20 The Thorax of the Horse
CONFORMATION AND SURFACE ANATOMY
In the horse the difficulties in obtaining a reliable impression of the thoracic cavity from simple inspection of the exterior are increased by the height of the withers and the caudal prolongation of the rib cage. The narrow cranial part of the thorax is completely covered by the shoulder and arm. Some variation in the projection of the limb bones on the thoracic skeleton is due to the inconstant slope of the scapula. As a general guide, the caudal angle of this bone lies over the upper end of the seventh rib, while the supraglenoid tubercle projects in front of the first rib, a little above the manubrium of the sternum (Figure 20–1). The humerus forms a lesser angle with the horizontal than in the smaller species, and this brings the elbow within the skin of the trunk. The precise position of the elbow joint is not immediately apparent but may be inferred from its relation to the olecranon whose summit (point of elbow) lies over the lowest part of the fifth rib or succeeding intercostal space. The triangle between the scapula and humerus is completely occupied by the massive triceps muscle, which severely restricts clinical access to the cranial part of the thorax.
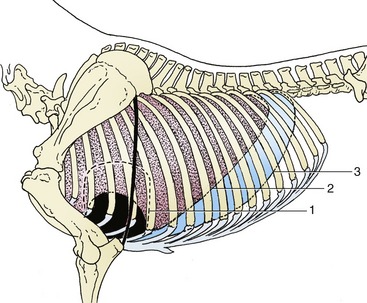
Figure 20–1 Projections of the heart and lung on the left thoracic wall. The heavy line indicates the caudal border of the triceps. 1, Outline of heart; 2, basal border of lung; 3, line of pleural reflection.
There are 18 pairs of ribs. Those behind the triceps, that is, those from the seventh rib caudally, are individually identifiable on palpation even though they are covered in varying degree by certain muscles: cataneus trunci, latissimus dorsi, serratus ventralis, and obliquus externus abdominis. The most caudal ribs may even provide visible landmarks; this is most often true of the upper part of the last rib, which prominently marks the cranial limit of the flank. Palpation of the ribs reveals their changing orientation. The last two or three, which are relatively short, have a pronounced caudal inclination; the half-dozen or so (R9–15) in front of these are longer and of equal length and curvature. The more cranial ribs are both shorter and less strongly curved. The first rib, the shortest of all, is almost vertical. The increasing slope of the ribs as the series is followed caudally brings the last rib remarkably close to the coxal tuber (see Figure 19–1).
Between the forelimbs the thorax is covered by the powerful pectoral muscles that form paired swellings separated by a prominent groove along the line of the sternum (see Figure 23–4). The cranial part of this bone, the manubrium, projects as a readily found landmark. The caudal xiphoid process is also palpable, though it is not quite so easily found as the manubrium; it is broad and flexible and is enclosed between the converging costal arches. External inspection fails to suggest the tilt of the sternum, which slopes upward toward the manubrium; this, in combination with the ventral slope of the cranial thoracic vertebrae, reduces the depth of the cranial part of the thoracic cavity.
An exact appreciation of the position of the diaphragm is essential for the clinician. The vertex is level with the sixth intercostal space (or even the sixth rib) and thus comes to within a short distance of the point of the elbow in an animal standing square (Figure 20–3). The inexperienced find it particularly hard to accept this crucial fact.
