 wk) | 8–10 mo2,6 | |
| Proximal epiphysis (3 wk) | 6–15 mo2,5 | 12–18 mo |
| Diaphysis | ||
| Distal epiphysis (3 wk) | 5–11 mo2,5 | 10–13 mo |
| Medial malleolus (3 mo) | 4–5 mo2,6 | |
| Fibula | ||
| Proximal epiphysis (9 wk) | 6–12 mo2,6 | 13–18 mo |
| Diaphysis | ||
| Distal epiphysis (2–7 wk) | 5–13 mo2-5 | 10–14 mo |
| Sesamoids | ||
| Gastrocnemius (3 mo dog; 2.5–4.0 mo cat) | ||
| Popliteus (3 mo dog; 4–5 mo cat) | ||
| Tarsus | ||
| Calcaneus | ||
| Calcanean tuber (6 wk) | 3–8 mo2,4-6 | 7–13 mo |
| Diaphysis | ||
| Other tarsal bones (2–4 wk), 1 center each | ||
| Metatarsus | ||
| Diaphysis | ||
| Distal epiphysis (4 wk) | 5–7 mo2,5 | 8–11 mo |
| Digit similar to forelimb | ||
1 Based on Chapman WL: Appearance of ossification centers and epiphyseal closures as determined by radiographic techniques, J Am Vet Med Assoc 147:138–141, 1965.
2 Based on Hare WCD: The age at which epiphyseal union takes place in the limb bones of the dog, Wien Tierärztl Monatsschr 9:224–245, 1972.
3 Based on Smith RN: Fusion of ossification centers in the cat, J Small Anim Pract 10:523–530, 1969.
4 Based on Smith RN, Allcock J: Epiphyseal fusion in the Greyhound, Vet Rec 72:75–79, 1960.
5 Based on Sumner-Smith G: Observations on the epiphyseal fusion of the canine appendicular skeleton, J Small Anim Pract 7:303–311, 1966.
6 Based on Ticer JW: Radiographic Technique in Small Animal Practice, Philadelphia, 1975, Saunders, p 101.
From de Lahunta A, Habel RE: Applied Veterinary Anatomy, Philadelphia, 1986, Saunders.
A special position, in which the hindlimbs of the supine animal are rotated inward until the femoral trochleae and patellae face directly upward, is used for the better depiction of the contours of the femoral head when hip dysplasia is suspected. In this view it is easier to gauge the congruence of the femoral head with the acetabulum and to recognize any flattening or distortion of its contours. Progressive deformation of the head and worsening of fit characterize the progress of the condition.
The etiology of hip dysplasia, very common in certain larger breeds and with a familial tendency, is uncertain. Several once promising theories have been abandoned, and much work has concentrated on the belief that the dysplasia, which inevitably leads to osteoarthritic changes, is a consequence of the instability permitted by abnormally lax soft articular tissues. It has been shown that many affected dogs exhibit similar but milder features of the capsules of certain other joints. This prompts the suspicion that the dysplasia is not so much a unique affection of the hip but a particularly severe local manifestation of a more widespread developmental disorder.
The shaft of the femur is so deeply embedded among the muscles of the thigh that only a general impression of its presence may be obtained on palpation (Figure 17–2/9). Despite this protection, the femur is the most commonly fractured bone, and most breaks occur about or below midshaft level. Such fractures are often complicated by considerable overriding; the lower fragment is commonly displaced caudally by the pull of the gastrocnemius. They are often repaired by intramedullary pinning, a procedure usually requiring direct exposure of the break. A lateral approach is most convenient: after incision of the fascia lata, the biceps, whose cranial margin is often palpable through the skin, is reflected, which completes the exposure of the vastus lateralis; the path is now open to the bone along the attachment of the latter muscle (Figure 17–2/8,9,10).
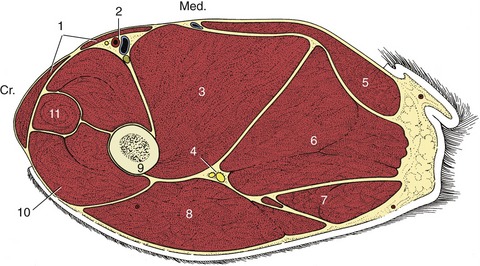
Figure 17–2 Transverse section of the canine left thigh. 1, Sartorius; 2, femoral vessels; 3, adductor; 4, sciatic nerve; 5, gracilis; 6, semimembranosus; 7, semitendinosus; 8, biceps; 9, femur; 10, vastus lateralis (of quadriceps); 11, rectus femoris.
