24 The Hindlimb of the Horse
CONFORMATION OF THE CROUP
Although the hindlimbs support little more than 40% of the body weight, they supply by far the greater part of the forward impetus in locomotion. This thrust is delivered through the hip and sacroiliac joints, which are intrinsically more stable than the shoulder and scapulothoracic synsarcosis, the corresponding “joints” of the forelimb. The sacroiliac joint is strengthened by tight ligaments, and both it and the hip joint are well supported by the muscles of the croup and thigh. These muscles are particularly massive in the horse, in which they round the contours in a distinctive fashion. In consequence, it is more difficult to appreciate the features and orientation of the pelvis of the horse than those of the pelves of other domestic species.
Despite the muscular development, the coxal tuber remains a conspicuous landmark that is palpable in its whole extent and visible in its upper part (Figure 24–1/2). The sacral tuber (Figure 24–1/2′), difficult to palpate in most animals, rises a little above the level of the adjacent spinous processes. The ischial tuber (Figure 24–1/3) is also not always easy to appreciate, although its location and a general impression of its form may be obtained on deep palpation over the muscles that form the caudal contour of the croup and thigh. If this is done, the slope of the pelvis may be estimated by visualizing the line joining the coxal and ischial projections. In the standard, generally approved conformation, this line forms an angle of about 30° with the horizon; from this it may be inferred that the sacrum is more or less horizontal. When the angle is significantly smaller—and the two tubers come close to sharing the same horizontal plane—the tail appears to be set high. When the angle is significantly greater, the animal is said to be goose-rumped. The croup is short in such animals, and the hamstring muscles are reduced in length and in the leverage they may exert. Although this is clearly disadvantageous, some compensation is obtained from the more stable support the limbs afford the trunk, and many horsemen and horsewomen find a gently sloping croup acceptable in a saddle horse. Undue prominence of the sacral tubers (“hunter’s bumps”) sometimes develops, especially in show jumpers and other horses subjected to similar repeated stress. The deformity is commonly ascribed to subluxation of the sacroiliac joints.
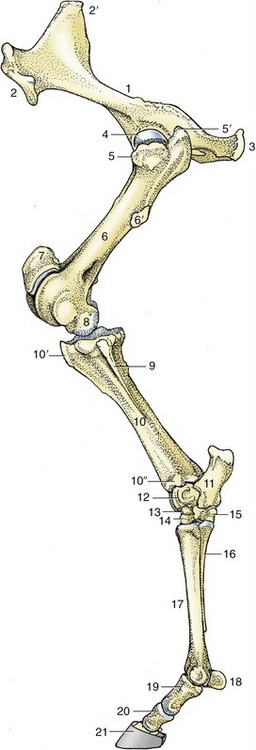
Figure 24–1 The skeleton of the left hindlimb, lateral view.
1, Hip bone (os coxae); 2, coxal tuber; 2′, sacral tuber; 3, ischial tuber; 4, head of femur; 5, 5′, cranial and caudal parts of greater trochanter; 6, femur; 6′, third trochanter; 7, patella; 8, femoral condyle; 9, fibula; 10, tibia; 10′, tibial tuberosity; 10″, lateral malleolus; 11, calcaneus; 12, talus; 13, central tarsal; 14, third tarsal; 15, fourth tarsal; 16, metatarsal IV (lateral splint bone); 17, metatarsal III (cannon bone); 18, proximal sesamoid bones; 19, 20, 21, proximal, middle, and distal phalanges, the last within the hoof.
The position of the hip joint cannot be determined directly but may be deduced from its relationship to the greater trochanter of the femur. This protuberance is divided into low cranial and high caudal parts, separately identifiable on palpation (Figure 24–1/5,5′). At more distal levels, the third trochanter (prominent only in this species) and the lateral epicondyle are easily distinguished and may be used to reveal the orientation of the femur. This bone is more nearly vertical than is often supposed (see Figure 19–1).
THE HIP JOINT
The stability of the hip joint owes much to the depth and extent of the acetabulum, which is considerably increased by a fibrocartilaginous rim; it embraces a large part of the femoral head (Figure 24–1/4). The head is additionally secured against luxation by two ligaments. One, the ligament of the femoral head, is short and stout but is not peculiar in any important way. The other, the accessory ligament, is unique to the horse (and donkey) among domestic species. It begins as a detachment from the prepubic tendon and reaches the joint by following a shallow groove on the ventral aspect of the pubis; this leads it to the acetabular notch through which it passes to insert on the head (see Figure 21–2/5′). The two ligaments together restrict both the range and the versatility of movement permitted to the joint. The restrictions on rotation and abduction are most severe; in practice, movement is almost confined to flexion and extension in a sagittal plane, which is a much more limited repertory than the geometry of the articular surfaces suggests. The stability of the joint is partly dependent on the tension exerted by the weight of the abdominal viscera pulling on the prepubic tendon and thus on the accessory ligament (p. 547).
Although the joint capsule is quite capacious, its deep location makes it relatively difficult to access. When it must be punctured, the needle is introduced between the two parts of the greater trochanter and is directed horizontally and craniomedially, at an angle of about 40° to the transverse plane.
THE MUSCLES OF THE HIP AND THIGH
These are conveniently regarded as comprising gluteal, hamstring, medial, and cranial groups.
THE GLUTEAL MUSCLES
The superficial and deep fasciae of the croup and thigh continue the corresponding coverings of the loins. The deep fascia detaches various septa that find anchorage on the pelvic girdle and the caudal edge of the sacrosciatic ligament after passing between certain muscles. The most substantial of these separate the gluteus superficialis and biceps femoris, the biceps and semitendinosus, and the semitendinosus and semimembranosus, thus molding the muscles so that their individual contours are often clearly visible through the skin; this is especially so in animals in “hard” training and when the muscles are contracted. The inner surface of this fascia itself, including the sides of the septa, gives origin to many fascicles of the muscles it covers.
The tensor fasciae latae (Figure 24–2/3) radiates from its origin on the coxal tuber to end by a broad aponeurosis (fascia lata) that inserts on the patella, the lateral patellar ligament, and the cranial border of the tibia. The cranial border of the fleshy part is related to the subiliac lymph nodes. The tensor is a flexor of the hip that helps to advance the limb during the swing phase of the stride. It is supplied by the cranial gluteal nerve.
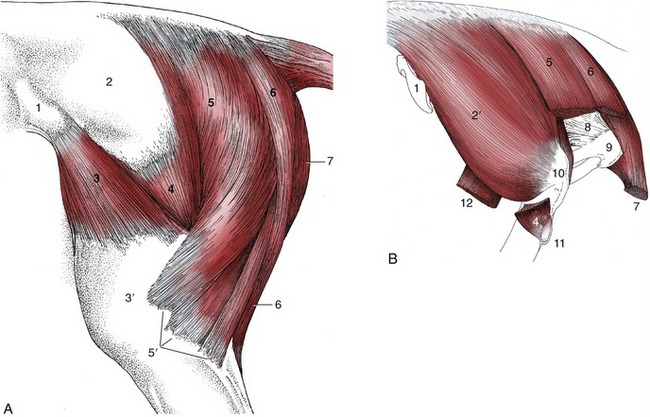
Figure 24–2 A, Muscles of the croup and thigh, lateral view. B, Croup muscles, resected to expose the ischial tuber, lateral view.
1, Coxal tuber; 2, deep gluteal fascia; 2′, gluteus medius; 3, tensor fasciae latae; 3′, fascia lata; 4, gluteus superficialis; 5, vertebral head of biceps; 5′, the three distal divisions of the biceps; 6, semitendinosus; 7, semimembranosus; 8, sacrosciatic ligament; 9, ischial tuber; 10, caudal part of greater trochanter; 11, third trochanter; 12, stump of rectus femoris.
The gluteus superficialis lies between the tensor and biceps (Figure 24–2/4). It has separate origins from the coxal tuber and the gluteal fascia, but the two parts combine at a common insertion on the third trochanter. Occasionally, this projection is broken off, and it is then displaced dorsally by the attaching muscle. The gluteus superficialis is potentially a flexor of the hip and abductor of the thigh. Its two parts are separately supplied by the cranial and caudal gluteal nerves.