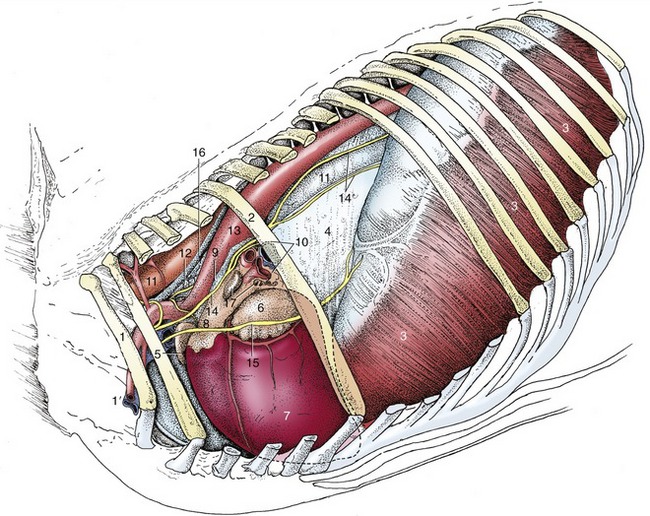
Figure 20–3 Structures within the mediastinum. The mediastinal pleura cranial to the heart has been removed, which exposes the cranial lobe of the right lung. 1, First rib; 1′, axillary vessels; 2, sixth rib; 3, diaphragm; 4, caudal mediastinum covering right lung; 5, right auricle; 6, left auricle; 7, left ventricle; 8, pulmonary trunk; 9, ligamentum arteriosum; 10, root of lung; 11, esophagus; 12, trachea; 13, aorta; 14, vagus; 14′, dorsal and ventral vagal trunks; 15, phrenic nerve; 16, thoracic duct; 17, tracheobronchial lymph nodes.
There are naturally considerable breed and individual variations in conformation. Without considering these in detail, it may be said that a deep chest is generally favored. In saddle horses it is desirable that the ribs slope caudally without excessive lateral bowing because too pronounced a “barrel” makes for an uncomfortable seat.
THE THORACIC WALL
Removal of the forelimbs exposes the contrasting form of the cranial and caudal parts of the thorax. The cranial part (formed by the sternal ribs) is narrow and bilaterally compressed and shows little movement; the caudal part (formed by the asternal ribs) is conspicuously wider and more rounded and makes a substantial contribution to the respiratory excursions (see Figure 20–8). In comparison with the bovine chest, the ribs are narrow and the intercostal spaces markedly wide, especially in their ventral parts. The arrangement of the structures within the spaces follows the usual pattern.
The short, stout first rib is almost immobile, as it is stabilized by tight joints with the vertebral column and sternum and by anchorage to the cervical vertebrae through the scalenus. The brachial plexus divides this muscle into ventral and (small) middle parts, while the axillary vessels emerge ventral to it. These vessels wind around the cranial margin of the first rib, where the artery may be palpated against the bone. Previously, the artery was punctured at this site when a sample of arterial blood was required (Figure 20–3/1′), but currently the carotid artery is preferred.
In conformity with the length of the thorax the diaphragm is more oblique than in other domestic species. It has the same general form and bulges forward from its peripheral attachments to the lumbar vertebrae, ribs, and sternum. Its most cranial part, the vertex, is situated directly above the sternum and, as already emphasized, projects on the lower part of the sixth space or preceding rib. The dorsal part of the diaphragm is molded to present right and left elevations between which the median portion is retracted by the crura to form a recess. The middle and ventral parts are uniformly curved from side to side. The openings within the diaphragm show no important specific features (Figure 20–4).
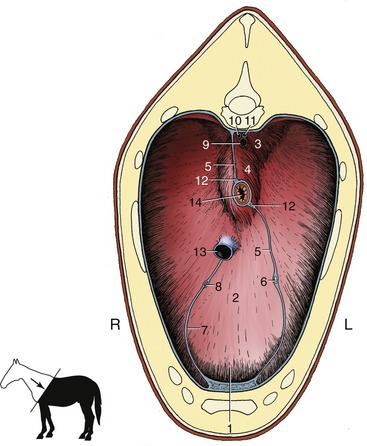
Figure 20–4 Cranial surface of the diaphragm. 1, Sternal and costal parts of diaphragm; 2, tendinous center; 3, left crus; 4, right crus; 5, caudal mediastinum; 6, left phrenic nerve; 7, plica venae cavae; 8, right phrenic nerve; 9, aorta; 10, right azygous vein; 11, thoracic duct; 12, dorsal and ventral vagal trunks; 13, caudal vena cava; 14, esophagus.