Although the caudal thigh muscles appear to lend themselves to intramuscular injection, they should be avoided for this purpose because of possible damage to the sciatic nerve; a better alternative is injection into the muscles of the back.
The gluteal muscles have been described. Caudal to these, the cat presents the gluteofemoral, a long and relatively strong muscle that arises from the second to fourth caudal vertebrae and runs caudal to the superficial gluteal and cranial to the biceps to insert lateral to the patella in the fascia lata. It retracts the hindlimb and may also draw the tail to the side. The biceps femoris covers the abductor cruris caudalis, a small, thin muscle strap that emerges over the lateral head of the gastrocnemius in the lower leg.
The most important palpable structure of the thigh is the femoral artery (Figure 17–2/2), which is subcutaneous on the medial aspect of the limb toward the groin. It lies within the femoral triangle, a pyramidal space whose base lies toward the vascular lacuna (the passage to and from the abdomen for the femoral artery and vein) and whose tip is closed distally by the convergence of the sartorius and pectineus muscles that form its cranial and caudal walls. The pectineus forms so obtrusive a fusiform swelling that it immediately guides the fingers to the adjacent artery, which is the first choice for the evaluation of the circulation. Pulsation may still be perceived in a stretch of the artery after it dives more deeply among the muscles of the thigh. Its course leads it across the medial aspect of the femur to reach the popliteal fossa, where it is renamed the popliteal artery (see Figure 17–6/1,2). The accompanying vein is less conspicuous, but its constant relationship to the caudal border of the artery makes it easily found and convenient for intravenous injection in the supine, anesthetized subject. The saphenous artery (see Figure 17–6/4) branches from the concealed part of the femoral but soon becomes subcutaneous and runs over the medial aspect of the thigh toward the stifle. Both it and a large, more proximal branch (running caudally toward the gracilis) may be palpated.
Unlike the larger species, the dog and cat have no subiliac lymph nodes. However, the popliteal node is usually palpable within the popliteal fossa, between the distal parts of the biceps and semitendinosus as they diverge toward their insertions at the stifle (see Figure 17–3/10 and Figure 17–5/6).
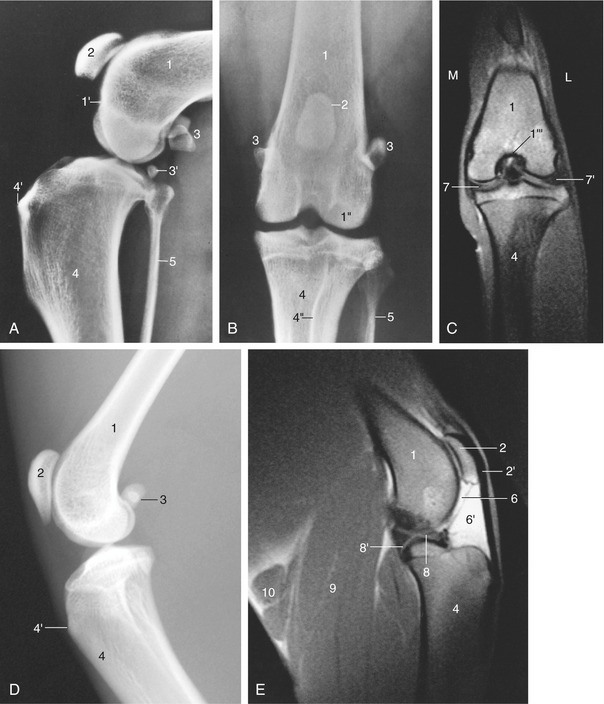
Figure 17–3 Lateral (A) and craniocaudal (B) radiographic views of the canine and feline (D, lateral only) stifles. C and E, Frontal and axial images of 4-mm-thick T1-weighted spin-echo magnetic resonance slices of the left canine stifle. 1, Femur; 1′, extensor fossa; 1″, lateral condyle; 1′″, intercondylar fossa; 2, patella; 2′, patellar ligament; 3, sesamoid bones in gastrocnemius; 3′, popliteal sesamoid bone; 4, tibia; 4′, tibial tuberosity; 4″, tibial crest; 5, fibula; 6, femoropatellar joint cavity; 6′, infrapatellar fat; 7, 7′, medial and lateral menisci; 8, 8′, cranial and caudal cruciate ligaments; 9, gastrocnemius; 10, popliteal lymph nodes.