The gluteus medius is a muscle of exceptional size and power (24–2, B/2′). Its wide origin spreads from a depression scooped in the surface of the longissimus dorsi, over the coxal tuber and iliac wing, to the sacrum and adjacent part of the sacrosciatic ligament. The principal insertion is to the caudal part of the greater trochanter, but a deep division—gluteus accessorius—has a separate aponeurotic attachment to the intertrochanteric line of the femur. This aponeurosis passes over the cranial part of the trochanter, where its passage is eased by the interposition of a synovial (trochanteric) bursa. This bursa may become inflamed, which is a condition detectable by the animal’s flinching when pressure is exerted over it. Horses so afflicted obtain relief by standing with the affected limb somewhat abducted; when they move, they tend to adopt an oblique doglike gait, swinging the limb in an arc.
This muscle is primarily an extensor of the hip, but it has a secondary use as an abductor of the thigh. Its association with the longissimus dorsi makes it an effective participant in rearing. It is supplied by the cranial gluteal nerve.
The gluteus profundus lies deep to the caudal part of the gluteus medius. It arises from and around the ischial spine and passes more or less transversely to insert on the cranial part of the greater trochanter. An abductor of the thigh, it is supplied by the cranial gluteal nerve.
THE CAUDAL (HAMSTRING) MUSCLES
In the horse, the three muscles of this group possess well-developed vertebral heads of origin (in addition to the usual pelvic heads); it is these vertebral heads that account for the characteristic filling and rounding of the croup (Figure 24–2/5,6). The vertebral head of the biceps arises from the sacrum and adjacent part of the sacrosciatic ligament. It descends behind and partly covers the gluteal muscles before it crosses the ischial tuber to be joined by the smaller pelvic head that arises from that process. The muscle inserts by three divisions (Figure 24–2/5′): the first in the fascia lata and on the patella, the second on the lateral patellar ligament and tibial crest, and the third, the tarsal tendon, on the common calcanean tendon. The vertebral head is supplied by the caudal gluteal nerve, and the pelvic head is supplied by the sciatic nerve.
The vertebral head of the semitendinosus (Figure 24–2/6) has an origin adjoining that of the biceps. After this merges with the pelvic head, the combination edges medially to insert on the medial aspect of the tibia and the crural fascia. It also detaches a tarsal tendon that joins the common calcanean tendon. The vertebral and pelvic heads are supplied by the caudal gluteal and sciatic nerves, respectively.
The semimembranosus (Figure 24–2/7) is included in the hamstring group, although topographically it is a muscle of the medial aspect of the thigh. The vertebral head is relatively weak, and the pelvic head is more substantial. The combined muscle is largely covered by the gracilis and follows the caudal margin of the adductor to which it is closely bound. It inserts by two divisions. The cranial division inserts on the medial epicondyle of the femur and the medial collateral ligament of the stifle joint; the caudal division proceeds distally to the medial condyle of the tibia. The principal nerve supply is from the sciatic nerve.
The actions and uses of the hamstring muscles are complicated and in certain respects enigmatic. It is clear that all three units are well placed to extend the hip. Rather than looking at the three muscles individually when considering the actions on the stifle, one will find it more useful to divide the hamstring group into two functional units: one that inserts proximal to the axis of rotation of the joint and the other distal to it. The “proximal unit” comprises parts of the muscles that are potentially extensor, as they may straighten the stifle by drawing the femur caudally when the limb bears weight. The “distal unit” will flex the stifle when the hoof is raised from the ground but will extend it when the hoof is firmly planted. The contributions of the biceps and semitendinosus to the common calcanean tendon must not be forgotten; these parts are extensor to the hock.
Some of these actions are clearly incompatible because the movements of the stifle and hock joints are linked in their actions by the reciprocal mechanism (see p. 638). It follows that the entire hamstring group, which includes parts that may flex the stifle, cannot always contract en masse.
THE MEDIAL MUSCLES
The medial muscles are disposed in the same three layers as in other species. The superficial layer comprises the gracilis and sartorius (Figure 24–3/8,14). The gracilis exhibits no specific features that require notice. The sartorius arises from the psoas fascia and the insertion tendon of the psoas minor and gains the thigh by passing through the gap between the caudal margin of the flank and the ilium. It is related to the deep inguinal lymph nodes, where it forms the cranial margin of the femoral triangle. The sartorius inserts on medial structures of the stifle joint, including the condyle of the tibia. Both muscles may adduct the thigh, but the sartorius is probably more important as a hip flexor. The gracilis is supplied by the obturator nerve, the sartorius by the saphenous nerve.
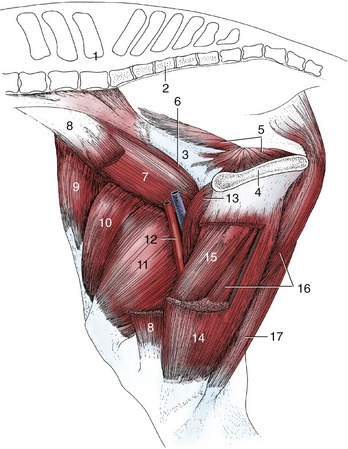
Figure 24–3 Muscles of the thigh, medial view.
1, Last lumbar vertebra; 2, sacrum; 3, shaft of ilium; 4, pelvic symphysis; 5, internal obturator; 6, psoas minor; 7, iliopsoas; 8, sartorius, resected; 9, tensor fasciae latae; 10, rectus femoris; 11, vastus medialis; 12, femoral vessels in femoral triangle; 13, pectineus; 14, gracilis, fenestrated; 15, adductor; 16, semimembranosus; 17, semitendinosus.
The pectineus and adductor constitute the middle layer. The pectineus (Figure 24–3/13) is a small fusiform muscle that arises from the margin of the pubis and inserts on the medial surface of the femur. A part of the tendon of origin is from the contralateral side, and the resulting decussation contributes a transverse strengthening to the prepubic tendon (p. 547). The pectineus is placed to flex the hip and adduct the thigh. It is supplied by the obturator nerve.
The much larger adductor (Figure 24–3/15) fills the space between the pectineus and semimembranosus. It arises from the floor of the pelvis and symphysial tendon and inserts on the caudal surface and medial epicondyle of the femur and the medial collateral ligament of the stifle. Although adduction of the thigh is the primary function, a subsidiary extensor action is possible. Innervation is from the obturator nerve.
The small muscles of the hip—quadratus femoris, gemelli, obturator internus, and obturator externus—are of little importance. The tendon of the obturator internus crosses the margin of the ischium as in the dog. The first three are supplied by the sciatic nerve; while the obturator externus is supplied by the obturator.
THE CRANIAL MUSCLES
This group comprises the quadriceps femoris, which possesses the usual four individually named heads of origin, and the insignificant capsularis.
The four heads of the quadriceps combine in a common insertion on the patella, and the intermediate patellar ligament (Figure 24–4/8) supplies the functional continuation to the tibial tuberosity. The rectus femoris is a potential flexor of the hip, but the principal action of the group is extension of the stifle. Extension, of course, embraces stabilization of the joint to prevent its collapse when the limb bears weight during the support phase of the stride. It can be observed (and confirmed by palpation) that the muscle appears relaxed when the animal stands quietly. This suggests that, once the patella has been brought into its resting position, no considerable further effort is required of the quadriceps. Quadriceps paralysis is a very severe handicap. The animal is unable to stabilize the stifle; it is also unable to stabilize the hock joint, whose movements are linked to those of the stifle by the reciprocal mechanism (p. 638). The group is supplied by the femoral nerve.
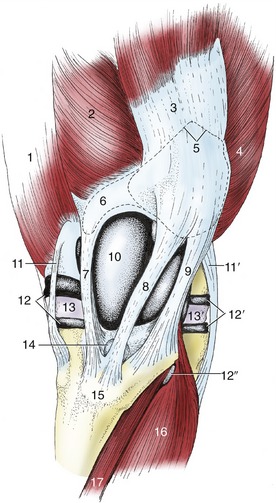
Figure 24–4 The left stifle joint, cranial view.
1, Adductor; 2, vastus medialis; 3, rectus femoris; 4, vastus lateralis; 5, outline of patella; 6, outline of patellar fibrocartilage; 7, 8, 9, medial, intermediate, and lateral patellar ligaments; 10, joint capsule over medial ridge of femoral trochlea; 11, 11′, medial and lateral collateral ligaments; 12, 12′, medial and lateral femorotibial joint capsules; 12″, recess of 12′ under combined tendon of peroneus tertius and long digital extensor; 13, 13′, medial and lateral menisci; 14, distal infrapatellar bursa; 15, tibial tuberosity; 16, long digital extensor; 17, tibialis cranialis.