THE PLEURAL CAVITIES
The arrangement of the pleura follows the usual pattern in that the thoracic interior is divided into two pleural cavities by an intermediate septum, the mediastinum. The subpleural connective tissue is poorly developed; as a result, the mediastinum is weak.
The projection of the pleural cavities on the chest wall is always a matter of clinical significance. The mediastinal pleura is reflected onto the thoracic wall within the costovertebral gutter, and the costal pleura thus extends above the ventral border of the vertebral bodies; the ventral limit of the costal pleura follows an irregular line that passes over the costal cartilages. Cranially, the pleural sac extends medially to the first rib and beyond this on the right side where an outpouching (cupula pleurae) passes several centimeters into the neck; this prolongation of the right sac is of potential importance because it may be punctured by penetrating wounds that appear to spare the thorax. The caudal reflection of the costal pleura onto the diaphragm has an unusual line. It begins at the vertebral end of the seventeenth rib and is then deflected caudally to reach the middle of the last rib before turning forward. It then follows a more conventional course that intersects successive ribs at progressively lower levels until it continues along the eighth rib cartilage to the sternum. This line traces a slight dorsocranial concavity (see Figures 20–1 and 20–2/3).
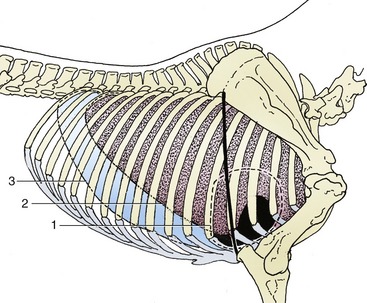
Figure 20–2 Projections of the heart and lung on the right thoracic wall. The heavy line indicates the caudal border of the triceps. 1, Outline of heart; 2, basal border of lung; 3, line of pleural reflection.
As always, the pleural cavities are considerably larger than the lungs, even when there is maximal inflation. There thus exist potential spaces along the ventral and caudal margins of the lung that are never utilized. The breadths of these spaces (the costomediastinal and costodiaphragmatic recesses) vary with the phase of respiration. The costodiaphragmatic recess lies over the intrathoracic part of the abdomen and provides a potential route for the puncture of certain abdominal organs. Obviously the risk of injury to the lung is minimized if the needle is introduced during full expiration (see Figure 20–8/13′).
THE LUNGS
The lungs are elongated and shallow, corresponding to the general form of the pleural cavities. The right and left lungs are more nearly equal in size than in other species (Figure 20–5), and because the difference lies mainly in the greater thickness of the right lung, the asymmetry that does exist may easily escape notice (Figure 20–6). There is no external evidence of lobation other than the presence of the accessory lobe appended to the base of the right lung. However, the cranial part of each lung is somewhat separated from the caudal mass by a relatively attenuated region (see Figures 4–24 and 20–1). The two lungs are extensively joined by connective tissue caudal to the bifurcation of the trachea.
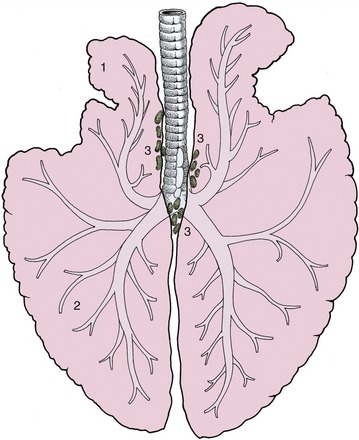
Figure 20–5 Dorsal view of the lungs and bronchial tree, schematic. 1, Apex (cranial lobe) of left lung; 2, base (caudal lobe) of left lung; 3, tracheobronchial lymph nodes.
