CHAPTER 80 Pelvic girdle, gluteal region and thigh
The pelvic girdle consists of the paired hip bones (each consisting of the ilium, ischium and pubis) and a single sacrum. The two pubic bones articulate anteriorly at the pubic symphysis and the sacrum articulates posteriorly with the two iliac bones: however, the bones are virtually incapable of independent movement except in the female during parturition. The pelvic girdle is massively constructed and serves as a weightbearing and protective structure, as an attachment for trunk and limb muscles, and as the skeletal framework of a birth canal capable of accommodating a large-headed fetus.
The gluteal region or buttock is an area demarcated by the gluteal fold inferiorly, a line joining the greater trochanter and the anterior superior iliac spine laterally, the iliac crest superiorly and the midline medially. It contains a large bulk of skeletal muscle that covers several vulnerable neurovascular structures, and incorporates junctional zones between the lower limb, pelvis and perineum at the sciatic foramina. Direct and indirect musculoskeletal injuries in this region may damage the sciatic nerve and gluteal vessels.
The thigh consists of a cylinder of compact bone, the femoral shaft, surrounded by muscle groups traversed by important neurovascular structures. The muscles are grouped according to function and lie within osteofascial compartments that are defined by fascial septa running between the femur and an enveloping tube of tough fascia, the fascia lata. The femoral artery gives off its major branch, the profunda femoris, in the anterior compartment, and the sciatic nerve usually divides into its main branches, the tibial and common fibular nerves, as it passes through the posterior compartment. The femoral nerve divides soon after entering the anterior compartment beneath the inguinal ligament; the obturator nerve enters the region proximally and medially from the pelvis and divides into its main branches which run anterior and posterior to adductor brevis.
SKIN AND SOFT TISSUES
SKIN
See also Chapter 79.
Vascular supply and lymphatic drainage
Buttock
Most of the skin of the buttock is supplied by musculocutaneous perforating vessels from the superior and inferior gluteal arteries. There are also small peripheral contributions from similar branches of the internal pudendal, iliolumbar and sacral arteries.
Cutaneous veins are tributaries of vessels that correspond to the named arteries. Cutaneous lymphatic drainage is to the superficial inguinal nodes.
Thigh
The skin of the thigh distal to the inguinal ligament and gluteal fold is supplied mainly by branches of the femoral and profunda femoris arteries. There is some contribution from the obturator, inferior gluteal and popliteal arteries and from direct cutaneous, musculocutaneous and fasciocutaneous vessels. For further details consult Cormack & Lamberty (1994).
Cutaneous veins are tributaries of vessels that correspond to the named arteries. Cutaneous lymphatic drainage is to the superficial inguinal nodes, mainly via collecting trunks accompanying the long saphenous vein.
SOFT TISSUE
Fascia
Superficial fascia
The superficial fascia of the buttock is continuous superiorly with that over the low back, and contains a variable quantity of coarse fat. The superficial fascia of the thigh consists, as elsewhere in the limbs, of loose areolar tissue containing fat. In some regions, particularly near the inguinal ligament, it splits into recognizable layers, between which may be found the branches of superficial vessels and nerves. It is thick in the inguinal region, where its two layers enclose the superficial inguinal lymph nodes, long saphenous vein and other smaller vessels. Here the superficial layer is continuous with that of the abdominal fascia. The deep layer, a thin fibroelastic stratum, is most marked medial to the long saphenous vein and inferior to the inguinal ligament, and is interposed between the subcutaneous vessels and nerves and the deep fascia, fusing with the latter a little below the ligament. (The line of fusion lies in the floor of the ventral flexure line, otherwise known as the groove associated with the hip joint.) This membranous layer of superficial fascia overlies the saphenous opening, blending with its circumference and with the femoral sheath. Over the opening it is perforated by the long (great) saphenous vein, by the superficial branches of the femoral artery other than the superficial circumflex iliac (which perforates the fascia lata separately), and lymphatic vessels, hence the term cribriform fascia (Latin cribrum = a sieve).
Deep fascia
The deep fascia covering the gluteal muscles varies in thickness. Over the maximus it is thin, but over the anterior two-thirds of the medius it forms the thick, strong gluteal aponeurosis. This is attached to the lateral border of the iliac crest superiorly, and splits anteriorly to enclose tensor fasciae latae and posteriorly to enclose gluteus maximus.
Fascia lata
The fascia lata, the wide deep fascia of the thigh, is thicker in the proximal and lateral parts of the thigh where tensor fasciae latae and an expansion from gluteus maximus are attached to it. It is thin posteriorly and over the adductor muscles, but thicker around the knee, where it is strengthened by expansions from the tendon of biceps femoris laterally, sartorius medially, and quadriceps femoris anteriorly. The fascia lata is attached superiorly and posteriorly to the back of the sacrum and coccyx, laterally to the outer margin of the iliac crest, anteriorly to the inguinal ligament and superior ramus of the pubis, and medially to the inferior ramus of the pubis, the ramus and tuberosity of the ischium, and the lower border of the sacrotuberous ligament. From the iliac crest it descends as a dense layer over gluteus medius to the upper border of gluteus maximus, where it splits into two layers, one passing superficial and the other deep to the muscle, the layers reuniting at the lower border of the muscle.
Iliotibial tract
Over the flattened lateral surface of the thigh, the fascia lata thickens to form a strong band, the iliotibial tract. The upper end of the tract splits into two layers, where it encloses and anchors tensor fasciae latae and receives, posteriorly, most of the tendon of gluteus maximus. The superficial layer ascends lateral to tensor fasciae latae to the iliac crest; the deeper layer passes up and medially, deep to the muscle, and blends with the lateral part of the capsule of the hip joint. Distally, the iliotibial tract is attached to a smooth, triangular facet (Gerdy’s tubercle) on the anterolateral aspect of the lateral condyle of the tibia where it is superficial to, and blends with, an aponeurotic expansion from vastus lateralis. When the knee is extended against resistance it stands out as a strong, visible ridge on the anterolateral aspect of the thigh and knee.
Distally, the fascia lata is attached to all exposed bony points around the knee joint, such as the condyles of the femur and tibia, and the head of the fibula. On each side of the patella the deep fascia is reinforced by transverse fibres, which receive contributions from the lateral and medial vasti. The stronger lateral fibres are continuous with the iliotibial tract.
Intermuscular septa
The deep surface of the fascia lata yields two intermuscular septa, which are attached to the whole of the linea aspera and to its proximal and distal prolongations. The lateral septum, thicker and stronger than the medial one, extends from the attachment of gluteus maximus to the lateral femoral condyle, and lies between vastus lateralis in front and the short head of biceps femoris behind and provides partial attachment for them. The medial, thinner and weaker septum lies between vastus medialis and the adductors and pectineus. Numerous smaller septa, such as that separating the thigh adductors and flexors, pass between the individual muscles, ensheathing them and sometimes providing partial attachment for their fibres.
Saphenous opening
The saphenous opening is an aperture in the deep fascia, inferolateral to the medial end of the inguinal ligament, which allows passage to the long saphenous vein and other smaller vessels (Fig. 80.1). The cribriform fascia, which is pierced by these structures, fills in the aperture and must be removed to reveal it. Adjacent subsidiary openings may exist to transmit venous tributaries. In the adult the approximate centre of the opening is 3 cm lateral to a point just distal to the pubic tubercle. The length and width of the opening vary considerably. The fascia lata in this part of the thigh displays superficial and deep strata (not to be confused with the superficial and deep layers of the superficial fascia described above). They lie, respectively, anterior and posterior to the femoral sheath; with the saphenous opening situated where the two layers are in continuity. This serves to explain the somewhat oblique and spiral configuration of the saphenous opening.
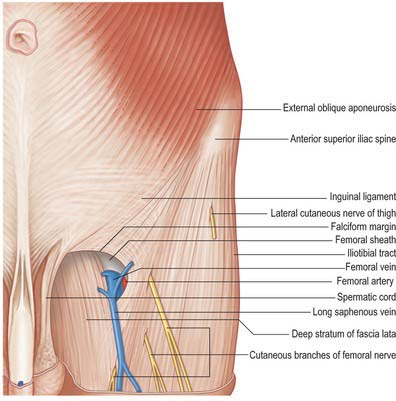
The superficial stratum, lateral and superior to the saphenous opening, is attached, in continuity, to the crest and anterior superior spine of the ilium, to the whole length of the inguinal ligament, and to the pecten pubis and lacunar ligament. It is reflected inferolaterally from the pubic tubercle as the arched falciform margin, which forms the superior, lateral and inferior boundaries of the saphenous opening: this margin adheres to the anterior layer of the femoral sheath, and the cribriform fascia is attached to it. The falciform margin is considered to have superior and inferior cornua. The inferior cornu is well defined, and is continuous behind the long saphenous vein with the deep stratum of the fascia lata.
The deep stratum is medial to the saphenous opening and is continuous with the superficial stratum at its lower margin. Traced upwards, it covers pectineus, adductor longus and gracilis, passes behind the femoral sheath, with which it is blends, and continues to the pecten pubis.
Fascial compartments
There are three functional groups of muscle in the thigh, namely, anterior (extensor), posterior (flexor) and medial (adductor). The anterior and posterior groups occupy separate osteofascial compartments that are limited peripherally by the fascia lata and separated from each other by the femur and the medial and lateral intermuscular septa (Fig. 80.2). The adductor muscles, though distinct in terms of function and innervation, do not possess a separate compartment limited by fascial planes. Nevertheless it is customary to speak of three compartments: anterior, posterior and medial. The muscles of the three compartments are described below. Adductor magnus, adductor longus and pectineus could each be considered to be constituents of two compartments, i.e. adductor magnus in the posterior and the medial compartments, and adductor longus and pectineus in the anterior and the medial compartments.
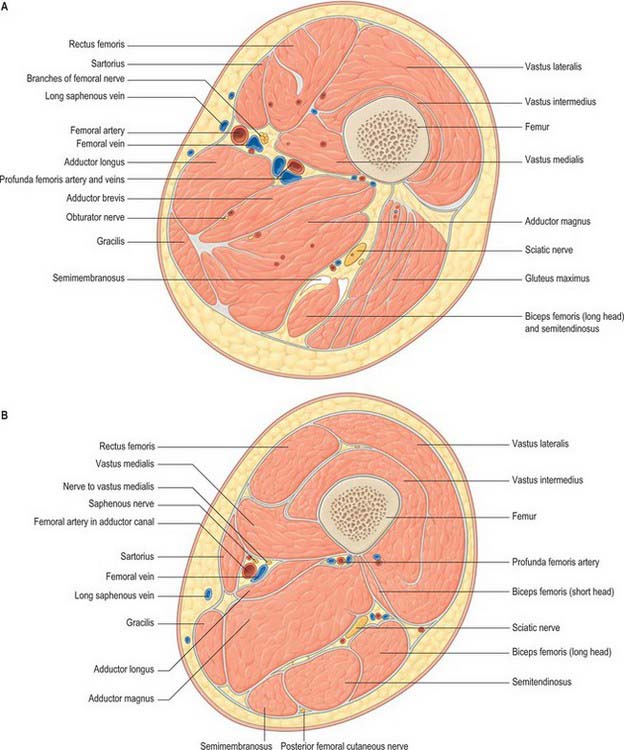
Fig. 80.2 Transverse (axial) section through the thigh. A, At the level of the apex of the femoral triangle; B, at the level of the mid-thigh.
The nerve supply to the compartments of the thigh mainly follows the ‘one compartment – one nerve’ principle. Thus the femoral nerve supplies the anterior compartment muscles, the obturator nerve supplies the medial compartment muscles, and the sciatic nerve supplies those in the posterior compartment. The dual functional and compartmental attribution of adductor magnus and adductor longus and pectineus are reflected in their dual nerve supplies.