THE STIFLE JOINT
Although generally conforming to the common pattern, the equine stifle also exhibits several important features of distinction. The most remarkable provide the means of “locking” the joint so that one hindlimb may support a disproportionate part of the body weight and allow the other to be rested while the animal remains standing. The arrangement is a major component of the passive stay-apparatus (p. 636).
The locking mechanism relies on certain peculiarities of the articular surfaces. The femoral trochlea is markedly asymmetrical. The medial ridge is larger than the lateral one and is prolonged proximally to a terminal protuberance that is easily identifiable on palpation (Figures 24–4/10, 24–5/4, and 24–6/2). The trochlear surface comprises two distinct areas. The larger one, known as the gliding surface, corresponds to the whole trochlea of most species and faces in a predominantly cranial direction; the smaller one, known as the resting surface, forms a narrow shelf above the gliding surface, from which it is sharply angled to face proximally (Figure 24–15/18). The patella is broadly diamond-shaped when viewed from in front (Figure 24–15, B/2); in the fresh state it is extended medially by a patellar fibrocartilage (Figure 24–15/3). The articular surface of the patella is also divided. The more extensive backward-facing area engages with the trochlea during the greater part of the normal range of movement; a narrow strip at the apex is directed distally and makes contact with the femur only at the limit of extension.
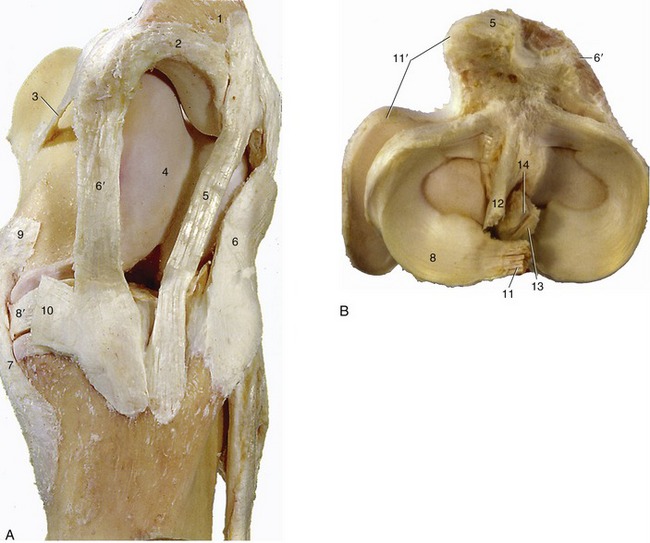
Figure 24–5 The ligaments of the left stifle joint. A, Medial view. B, Proximal view of the left tibia and the menisci.
1, Patella; 2, patellar fibrocartilage; 3, medial femoropatellar ligament; 4, medial ridge of trochlea; 5, intermediate patellar ligament; 6, 6′, lateral and medial patellar ligaments; 7, 7′, lateral and medial collateral ligaments; 8, 8′, lateral and medial menisci; 9, insertion of semimembranosus; 10, insertion of gracilis and sartorius; 11, meniscofemoral ligament; 11′, tendon of popliteus; 12, 13, cranial and caudal cruciate ligaments; 14, intercondylar eminence.
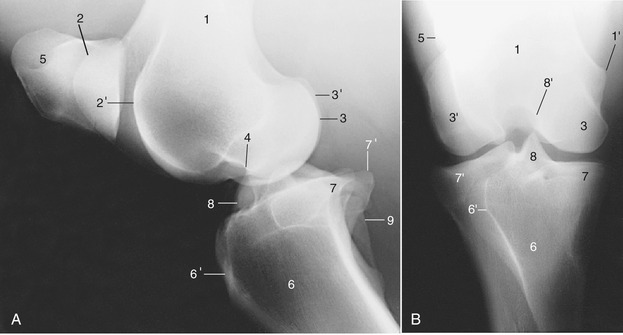
Figure 24–6 Lateral (A) and caudocranial (B) radiographs of the stifle joint.
1, Femur; 1′, medial epicondyle; 2, 2′, medial and lateral ridges of the trochlea; 3, 3′, medial and lateral condyles; 4, extensor fossa; 5, patella; 6, tibia; 6′, tibial tuberosity; 7, 7′, medial and lateral condyles; 8, intercondylar eminence; 8′, intercondylar fossa; 9, fibula.
In this species there are three patellar ligaments joined by a retinaculum in which the insertion tendons of several thigh muscles merge. The intermediate ligament (Figure 24–4/8), the homologue of the single structure of the smaller species, runs from the apex of the patella to the tibial tuberosity. The lateral and medial ligaments run from the angles of the patella or, more accurately where the medial one is concerned, from the parapatellar cartilage. The three ligaments are thus quite widely separated at their origins but converge distally and insert close together. The gap between the proximal parts of the medial and intermediate ligaments is especially wide and is occupied by the medial ridge of the trochlea (Figure 24–4/10).
The patella slides up and down over the femoral trochlea during the greater part of the normal excursions of the joint. Only in extreme extension, as momentarily during the support phase of a walking stride, do the resting surfaces engage. The resting position is also adopted when the animal is standing squarely with its weight evenly distributed over the two hindlimbs. This is easily verified on palpation, and it can be found that the medial ligament then runs even with the edge of the corresponding ridge of the trochlea. This position is maintained without the assistance of the main extensor (quadriceps femoris) of the stifle but does require some effort on the part of the muscles that converge on the medial and lateral patellar ligaments: the biceps and tensor fasciae latae laterally and the gracilis and sartorius medially. The position is unstable and the patella is easily dislodged; it then slips back onto the gliding surface of the trochlea.
The joint cavity is capacious, and its division into compartments is relatively complete. The extensive femoropatellar compartment is mainly contained between the femur, the patella, and the quadriceps. The part distal to the patella is more accessible, though separated from the patellar ligaments (and retinaculum) by a thick cushion of fat. It communicates with the medial femorotibial compartment in the large majority of horses but with the corresponding lateral compartment in far fewer, perhaps 25%. The partition between the medial and lateral compartments is almost always imperforate. The inconstancy of these arrangements has considerable practical importance: it must be assumed that any infection spreads readily among the three compartments, while prudence dictates that therapeutic substances be separately injected into each.
Such injections require familiarity with the disposition of the ligaments and the ability to recognize them on palpation. The medial collateral ligament can be picked out close to its origin from the femoral epicondyle and provides a convenient landmark in puncture of the medial femorotibial compartment. The needle is introduced close to its cranial border, between it and the medial patellar ligament (Figures 24–4/7,11 and 24–5/6′,7′). The lateral collateral ligament is palpable along its whole length but is most easily found close to its insertion on the head of the fibula. The lateral femorotibial compartment is punctured between this ligament and the more cranial, and also palpable, tendon of origin of the long digital extensor (Figure 24–4/11′,16). The femoropatellar compartment is also easily entered from the side, behind the proximal part of the lateral patellar ligament (Figure 24–4/9). Alternatively, this compartment can be approached from in front, between the patellar ligaments, but this requires that the needle be passed through a considerable thickness of fat.
THE SKELETON OF THE LEG AND HOCK; THE HOCK JOINT
The tibia is the only functional component of the skeleton of the leg. Its shaft is thickly covered by muscle on its craniolateral and caudal aspects but is subcutaneous medially (Figure 24–7/1). The distal articular surface, known as the cochlea, comprises two grooves separated by a ridge, all with a craniolateral inclination. The cochlea is flanked by medial and lateral malleoli (Figure 24–8/2,2′).
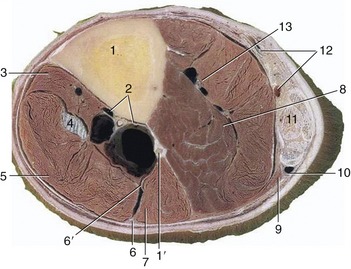
Figure 24–7 Transverse section of the left leg slightly above its middle.