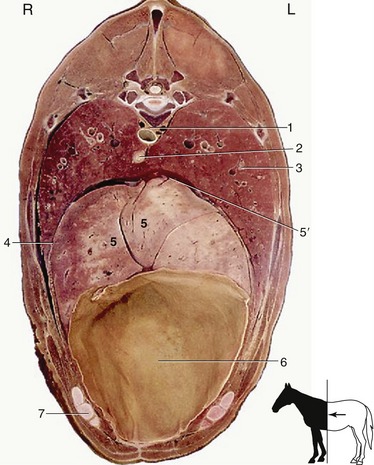
Figure 20–6 Transverse section of the trunk at the level of T12 and the middle of the ninth rib. 1, Aorta; 2, esophagus; 3, lung; 4, diaphragm; 5, liver; 5′, caudal vena cava; 6, diaphragmatic flexure of the ascending colon; 7, costal arch.
The left lung exhibits a deep cardiac notch that allows the pericardium extensive contact with the chest wall between the third and sixth ribs (see Figure 20–1). The notch is margined by a thinned region so that the lung provides little cover to the pericardium over a much larger area (Figure 20–7). The arrangement on the right side is similar, although the asymmetry of the heart reduces the size of the cardiac notch, which extends from the third rib to the fourth intercostal space (see Figure 20–2). When moderately expanded, the base of each lung reaches to a line passing through the upper part of the sixteenth, the middle of the eleventh, and the costochondral junction of the sixth rib; the upper part of this line is almost vertical, and the lower part sweeps cranioventrally. This margin of the lung is separated from the line of pleural reflection by about 5 cm dorsally and ventrally but by as much as 15 cm in its middle part (see Figures 20–1 and 20–2). In young foals the extent of the lung is more restricted and the caudal limit is at about the thirteenth rib.
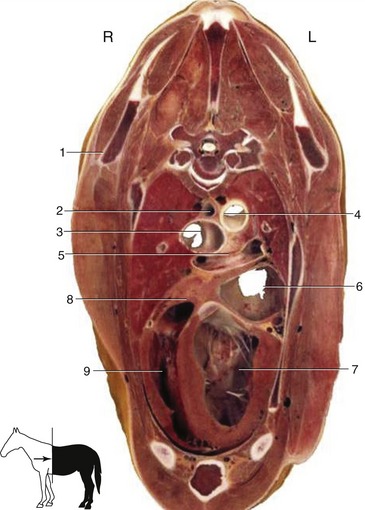
Figure 20–7 Transverse section of the thorax at the level of T5. 1, Caudal angle of scapula; 2, esophagus; 3, bifurcation of the trachea; 4, aorta; 5, bifurcation of the pulmonary trunk; 6, left atrium; 7, left ventricle; 8, right atrium; 9, right ventricle.
The projection of the lung on the chest wall is considerably larger than the clinically useful area for percussion and auscultation, as examination of the thin margins of the lung will not provide useful information. The area for such examination is triangular and is defined by the caudal angle of the scapula, the point of the elbow, and the upper end of the sixteenth rib. Two sides of this triangle are more or less straight, but the caudoventral side, the hypotenuse, is slightly bowed.
Tapping of pleural fluid is most safely performed in the lower part of the seventh intercostal space, ventral to the margin of the lung. Care is required to avoid puncturing the superficial thoracic (“spur”) vein that crosses the site (see Figure 23–3/11″).
The lobulation of the lungs is not obtrusive but can be detected on careful examination of the expanded lung. It is less obvious in the collapsed state when the covering pleura is wrinkled. It is also evident on section. However, it is accepted that the septa are incomplete and that the possibility of collateral ventilation between neighboring lobules exists.
The chief bronchus, the pulmonary artery, and the pulmonary vein combine to form the root of the lung before entering at the hilus in a region deprived of pleura and directly adherent to the same part of the other lung. The chief bronchus separates within the lung into a small cranial division that passes toward the cranial lobe and a larger caudal division that attends to the ventilation of the remainder of the organ. There are difficulties in homologizing the bronchi of lower orders with those in other domestic species, but at the present time these details are not of great importance; lung surgery is rarely performed in horses.