In contrast to the motor innervation, the arterial supply to the compartmental muscle groups does not exhibit such a direct relationship. All groups receive a supply from the femoral system, particularly from the profunda femoris and its branches. The adductors receive a contribution from the obturator artery, and the hamstrings receive a proximal supply from the inferior gluteal artery. Further details are given in the descriptions of the individual muscles.
Femoral sheath
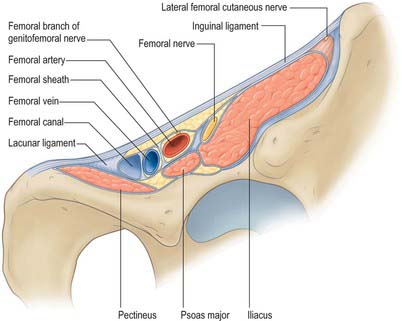
The femoral sheath is a funnel-shaped distal prolongation of extraperitoneal fascia, formed of transversalis fascia anterior to the femoral vessels, and of the iliac fascia posteriorly (Fig. 80.3). It is wider proximally and its tapered distal end fuses with the vascular adventitia 3 or 4 cm distal to the inguinal ligament. At birth the sheath is shorter; it elongates when extension at the hips becomes habitual. The femoral branch of the genitofemoral nerve perforates its lateral wall. The medial wall slopes laterally and is pierced by the long (great) saphenous vein and lymphatic vessels. Like the carotid sheath, the femoral sheath encloses a mass of connective tissue in which the vessels are embedded. Three compartments are described: a lateral one containing the femoral artery, an intermediate one for the femoral vein, and a medial compartment, the femoral canal, which contains lymph vessels and an occasional lymph node embedded in areolar tissue. The presence of this canal allows the femoral vein to distend. The canal is conical and approximately 1.25 cm in length. Its proximal (wider) end, termed the femoral ring, is bounded in front by the inguinal ligament, behind by pectineus and its fascia and the pectineal ligament, medially by the crescentic, lateral edge of the lacunar ligament and laterally by the femoral vein. The spermatic cord, or the round ligament of the uterus, is just above its anterior margin, while the inferior epigastric vessels are near its anterolateral rim. It is larger in women than in men: this is due partly to the relatively greater width of the female pelvis and partly to the smaller size of the femoral vessels in women. The ring is filled by condensed extraperitoneal tissue, the femoral septum, which is covered on its proximal aspect by the parietal peritoneum. The femoral septum is traversed by numerous lymph vessels that connect the deep inguinal to the external iliac lymph nodes.
Iliac fascia
The iliac fascia covers psoas and iliacus. It is thin above, but thickens progressively towards the inguinal ligament. The part covering psoas is thickened above as the medial arcuate ligament. Medially, the fascia over psoas is attached by a series of fibrous arches to the intervertebral discs, the margins of vertebral bodies, and the upper part of the sacrum. Laterally, it blends with the fascia anterior to quadratus lumborum above the iliac crest, and with the fascia covering iliacus below the crest.
The iliac part is connected laterally to the whole of the inner lip of the iliac crest and medially to the pelvic brim, where it blends with the periosteum. It is attached to the iliopubic ramus, where it receives a slip from the tendon of psoas minor, when that muscle is present. The external iliac vessels are anterior to the fascia but the branches of the lumbar plexus are posterior. The fascia is separated from the peritoneum by loose extraperitoneal tissue. Lateral to the femoral vessels, the iliac fascia is continuous with the posterior margin of the inguinal ligament and the transversalis fascia. Medially it passes behind the femoral vessels to become the pectineal fascia, attached to the pecten pubis. At the junction of its lateral and medial parts it is attached to the iliopubic ramus and the capsule of the hip joint. It thus forms a septum between the inguinal ligament and the hip bone, dividing the space here into a lateral part, the muscular space, containing psoas major, iliacus and the femoral nerve, and a medial part, the vascular space, transmitting the femoral vessels (Fig. 80.3). The iliac fascia continues downward to form the posterior wall of the femoral sheath.
Obturator membrane
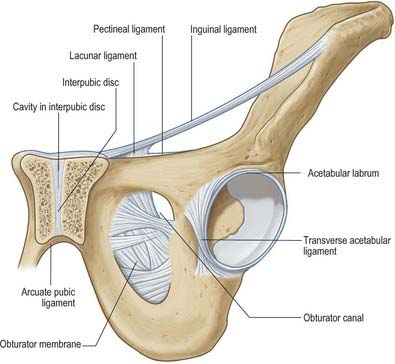
The obturator membrane (Fig. 80.4) is a thin aponeurosis that closes (obturates) most of the obturator foramen, leaving a superolateral aperture, the obturator canal, through which the obturator vessels and nerve leave the pelvis and enter the thigh. The membrane is attached to the sharp margin of the obturator foramen except at its inferolateral angle, where it is fixed to the pelvic surface of the ischial ramus, i.e. internal to the foramen. Its fibres are arranged mainly transversely in interlacing bundles; the uppermost bundle, which is attached to the obturator tubercles, completes the obturator canal. The outer and inner surfaces of the obturator membrane provide attachment for the obturator externus and internus respectively. Some fibres of the pubofemoral ligament of the hip joint are attached to the outer surface.
BONE
The pelvic girdle is an entity consisting of the two hip bones and the sacrum (strictly speaking, the sacrum is part of the vertebral column). These bones are virtually incapable of independent movement except in the female during parturition. The pelvic girdle is massively constructed and serves as a weightbearing and protective structure, an attachment for trunk and limb muscles, and as the skeletal framework of the birth canal.
HIP BONE
The hip bone is large, irregular, constricted centrally and expanded above and below (Fig. 80.5A,B). Its lateral surface has a deep, cup-shaped acetabulum, articulating with the femoral head, anteroinferior to which is the large, oval or triangular obturator foramen. Above the acetabulum the bone widens into an undulant plate surmounted by a sinuously curved iliac crest.
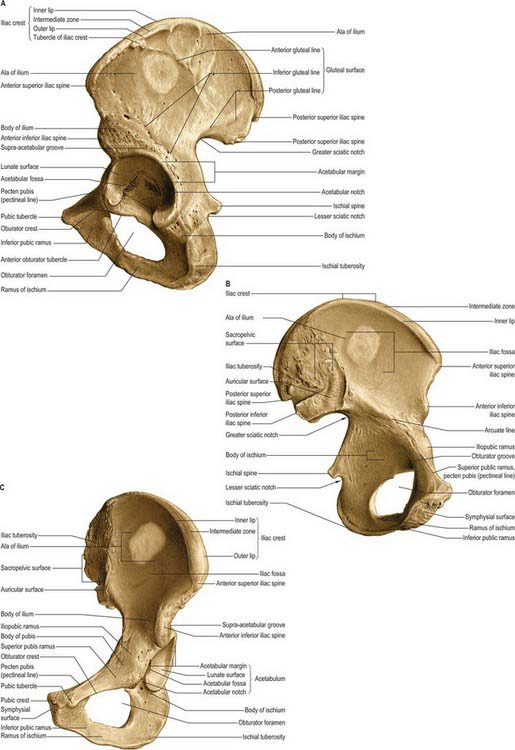
Fig. 80.5 Left hip bone. A, Outer aspect. B, Inner aspect. C, Anterosuperior view.
(From Sobotta 2006.)
The bone articulates in front with its fellow and posteriorly with the side of the sacrum to form the pelvic girdle. Each hip bone has three parts, ilium, ischium and pubis, connected to each other by cartilage in youth but united as one bone in adults. The principal union is in the acetabulum. The ilium includes the upper acetabulum and expanded area above it; the ischium includes the lower acetabulum and bone posteroinferior to it; the pubis forms the anterior acetabulum, separating the ilium from ischium, and the anterior median region where the pubes meet.
Acetabulum
The acetabulum (Fig. 80.5A,C) is an approximately hemispherical cavity situated about the centre of the lateral aspect of the hip bone. It faces anteroinferiorly and is circumscribed by an irregular margin deficient inferiorly at the acetabular notch. The acetabular fossa forms the central floor and is rough and non-articular. The articular lunate surface is widest above (the ‘dome’), where weight is transmitted to the femur. Consequently, fractures through this region tend to be associated with unsatisfactory outcomes. All three components of the hip bone contribute to the acetabulum, although unequally. The pubis forms the anterosuperior fifth of the articular surface, the ischium forms the floor of the fossa and rather more than the posteroinferior two-fifths of the articular surface, and the ilium forms the remainder. Occasionally a linear defect may be seen to cross the acetabular surface from the superior border to the acetabular fossa This does not correspond to any junction between the main morphological parts of the hip bone.
Obturator foramen
The obturator foramen lies below and slightly anterior to the acetabulum, between the pubis and ischium. It is bordered above by the grooved obturator surface of the superior pubic ramus, medially by the pubic body and its inferior ramus, below by the ischial ramus, and laterally by the anterior border of the ischial body, including the margin of the acetabulum. The foramen is almost closed by the obturator membrane (see above), which is attached to its margins, except superolaterally, where a communication remains between the pelvis and thigh. This free edge of the membrane is attached to an anterior obturator tubercle at the anterior end of the inferior border of the superior pubic ramus, and a posterior obturator tubercle on the anterior border of the acetabular notch; these tubercles are sometimes indistinct. Since the tubercles lie in different planes and the obturator groove crosses the upper border of the foramen, the acetabular rim is in fact a spiral. The foramen is large and oval in males, but smaller and nearly triangular in females.
Structure
The thicker parts of the hip bone are trabecular, encased by two layers of compact bone, while the thinner parts, as in the acetabulum and central iliac fossa, are often translucent and consist of a single lamina of compact bone. In the upper acetabulum and along the arcuate line, i.e. the route of weight transmission from the sacrum to the femur, the amount of compact bone is increased and the subjacent trabecular bone displays two sets of pressure lamellae. These start together near the upper auricular surface and diverge to meet two strong buttresses of compact bone, from which two similar sets of lamellar arches start and converge on the acetabulum. The anterior part of the iliac crest has been much studied with regard to distribution of cortical and trabecular bone. For a survey of these studies consult Whitehouse (1977). Additionally, Whitehouse’s observations, based on scanning electron micrography, indicate that the cortical bone is very porous, being only 75% bone, decreasing to 35% near the anterior superior iliac spine. Denser cortical bone starts at the margins of the crest and thickens rapidly below it on both aspects of the iliac blade.
Studies of the internal stresses within the hip bone have revealed a pattern of trabeculae that corresponds well with the theoretically expected patterns of stress trajectories (Holm 1980). These patterns are considerably more complex than in any other major bone. Stresses are higher in the acetabular than in the iliac region. In the ilium, the pelvic surface is subjected to considerably less stress than is the gluteal surface.
Vascular supply
In the infant, nutrient arteries are clearly demonstrable for each component of the hip bone. Each nutrient artery branches in fan-like fashion within its bone of supply (Crock 1996). Later, a periosteal arterial network develops, with contributions from numerous local arteries (see under individual bones).
Innervation
Periosteal innervation is by a network of nerves derived from branches of local nerves. These nerves also supply muscles attaching to the periosteum and the joints involving the hip bone. Autonomic nerves accompany nutrient arteries and branch within the bone.
Ossification
Ossification (Figs 80.6, 80.7) is by three primary centres, one each for the ilium, ischium and pubis. The iliac centre appears above the greater sciatic notch prenatally at about the ninth week; the ischial centre in its body in the fourth month, and the pubic centre in its superior ramus between the fourth and fifth months. At birth the whole iliac crest, the acetabular floor and inferior margin are cartilaginous. Gradual ossification of the three components of the acetabulum results in a triradiate cartilaginous stem extending medially to the pelvic surface as a Y-shaped epiphysial plate between the ilium, ischium and pubis, and including the anterior inferior iliac spine. Cartilage along the inferior margin also covers the ischial tuberosity, forms conjoined ischial and pubic rami, and continues to the pubic symphysial surface and along the pubic crest to its tubercle.