THE STIFLE JOINT AND LEG
(See also pp. 91–93 and 97–99.)
The stifle joint is flexed in the standing posture. Although it is more fully extended in certain phases of locomotion, the femur and tibia are never brought into line, and the caudal angle of the joint does not open beyond 150° or so in dogs; considerably greater extension is permitted in the cat. Some lateral or medial angulation of the joint may often be observed when the limb is viewed from the front or behind. In the “bowlegged” version common in certain toy breeds, the pull of the quadriceps does not coincide with the axis of the femoral trochlea and there is a tendency to medial luxation of the patella. The luxation, which may be intermittent or permanent, causes the limb to be carried and, if left uncorrected, leads to deformity of other parts. The trochlea can be brought into alignment with the axis of the tibia by translation of the tibial tuberosity. The same procedure can be used to correct any medial or lateral patellar luxation, whatever its origin.
Palpation of the stifle joint reveals the following features of the skeleton: the patella; the ridges of the trochlea and the outer surfaces of the condyles of the femur; the sesamoid bones within the origin of the gastrocnemius; the head of the fibula; the edge of the lateral condyle adjacent to the fibula; the tuberosity; the extensor groove; and the medial surface of the tibia. The single patellar ligament and the medial and lateral collateral ligaments may also be distinguished; however, the femoropatellar ligaments cannot be distinguished because they are overlain by the aponeuroses of the sartorius and semimembranosus on the medial side and by that of the biceps laterally.
The most distinctive internal feature of the joint is the free communication of the various synovial compartments, which ensures that a single injection will reach all parts of the cavity. The most convenient entry is from the lateral side, caudal to the thick pad of fat interposed between the patellar ligament (and adjoining retinaculum) and the synovial membrane. The cruciate ligaments are set well back (see Figure 2–63/15,16). They assist the collateral ligaments in opposing rotation and medial or lateral deviation of the leg and are most susceptible to injury when tautened. The cranial cruciate ligament, named for the relative position of its tibial attachment (Figure 2–63/16), is therefore at greatest risk when strained in overextension of the joint; its rupture allows abnormally free forward displacement of the tibia in relation to the femur (the “cranial drawer” sign). A short cranial drawer movement (1 to 3 mm) brought to an abrupt stop is normal in young dogs. A deterioration in the strength of this ligament is correlated with age and is due to fiber bundle disruption and metaplastic cellular changes; the central part of the ligament is most affected. The changes are more pronounced and appear at earlier ages in larger dogs.
The caudal cruciate ligament is at greatest risk in the flexed position of the joint, and its rupture allows excessive caudal displacement of the tibia (the “caudal drawer” sign). Various surgical techniques for the restoration or replacement of these ligaments use fascial or artificial substitutes. The lateral collateral ligament can be used as a substitute for the cranial cruciate ligament after transposition of the head of the fibula cranially.
The lateral femorotibial joint has two pouches: one is under the tendon of the long digital extensor muscle at its origin from the extensor fossa, and the other invests the tendon of origin of the popliteal muscle, which contains a sesamoid bone close to the lateral tibial condyle.
The menisci, joined cranially by an intermeniscal ligament (also caudally, in the cat), provide additional restraints and are also prone to injury. They are most vulnerable when torsion is imposed on a limb in which the stifle is extended and the foot fixed—a combination of circumstances found when an abrupt change in direction is attempted by a dog traveling at speed. Most often a meniscal tear is found in combination with a rupture of the cranial cruciate ligament. The meniscal horns are more richly supplied with blood vessels and nerves in comparison with the more central parts. The involved portion, or sometimes the whole meniscus, is often removed after injury; an imperfect replacement may ultimately be formed from granulation tissue produced by the capsule at the site of the original attachment. The medial meniscus has an additional restraint imposed by a connection with the medial femorotibial ligament by way of the joint capsule; this may limit the range of its excursions when these are compared with those of its fellow and may thus be a factor in determining the incidence of injuries.