In standing animals the ventilation and perfusion of different regions and lobes of the lungs are reasonably well matched, although in larger species, such as the horse, there must be some tendency for gravity to favor the perfusion of more ventral parts. The spatial relationship of ventilation and perfusion is disturbed in animals placed in dorsal or lateral recumbency, and the disturbance becomes significant when the recumbent posture is long maintained—as during major surgery. In these circumstances there is compression of whichever part of the lung is at the bottom. This reduces the tensile forces that ordinarily hold airways open in that part of the lung. The ensuing airway closure permits complete collapse of the alveoli served by such airways; blood perfusing these alveoli cannot take part in respiratory gas exchange.
The pattern of division of the pulmonary artery corresponds to that of the bronchi. A separate bronchial artery attends to the supply of the bronchial and peribronchial tissue, but the blood is returned by the single set of pulmonary veins.*
The lymphatic drainage leads first through very small pulmonary nodes embedded in the substance of the organ and then to larger tracheobronchial nodes about the bifurcation of the trachea (Figure 20–3/17). From here most lymph is drained via the cranial mediastinal nodes.
The nerves that enter at the hilus derive from the pulmonary plexus to which both sympathetic and parasympathetic fibers contribute.
THE MEDIASTINUM
The heart divides the mediastinum into the familiar parts (Figure 20–8/4,4′).
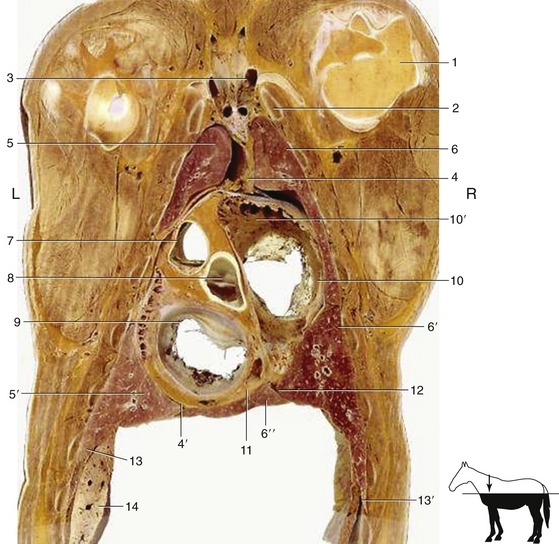
Figure 20–8 Dorsal section of the thorax at the level of the atrioventricular valves. 1, Head of humerus; 2, first rib; 3, formation of cranial vena cava; 4, 4′, cranial and caudal mediastinum; 5, 5′, cranial and caudal lobes of the left lung; 6, 6′, 6″, cranial, caudal, and accessory lobes of the right lung; 7, pulmonary valve; 8, aortic valve; 9, left atrioventricular valve; 10, right atrioventricular valve; 10′, right auricle; 11, coronary sinus; 12, plica venae cavae; 13, diaphragm; 13′, costodiaphragmatic recess; 14, part of the liver.
The cranial part is markedly asymmetrical; it attaches to the left first rib and gradually shifts to reach a more or less median situation directly in front of the heart. The dorsal part is thick, the ventral part much thinner, especially after the thymus has regressed. The dorsal part occupies about half the transverse diameter of the thorax and includes the esophagus and trachea, the brachiocephalic trunk and cranial vena cava with their respective branches and tributaries, the cranial mediastinal lymph nodes, the thoracic duct, and the phrenic, vagus, and sympathetic nerves. The interstices between these structures are occupied by fat, sometimes present in large amounts. The thymus is the sole content of the ventral portion.
The ventral part of the middle mediastinum is very broad because it contains the heart and pericardium (Figure 20-7). The dorsal part is paper-thin except where it contains the esophagus, the continuation of the trachea to its bifurcation, the aorta, and certain nerves (including vagal branches).
In lateral view the caudal mediastinum is triangular (Figure 20–3/4). It is divided into two parts by adhesion between the lungs about and caudal to their roots. The ventral part, whose sole occupant is the left phrenic nerve, is diverted far to the left before it merges with the pleura covering the diaphragm (Figure 20–4/6). The dorsal part is thin except where it encloses the esophagus and aorta.