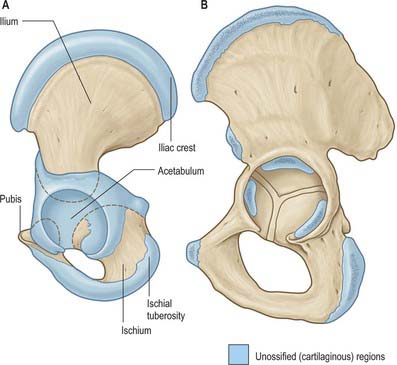
Fig. 80.6 The hip bone. A, At birth. B, In adolescence; more heavily stippled areas indicate the secondary centres of ossification.
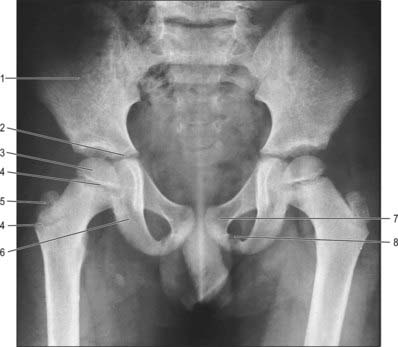
Fig. 80.7 Anteroposterior radiograph of the pelvis of a boy aged 7. 1. Ilium. 2. Part of triradiate growth cartilage. 3. Superior femoral epiphysis. 4. Cartilaginous growth plates. 5. Ossifying greater trochanter. 6. Ischium. 7. Pubis. 8. Cartilage between pubic and ischial rami.
The ossifying ischium and pubis fuse to form a continuous ischiopubic ramus at the seventh or eighth year. Secondary centres, other than for the acetabulum, appear about puberty and fuse between the 15th and 25th years. There are usually two for the iliac crest (which fuse early), and single centres for the ischial tuberosity (in cartilage close to the inferior acetabular margin and spreading forwards), anterior inferior iliac spine (although it may ossify from the triradiate cartilage), and symphysial surface of the pubis (the pubic tubercle and crest may have separate centres). Progression of ossification of the iliac crest in girls is an index of skeletal maturity and is useful in determining the optimal timing of surgery for spinal deformity.
Between the ages of 8 and 9 years three major centres of ossification appear in the acetabular cartilage. The largest appears in the anterior wall of the acetabulum and fuses with the pubis, the second in the iliac acetabular cartilage superiorly, fusing with the ilium, and the third in the ischial acetabular cartilage posteriorly, fusing with the ischium. At puberty these epiphyses expand towards the periphery of the acetabulum and contribute to its depth (Ponseti 1978). Fusion between the three bones within the acetabulum occurs between the 16th and 18th years. Delaere et al (1992) have suggested that ossification of the ilium is similar to that of a long bone, possessing three cartilaginous epiphyses and one cartilaginous process, although it tends to undergo osteoclastic resorption comparable with that of cranial bones. During development the acetabulum increases in breadth at a faster rate than it does in depth.
Avulsion fractures of pelvic apophyses may occur from excessive pull on tendons, usually in athletic adolescents. The most frequent examples of such injuries are those to the ischial tuberosity (hamstrings) and anterior inferior iliac spine (rectus femoris).
Pubis
Topography
The pubis (Figs 80.5, 80.6) is the ventral part of the hip bone and forms a median cartilaginous pubic symphysis with its fellow. The body of the pubis occupies the anteromedial part of the bone, and from the body a superior ramus passes up and back to the acetabulum and an inferior ramus passes back, down and laterally to join the ischial ramus inferomedial to the obturator foramen.
Body
The body, anteroposteriorly compressed, has anterior, posterior and symphysial (medial) surfaces and an upper border, the pubic crest. The anterior surface also faces inferolaterally; it is rough superomedially and smooth elsewhere, giving attachment to medial femoral muscles. The smooth posterior surface faces upwards and backwards as the oblique anterior wall of the lesser pelvis and is related to the urinary bladder. The symphysial surface is elongated and oval, united by cartilage to its fellow at the pubic symphysis. Denuded of cartilage it has an irregular surface of small ridges and furrows or nodular elevations, varying considerably with age, features which are of forensic interest. The pubic crest is the rounded upper border of the body which overhangs the anterior surface; its lateral end is the rounded pubic tubercle. Both crest and tubercle are palpable, the latter partly obscured in males by the spermatic cord that crosses above it from the scrotum to the abdomen. The pubic rami diverge posterolaterally from the superolateral corners of the body.
The anterior surface of the pubic body faces the femoral adductor region. The anterior pubic ligament attaches to its medial part along a rough strip, which is wider in females. The posterior surface is separated from the urinary bladder by retropubic fat. The puboprostatic ligaments are attached to this surface medial to levator ani.
Superior pubic ramus
The superior pubic ramus passes upwards, backwards and laterally from the body, superolateral to the obturator foramen, to reach the acetabulum. It is triangular in section and has three surfaces and borders. Its anterior, pectineal surface, tilted slightly up, is triangular in outline and extends from the pubic tubercle to the iliopubic ramus. It is bounded in front by the rounded obturator crest and behind by the sharp pecten pubis (pectineal line) which, with the crest, is the pubic part of the linea terminalis (i.e. anterior part of the pelvic brim). The posterosuperior, pelvic surface, medially inclined, is smooth and narrows into the posterior surface of the body, which is bounded above by the pecten pubis and below by a sharp inferior border. The obturator surface, directed down and back, is crossed by the obturator groove sloping down and forwards. Its anterior limit is the obturator crest and its posterior limit is the inferior border.
Inferior pubic ramus
The inferior pubic ramus, an inferolateral process of the body, descends inferolaterally to join the ischial ramus medial to, and below, the obturator foramen. The union may be locally thickened, but not obviously so in adults. The ramus has two surfaces and borders. The anterolateral surface, continuous above with that of the pubic body, faces the thigh and is marked by muscles. It is limited laterally by the margin of the obturator foramen and, medially, by the rough anterior border. The posteromedial surface is continuous above with that of the body and is transversely convex: its medial part is often everted in males and gives attachment to the crus of the penis. This surface faces the perineum medially, its smooth lateral part tilted up towards the pelvic cavity.
The internal surface is indistinctly divided into medial, intermediate and lateral areas. The medial area faces inferomedially in direct contact with the crus of the penis or clitoris and is limited above and behind by an indistinct ridge for attachment of the fascia overlying the superficial perineal muscles. The medial margin of the ramus, strongly everted in males, provides attachment for the fascia lata and the membranous layer of the superficial perineal fascia.
Pubic tubercle
The pubic tubercle provides attachment to the medial end of the inguinal ligament. It forms part of the floor of the superficial inguinal ring and is crossed by the spermatic cord.
Pecten pubis
The pecten pubis is the sharp, superior edge of the pectineal surface. The conjoint tendon and lacunar ligament are attached at its medial end and a strong, fibrous, pectineal ligament is attached along the rest of its surface. The smooth pelvic surface is separated from parietal peritoneum only by areolar tissue, in which the lateral umbilical ligament descends forwards across the ramus and, laterally, the vas deferens passes backwards. The obturator groove, which is converted to a canal by the upper borders of the obturator membrane and obturator muscles, transmits the obturator vessels and nerve from the pelvis to the thigh. Some fibres of the pubofemoral ligament are attached to the lateral end of the obturator crest.
Muscle attachments
The tendon of adductor longus is attached on the anterior (external) surface of the body, below the pubic crest. Below adductor longus, gracilis is attached to a line near the lower margin extending down onto the inferior ramus. Above gracilis, adductor brevis is attached to the body and inferior ramus. Above again, obturator externus is attached to the anterior surface, spreading onto inferior pubic and ischial rami. Adductor magnus usually extends from the ischial ramus on to the lower part of the inferior pubic ramus between adductor brevis and obturator externus. Pectineus is attached to the pectineal surface of the superior ramus along its upper part. Ascending loops of cremaster are attached to the pubic tubercle. The lateral part of rectus abdominis and, inferiorly, pyramidalis, are attached lateral to the tubercle, on the pubic crest. Medially the crest is crossed by the medial part of rectus abdominis, ascending from ligamentous fibres that interlace in front of the pubic symphysis. Anterior fibres of levator ani are attached on the posterior (internal) surface of the body near its centre. More laterally, obturator internus is attached on this surface, extending onto both rami. Psoas minor, when present, is attached near the centre of the pecten pubis.
Vascular supply
The pubis is supplied by a periosteal anastomosis of branches from the obturator, inferior epigastric and medial circumflex femoral arteries. The superficial and deep external pudendal arteries may also contribute. Multiple vascular foramina are present, mainly at the lateral (acetabular) end of the bone, but there is no consistently placed nutrient foramen.
Ilium
Topography
The ilium has upper and lower parts and three surfaces (Figs 80.5, 80.6). The smaller, lower part forms a little less than the upper two-fifths of the acetabulum. The upper part is much expanded, and has gluteal, sacropelvic and iliac (internal) surfaces. The posterolateral gluteal surface is an extensive rough area; the anteromedial iliac fossa is smooth and concave; the sacropelvic surface is medial and posteroinferior to the fossa, from which it is separated by the medial border.
Iliac crest
The iliac crest is the superior border of the ilium. It is convex upwards but sinuous from side to side, being internally concave in front, and convex behind. Its ends project as anterior and posterior superior iliac spines. The anterior superior iliac spine is palpable at the lateral end of the inguinal fold; the lateral end of the inguinal ligament is attached to the anterior superior iliac spine. The posterior superior iliac spine is not palpable but is often indicated by a dimple, approximately 4 cm lateral to the second sacral spinous process, above the medial gluteal region (buttock).
The crest has ventral and dorsal segments. The ventral occupies slightly more than the anterior two-thirds of the crest and its prominence is associated with changes in iliac form as a result of the emergence of the upright posture; the dorsal segment, which occupies approximately the posterior third in man, is a feature of all land vertebrates. The ventral segment of the crest has internal and external lips and the rough intermediate zone is narrowest centrally. The tubercle of the crest projects outwards from the outer lip approximately 5 cm posterosuperior to the anterior superior spine. The dorsal segment has two sloping surfaces separated by a longitudinal ridge ending at the posterior superior spine. The summit of the crest, a little behind its midpoint, is level with the fourth lumbar vertebral body. The interosseous and posterior sacroiliac ligaments arise from the medial margin of the dorsal segment.
Anterior border
The anterior border descends to the acetabulum from the anterior superior spine. Superiorly it is concave forwards. Inferiorly, immediately above the acetabulum, is a rough anterior inferior iliac spine, which is divided indistinctly into an upper area for the straight part of rectus femoris and a lower area extending laterally along the upper acetabular margin to form a triangular impression for the proximal end of the iliofemoral ligament.
Posterior border
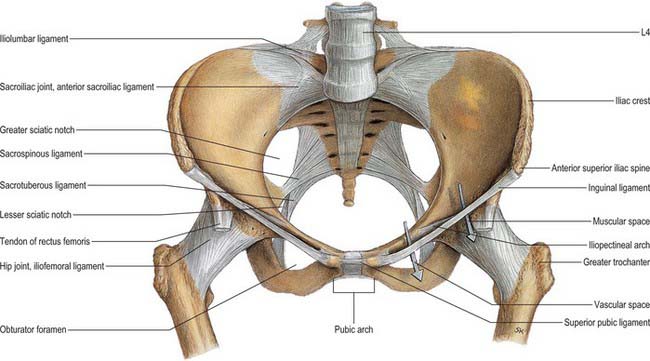
The posterior border is irregularly curved and descends from the posterior superior spine, at first forwards, with a posterior concavity forming a small notch. At the lower end of the notch is a wide, low projection, the posterior inferior iliac spine. Here the border turns almost horizontally forwards for approximately 3 cm, then down and back to join the posterior ischial border. Together these borders form a deep notch, the greater sciatic notch, that is bounded above by the ilium and below by the ilium and ischium. The upper fibres of the sacrotuberous ligament are attached to the upper part of the posterior border. The superior rim of the notch is related to the superior gluteal vessels and nerve. The lower margin of the greater sciatic notch is covered by piriformis and related to the sciatic nerve.