1, Tibia; 1′, fibula; 2, cranial tibial vessels; 3, tibialis cranialis; 4, peroneus tertius; 5, long digital extensor; 6, 6′, superficial and deep peroneal nerves; 7, lateral digital extensor; 8, deep digital flexors; 9, soleus; 10, lateral saphenous vein and caudal cutaneous sural nerve; 11, superficial digital flexor surrounded by the other components of the common calcanean tendon (gastrocnemius and tarsal tendons of semitendinosus and biceps); 12, caudal branch of medial saphenous vein, tibial nerve, and saphenous artery; 13, caudal tibial vessels.
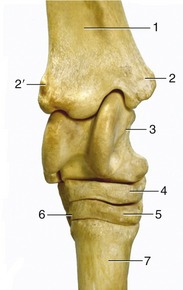
Figure 24–8 Dorsal view of the right hock.
1, Tibia; 2, medial malleolus; 2′, lateral malleolus; 3, talus with trochlea; 4, central tarsal bone; 5, third tarsal bone; 6, fourth tarsal bone; 7, third metatarsal (cannon) bone.
The fibula is much reduced. The proximal extremity or head forms a tight articulation with the lateral condyle of the tibia (Figure 24–1/9). The head usually continues into a short and rodlike shaft, but sometimes a band of soft tissue intervenes; this may simulate a fracture when depicted in a radiograph. In later embryonic life the isolated distal extremity of the fibula becomes assimilated within the tibia to which it furnishes the lateral malleolus (see Figure 2–59, D,E/6′). The independence of the malleolar center of ossification is clearly evident in radiographs of young animals, and the line of union may be evident in the adult bone.
The hock (Figure 24–8) comprises the following elements: talus and calcaneus in the proximal row, a central tarsal bone in the intermediate row, and fused first and second bones and separate third and fourth bones in the distal row. The proximodorsal surface of the talus (Figure 24–8/3) carries an oblique trochlea corresponding to the cochlea of the tibia. The distal surface is more or less flat and rests on the central bone. The calcaneus (Figure 24–9/4,4′) lies largely to the plantar aspect of the talus; the tuber surmounting the calcanean process rises about 5 cm above the tarsocrural joint space and serves as a rough guide to that feature. The composite bone formed by the first and second tarsal bones is relatively small and lies mainly behind the much larger, wedge-shaped third tarsal (Figure 24–8/5). The fourth bone (Figure 24–8/6; on the lateral side) is cuboidal, unlike the other bones in the distal row, which are flattened; its greater depth causes it to occupy both the intermediate and the distal tiers. The bones of the distal row articulate with the metatarsal bones, that is, the third (cannon) bone centrally and the much smaller second and fourth (splint) bones to the sides.
Even cursory examination of the tarsal skeleton is sufficient to make it plain that while free movement is allowed at the tarsocrural joint, there can be almost no play at any other level. The obliquity of the articular surfaces of the tibia and talus ensures that the distal part of the limb is carried outward as well as forward when the hock joint is flexed.
The fibrous layer of the joint capsule extends from the tibia to the metatarsus. It is firmly attached over various parts of the skeleton but is free elsewhere and then varies considerably in strength; the weaker parts (pouches) bulge when the synovial sac is distended. Numerous ligaments are associated with the hock, but the majority are short and are conveniently regarded as mere local thickenings of the capsule. Three that are larger and more discrete are of greater importance. Paired collateral ligaments extend from the malleoli to the corresponding splint bones and may be palpated along their whole lengths (see Figure 24–10, D/9,9′). They have intermediate attachments to the bones they cross, and these help ensure that movements of the hock are restricted to flexion and extension at the tarsocrural level. A long plantar ligament (Figure 24–10, D/10) follows the plantar aspect of the calcaneus, passes over the fourth tarsal, and then continues distally onto the proximal part of the metatarsus. It is largely covered by the tendon of the superficial digital flexor but may be palpated to each side of this. It is commonly strained about the middle of its length, and in lateral view the resulting thickening gives a convex profile to the plantar aspect of the hock. The condition is known as a curb (from the French courbe, curve, contour).
The hock is a compound joint with three joint sacs: one common to the tarsocrural and proximal intertarsal levels, one for the distal intertarsal level, and one for the tarsometatarsal level (Figure 24–9/10-13). The two more distal sacs are small and occasionally communicate; the distal intertarsal sac may be punctured from the medial side, while access to the tarsometatarsal sac may be gained between the fourth tarsal and the head of the splint bone. The novice will find neither technique very reliable. The proximal part of the talocrural sac is capacious and is prone to overdistention, which causes the capsule to pouch at its weakest points. There are three such pouches. One, at the dorsomedial aspect of the hock, is bounded by the tendon of the peroneus tertius, the medial collateral ligament, the medial malleolus, and the medial branch of the tendon of the tibialis cranialis (Figure 24–10, D/8 and Figure 24–10, A-C). The sac is easily punctured here, even when it is not distended; care must be taken to avoid the cranial branch of the medial saphenous vein, which crosses the site. The second and third pouches are on the plantar aspect. One is found between the medial collateral ligament and the deep flexor tendon at the level of the medial malleolus; the other is behind the lateral collateral ligament, between the calcaneus and the lateral malleolus. Unless the joint sac is considerably distended, puncture at either of these sites may prove difficult.

Figure 24–9 Sagittal section of the hock joint.
1, Superficial digital flexor; 1′, subtendinous calcanean bursa; 2, gastrocnemius; 3, tibia; 4, calcaneus; 4′, sustentaculum tali; 5, talus; 6, central tarsal; 7, third tarsal; 8, fourth tarsal (mainly on lateral side); 9, large metatarsal (cannon) bone; 10, tarsocrural joint; 11, proximal intertarsal joint (communicates with 10); 12, distal intertarsal joint; 13, tarsometatarsal joint; 14, deep digital flexor; 14′, tarsal sheath.
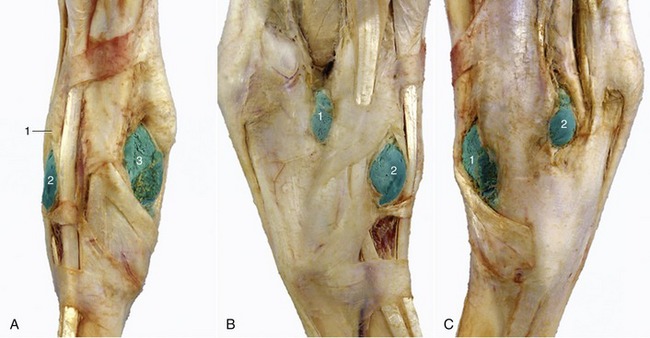
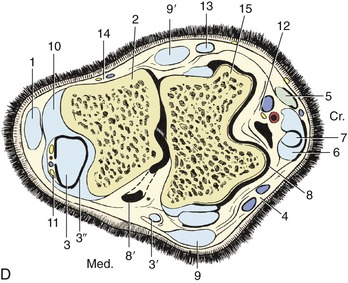
Figure 24–10 A–C, All illustrations of hock. A, Dorsal view of right hock.
1, Long digital extensor; 2, 3, laterodorsal and mediodorsal pouches of tarsocrural joint, respectively, are filled with latex.
B, Lateral view of right hock.
1, 2, Lateral plantar and laterodorsal pouches of tarsocrural joint, respectively, filled with latex.
1, 2, Mediodorsal and medioplantar pouches of tarsocrural joint, respectively.
D, Bursae, tendon sheaths, and joint pouches of the left hock.
1, Superficial digital flexor; 2, calcaneus; 3, lateral deep digital flexor and tibialis caudalis (combined tendon in B); 3′, tendon of medial deep digital flexor; 3″, tarsal sheath; 4, cranial branch of medial saphenous vein; 5, long digital extensor; 6, peroneus tertius; 7, tibialis cranialis and underlying bursa; 8, 8′, dorsal and medioplantar pouches of tarsocrural joint; 9, 9′, medial and lateral collateral ligaments (superficial parts); 10, long plantar ligament; 11, plantar nerves and saphenous vessels; 12, cranial tibial vessels and deep peroneal nerve; 13, lateral digital extensor; 14, caudal cutaneous sural nerve and lateral saphenous vein; 15, talus.