Except in foals, small openings in the mediastinum place the two pleural cavities in communication. The mediastinum is very fragile, and exposure during dissection inevitably increases the number of visible openings, which leaves it unclear whether any were present when the thorax was intact and suggests that the mediastinum might be an ineffectual partition. However, small openings in the thoracic wall such as are made for the purpose of thoracoscopy (when the influx of air can be controlled) result in incomplete unilateral pneumothorax and are survived without obvious adverse effects.
THE HEART
The heart lies in the ventral part of the middle mediastinum, directly cranial to the diaphragm and largely covered by the forelimbs (see Figure 20–1). It forms an irregular and laterally compressed cone; the larger part lies left of the median plane and is so disposed that the axis slopes caudoventrally and to the left (see Figure 20–3). There is significant variation in heart size: that of a Thoroughbred is conspicuously larger, both relatively and absolutely, than that of other horses of comparable body weight. The difference is mostly inherited, and partly conditioned by training. Such variation inevitably affects the topography. Most commonly the heart extends between the planes of the second to sixth intercostal spaces, which places the apex directly caudal to the level of the point of the elbow. The cranial margin is strongly curved and is arranged with its upper part vertical; its lower part follows the dorsal surface of the sternum. The caudal border, though sinuous in profile, is more or less upright (see Figure 20–3). The flattened lateral surfaces are related through the pericardium to the mediastinal surfaces of the lungs, except where the cardiac notches allow direct contact with the thoracic wall; as already stated, this contact is greater on the left side. A strong sternopericardiac ligament attaches the pericardium to the sternum, and this, with the anchorage of the great vessels, limits the displacement allowed to the heart. A slight shift, however, does occur with the movement of the diaphragm.
Apart from the general form there is little of significance to distinguish the heart of the horse. Mention should be made, however, of two features of the aortic and pulmonary valves, especially the former. The cusps commonly develop nodules at the free margins, and these can be quite striking in older animals; in addition, fenestrations may appear in the middle region of the cusps. Neither development appears to have much, if any, functional significance. The puncta maxima, the sites at which the valve sounds are most clearly heard, do not correspond exactly to the projections of the openings on the chest wall. The left atrioventricular valve is auscultated to most advantage in the fifth intercostal space, a little caudodorsal to the point of the elbow; the aortic valve at a somewhat higher level in the fourth space; and the pulmonary valve lower within the third space—all of course on the left side. The right atrioventricular valve is best heard in the lower parts of the third and fourth right intercostal spaces. These directions are perhaps overprecise as the skeletal topography is not always easy to appreciate in practice. It is perhaps more useful to be aware that the puncta lie within a band of a few centimeters’ depth about midway between the horizontal planes that intersect the points of the shoulder and the elbow. Within this band the punctum maximum of the left atrioventricular valve is at the intersection of the vertical line that falls a couple of fingerbreadths behind the point of the elbow. The approach to the other valves follows from the relative positions indicated and requires the introduction of the stethoscope between the limb and the chest wall.
The coronary arteries share the supply of the heart wall in more equal fashion than in many other species because the right one ends by descending within the right (subsinuosal) interventricular groove (see Figure 7–19/2′).
THE ESOPHAGUS, TRACHEA, AND THYMUS
Although the esophagus still lies partly to the left on entering the chest, it quickly regains a position dorsal to the trachea; thereafter, it pursues a median course, apart from slight deflections as it passes the aortic arch and again just before the esophageal hiatus. The striated muscle of the cranial part of the esophagus is gradually replaced by smooth muscle as the heart is approached; the color change makes the transformation obvious. The muscle is somewhat thicker immediately before the diaphragm, and this part of the tube is commonly contracted in the dead specimen. There is no evidence that the diaphragm embraces the esophagus tightly at the hiatus, as sometimes alleged. Indeed the free movement of the diaphragm over the esophagus is facilitated by the peritoneum pouching through the hiatus on the right and ventral side of the esophagus.