Medial border
The medial border separates the iliac fossa and the sacropelvic surface. It is indistinct near the crest, rough in its upper part, then sharp where it bounds an articular surface for the sacrum, and finally rounded. The latter part is the arcuate line, which inferiorly reaches the posterior part of the iliopubic ramus, marking the union of the ilium and pubis.
Gluteal surface
The gluteal surface, facing inferiorly in its posterior part and laterally and slightly downwards in front, is bounded above by the iliac crest, below by the upper acetabular border and by the anterior and posterior borders. It is rough and curved, convex in front, concave behind, and marked by three gluteal lines. The posterior gluteal line is shortest, descending from the external lip of the crest approximately 5 cm in front of its posterior limit and ending in front of the posterior inferior iliac spine. Above, it is usually distinct, but inferiorly it is ill-defined and frequently absent. The anterior gluteal line, the longest, begins near the midpoint of the superior margin of the greater sciatic notch and ascends forwards into the outer lip of the crest, a little anterior to its tubercle. The inferior gluteal line, seldom well-marked, begins posterosuperior to the anterior inferior iliac spine, curving posteroinferiorly to end near the apex of the greater sciatic notch. Between the inferior gluteal line and the acetabular margin is a rough, shallow groove. Behind the acetabulum the lower gluteal surface is continuous with the posterior ischial surface, the conjunction marked by a low elevation.
The articular capsule is attached to an area adjoining the acetabular rim, most of which is covered by gluteus minimus. Posteroinferiorly, near the union of the ilium and ischium, the bone is related to piriformis.
Iliac fossa
The iliac fossa, the internal concavity of the ilium, faces anterosuperiorly. It is limited above by the iliac crest, in front by the anterior border and behind by the medial border, separating it from the sacropelvic surface. It forms the smooth and gently concave posterolateral wall of the greater pelvis. Below it is continuous with a wide shallow groove which is bounded laterally by the anterior inferior iliac spine and medially by the iliopubic ramus.
The wide groove between the anterior inferior iliac spine and the iliopubic ramus is occupied by the converging fibres of iliacus laterally and the tendon of psoas major medially: the tendon is separated from the underlying bone by a bursa. The right iliac fossa contains the caecum, and often the vermiform appendix and terminal ileum. The left iliac fossa houses the terminal part of the descending colon and the proximal sigmoid colon.
Sacropelvic surface
The sacropelvic surface, the posteroinferior part of the medial iliac surface, is bounded posteroinferiorly by the posterior border, anterosuperiorly by the medial border, posterosuperiorly by the iliac crest and anteroinferiorly by the line of fusion of the ilium and ischium. It is divided into iliac tuberosity, auricular and pelvic surfaces. The iliac tuberosity, a large, rough area below the dorsal segment of the iliac crest, shows cranial and caudal areas separated by an oblique ridge and connected to the sacrum by the interosseous sacroiliac ligament. The sacropelvic surface gives attachment to the posterior sacroiliac ligaments and, behind the auricular surface, to the interosseous sacroiliac ligament. The iliolumbar ligament is attached to its anterior part. The auricular surface, immediately anteroinferior to the tuberosity, articulates with the lateral sacral mass. Shaped like an ear, its widest part is anterosuperior, its ‘lobule’ posteroinferior and on the medial aspect of the posterior inferior spine. Its edges are well defined, but the surface, though articular, is rough and irregular. It articulates with the sacrum and is reciprocally shaped. The anterior sacroiliac ligament is attached to its sharp anterior and inferior borders. The narrow part of the pelvic surface, between the auricular surface and the upper rim of the greater sciatic notch, often shows a rough preauricular sulcus (that is usually better defined in females) for the lower fibres of the anterior sacroiliac ligament. For the reliability of this feature as a sex discriminant, refer to Finnegan (1978) and Brothwell & Pollard (2001). The pelvic surface is anteroinferior to the acutely recurved part of the auricular surface, and contributes to the lateral wall of the lesser pelvis. Its upper part, facing down, is between the auricular surface and the upper limb of the greater sciatic notch. Its lower part faces medially and is separated from the iliac fossa by the arcuate line. Anteroinferiorly it extends to the line of union between the ilium and ischium. Though usually obliterated, it passes from the depth of the acetabulum to approximately the middle of the inferior limb of the greater sciatic notch.
Muscle attachments
The attachment of sartorius extends down the anterior border below the anterior superior iliac spine. The iliac crest gives attachment to the anterolateral and dorsal abdominal muscles, and to the fasciae and muscles of the lower limb. The fascia lata and iliotibial tract are attached to the outer lip and tubercle of its ventral segment. Tensor fasciae latae is attached anterior to the tubercle. The lower fibres of external oblique and, just behind the summit of the crest, the lowest fibres of latissimus dorsi are attached to its anterior two-thirds. A variable interval exists between the most posterior attachment of external oblique and the most anterior attachment of latissimus dorsi, and here the crest forms the base of the lumbar triangle. Internal oblique is attached to the intermediate area of the crest. Transversus abdominis is attached to the anterior two-thirds of the inner lip of the crest, and behind this to the thoracolumbar fascia and quadratus lumborum. The highest fibres of gluteus maximus are attached to the dorsal segment of the crest on its lateral slope. Erector spinae arises from the medial slope of the dorsal segment. The straight part of rectus femoris is attached to the upper area of the anterior inferior spine. Some fibres of piriformis are attached in front of the posterior inferior spine on the upper border of the greater sciatic notch.
The gluteal surface is divided by three gluteal lines into four areas. Behind the posterior line, the upper rough part gives attachment to the upper fibres of gluteus maximus and the lower, smooth region to part of the sacrotuberous ligament and iliac head of piriformis. Gluteus medius is attached between the posterior and anterior lines, below the iliac crest, and gluteus minimus is attached between the anterior and inferior lines. The fourth area, below the inferior line, contains vascular foramina.
The reflected head of rectus femoris attaches to a curved groove above the acetabulum. Iliacus is attached to the upper two-thirds of the iliac fossa and is related to its lower third. The medial part of quadratus lumborum is attached to the anterior part of the sacropelvic surface, above the iliolumbar ligament. Piriformis is sometimes partly attached lateral to the preauricular sulcus, and part of obturator internus is attached to the more extensive remainder of the pelvic surface.
Vascular supply
Branches of the iliolumbar artery run between iliacus and bone; one or more enter large nutrient foramina lying posteroinferiorly in the iliac fossa. The superior gluteal, obturator and superficial circumflex iliac arteries contribute to the periosteal supply. The obturator artery may supply a nutrient branch. Vascular foramina on the iliac gluteal aspect may lead into large vascular canals in the bone.
Ischium
Topography
The ischium, the inferoposterior part of the hip bone, has a body and ramus. The body has upper and lower ends and femoral, posterior and pelvic surfaces (Figs 80.5, 80.6). Above, it forms the posteroinferior part of the acetabulum; below, its ramus ascends anteromedially at an acute angle to meet the inferior pubic ramus, thereby completing the boundary of the obturator foramen. The ischiofemoral ligament is attached to the lateral border below the acetabulum.
The femoral surface faces downwards, forwards and laterally towards the thigh. It is bounded in front by the margin of the obturator foramen. The lateral border, indistinct above but well defined below, forms the lateral limit of the ischial tuberosity. At a higher level the femoral surface is covered by piriformis, from which it is partially separated by the sciatic nerve and the nerve to quadratus femoris. The posterior surface, facing superolaterally, is continuous above with the iliac gluteal surface, and here a low convexity follows the acetabular curvature. Inferiorly, this surface forms the upper part of the ischial tuberosity, above which is a wide, shallow groove on its lateral and medial aspects. Above the ischial tuberosity the posterior surface is crossed by the tendon of obturator internus and the gemelli. The nerve to quadratus femoris lies between these structures and the bone. The ischial tuberosity is a large, rough area on the lower posterior surface and inferior extremity of the ischium. Though obscured by gluteus maximus in hip extension, it is palpable in flexion. It is 5 cm from the midline and about the same distance above the gluteal fold. It is elongated, widest above, and tapers inferiorly. The ischial posterior aspect lies between the lateral and posterior borders. The posterior border blends above with that of the ilium, helping to complete the inferior rim of the greater sciatic notch, the posterior end of which has a conspicuous ischial spine. Below this, the rounded border forms the floor of the lesser sciatic notch, between the ischial spine and tuberosity. The pelvic surface is smooth and faces the pelvic cavity; inferiorly it forms part of the lateral wall of the ischio-anal fossa.
Ischial ramus
The ischial ramus has anteroinferior and posterior surfaces continuous with the corresponding surfaces of the inferior pubic ramus: the anteroinferior surface is roughened by the attachment of the medial femoral muscles. The smooth posterior surface is partly divided into perineal and pelvic areas, like the inferior pubic ramus. The upper border completes the obturator foramen; the rough lower border, together with the medial border of the inferior pubic ramus, contributes to the pubic arch. The fascia overlying the superficial perineal muscles is attached below the ridge between the perineal and pelvic areas of the posterior surface of the ischial ramus. Above the ridge areas give attachment to the crus of the penis or clitoris and the sphincter urethrae. The lower border of the ramus provides an attachment for the fascia lata and a membranous layer of the superficial perineal fascia.
Ischial tuberosity
The ischial tuberosity is divided nearly transversely into upper and lower areas. The upper area is subdivided by an oblique line into a superolateral and an inferomedial part. The lower area, narrowing as it curves onto the inferior ischial aspect, is subdivided by an irregular vertical ridge into lateral and medial areas. The medial is covered by fibroadipose tissue that usually contains the ischial bursa of gluteus maximus, which supports the body in sitting. Medially the tuberosity is limited by a curved ridge that passes on to the ramus and which gives attachment to the sacrotuberous ligament and its falciform process.
Ischial spine
The ischial spine projects downwards and a little medially (Fig. 80.8). The sacrospinous ligament is attached to its margins, separating the greater from the lesser sciatic foramen. The ligament is crossed posteriorly by the internal pudendal vessels, pudendal nerve and the nerve to obturator internus.
Muscle attachments
Part of obturator externus is attached to the lower femoral surface of the ischial body. Part of obturator externus, the anterior fibres of adductor magnus and, near the lower border, gracilis, are all attached to the anterior surface of the ischial ramus. Between adductor magnus and gracilis the attachment of adductor brevis may descend from the inferior pubic ramus. The posterior surface is divided into pelvic and perineal areas. The pelvic area, facing back, has part of obturator internus attached to it. The perineal area faces medially; its upper part is related to the crus of the penis or clitoris, and its lower part gives attachment to sphincter urethrae, ischiocavernosus and the transverse superficial perineal muscle.
The ischial tuberosity gives attachment to the posterior femoral muscles. Quadratus femoris is attached along the upper part of its lateral border. The upper area of the tuberosity is subdivided by an oblique line into a superolateral part for semimembranosus and an inferomedial part for the long head of biceps femoris and semitendinosus. The lower area is subdivided by an irregular vertical ridge into lateral and medial areas. The larger lateral area is for part of adductor magnus. Superomedial to the tuberosity the posterior surface has a wide, shallow groove, usually covered by hyaline cartilage, with a bursa between it and the tendon of obturator internus. Gemellus inferior is attached to the lower margin of the groove, near the tuberosity. Gemellus superior is attached to the upper margin, near the ischial spine.