Both lateral and craniocaudal radiographic projections are commonly used in the diagnosis of stifle injuries (Figure 17–3). In the latter view the patella is superimposed on the distal end of the femur, where it is flanked by the ridges of the trochlea, which appear as thin radiodense lines. The tibial condyles are relatively flat because they are not separated by the conspicuous intercondyloid tubercles found in the larger species. The head of the fibula falls short of the extremity of the tibia. In the lateral view the femoral and tibial condyles are seen to have only limited, rather caudal contact and the joint appears unstable because the menisci that maintain its congruence are not revealed. The patellar ligament, the most prominent soft tissue shadow, runs at some distance from the femur, and the space behind it is occupied by the infrapatellar fat cushion. Displacement of this fat may be evident in radiographs and may suggest either thickening of the capsule or effusion into the joint cavity. The same view best depicts the associated sesamoid bones. The pair within the heads of the gastrocnemius are large and well defined (Figure 17–3/3); they articulate with small facets on the upper parts of the corresponding femoral condyles. That within the popliteus tendon is smaller, less sharply outlined, and occasionally duplicated; it is related to the margin of the tibia (Figure 17–3/3′). A relatively radiolucent area between the trochlea and lateral femoral condyle indicates the position of the extensor fossa (Figure 17–3/1′); it has occasionally been mistaken for an osteolytic lesion.
In dogs, both the distal femoral and proximal tibial epiphyses generally fuse with their respective shafts between the 6th and 12th months. The center for the tibial tuberosity fuses between the 8th and 10th months; while it persists, the cartilage line between it and the shaft is rather wide and irregular, presenting an appearance that simulates avulsion of the tuberosity. The onset and completion of these fusions are somewhat delayed in cats.
Few features of the leg require further comment. The subcutaneous surface of the tibia divides the cranial and caudal crural muscles medially, while the fibula makes the same division laterally (Figure 17–4). In lean dogs the fibula may be palpated along its length, but in fatter and particularly well-muscled animals only the head and the distal half of the shaft may be felt with certainty. The superficial flexor and gastrocnemius components of the common calcaneal tendon may be identified separately, distal to the belly of the latter. The lateral saphenous vein is a very conspicuous surface feature of the lateral aspect (Figure 17–5/9). It runs proximocaudally over the lower part of the leg before following the gastrocnemius on the caudal border to join the femoral vein within the popliteal fossa. The vein is much used for intravenous injections; its proximal part is better suited to this use because it is both relatively fixed and straight; the more mobile distal part undulates, dipping between the caudal crural muscles and the common calcanean tendon.
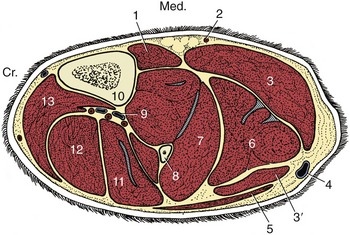
Figure 17–4 Transverse section of the canine left leg. 1, Popliteus; 2, saphenous artery; 3, 3′, medial and lateral heads of the gastrocnemius; 4, lateral saphenous vein; 5, biceps femoris; 6, superficial digital flexor; 7, deep digital flexor; 8, fibula; 9, cranial tibial vessels; 10, tibia; 11, peroneus longus; 12, long digital extensor; 13, tibialis cranialis.

Figure 17–5 Left canine hindlimb; the inset shows the actual appearance of the lateral saphenous vein (9); lateral view. 1, Patella; 2, patellar ligament; 3, peroneus longus; 4, tibialis cranialis; 5, biceps femoris; 6, popliteal lymph node; 7, common peroneal nerve; 8, lateral head of gastrocnemius; 9, lateral saphenous vein; 10, deep digital flexor; 11, superficial peroneal nerve; 12, calcanean tendon; 13, calcaneus; 14, peroneus longus tendon.