The trachea becomes median soon after entering the thorax. It then lies against the longus colli muscles but soon diverges to run lower within the mediastinum. After passing over the left atrium, it bifurcates at about the level of the fifth rib (or space) (Figure 20–7/3). The bifurcation is not symmetrical; the right bronchus is larger.
The thymus is prominent in early life but soon regresses. Its formation from right and left parts is not obvious because they are closely applied together. In the young foal it completely fills the ventral part of the mediastinum cranial to the heart and may even extend over the left side of the pericardium; part may also pass into the neck beside the trachea, very occasionally reaching the thyroid gland. At this stage the thymus is clearly lobulated and bright pink. It is largest about 2 months after birth and thereafter regresses, although the rate is variable. Usually little remains after 3 years, when the vestige consists largely of fatty fibrous tissue. At its apogee the thymus makes contact with most structures within the cranial mediastinum.
THE GREAT VESSELS AND NERVES WITHIN THE THORAX
The pattern of arterial branching is shown in Figure 7–37 and need not be further described because the details are altogether without clinical significance. Rupture of the aortic wall in the sinus region or at the origin of the brachiocephalic trunk is not too uncommon in conditions of stress; the resulting hemorrhage is rapidly fatal. It appears to indicate inherent weakness at these sites because pathological change is rarely evident.
The presence of a single right azygous vein may be used to distinguish the equine from the bovine heart.
The formations, the courses, and the ramifications of the phrenic, sympathetic, and vagus nerves conform to the usual patterns; none are of great practical importance. The relationship of the left recurrent laryngeal nerve to the aortic arch, though not specific to the horse, deserves emphasis since intermittent stretching of the nerve with the pulsation of the vessel has been postulated as a factor in the etiology of laryngeal hemiplegia (roaring). The closer association of the left nerve to the tracheobronchial lymph nodes is a second factor of alleged but unproven significance (p. 526).
THE LYMPHATIC STRUCTURES OF THE THORAX
There are very numerous lymph nodes within the thorax. Although most are collected in groups, these are sometimes less discrete than is often suggested, and the provenance of the more scattered nodes may be difficult to determine. The following are the principal groups.
Small intercostal nodes lie in the dorsal part of some intercostal spaces. They receive lymph from the vertebrae and the adjacent muscles, the dorsal part of the diaphragm, and the local costal and mediastinal pleura. The efferent flow is to the thoracic duct.
The cranial mediastinal nodes are numerous and scattered about the esophagus, trachea, and vessels at the entrance to the thorax; usually some form a discontinuous chain that joins the caudal deep cervical nodes within the neck. The most caudal members reach the pericardium, where they overlap the nodes about the tracheal bifurcation that are assigned to the tracheobronchial and caudal mediastinal groups. Most efferent vessels pass to the thoracic duct; those from the most cranial nodes in the series may first perfuse deep cervical nodes.
The tracheobronchial group is scattered about the caudal part of the trachea and the chief bronchi (Figure 20–5/3); left, middle, and right subdivisions are commonly distinguished. Small nodes within the peribronchial tissue of the lung may be regarded as members of this series. Most lymph passing through this group has origins within the lungs, but some comes from the pericardium, the heart, and the caudal mediastinal nodes. The efferent vessels are divided between those that go directly to the thoracic duct and those that first perfuse the cranial mediastinal nodes.
A number of small caudal mediastinal nodes lie directly in front of the diaphragm and between the esophagus and aorta. Lymph is received from the esophagus, the diaphragm, the liver, the mediastinal and diaphragmatic pleura, and, apparently, the lungs. The efferent lymph flow is divided between the thoracic duct and the tracheobronchial and cranial mediastinal lymph nodes.
The few ventral mediastinal lymph nodes are without significance.
The thoracic duct exhibits no important distinctive features. It drains into one or another of the large veins at the entrance to the thorax, most commonly the cranial vena cava.