The pelvic surface of the ischial spine gives attachment to coccygeus (coextensive with the sacrospinous ligament) and to the most posterior fibres of levator ani. Obturator internus is attached to the upper part of the smooth pelvic ischial surface and converges on the lesser sciatic notch, covering the rest of this surface other than the pelvic aspect of the ischial spine: the muscle and its fascia separate the bone from the ischiorectal fossa.
Vascular supply
There are multiple vascular foramina at the acetabular margins, and a few are usually present on the pelvic surface. The bone is supplied by branches of the obturator, medial circumflex femoral and inferior gluteal arteries.
SKELETAL PELVIS AS A WHOLE
The term pelvis (‘basin’) is applied variously to the skeletal ring formed by the hip bones and the sacrum, the cavity therein, and even the entire region where the trunk and lower limbs meet. It is used here in the skeletal sense, to describe the irregular osseous girdle between the femoral heads and fifth lumbar vertebra. It is large because its primary function is to withstand the forces of body weight and musculature. In this section, its obstetric, forensic and anthropological significance will be considered.
The pelvis can be regarded as having greater and lesser segments, the true and false pelves. The segments are arbitrarily divided by an oblique plane passing through the sacral promontory posteriorly and the lineae terminales elsewhere. Each linea terminalis includes the iliac arcuate line, pectineal line (pecten), and pubic crest. The segments are continuous, and the parts of the body cavity that they enclose are also continuous through the pelvic inlet (pelvic inlet).
Greater pelvis
The greater pelvis consists of the ilium and pubis above the lineae terminales and the sacral base. This junctional zone is structurally massive and forms powerful arches from the acetabular fossae to the vertebral column around the visceral cavity, which is part of the abdomen. It has little anterior wall because of the pelvic inclination.
Pelvic inlet (superior pelvic aperture)
The pelvic inlet or brim may be round or oval, and is indented posteriorly by the sacral promontory. The pelvic brim is obstetrically important and has also long been measured for anthropological reasons, as has the pelvic cavity.
By convention, the pelvic inlet is described in three dimensions. The anteroposterior diameter (true conjugate) is measured between the midpoints of the sacral promontory and upper border of the symphysis pubis and on average is 10 cm in the adult male and 11.2 cm in the adult female. The transverse diameter is the maximum distance between similar points (assessed by eye) on opposite sides of the pelvic brim and is on average 12.5 cm in the male and 13.1 cm in the adult female. The oblique diameter is measured from the iliopubic ramus to the opposite sacroiliac joint and is on average 12 cm in the adult male and 12.5 cm in the adult female. These measurements vary with the individual and racial group.
Articulated bony pelvis
The lesser pelvis encloses a true basin when soft tissues of the pelvic floor are in place. Skeletally it is a narrower continuation of the greater pelvis, with irregular but more complete walls around its cavity. Of obstetric importance, it has a curved median axis, and superior and inferior openings. The superior opening is occupied by viscera. The pelvic floor, viscera and subjacent perineal sphincters close the inferior opening.
Cavity of the lesser pelvis
The cavity of the lesser pelvis is short, curved, and markedly longer in its posterior wall. Anteroinferiorly it is bounded by pubic bones, their rami and symphysis. Posteriorly it is bounded by the concave anterior sacral surface and coccyx. Laterally on each side its margins are the smooth quadrangular pelvic aspect of the fused ilium and ischium. The region so enclosed is the pelvic cavity proper, through which pass the rectum, bladder and parts of the reproductive organs. The cavity in females must also permit passage of the fetal head.
The pelvic cavity diameters are measured at approximately the mid level. The anteroposterior diameter is measured between the midpoints of the third sacral segment and posterior surface of the symphysis pubis and is about 10.5 cm in the male and 13 cm in the adult female. The transverse diameter is the widest transverse distance between the side walls of the cavity, and often the greatest transverse dimension in the whole cavity. It measures about 12 cm in the adult male and 12.5 cm in the adult female. The oblique diameter is the distance from the lowest point of one sacroiliac joint to the midpoint of the contralateral obturator membrane and measures about 11 cm in the male and 13.1 cm in the adult female. All measurements vary with the individual and racial group.
Pelvic outlet (inferior pelvic aperture)
Less regular in outline than the pelvic inlet, the pelvic outlet is indented behind by the coccyx and sacrum and bilaterally by the ischial tuberosities. Its perimeter thus consists of three wide arcs. Anteriorly is the pubic arch, between the converging ischiopubic rami. Posteriorly and laterally on both sides are the sciatic notches between the sacrum and ischial tuberosities. These are divided by the sacrotuberous and sacrospinous ligaments into greater and lesser sciatic foramina.
With ligaments included, the pelvic outlet is rhomboidal. Its anterior limbs are the ischiopubic rami (joined by the inferior pubic ligament) and its posterior margins are the sacrotuberous ligaments, with the coccyx in the midline. The outlet is thus not rigid in its posterior half, being limited by ligaments and the coccyx, all slightly yielding. Even with the sacrum taken as the posterior midline limit (more reliable for measurement), there may be slight mobility at the sacroiliac joints. Note also that a plane of the pelvic outlet is merely conceptual. The anterior, ischiopubic part has a plane which is inclined down and back to a transverse line between the lower limits of the ischial tuberosities, and the posterior half has a plane approximating to the sacrotuberous ligaments, sloping down and forwards to the same line.
Three measurements are made for the pelvic outlet. The anteroposterior diameter is usually measured from the coccygeal apex to the midpoint of the lower rim of the symphysis. The lowest sacral point may also be used (on average male 8 cm, female 12.5 cm). The transverse (bituberous) diameter is measured between the ischial tuberosities at the lower borders of their medial surfaces (on average male 8.5 cm, female 11.8 cm). The oblique diameter extends from the midpoint of the sacrotuberous ligament on one side to the contralateral ischiopubic junction (on average male 10 cm, female 11.8 cm). All measurements vary with the individual and racial group.
Other measurements
Apart from these main measurements, by consensus the basis of pelvic osteometry, other planes and measurements are used in obstetric practice. The plane of greatest pelvic dimensions is an obstetric concept. It represents the most capacious pelvic level, between the pelvic brim and midlevel plane, and corresponds with the latter anteriorly at the middle part of the symphysis pubis and posteriorly at the level of the second and third sacral segments.
The plane of least dimensions is said to be at about midpelvic level. Its transverse diameter is between the apices of the ischial spines. This measurement is about 9.5 cm in an adult female and is just wide enough to allow passage of the biparietal diameter of a fetal head (about 9 cm). Not surprisingly, most difficulty in parturition occurs here.
The above measurements are sometimes made in clinical practice using X-ray or MRI pelvimetry. Precise measurement is not possible without radiological techniques, and even these do not take into account the soft tissues. In the past, measurements were made at physical and vaginal examination. However, these manual measurements have proved to be of little clinical value and are now obsolete.
Morphological classification of pelves
Interest in the dimensions described above is primarily obstetric and, less frequently, forensic. All pelvic measurements display individual variation and the values quoted are means from limited surveys. Sexual and racial differences also occur. These measurements have been analysed by many anatomists, anthropologists, obstetricians and radiologists in attempts to classify human pelves, especially female. The four most common terms used today are gynaecoid, anthropoid, platypelloid and android. The gynaecoid pelvis is the traditional Western female pelvis with a heart-shaped brim and the range of measurements quoted above. An anthropoid pelvis has a larger midcavity and a wide anteroposterior inlet which is oval in shape: it is more common in women of African origin and may be associated with a ‘high assimilation’ pelvis where there is an additional lumbar vertebra. A platypelloid pelvis is flat and oval from side to side at the brim: it is a contracted pelvis that is rarely seen nowadays, having previously been associated with rickets. An android pelvis has a triangular brim and is the shape of a male pelvis.
Pelvic axes and inclination
The axis of the superior pelvic aperture traverses its centre at right angles to its plane, directed down and backwards (Fig. 80.9). When prolonged (projected) it passes through the umbilicus and midcoccyx. An axis is similarly established for the inferior aperture: projected upwards it impinges on the sacral promontory. Axes can likewise be constructed for any plane, and one for the whole cavity is a concatenation of an infinite series of such lines (Fig. 80.9). The fetal head, however, descends in the axis of the inlet as far as the level of the ischial spines; it is then directed forwards into the axis of the vagina at right angles to that axis. The form of this pelvic axis and the disparity in depth between the anterior and posterior contours of the cavity are prime factors in the mechanism of fetal transit in the pelvic canal.
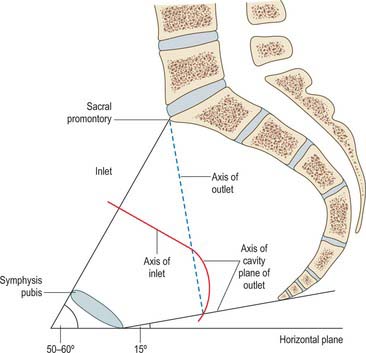
Fig. 80.9 Median sagittal section through the female pelvis, showing the planes of the inlet and outlet and the axis of the pelvic cavity.
In the standing position the pelvic canal curves obliquely backwards relative to the trunk and abdominal cavity. The whole pelvis is tilted forwards, the plane of the pelvic brim making an angle of 50–60° with the horizontal. The plane of the pelvic outlet is tilted to about 15°. Strictly, the pelvic outlet has two planes, an anterior passing backwards from the pubic symphysis and a posterior passing forwards from the coccyx, both descending to meet at the intertuberous line. In standing, the pelvic aspect of the symphysis pubis faces nearly as much upwards as backwards and the sacral concavity is directed anteroinferiorly. The front of the symphysis and anterior superior iliac spines are in the same vertical plane. In sitting, body weight is transmitted through inferomedial parts of the ischial tuberosities, with variable soft tissues intervening. The anterior superior iliac spines are in a vertical plane through the acetabular centres, and the whole pelvis is tilted back with the lumbosacral angle somewhat diminished at the sacral promontory.
Pelvic mechanism
The skeletal pelvis supports and protects the contained viscera, but is primarily part of the lower limbs, affording wide attachment for leg and trunk muscles. It constitutes the major mechanism for transmitting the weight of the head, trunk and upper limbs to the lower limbs. It may be considered as two arches divided by a coronal transacetabular plane. The posterior arch, chiefly concerned in transmitting weight, consists of the upper three sacral vertebrae and strong pillars of bone from the sacroiliac joints to the acetabular fossae. The anterior arch, formed by the pubic bones and their superior rami, connects these lateral pillars as a tie beam to prevent separation; it also acts as a compression strut against medial femoral thrust. The sacrum, as the summit of the posterior arch, is loaded at the lumbosacral joint. Theoretically this force has two components, one thrusting the sacrum downwards and backwards between the iliac bones, the other thrusting its upper end downwards and forwards. Sacral movements are regulated by osseous shape and massive ligaments. The first component therefore acts against the wedge, its tendency to separate iliac bones resisted by the sacroiliac and iliolumbar ligaments and symphysis pubis.