The vascularization of the leg and more distal parts depends on the cranial tibial and saphenous arteries because the caudal tibial is quite insignificant (Figure 17–6/3,4). The cranial tibial continues the popliteal artery, which runs deep to the popliteal muscle on the caudal aspect of the stifle. The artery then passes between the tibia and fibula in the proximal part of the leg before penetrating the dorsal muscles. It reappears toward the hock and then follows the long extensor tendon across the joint into the paw. The saphenous artery, which broadly serves the territory assigned to the caudal tibial in many species, crosses the medial aspect of the stifle before dividing into cranial and caudal branches. The cranial branch (Figure 17–6/6) remains superficial and continues into the paw, where it supplements the cranial tibial in supplying dorsal structures; the caudal branch (Figure 17–6/5) accompanies the tibial nerve and, after supplying caudal crural muscles, follows the flexor tendons into the plantar aspect of the paw.

THE HOCK AND HINDPAW
(See also pp. 90–91 and 93.)
Inspection of the distal part of the limb reveals the distinctive conformation of the hock; when it is taken apart, there is little external difference between the forepaws and hindpaws beyond the absence of any analogue of the carpal pad. A dewclaw is commonly present at birth in dogs but is routinely removed at an early age in puppies of many breeds. Duplication of this digit is known to occur in the French Shepherd breeds. Dewclaws of the hindlimb are not found in cats.
Although the hock skeleton is complete—there is no suppression or fusion of the standard elements—most bones cannot be individually identified on palpation. The most distinctive feature is the long, rather slender calcaneus, which provides the leverage for the effective extension of the hock; the arrangement carries an intrinsic risk, and the bone is occasionally fractured by the force exerted by the powerful muscles attaching to its slightly swollen tip. The calcaneus extends a medial process, the sustentaculum tali, over the plantar aspect of the talus, where it may be felt despite being covered by the deep flexor tendon (Figure 17–7/3′). The more distal tarsal bones do not have identifying surface features, but their locations and extents may be deduced after reference to a skeleton or to radiographs. The other prominent surface features of the region are the projections of the tibial and fibular malleoli at the lower limit of the leg and the equally prominent swellings at the proximal ends of the second and fifth metatarsal bones. A long collateral ligament may be traced from the malleolar to the metatarsal thickening on each side of the limb. The extensor tendons can be followed over the dorsal surface of the hock; the retinacula that hold them in place over the distal tibia and again at the proximal end of the metatarsus can also be appreciated in many dogs.

Figure 17–7 Dorsoplantar and lateral radiographic views of the canine (A and B) and feline (C and D) hocks and hindpaws. 1, Tibia; 2, fibula; 3, calcaneus; 3′, sustentaculum tali; 4, talus; 5, fourth tarsal bone; 6, central tarsal bone; 7, 7′, second and fifth metatarsal bones.
Only the tarsocrural joint is large enough to be punctured in the live animal. This is done on the lateral side just distal to the malleolus; the needle is directed distally toward the lateral surface of the palpable lateral trochlear ridge of the talus.
Similar impressions of the bones and soft structures are obtained as on a palpation of the hind as of the forepaw.
Although a complete radiographic examination of the hock calls for exposures in dorsoplantar, mediolateral, and oblique projections, the most useful general picture is obtained from the dorsoplantar view because it permits identification of all the bones, of which some are more easily identified than others as there is considerable superimposition (Figure 17–7, A). Both the talus and the calcaneus are well outlined despite the overlap of the sustentaculum tali. The two bones in the subjacent tier, the fourth and central tarsals (Figure 17–7/5,6), are also generally well outlined, although the mediodistal part of the fourth is superimposed on the third. The second tarsal is clearly shown with the smaller first tarsal superimposed on it. The distal extremities of the tibia and fibula appear closely related in this projection; the gap between them is unexpectedly wide in slightly oblique projections obtained of the cat’s hock and is a feature that is occasionally misinterpreted as evidence of luxation.
The lateral projection (Figure 17–7, B) depicts the calcaneus and talus clearly, although they overlap toward the center of the field. The more distal bones are less easily identified in this view, apart from the fourth, which is betrayed by a protuberance on its plantar aspect (Figure 17–8/4′). Because the central bone is occasionally dislocated, it is important to note the normal alignment of the dorsal borders of the bones of successive tiers. Two previously unrecorded sesamoid bones have recently been described in the Greyhound at the plantar aspect of the hock about the level of the tarsometatarsal joint. They appear to have the usual significance of such sesamoids; that is, they can potentially be misinterpreted as chips fractured from the major bones.