Swelling of the joint sac may be confused with swelling of the synovial (tarsal) sheath around the deep flexor tendon (Figure 24–10/3″). The differential diagnosis is simple. When the joint sac is distended, pressure applied to either plantar pouch is transmitted to the dorsal pouch (and vice versa). Swelling of the tarsal sheath is transmitted from plantaromedial to plantarolateral (or vice versa) if local pressure is applied; it is not transmitted to the dorsal aspect of the joint. Moreover, the swelling of the tendon sheath is evident about 5 cm proximal to the plantar swelling of the joint.
Arthritic changes (spavin) commonly affect the bones of the hock. The changes most commonly begin on the medial aspect, near the meeting of the third and central tarsal and third metatarsal bones. This region, the “seat of spavin,” is crossed by the medial branch of the tibialis cranialis tendon (the cunean tendon of clinical authors) (Figure 24–10/7) en route to its insertion on the combined first and second tarsal bones. The tendon is a useful reference point because it is palpable. A portion is sometimes resected for the purposes of reducing pressure over the lesion and eliminating movement between the distal tarsal elements. The treatment is often effective in reducing pain, although obviously it does not cure the condition.
THE MUSCLES OF THE LEG
The leg is enveloped by three layers of fascia. The superficial layer continues the corresponding fascia of the thigh. The middle layer is formed by the aponeuroses of the tensor fasciae latae, biceps, semitendinosus, gracilis, and sartorius. Its lateral and medial parts combine on the caudal aspect to form a stout plate that bridges the space between the deep flexor and the common calcanean tendon. The plate receives the tarsal tendons of the biceps and semitendinosus and attaches to the calcaneus as part of the common calcanean tendon formation. The saphenous artery, medial and lateral saphenous veins, and lateral and caudal sural nerves are enclosed between the superficial and middle fasciae. The deep fascial layer extends septa that pass between the muscles to attach to the tibia. It thus divides the leg into a number of osteofascial compartments.
THE CRANIOLATERAL MUSCLES
This group comprises the tibialis cranialis, peroneus tertius, and the long and lateral digital extensors. All are flexors of the hock, and those that proceed farther are extensors of the digit. The tibialis cranialis arises from the lateral condyle and tuberosity of the tibia and continues distally, closely applied to the bone (Figure 24–7/3). The insertion tendon begins just above the level of the hock and passes through a split in the tendon of the peroneus tertius before dividing itself. The larger dorsal branch continues to the metatarsal tuberosity. The smaller medial branch diverges to cross the medial collateral ligament before inserting on the combined first and second tarsal bones (Figure 24–10). When the muscle contracts, it presses on the seat of spavin. Although the tibialis cranialis appears to be a flexor of the hock, it is difficult to be certain of its function. According to one view, its prime role is to counteract the bending moment applied to the tibia by the action of other muscles and by gravity.
The peroneus tertius is almost exclusively tendinous (Figure 24–7/4). It arises from the lower end of the femur together with the long extensor; for much of its course it is recessed in the deep surface of that muscle. It bifurcates at the hock; the lateral branch inserts on the calcaneus and fourth tarsal bone, the dorsal one on the proximal part of the third tarsal and third metatarsal bones (Figure 24–12/1). The tendon links the actions of the stifle and hock joints, which is a function convincingly demonstrated should it be ruptured (see Figure 24–16); there then appears an ability to extend the hock while retaining a flexed stifle, which is a combination of movements normally impossible.
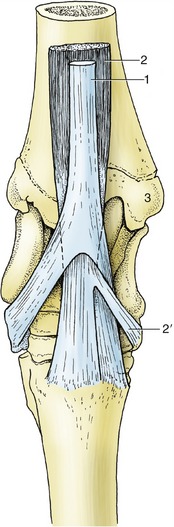
Figure 24–12 The insertion of the flexors of the right hock, dorsal view.
1, Peroneus tertius, splitting into dorsal and lateral branches; 2, tibialis cranialis, splitting into dorsal and medial (cunean, 2′) branches; 3, medial malleolus.
The long digital extensor, the largest muscle of the group, arises in common with the peroneus tertius by a short tendon. This is soon succeeded by a broad belly that covers the tibialis cranialis (Figure 24–13/5). The insertion tendon begins in the lower leg and continues to the extensor process of the distal phalanx, with passing attachments to the proximal and middle phalanges. It is joined by the smaller tendon of the lateral extensor (Figure 24–13/6) near the middle of the cannon. As it descends on the dorsal surface of the limb it is surrounded by a synovial sheath from midtarsal level to the tendon union and is held in place by three retinacula where it crosses the hock. This muscle is capable of flexion of the hock and extension of the digit.
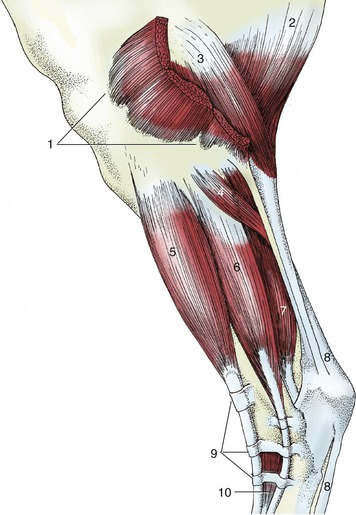
Figure 24–13 The stifle and leg, lateral view.
1, Distal divisions of biceps; 2, semitendinosus; 3, gastrocnemius; 4, soleus; 5, long digital extensor; 6, lateral digital extensor; 7, deep digital flexors; 8, superficial digital flexor; 9, proximal, middle, and distal extensor retinacula; 10, extensor digitalis brevis.
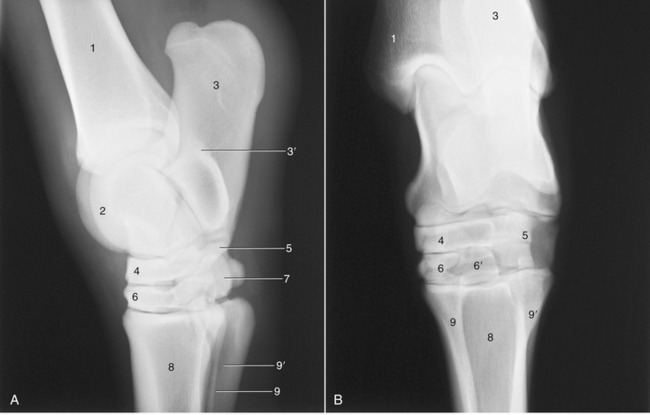
Figure 24–11 Lateral (A) and dorsoplantar (B) radiographs of the hock joint.
1, Tibia; 1′, medial malleolus (outlined); 2, talus; 3, calcaneus; 3′, sustentaculum tali; 4, central tarsal; 5, fourth tarsal; 6, third tarsal (in B superimposed on tarsal 1 and 2); 6′, plantar projection of third tarsal; 7, tarsal 1 and 2; 8, large metatarsal bone; 9, 9′, medial and lateral splint bones.
The lateral digital extensor runs between the long extensor and the deep flexor on the lateral aspect of the limb. It arises from the lateral collateral ligament of the stifle and adjacent parts of both tibia and fibula and ends by joining the long extensor tendon. Its tendon is also held down by retinacula and protected by a synovial sheath where it crosses the hock. A very small, short digital extensor muscle (extensor digitalis brevis) occupies the angle between the converging tendons of the larger muscles (Figure 24–13/10). It is of no importance.
All muscles of the craniolateral group are supplied by the peroneal nerve.
THE CAUDAL MUSCLES
This group comprises the popliteus, whose action is confined to the stifle, and the gastrocnemius, soleus, and superficial and deep digital flexors, which all extend the hock; the last two also flex the digit.
The popliteus is a relatively small triangular muscle placed directly over the caudal aspect of the stifle joint (Figure 24–14, B/7). It arises from the lateral condyle of the femur and inserts on the caudomedial border of the tibia. The popliteus flexes the stifle and rotates the leg inward.
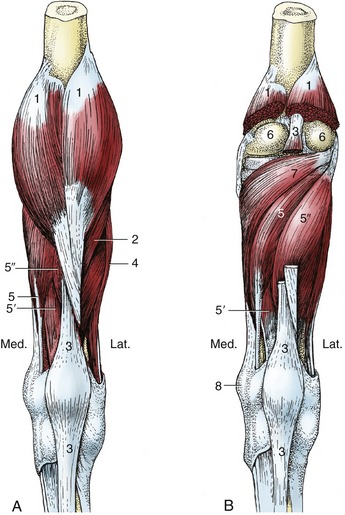
Figure 24–14 Superficial (A) and deep (B) muscles of the right leg, caudal view.