Vertical coronal sections through the sacroiliac joints suggest division of the (synovial) articular region of the sacrum into three segments. In the anterosuperior segment, involving the first sacral vertebra, the articular surfaces are slightly sinuous and almost parallel. In the middle segment the posterior width between the articular markings is greater than the anterior, and centrally a sacral concavity fits a corresponding iliac convexity, an interlocking mechanism relieving the strain on the ligaments produced by body weight. In the posteroinferior segment the anterior sacral width is greater than the posterior and here its sacral surfaces are slightly concave. Anteroinferior sacral dislocation by the second component (of force) is prevented, therefore, mainly by the middle segment, owing to its cuneiform shape and interlocking mechanism. However, some rotation occurs, in which the anterosuperior segment tilts down and the posteroinferior segment up. ‘Superior’ segmental movement is limited to a small degree by wedging but primarily by tension in the sacrotuberous and sacrospinous ligaments. In all movements the sacroiliac and iliolumbar ligaments and symphysis pubis resist separation of the iliac bones.
SEXUAL DIFFERENCES IN THE PELVIS
The pelvis provides the most marked skeletal differences between male and female. Distinction can be made even during fetal life, particularly in the subpubic arch. In infancy, dimensions of the whole pelvis are greater in males than in females, but the size of the pelvic cavity is usually greater in females. This distinction prevails in childhood, but the difference is maximal at about 22 months. Sexual differences in adults are divisible into metrical and non-metrical features: the range of most features overlaps between the sexes.
Differences are inevitably linked to function. While the primary pelvic function in both sexes is locomotor, the pelvis, particularly the lesser pelvis, is adapted to parturition in females, and these changes variably affect the proportions and dimensions of the greater pelvis. Since males are distinctly more muscular and therefore more heavily built, overall pelvic dimensions, such as the intercristal measurement (distance between the iliac crests), are greater, markings for muscles and ligaments more pronounced, and general architecture heavier. The male iliac crest is more rugged and more medially inclined at its anterior end; in females the crests are less curved in all parts. The iliac alae are more vertical in females, but do not ascend so far; the iliac fossae are therefore shallower and each iliopectineal line more vertical. These iliac peculiarities probably account for the greater prominence of female hips.
The male is relatively and absolutely more heavily built above the pelvis, with consequent differences at the lumbosacral and hip joints. The sacral basal articular facet for the fifth lumbar vertebra and intervening disc is more than a third of the total sacral basal width in males but less than a third in females, in whom the sacrum is also relatively broader, accentuating this difference. The female has relatively broader sacral alae. The male acetabulum is absolutely larger, and its diameter is approximately equal to the distance between its anterior rim and symphysis pubis. In females, acetabular diameter is usually less than this distance, not only because it is absolutely smaller but also because the anterolateral wall of the cavity is comparatively and often absolutely wider. The height of the female symphysis and adjoining parts of the pubis and ischium, which form the anterior pelvic wall, are also absolutely less, producing a somewhat triangular obturator foramen, which is more ovoid in males. Differing pubic growth is also expressed in the subpubic arch below the symphysis and between the inferior pubic rami. It is more angular in males, being 50–60°; in females it is rounded, less easy to measure and usually 80–85°. A greater separation of the pubic tubercles in females contributes to the pubic width. The ischiopubic rami are also much more lightly built and narrowed near the symphysis; in males they bear a distinctly rough, everted area for attachment of the penile crura, the corresponding attachment for the clitoris being poorly developed. Ischial spines are closer in males and are more inturned. The greater sciatic notch is usually wider in females: mean values for males and females are 50.4° and 74.4°, respectively. The greater female values for angle and width are associated with increased backward sacral tilt and greater anteroposterior pelvic diameter, especially at lower levels.
The sacrum also displays metrical sexual differences. Female sacra are less curved, the curvature being most marked between the first and second segments and the third and fifth, with an intervening flatter region. Male sacra are more evenly curved, relatively long and narrow and more often exceeding five segments (by addition of a lumbar or coccygeal vertebra). The sacral index compares sacral breadth (between the most anterior points on the auricular surfaces) with length (between midpoints on the anterior margins of the promontory and apex): average values for males and females are 105% and 115%. Auricular surfaces are relatively smaller and more oblique in females, but extend onto the upper three sacral vertebrae in both sexes. The dorsal auricular border is more concave in females. Many differences may be summarized in the generalization that the pelvic cavity is longer and more conical in males, shorter and more cylindrical in females; the axis is curved in both. Differences are greater at the inferior aperture than at the brim, where in absolute measurements males are not as different from females as sometimes stated. The superior aperture is more likely to be anthropoid or android in males and gynaecoid or android in females.
In forensic practice, identification of human skeletal remains (which are sometimes fragmentary) usually involves determination of sex, and this is most reliably established from an examination of the pelvis. Even fragments of the pelvis may be useful in this respect. Several studies of metrical characteristics in various pelvic regions have been made, leading to the establishment of various indices. The ilium has received particular attention, e.g. one index compares the pelvic and sacroiliac parts of the bone. A line is extended back from the iliopubic ramus to the nearest point on the anterior auricular margin and thence to the iliac crest. The auricular point divides this chilotic line into anterior (pelvic) and posterior (sacral) segments, each expressed as a percentage of the other. Chilotic indices display reciprocal values in the sexes: the pelvic part of the chilotic line is predominant in females, and the sacral part in males. Detailed metrical studies of the ilium have indicated its limited reliability in ‘sexing’ pelves. However, the higher incidence and definition of the female preauricular sulcus is recognized. The desirability of correlating all available metrical data is to be emphasized; when a range of pelvic data can be combined, especially if they are metrical, 95% accuracy should be achieved. Complete accuracy has been claimed when the rest of the skeleton is available. Assessment of sex from isolated and often incomplete human remains is less reliable. For further details, consult Mays (1998) and Brothwell & Pollard (2001).
FEMUR
Topography
The femur is the longest and strongest bone in the human body (Figs 80.10, 80.11). Its length is associated with a striding gait, its strength with the weight and muscular forces it is required to withstand. Its shaft, almost cylindrical along most of its length, is bowed forward. It has a proximal rounded, articular head projecting medially from its short neck, which, in turn, is a medial extension of the proximal shaft. The distal extremity is wider and more substantial, and presents a double condyle that articulates with the tibia. In standing, the femoral shafts show an inclination upwards and outwards from their tibial articulations, with the femoral heads being separated by the pelvic width. Since the tibia and fibula descend vertically from the knees, the ankles are also in the line of body weight in standing or walking. The degree of femoral obliquity varies between individuals, but is generally greater in women, reflecting the relatively greater pelvic breadth and shorter femora. Proximally the femur consists of a head, neck, and greater and lesser trochanters.
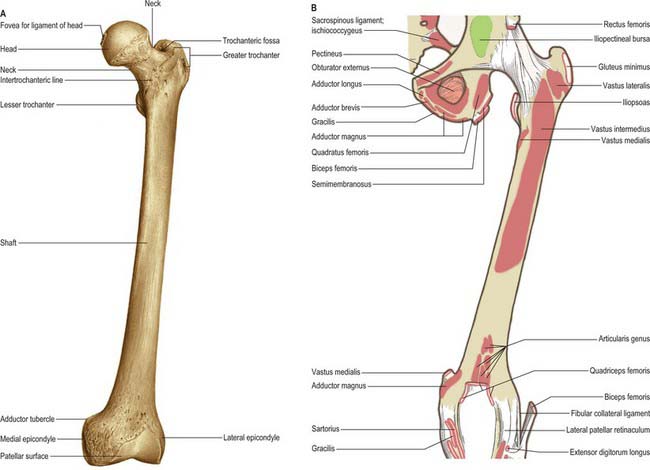
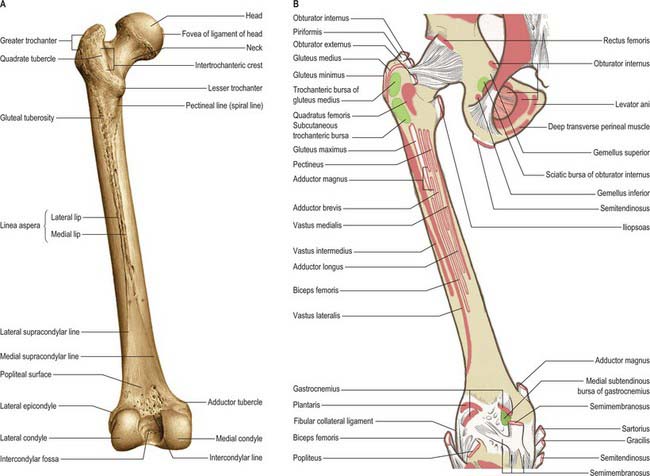
Fig. 80.11 Femur: posterior aspect. A, Osseous features. B, Muscle attachments.
(From Sobotta 2006.)
Femoral head
The femoral head faces anterosuperomedially to articulate with the acetabulum (Fig. 80.12). The head, often described as rather more than half a ‘sphere’, is not part of a true sphere but is spheroidal. Its smoothness is interrupted posteroinferior to its centre by a small, rough fovea. The head is intracapsular and is encircled, distal to its equator, by the acetabular labrum. Its articular margin is distinct, except anteriorly, where the articular surface extends on to the neck. The ligamentum teres is attached to the fovea. The anterior surface of the head is separated inferomedially from the femoral artery by the tendon of psoas major, the psoas bursa, and the articular capsule.
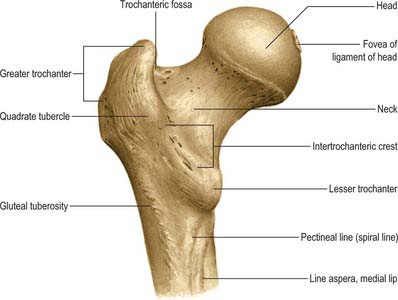
Femoral neck
The femoral neck (Fig. 80.12) is approximately 5 cm long, narrowest in its mid part and widest laterally, and connects the head to the shaft at an average angle of 135° (angle of inclination; neck–shaft angle): this facilitates movement at the hip joint, enabling the limb to swing clear of the pelvis. The neck also provides a lever for the action of the muscles acting about the hip joint, which are attached to the proximal femur. The neck–shaft angle is widest at birth and diminishes gradually until adolescence; it is smaller in females. The neck is laterally rotated with respect to the shaft (angle of anteversion) some 10–15°, although values of this angle vary between individuals and between populations (Eckhoff et al 1994). The contours of the neck are rounded: the upper surface is almost horizontal and slightly concave, the lower is straighter but oblique, directed inferolaterally and backwards to the shaft near the lesser trochanter. On all aspects the neck expands as it approaches the articular surface of the head. The anterior surface of the neck is flat and marked at the junction with the shaft by a rough intertrochanteric line. The posterior surface, facing posteriorly and superiorly, is transversely convex, and concave in its long axis; its junction with the shaft is marked by a rounded intertrochanteric crest. There are numerous vascular foramina, especially anteriorly and posterosuperiorly.
The anterior surface is intracapsular, the capsule attaching laterally to the intertrochanteric line. Facets, often covered by extensions of articular cartilage, and various imprints frequently occur here. These facets may sometimes be associated with squatting. One such feature, the cervical fossa, may be a racial characteristic. On the posterior surface the capsule does not reach the intertrochanteric crest; little more than the medial half of the neck is intracapsular. The anterior surface adjoining the head and covered by cartilage is related to the iliofemoral ligament. A groove, produced by the tendon of obturator externus as it approaches the trochanteric fossa, spirals across the posterior surface of the neck in a proximolateral direction.
Greater trochanter
The greater trochanter is large and quadrangular, projecting up from the junction of the neck and shaft (Fig. 80.12). Its posterosuperior region projects superomedially to overhang the adjacent posterior surface of the neck and here its medial surface presents the rough trochanteric fossa. The proximal border of the trochanter lies approximately a hand’s breadth below the iliac tubercle, level with the centre of the femoral head. It has an anterior rough impression. Its lateral surface, continuous distally with the lateral surface of the femoral shaft, is crossed anteroinferiorly by an oblique, flat strip, which is wider above. This surface is palpable, especially when the muscles are relaxed. The trochanteric fossa occasionally presents a tubercle or exostosis.