Figure 17–8 Lateral radiographic views of the canine (A) and young feline (B) hocks. 1, Tibia and fibula; 2, calcaneus; 2′, sustentaculum tali; 2″, coracoid process; 3, talus; 3′, trochlea of talus; 4, superimposed fourth and central tarsal bones; 4′, plantar tubercle on fourth tarsal bone; 5, distal row of tarsal bones; 6, metatarsal bones.
There are no distinctive features of the radiological anatomy of the metatarsal bones and phalanges. The short digital muscles are comparable with those in the front limb.
THE MAJOR NERVES OF THE HINDLIMB
It is only necessary to deal briefly with the course, relations, and distribution of those nerves that extend substantially into the free limb because a general account of the lumbosacral plexus (usually formed by the nerves L4–S2) and its divisions has been presented (pp. 323–325).
The femoral nerve (L4–L6) has a very short course within the thigh before it ends by ramifying within the quadriceps femoris, the principal extensor of the stifle and an ancillary flexor of the hip. Shortly before disappearing into this muscle, it detaches the saphenous nerve, which descends subcutaneously over the medial aspect of the limb accompanied by the palpable saphenous artery. Although the saphenous nerve supplies the sartorius, it is largely sensory, serving the skin of the medial surface of the thigh, stifle, leg, and hock (Figure 17–9). Dysfunction of the femoral nerve paralyzes the quadriceps, resulting in the collapse of the stifle and disabling the entire limb. Compensation is not available. The skin of the medial surface of the limb is deprived of sensation.

Figure 17–9 Autonomous zones of the cutaneous innervation on the lateral (A) and medial (B) surfaces of the canine hindlimb. 1, Caudal cutaneous femoral nerve (purple); 2, lateral cutaneous femoral nerve (green); 3, genitofemoral nerve (teal); 4, saphenous nerve (blue); 5, sciatic nerve (yellow); 6, peroneal nerve (orange); 7, tibial nerve (red). a, Position of greater trochanter; b, ischial tuber; c, lateral tibial condyle; d, medial tibial condyle.
The sciatic nerve (L6–S1) crosses the dorsal border of the hip bone to enter the limb together with the caudal gluteal vessels. After passing the dorsocaudal aspect of the hip joint deep to the greater trochanter, where it is susceptible to injury in trauma or surgery of the joint, the nerve and accompanying vessels supply branches to the hamstring muscles. The nerve then continues distally in a central position within the thigh, caudal to the femur and cushioned between the biceps laterally, the adductor, and later the semimembranosus medially (Figure 17–2/4). At a rather variable point it divides into common peroneal and tibial nerves that continue the course of the parent trunk until they diverge caudal to the stifle. The sciatic nerve and its peroneal and tibial branches collectively supply the skin of the entire limb distal to the stifle with the exception of the medial strip claimed by the saphenous.
The common peroneal nerve, the more lateral of the terminal divisions of the sciatic, can be palpated in lean dogs where it passes over the lateral head of the gastrocnemius (Figure 17–5/7). It then dives deeply among the dorsal crural muscles (the extensors of the digits and flexors of the hock), which it supplies. It is continued by superficial and deep (peroneal) branches that enter the paw over the dorsal aspect of the hock; they supply the skin of the dorsal surface. Paralysis of the common peroneal nerve produces slight overextension of the hock and inability to extend the digits, which may be rested on their dorsal surfaces. In time, affected dogs learn to flick their paws forward before putting them down, enabling their limbs to support weight. The dorsal surface of the paw is without sensation.
The tibial nerve passes between the two heads of the gastrocnemius, where it detaches branches to the muscles behind the tibia (the flexors of the digits and extensors of the hock). The depleted nerve, now largely sensory but retaining a small motor component for the intrinsic muscles of the paw, continues distally within the web of skin between the caudal crural muscles and the common calcanean tendon. It crosses the hock beside the deep flexor tendon before branching to supply the plantar structures of the paw. Tibial nerve injuries cause the hock to be flexed and lowered closer to the ground when the limb bears weight. The paralysis of the digital flexors elevates the toes; their plantar aspect is without sensation.