1, Gastrocnemius; 2, soleus; 3, superficial digital flexor; 4, lateral digital extensor; 5, 5′, 5″, medial and lateral deep digital flexors and tibialis caudalis; 6, femoral condyles; 7, popliteus; 8, medial malleolus.
The gastrocnemius, the most superficial and largest muscle of the group, arises by two heads from the supracondylar tuberosities of the femur (Figure 24–14/1). The heads, which are first covered by the hamstring muscles, soon unite in a single strong tendon that is a major component of the calcanean tendon. The gastrocnemius tendon inserts on the point of the hock where it is covered by the tendon of the superficial flexor. To attain this deep position, it must first wind around the lateral border of the flexor tendon, where it is cushioned by the interposition of a synovial bursa (see below). Theoretically, the gastrocnemius is a flexor of the stifle and extensor of the hock, but because the tendons of the peroneus tertius and superficial flexor ensure that these joints extend or flex together, it is difficult to envisage its action. It has been asserted that its prime function is comparable to that of the tibialis cranialis, that is, adjustment of the load on the tibia. A ribbonlike soleus runs from the head of the fibula to the gastrocnemius tendon but is of no importance.
The superficial digital flexor (Figure 24–14, B/3) is largely tendinous, although it has a slightly greater content of flesh than the peroneus tertius. It arises from the supracondylar fossa of the femur under cover of the gastrocnemius and, twisting around the medial surface of the tendon of that muscle, passes toward the calcanean tuber where it expands to form a cap; the medial and lateral edges attach here, but the main part continues over the plantar aspect of the hock to enter the cannon. It inserts on the first and second phalanges in similar fashion to the superficial flexor of the forelimb. A considerable synovial bursa protects the expanded tendon where it caps the tuber; the bursa also extends proximally between the flexor and gastrocnemius tendons, where they wind around each other (Figure 24–9, D/1′). A second, smaller, subcutaneous bursa (Figure 24–9/1″) may form over the expanded tendon where it caps the calcaneus (“capped hock”). Both bursae are liable to inflammation and distention. The proximal part of the muscle is a main constituent of the so-called reciprocal mechanism (p. 638). The distal part supports the fetlock and pastern joints in similar fashion to the superficial flexor of the forelimb.
The deep digital flexor arises by three separate and individually named heads—lateral digital flexor, medial digital flexor, and tibialis caudalis—which later unite to form a single stout tendon of insertion. The medial flexor arises from the lateral condyle of the tibia but soon swings to the medial side of the leg (Figure 24–14/5). The narrow tendon passes the hock, resting within a groove on the medial malleolus and medial collateral ligament, where it is protected by a synovial sheath. Once past the hock, the tendon unites with the tendon common to the other two bellies.
The lateral flexor and the tibialis caudalis have extensive origins from the caudal surface of the tibia, distal to the attachment of the popliteus (Figure 24–14/5′,5″). They are difficult to separate, and there is little merit in attempting the distinction because the tendons combine in the lower part of the leg. The common tendon crosses the plantar aspect of the hock over the sustentaculum tali of the calcaneus. A synovial (tarsal) sheath invests the tendon from the distal part of the leg to its junction with the tendon of the medial flexor in the upper part of the cannon (Figure 24–10, D/3″). A further tendinous slip (the accessory ligament) that passes from the joint capsule to join the common tendon is analogous to the forelimb formation but is usually less developed and may even be absent. The distal part of the tendon comports itself similarly to the corresponding part of the deep digital flexor of the forelimb.
The deep plantar metatarsal fascia resembles the corresponding forelimb fascia and offers the same obstruction to palpation of the flexor tendons in the proximal half, and more, of the cannon.
The tibial nerve supplies all muscles of the caudal group.
The remaining structures of the metatarsus and digit closely resemble the corresponding parts of the forelimb. Certain quantitative differences have been mentioned (p. 586 and Figure 23–36).
THE PASSIVE STAY-APPARATUS
The caudal end of the trunk rests on the head of the femur. A vertical line dropped from the center of the support passes caudal to the stifle joint and cranial to the hock, fetlock, and pastern joints before intersecting the hoof (Figure 24–15, A). If unsupported, the bony column of the hindlimb would collapse by flexion of the stifle and hock and overextension of the fetlock and pastern joints. The tendons and ligaments of the passive stay-apparatus enable the horse to prevent this collapse using only a minimum of muscular effort.
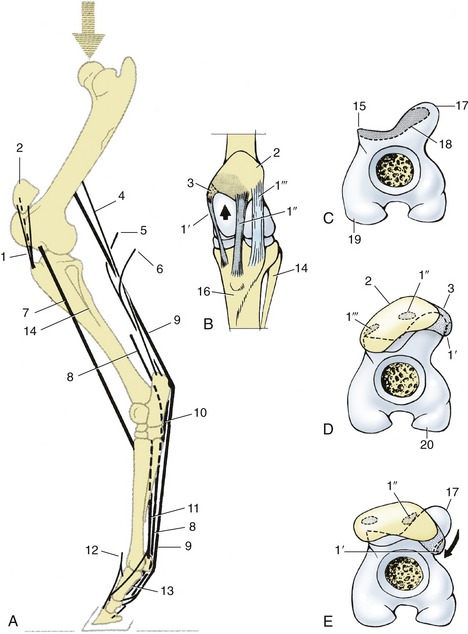
Figure 24–15 A, Stay-apparatus of the left hindlimb, lateral view. B, Left stifle joint, cranial view. C–E, Distal end of left femur, looking distally; in D, position of patella in horse standing square; in E, the stifle is locked.
1, patellar ligaments; 1′, medial; 1″, intermediate; 1′″, lateral patellar ligaments; 2, patella; 3, parapatellar fibrocartilage; 4, fibrous band associated with gastrocnemius; 5, tarsal tendon of semitendinosus; 6, tarsal tendon of biceps; 7, peroneus tertius; 8, deep digital flexor; 9, superficial digital flexor; 10, long plantar ligament; 11, interosseous; 12, long digital extensor; 13, sesamoidean ligaments; 14, fibula; 15, lateral trochlear ridge; 16, tibia; 17, tubercle on proximal end of medial trochlear ridge; 18, resting surface on proximal end of trochlea; 19, lateral condyle; 20, medial condyle.
The supportive mechanisms below the hock are very similar to those of the forelimb (p. 614). However, the accessory ligament of the deep digital flexor tendon, which arises from the caudal aspect of the hock, is weak and occasionally absent. This is compensated by the firm, intermediate attachment of the superficial digital flexor tendon to the point of the hock, which is broadly comparable in function to the accessory ligament of the corresponding tendon of the forelimb. The part of the superficial flexor tendon between its attachments proximal and distal to the fetlock joint is tensed when weight is on the limb and assists the interosseous in supporting the fetlock.
Fixation of the stifle and hock joints depends on the locking mechanism of the former joint and the existence of the so-called reciprocal mechanism, which associates the movements of the two joints. For the horse to “lock” the stifle, the patella is first brought into the resting position (by extending the joint) and then fixed by being rotated medially through about 15° (Figure 24–15, E/arrow). This hooks the parapatellar cartilage and medial patellar ligament securely over the protuberance of the medial trochlear ridge (Figure 24–15/17); palpation confirms that the medial ligament now runs more caudally than before, being displaced as much as 2 cm behind the crest of the medial ridge. Secured in this position, the patella firmly resists displacement, and a larger part of the body weight can be lowered onto the locked joint, which enables the other hindlimb to be rested with only the toe of the hoof on the ground. The “unlocking” is effected quite briskly: the patella is rotated laterally and snaps back into its usual place; the joint may now be flexed.
The reciprocal mechanism is provided by two tendinous cords—the peroneus tertius and the superficial flexor—that pass between the distal end of the femur and the hock, one on the cranial, the other on the caudal aspect of the tibia (Figure 24–15/7,9). (Figure 24–16, A, demonstrates the result of the rupture of the peroneus tertius.) These ensure that the two joints move in unison; flexion or extension of one necessitates a similar movement of the other joint. However, some looseness in the system renders it unnecessary for the angular changes at the two joints to be exactly the same, especially during fast gaits when large forces must be absorbed by the tendons.