Lesser trochanter
The lesser trochanter is a conical posteromedial projection of the shaft at the posteroinferior aspect of its junction with the neck. Its summit and anterior surface are rough, but its posterior surface, at the distal end of the intertrochanteric crest, is smooth. It is not palpable.
Intertrochanteric line
The intertrochanteric line, a prominent ridge at the junction of the anterior surfaces of the neck and shaft, descends medially from a tubercle on the upper part of the anterior aspect of the greater trochanter to a point on the lower border of the neck, anterior to the lesser trochanter, where there may also be a tubercle. This line is the lateral limit of the hip joint capsule anteriorly. The upper and lower bands of the iliofemoral ligament are attached to its proximal and distal ends and the associated tubercles. Distally it is continuous with the spiral line.
Intertrochanteric crest
The intertrochanteric crest, a smooth and prominent ridge at the junction of the posterior surface of the neck with the shaft, descends medially from the posterosuperior angle of the greater trochanter to the lesser trochanter. A little above its centre is a low, rounded quadrate tubercle. It is covered by gluteus maximus, from which it is separated, medial to the tubercle, by quadratus femoris and the upper border of adductor magnus.
Gluteal tuberosity
The gluteal tuberosity may be an elongated depression or a ridge. It may at times be prominent enough to merit the unofficial title of ‘third trochanter’.
Shaft
The shaft is surrounded by muscles and is impalpable (Figs 80.10, 80.11). The distal anterior surface, for 5–6 cm above the patellar articular surface, is covered by a suprapatellar bursa, between bone and muscle. The distal lateral surface is covered by vastus intermedius. The medial surface, devoid of attachments, is covered by vastus medialis.
The shaft is narrowest centrally, expanding a little at its proximal end, and substantially more at its distal end. Its long axis makes an angle of approximately 10° with the vertical, and diverges 5–7° from the long axis of the tibia. Its middle third has three surfaces and borders. The extensive anterior surface, smooth and gently convex, is between the lateral and medial borders, which are both round and indistinct. The posterolateral surface is bounded posteriorly by the broad, rough linea aspera, usually a crest with lateral and medial edges. Its subjacent compact bone is augmented to withstand compressive forces, which are concentrated here by the anterior curvature of the shaft. The linea aspera gives attachment to adductor longus, intermuscular septa and the short head of biceps femoris, all inseparably blended at their attachments. Perforating arteries cross the linea laterally under tendinous arches in adductor magnus and biceps femoris. Nutrient foramina, directed proximally, appear in the linea aspera, varying in number and site, one usually near its proximal end, a second usually near its distal end. The posteromedial surface, smooth like the others, is bounded in front by the indistinct medial border and behind by the linea aspera. In its proximal third the shaft has a fourth, posterior surface, bounded medially by a narrow, rough spiral line that is continuous proximally with the intertrochanteric line and distally with the medial edge of linea aspera. Laterally this surface is limited by the broad, rough, gluteal tuberosity, ascending a little laterally to the greater trochanter and descending to the lateral edge of the linea aspera. In its distal third the posterior surface of the shaft presents a further surface, the popliteal surface (see below) between the medial and lateral supracondylar lines. These lines are continuous above with the corresponding edges of the linea aspera. The lateral line is most distinct in its proximal two-thirds, where the short head of biceps femoris and lateral intermuscular septum are attached. Its distal third has a small rough area for the attachment of plantaris, often encroaching on the popliteal surface. The medial line is indistinct in its proximal two-thirds, where vastus medialis is attached. Distally, the medial line is crossed obliquely by the femoral vessels entering the popliteal fossa from the adductor canal. Further distally, the line is often sharp for 3 or 4 cm proximal to the adductor tubercle.
The popliteal surface, triangular in outline, lies between the medial and lateral supracondylar lines. In its distal medial part it is rough and slightly elevated. Forming the proximal part of the floor of the popliteal fossa, the popliteal surface is covered by a variable amount of fat that separates the popliteal artery from bone. The superior medial genicular artery, a branch of the popliteal artery, arches medially above the medial condyle. It is separated from bone by the medial head of gastrocnemius. The latter is attached a little above the condyle; further distally there may be a smooth facet underlying a bursa for the medial head of gastrocnemius. More medially, there is often an imprint proximal to the articular surface: in flexion this is close to a rough tubercle on the medial tibial condyle for the attachment of semimembranosus. The superior lateral genicular artery arches up laterally proximal to the lateral condyle but is separated from bone by the attachment of plantaris to the distal part of the lateral supracondylar line.
Distal end
The distal end of the femur is widely expanded as a bearing surface for transmission of weight to the tibia (Fig. 80.13). It bears two massive condyles, which are partly articular. Anteriorly the condyles are confluent and continue into the shaft; posteriorly they are separated by a deep intercondylar fossa and project beyond the plane of the popliteal surface. The articular surface is a broad area, like an inverted U, for the patella and the tibia. The patellar surface extends anteriorly on both condyles, especially the lateral. It is transversely concave, vertically convex and grooved for the posterior patellar surface. The tibial surface is divided by the intercondylar fossa but is anteriorly continuous with the patellar surface. Its medial part is a broad strip on the convex inferoposterior surface of the medial condyle, and is gently curved with a medial convexity. Its lateral part covers similar aspects of the lateral condyle but is broader and passes straight back. The tibial surfaces are convex in all directions. The medial and lateral tibial surfaces have dissimilar anteroposterior curvatures. However views differ as to the exact representation of these differences. One view holds that in both tibial portions of the femoral condyles the sagittal radius of curvature is ever decreasing (a ‘closing helix’). More recently it has been suggested that the medial articular surface describes arcs of two circles. The more posterior has a smaller radius. Laterally there may only be one arc of fixed curvature with a radius similar to that of the posterior arc of the medial femoral articular surface. These differences are believed to be important determinants of knee joint motion.
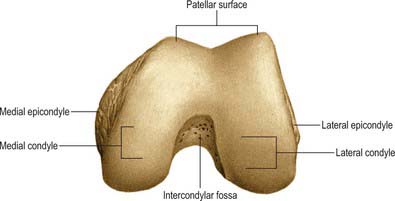
Patellar surface (trochlear groove)
The patellar surface extends more proximally on the lateral side. Its proximal border is therefore oblique and runs distally and medially, separated from the tibial surfaces by two faint grooves that cross the condyles obliquely. The lateral groove is the more distinct. It runs laterally and slightly forwards from the front of the intercondylar fossa and expands to form a faint triangular depression, resting on the anterior edge of the lateral meniscus when the knee is fully extended. The medial groove is restricted to the medial part of the medial condyle and rests on the anterior edge of the medial meniscus in full extension. Where it ceases, the patellar surface continues back to the lateral part of the medial condyle as a semilunar area adjoining the anterior region of the intercondylar fossa. This area articulates with the medial vertical facet of the patella in full flexion; its outline is indistinct in most femora. In habitual squatters articular cartilage may extend to the lateral aspect of the lateral condyle under vastus lateralis.
The trochlear groove helps to stabilize the patella. An abnormally shallow groove predisposes to instability.
The intercondylar fossa separates the two condyles distally and behind. In front it is limited by the distal border of the patellar surface, and behind by an intercondylar line, separating it from the popliteal surface. It is intracapsular but largely extrasynovial. Its lateral wall, the medial surface of the lateral condyle, bears a flat posterosuperior impression which spreads to the floor of the fossa near the intercondylar line for the proximal attachment of the anterior cruciate ligament. The medial wall of the fossa, i.e. the lateral surface of the medial condyle, bears a similar larger area, but far more anteriorly, for the proximal attachment of the posterior cruciate ligament. Both impressions are smooth and largely devoid of vascular foramina, whereas the rest of the fossa is rough and pitted by vascular foramina. A bursal recess between the ligaments may ascend to the fossa. The capsular ligament and, laterally, the oblique popliteal ligament, are attached to the intercondylar line. The ligamentum mucosum (infrapatellar synovial fold or plica) is attached to the anterior border of the fossa.
The lateral condyle (Figs 80.13, 80.14) is larger anteroposteriorly than the medial. Its most prominent point is the lateral epicondyle to which the lateral collateral ligament is attached. A short groove, deeper in front, separates the lateral epicondyle inferiorly from the articular margin. This groove allows the tendon of popliteus to run deep to the lateral collateral ligament and insert inferior and anterior to the ligament insertion. Adjoining the joint margin is a strip of condyle, 1 cm broad. It is intracapsular and covered by synovial membrane except for the attachment of popliteus.
Fig. 80.14 Distal end of femur: lateral aspect. 1. Attachment of plantaris. 2. Attachment of lateral head of gastrocnemius. 3. Lateral epicondyle. 4. Attachment of lateral collateral ligament. 5. Groove for popliteus in full flexion. 6. Attachment of popliteus.
The medial surface is the lateral wall of the intercondylar fossa. Its lateral surface projects beyond the shaft. Part of the lateral head of gastrocnemius is attached to an impression posterosuperior to the lateral epicondyle.
The medial condyle has a bulging, convex medial aspect which is easily palpable. Proximally its adductor tubercle, which may only be a facet rather than a projection, receives the tendon of adductor magnus. The medial prominence of the condyle, the medial epicondyle, is anteroinferior to the tubercle. The lateral surface of the condyle is the medial wall of the intercondylar fossa. The condyle projects distally so that, despite the obliquity of the shaft, the profile of the distal end is almost horizontal. A curved strip, 1 cm wide, adjoining the medial articular margin, is covered by synovial membrane and is inside the joint capsule. Proximal to this, the medial epicondyle receives the medial collateral ligament.
Structure
The femoral shaft is a cylinder of compact bone with a large medullary cavity. The wall is thick in its middle third, where the femur is narrowest and the medullary cavity most capacious. Proximally and distally the compact wall becomes progressively thinner, and the cavity gradually fills with trabecular bone. The extremities, especially where articular, consist of trabecular bone within a thin shell of compact bone, their trabeculae being disposed along lines of greatest stress. At the proximal end the main trabeculae form a series of plates orthogonal to the articular surface, converging to a central dense wedge, which is supported by strong trabeculae passing to the sides of the neck, especially along its upper and lower profiles (Fig. 80.15). Force applied to the femoral head is therefore transmitted to the wedge and thence to the junction of the neck and shaft. This junction is strengthened by dense trabeculae extending laterally from the lesser trochanter to the end of the superior aspect of the neck, thus resisting tensile or shearing forces applied to the neck through the head (Fig. 80.16). Tensile and compressive tests indicate that axial trabeculae of the femoral head withstand much greater stresses than peripheral trabeculae. A smaller bar across the junction of the greater trochanter with the neck and shaft resists shearing produced by muscles attached to it. These two bars are proximal layers of arches between the sides of the shaft and transmit to it forces applied to the proximal end. A thin vertical plate, the calcar femorale, ascends from the compact wall near the linea aspera into the trabeculae of the neck. Medially it joins the posterior wall of the neck; laterally it continues into the greater trochanter, where it disperses into general trabecular bone. It is thus in a plane anterior to the trochanteric crest and base of the lesser trochanter. Newell (1997), in a review of the calcar femorale, has described its three-dimensional anatomy in terms of the work of Dixon (1910), who was the first to recognize the deficiencies of the classic two-dimensional description of the architecture of the proximal femur. Dixon suggested that the trabecular framework of the proximal femur was spiral, and that the ‘arches’ were simplified sectional profiles of this spiral. At the distal end of the femur, trabeculae spring from the entire internal surface of compact bone, descending perpendicular to the articular surface. Proximal to the condyles these are strongest and most accurately perpendicular. Horizontal planes of trabecular bone, arranged like crossed girders, form a series of cubical compartments.