When the stifle is locked, the weight of the hindquarters tends to flex the hock joint; this is opposed by tension in the superficial flexor caudal to the tibia. The peroneus tertius is not involved at this time, and it seems that it is superfluous in the animal standing quietly.
The stifle joint is fully locked only when the horse takes most of the weight on that limb and rests the other on the toe of the hoof. It should be emphasized that while the arrangement conserves energy, it does not eliminate muscular effort; every few minutes the animal shifts its support from one side to the other as muscles tire or, perhaps, as tension in the passive supporting structures becomes uncomfortable.
Sometimes a neuromuscular disorder makes unlocking of the stifle difficult or even impossible (Figure 24–16, B). A temporary “lock” may be broken by startling a horse into sudden movement; a persistent “lock” may be alleviated by section of the medial ligament to break the retention loop (Figure 24–15, B/1′). The operation is easily and safely performed because a considerable thickness of fat lies deep to the ligament, protecting the synovial membrane.
VASCULARIZATION OF THE HINDLIMB
The chief artery of the limb, the femoral artery, directly continues the external iliac artery (Figure 24–17/1,3). It reaches the femoral triangle, traveling in company with the femoral vein and nerve and almost at once detaches the saphenous artery and several larger muscular branches. The saphenous artery (Figure 24–17/8) pursues a superficial course down the medial aspect of the limb, where it may be traced almost to the hock.
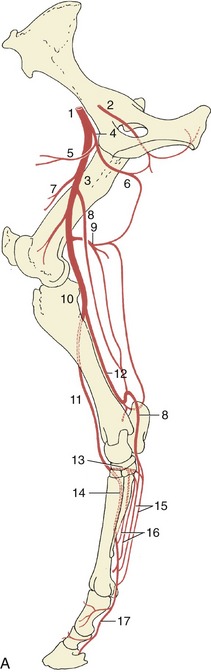
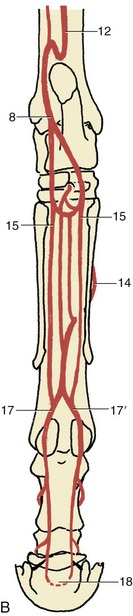
Figure 24–17 The principal arteries of the right hindlimb, medial view (A), caudal view (B).
1, External iliac a.; 2, obturator a.; 3, femoral a.; 4, deep femoral a.; 5, pudendoepigastric trunk; 6, medial circumflex femoral a.; 7, lateral circumflex femoral a.; 8, saphenous a.; 9, caudal femoral a.; 10, popliteal a.; 11, cranial tibial a.; 12, caudal tibial a.; 13, perforating tarsal a.; 14, dorsal metatarsal a.; 15, medial and lateral plantar aa.; 16, medial and lateral plantar metatarsal aa.; 17, 17′, medial and lateral digital aa.; 18, terminal arch, anastomosis of digital aa. within the distal phalanx.
The muscular branches include deep and caudal femoral arteries (Figure 24–17/4,9) that anastomose with each other and with other more proximal and more distal arteries, forming an alternative pathway available when the chief trunk is obstructed. The femoral artery then passes obliquely over the femur to gain the caudal aspect of the stifle, where it passes between the heads of the gastrocnemius. The segment at the stifle, known as the popliteal artery, divides into cranial and caudal tibial arteries in the upper part of the leg.
The larger cranial tibial artery (Figure 24–17/11) passes through the interosseous space between the fibula and tibia to gain the dorsolateral aspect where it turns distally between the muscles and the bone. It comes to the surface at the hock and continues as the dorsal pedal artery and then, on entering the groove between the cannon and lateral splint bones, as the dorsal metatarsal artery. A perforating branch (Figure 24–17/13) of the dorsal pedal passes between the tarsal bones to reach the plantar aspect of the limb, where it anastomoses with branches of the saphenous artery. The dorsal metatarsal artery, the major supply to the foot, is well placed at the proximal end of the cannon for evaluation of the pulse. Toward the fetlock, it passes under the free end of the splint bone to gain the plantar aspect of the cannon, where it is reinforced by small branches from the saphenous. It ends by dividing into medial and lateral digital arteries (Figure 24–17/17,17′) that replicate the pattern of the forelimb vessels.
The caudal tibial artery first runs distally in the deep flexor (Figure 24–17/12). Toward the hock it enters the space before the calcanean tendon and sends a short S-shaped anastomosis to the nearby saphenous artery and a longer branch that reascends the leg to join the caudal femoral. The saphenous artery, thus reinforced, divides into medial and lateral plantar arteries that descend toward the fetlock (Figure 24–17/15). These and the deeper plantar metatarsal arteries are individually of no great importance and may eventually fade away or join the dorsal metatarsal artery or its digital divisions.
The deep veins are largely satellite to the arteries. As in the forelimb, certain superficial trunks run alone; they include the medial and lateral saphenous veins. A branch of the former is often prominent where it crosses the dorsal aspect of the hock; the swelling (“blood spavin”) may occasionally be mistaken for a distention of the dorsal joint pouch (Figure 24–10/4,8). Within the leg, the saphenous veins run between the calcanean tendon and the caudal muscle mass, one to each side (Figure 24–7/10,12). The medial vein later crosses the medial aspect of the thigh to open into the femoral vein. The lateral vein joins the caudal femoral vein at the stifle.
Lymph draining from the distal part of the limb passes mainly to the group of popliteal nodes tucked within the popliteal fossa between the biceps and semitendinosus. Efferent vessels from this group and additional vessels that arise within the thigh proceed mainly to the deep inguinal nodes within the femoral triangle. Some lymph from superficial structures passes to the subiliac nodes, which drain into the lateral and medial iliac nodes. The courses of certain lymphatic vessels may be manifested as cords visible through the skin in some lymph-borne infections.
THE NERVES OF THE HINDLIMB
The formation and ramification of the lumbosacral plexus and the distribution of its peripheral branches follow the common pattern in broad outline; important species differences are confined to the innervation of the foot.
The cranial and caudal gluteal nerves attend to the innervation of the lateral muscles of the croup, including the vertebral heads of the hamstring muscles; the details have been given.
The distributions of the femoral, obturator, and sciatic nerves have greater clinical relevance. The femoral nerve (L4–L6) (Figure 24–18/1) passes through and also supplies the sublumbar muscles before entering the thigh by way of the vascular lacuna. It then splits into several branches, most of which at once enter the quadriceps. The one branch of more extended course, the saphenous nerve (Figure 24–18/1′), continues within the femoral triangle before penetrating the medial femoral fascia to obtain a more superficial position. It continues through the thigh, leg, and upper cannon, supplying skin over the medial aspect of the limb from thigh to fetlock. It also supplies the sartorius. Extensive damage to the femoral nerve is uncommon, but when it does occur, the consequences are severe; paralysis of the quadriceps removes the ability to fix the stifle and therefore the ability to support weight on the affected limb. In addition, skin sensibility is lost over a considerable area.
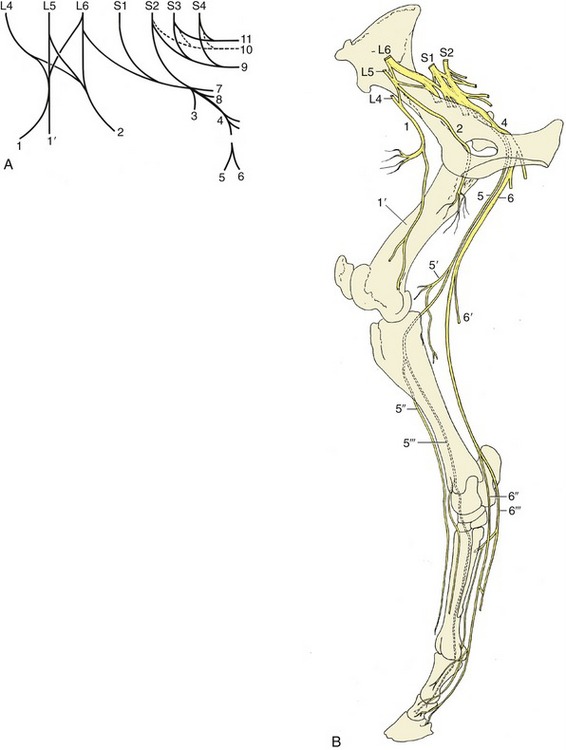
Figure 24–18 The nerves of the hindlimb. A, The lumbosacral plexus, schematic. B, The principal nerves, medial view.
1, Femoral n.; 1′, saphenous n.; 2, obturator n.; 3, cranial gluteal n.; 4, sciatic n.; 5, common peroneal n.; 5′, lateral cutaneous sural n.; 5″, 5′″, superficial and deep peroneal nn.; 6, tibial n.; 6′, caudal cutaneous sural n.; 6″, 6′″, medial and lateral plantar nn. (the lateral nerve gives rise to the plantar metatarsal nn.); 7, caudal gluteal n.; 8, caudal cutaneous femoral n.; 9, pudendal n.; 10, pelvic n.; 11, caudal rectal n.
The obturator nerve (L4–L6) (Figure 24–18/2) leaves the pelvis by way of the obturator foramen and innervates the adductor muscles (pectineus, gracilis, adductor, and obturator externus). Injury, which generally follows foaling or a pelvic fracture, results in partial or complete inability to adduct the limb. The severity of the dysfunction is rather unpredictable; it appears to depend on the weight of the animal, the nature of the terrain, and the extent of the lesion.
The sciatic nerve (L6–S2) (Figure 24–18/4) leaves the pelvis by the greater sciatic foramen and after a short course over the sacrosciatic ligament turns distally caudal to the hip joint to enter the thigh under cover of the biceps. It divides about the level of the joint into tibial and peroneal nerves that initially run together. They part company a little above the stifle, when the peroneal nerve moves laterally to pass between the biceps and the lateral head of the gastrocnemius; the tibial nerve holds its course and runs between the two heads of the gastrocnemius. Both divisions detach cutaneous branches while still within the thigh. That from the peroneal (lateral cutaneous sural nerve; Figure 24–18/5′) becomes subcutaneous by piercing the biceps and then spreads to supply skin over the lateral aspect of the leg. The corresponding tibial branch (caudal cutaneous sural nerve; Figure 24–18/6′) descends in the fascial plate between the calcanean tendon and deep flexor, following the lateral saphenous vein for part of its course. It supplies branches to the skin over the plantarolateral aspect of the hock and cannon, reaching to the fetlock.
The peroneal nerve divides caudal to the lateral collateral ligament of the stifle into deep and superficial branches. The superficial branch (Figure 24–18/5″) continues down the leg, slightly sunken within the groove between the long and lateral extensors, where it can be palpated below the middle of the leg. It supplies the lateral extensor, the skin over the lateral aspect of the leg, and more distal segments of the limb. The deep branch takes a parallel course after sinking deeply between the same two muscles to follow the cranial face of the intervening septum (Figure 24–18/5′″ and Figure 24–7/6′). It supplies branches to the remaining muscles of the dorsolateral group and then continues under cover of the long extensor tendon as a purely sensory nerve that splits into medial and lateral branches over the hock. These, the medial and lateral dorsal metatarsal nerves, edge toward the grooves between the cannon and splint bones (Figure 24–19/3,3′). The lateral nerve follows the palpable dorsal metatarsal artery (Figure 24–20/8). After detaching twigs to the skin and the fetlock and pastern joints, both finally fade within the hoof.
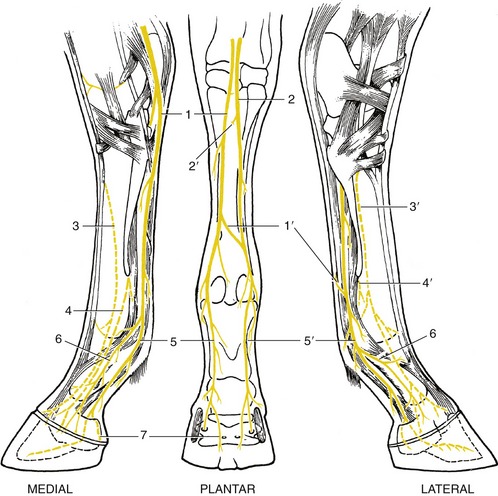
Figure 24–19 The nerves of the right hindfoot.
1, 2, Medial and lateral plantar nn. (from tibial); 1′, communicating branch; 2′, deep branch (for plantar metatarsal nn.), cut; 3, 3′, medial and lateral dorsal metatarsal nn. (from deep peroneal); 4, 4′, medial and lateral plantar metatarsal nn. (from lateral plantar, 2′); 5, 5′, medial and lateral digital nn.; 6, dorsal branch of digital n.; 7, branch to digital cushion.
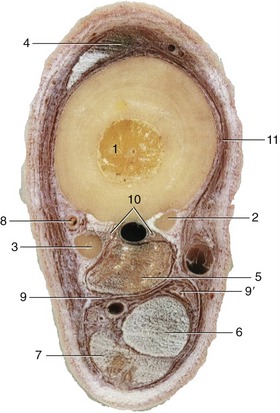
Figure 24–20 Transverse section of the middle of the left metatarsus.
1-3, Large and small metatarsal bones; 4, long digital extensor; 5, interosseous; 6, deep digital flexor; 7, superficial digital flexor; 8, dorsal metatarsal a. and lateral dorsal metatarsal n.; 9, 9′, lateral and medial plantar vessels and n.; 10, plantar metatarsal vessels and nn.; 11, medial dorsal metatarsal n.
Complete section of the peroneal nerve results in inability to extend the digit actively; the hoof rests on its dorsal surface unless the ground surface is passively set down. The posture invites comparison with that which occurs in radial paralysis. Afflicted animals may learn to compensate in a similar manner: they flick the foot forward and plant the hoof before the impetus is lost. In addition to the motor disability, skin sensation is lost over the dorsolateral aspect of the lower part of the limb. Peroneal lesions are most frequent in two circumstances: intrapelvic damage to the sciatic nerve (which is likely also to involve the tibial division) and as the result of trauma in the region of the fibula, where the nerve is superficial (Figure 31–13, shown on a cow).
The tibial nerve dives between the two heads of the gastrocnemius and crosses the stifle on the surface of the popliteus. It detaches branches to these muscles and to other muscles of the caudal group before continuing as a sensory trunk in the space between the calcanean tendon and the deep flexor, where it is easily palpated (Figure 24–7/12). When level with the calcaneus, it divides into medial and lateral plantar nerves that pass over the sustentaculum tali beside the deep flexor tendon. The lateral nerve diverges laterally, and just distal to the hock, it detaches the common trunk of the medial and lateral plantar metatarsal nerves (Figure 24–19/2′). These supply the interosseous muscle and associated structures and the plantar pouch of the fetlock joint (Figure 24–19/4,4′). The medial plantar nerve follows the line of the parent trunk. Although the plantar nerves generally resemble the palmar nerves of the forelimb, the communicating branch is relatively slight or even absent; when present, it can usually be palpated as it slopes in a laterodistal direction over the superficial aspect of the flexor tendons (Figure 24–19/1).
There is one other difference. The dorsal and plantar metatarsal nerves play a larger role in the sensory innervation of the hoof contents than do the corresponding forelimb trunks—the dorsal branch of the ulnar nerve and the palmar metacarpal nerves—which commonly fail to reach the coronet.
Tibial paralysis is manifested by a slight sagging of the hock when weight is borne on the affected limb. Despite the inability to flex the distal joints, the gait is not seriously disturbed. The sensory deficit is very considerable.
Lesions that affect the sciatic trunk involve the hamstring as well as the leg muscles. Despite this, the consequences are less disastrous than might well be supposed. Retention of activity by the quadriceps enables the animal to fix the stifle and, through the reciprocal apparatus, the hock. It is thus able to support weight on the limb. Cutaneous and deep sensations are lost below the stifle, except in the province of the saphenous nerve.
The tibial nerve may be blocked on the lateral side of the limb approximately 10 cm above the point of the hock.
Both superficial and deep branches of the peroneal nerve can be blocked by injecting, subcutaneously and then deeply from the same point of entry, between the long and lateral extensors a handbreadth or so proximal to the tarsocrural joint (Figure 24–7/6,6′). Apart from this, the local anesthetic techniques for surgical and diagnostic purposes generally resemble those prescribed for the forelimb; the one distinction of relevance is the distal extension of the dorsal metatarsal nerves. It is possible to block the undivided tibial nerve (level with the point of the hock) as an alternative to the plantar nerves (Figure 24–7/12).