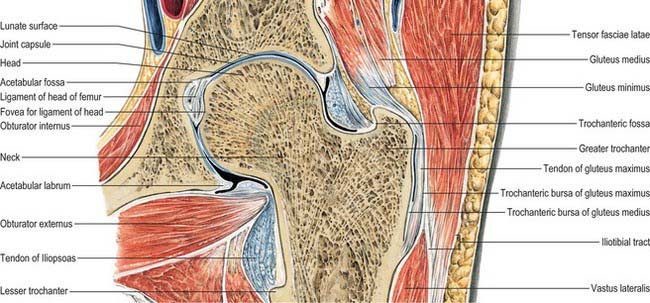
Muscle attachments
The greater trochanter provides attachment for gluteus minimus and medius. Gluteus minimus is attached to its rough anterior impression and gluteus medius to its lateral oblique strip. The bone is separated from the tendon of gluteus medius by a bursa. The area behind is covered by deep fibres of gluteus maximus, with part of its trochanteric bursa interposed. The tendon of piriformis is attached to the upper border of the trochanter and the common (tricipital) tendon of obturator internus and the gemelli is attached to its medial surface. The trochanteric fossa receives the tendon of obturator externus. Psoas major is attached to the summit and anteromedial surface of the lesser trochanter. Iliacus is attached to the medial or anterior surface of its base, descending a little behind the spiral line as its tendon fuses with that of psoas. Adductor magnus (upper part) passes over its posterior surface, sometimes separated by an interposed bursa.
The most proximal fibres of vastus lateralis are attached to the proximal end of the intertrochanteric line, and those of vastus medialis to the distal end. Quadratus femoris is attached to the quadrate tubercle and the immediately distal bone. Vastus intermedius is attached to the anterior and lateral surfaces of the proximal three-quarters of the femoral shaft. Slips of articularis genu are attached distal to this.
The gluteal tuberosity receives the deeper fibres of the distal half of gluteus maximus and, at its medial edge, the uppermost fibres of adductor magnus. Distal to this, adductor magnus is attached to the linea aspera and, by an aponeurosis, to the proximal part of the medial supracondylar ridge. Its remaining fibres form a large tendon attached to the adductor tubercle, with an aponeurotic expansion to the distal part of the medial supracondylar ridge.
Pectineus and adductor brevis are attached to the posterior femoral surface between the gluteal tuberosity and spiral line. The pectineal attachment is a line, sometimes slightly rough, from the base of the lesser trochanter to the linea aspera. Adductor brevis is attached lateral to pectineus and beyond this to the proximal part of the linea aspera, medial to adductor magnus. Adductor longus, intermuscular septa and the short head of biceps femoris are attached to the linea aspera. Vastus lateralis has a linear attachment from the anterior surface of the base of the greater trochanter to the proximal end of the gluteal tuberosity, and along the lateral margin of the latter to the proximal half of the lateral edge of the linea aspera. Vastus medialis is attached from the distal end of the intertrochanteric line along the spiral line to the medial edge of the linea aspera and thence to the medial supracondylar line, which also receives many fibres from the aponeurotic attachments of adductor magnus.
The medial head of gastrocnemius is attached to the posterior surface a little above the medial condyle. The short head of biceps femoris is attached to the proximal two-thirds of the lateral supracondylar line. Plantaris attaches to the line distally. Vastus medialis is attached to the proximal two-thirds of the medial supracondylar line.
Part of the lateral head of gastrocnemius is attached posterosuperiorly to the lateral epicondyle. Popliteus is attached anteriorly in the groove on the outer aspect of the lateral epicondyle. Its tendon passes deep to the lateral collateral ligament (Fig. 80.14). The tendon lies in the groove in full knee flexion; in extension it crosses the articular margin and may form an impression on it.
Vascular supply
The blood supply of the femoral head (Fig. 80.17) is derived from an arterial ring around the neck, just outside the attachment of the fibrous capsule, constituted by the medial and lateral circumflex femoral arteries with minor contributions from the superior and inferior gluteal vessels (see trochanteric anastomosis). From this ring, ascending cervical branches pierce the capsule (under its zona orbicularis) to ascend the neck beneath the reflected synovial membrane. These vessels become the retinacular arteries and form a subsynovial intracapsular anastomosis. Here the vessels are at risk with a displaced fracture of the femoral neck. Interruption of blood supply in this way can lead to avascular necrosis of the femoral head. If the fracture is intracapsular, not only is the intraosseous blood supply damaged but the retinacular vessels are also vulnerable. If the fracture is extracapsular, the retinacular vessels will remain intact and avascular necrosis of the femoral head is much less likely. The ascending cervical vessels give off metaphysial branches that enter the neck, while the intracapsular ring gives off lateral and inferior epiphysial branches. A small medial epiphysial supply, of importance in early childhood, reaches the head along the ligamentum teres by the acetabular branches of the obturator and medial circumflex femoral arteries, which anastomose with the other epiphysial vessels. During growth, the epiphysial plate separates the territories of the metaphysial and epiphysial vessels; these vessels anastomose freely after osseous union of the head and neck. Observations on developmental patterns of this supply in late fetal and early postnatal periods have revealed that although medial and lateral circumflex femoral arteries at first contribute equally, two major branches of the medial provide the final supply, both posterior to the neck. The supply from the lateral circumflex artery diminishes and the arterial ring is interrupted. As the femoral neck elongates, the extracapsular circle becomes more distant from the epiphysial part of the head.
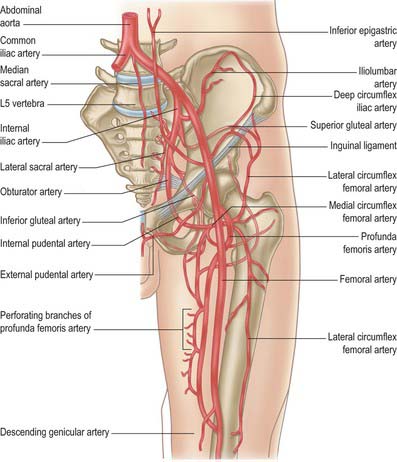
The trochanteric regions and subtrochanteric shaft are supplied by the trochanteric and cruciate arterial anastomoses. More distally in the shaft, nutrient foramina, directed proximally, are found in the linea aspera, varying in number and site: one is usually near its proximal end and a second usually near its distal end. The main nutrient artery is usually derived from the second perforating artery (see profunda femoris). If two nutrient arteries occur, they may branch from the first and third perforators. Periosteal vessels arise from the perforators and from the profunda, and run circumferentially rather than longitudinally. The distal metaphysis has many vascular foramina. Arterial supply here is from the genicular anastomosis. For further details consult Crock (1980, 1996).
The pattern of venous drainage of the head and neck corresponds in general to that of the arteries, though there may be a single large cervical vein posteroinferiorly.
Innervation
The periosteal innervation is derived proximally from nerves that supply the hip joint, distally from those supplying the knee, and in all areas from nerves that innervate muscles attached to the bone.
Ossification
The femur ossifies from five centres: in the shaft, head, greater and lesser trochanters and the distal end (Figs 80.10, 80.11, 80.18). Other than the clavicle, it is the first long bone to ossify. The process starts in the midshaft in the seventh prenatal week and extends to produce a miniature shaft that is largely ossified at birth. Secondary centres appear in the distal end (from which the condyles and epicondyles are formed) during the ninth month, in the head during the first six months after birth, in the greater trochanter during the fourth year and in the lesser between the 12th and 14th year. The centre in the cartilaginous head is restricted to it until the tenth year, so that the epiphysial line (Fig. 80.7) is horizontal and the inferomedial part of the articular surface is on the neck. The medial epiphysial margin later grows over this part of the articular surface. Thus, the mature epiphysis is a hollow cup on the summit of the neck. The epiphysial line follows the articular margin except where it is separated superiorly from the articular surface by a non-articular area where blood vessels enter the head (Trueta 1957). The epiphyses fuse independently: the lesser trochanter soon after puberty followed by the greater trochanter. The capital epiphysis fuses in the 14th year in females and 17th year in males,. The epiphysis at the distal end fuses in the 16th year in females, and 18th year in males. The distal epiphysial plate traverses the adductor tubercle.
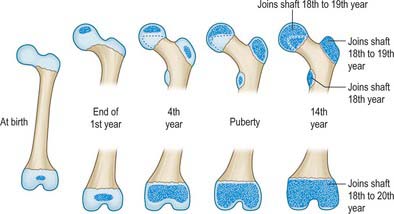
Growth plate considerations
Trauma to any epiphysial plate can lead to bony union between epiphysis and metaphysis, and so cause premature cessation of growth. Any surgery in the hip region in children can injure the growth plate, resulting in abnormal proximal femoral development. In the case of fractures involving the epiphysis, expeditious restoration of normal bony alignment is essential in order to minimize the risk of subsequent abnormal growth.
The growth plate represents a line of weakness and predisposes to fracture from injury. Such acute injuries affecting the capital epiphysis are uncommon. However, a more chronic fracture through the capital epiphysis occurs in ‘slipped upper femoral epiphysis’. The condition affects pubescent adolescents, especially males. Endocrinological abnormality may be related. The femoral head epiphysis displaces posteriorly off the femoral neck. If it heals in this position, lower limb deformity and restricted hip movement occur. A classic hallmark is obligatory external rotation of the femur as the hip is flexed. Treatment varies according to the time taken for the ‘slip’ to occur. Normal anatomical restoration is not attempted because of an increased incidence of avascular necrosis. The position of the femoral head may be accepted as it is and fixed with screws in this position to stop further displacement. This treatment will deliberately cause premature growth plate fusion and so prevent future ‘slippage’. Since the distal femoral growth plate accounts for most of the normal increase in longitudinal growth of the femoral shaft, an acceptable limb length difference usually results.
Infection of bone in neonates and young children tends to arise via bacteria in the blood stream which usually ‘seed’ in the metaphysial region, probably as a consequence of the vascular ‘arcade’ arrangement of arteries in this part of the bone. The proximal femoral growth plate is intra-articular. As a result infection in the proximal femoral metaphysis can spread into the joint and result in a septic arthritis that can destroy the hip joint permanently.
The distal end of the femur is the only epiphysis in which ossification consistently starts just before birth: the phenomenon therefore serves as a reliable indicator of the gestational maturity of a stillborn baby. Since the epiphysial plate is level with the adductor tubercle, the epiphysis is partly extra-articular. Operations here may damage the distal epiphysial cartilage in children and result in subsequent shortening of the leg.
JOINTS
PUBIC SYMPHYSIS
The pubic bones meet in the midline at the pubic symphysis, a secondary cartilaginous joint (Fig. 80.4).
The articulating surfaces are the medial (symphysial) surfaces of the pubic bones, each covered by a thin layer of tightly adherent hyaline cartilage (surface growth cartilage in the young). The junction is not flat but marked by reciprocal crests and papillae. Theoretically this would resist shearing. The surfaces of hyaline cartilage are connected by fibrocartilage, varying in thickness and constituting the interpubic disc. The symphysis often contains a cavity, probably due to absorption. It rarely appears before the tenth year and is non-synovial. The cavity, which is better developed in females, is usually posterosuperior but may reach the front or even occupy most of the cartilage.
The interpubic disc is strengthened anteriorly by several interlacing collagenous fibrous layers, passing obliquely from bone to bone, decussating with fibres of the external oblique aponeuroses and the medial tendons of the recti abdominis. These layers constitute the anterior pubic ligament. There are less well-developed posterior fibres, sometimes named the posterior pubic ligament. The main ligaments of the joint are the superior and arcuate pubic ligaments. The superior pubic ligament connects the bones above, extending to the pubic tubercles. The arcuate pubic ligament, a thick arch of fibres, connects the lower borders of the symphysial pubic surfaces bounding the pubic arch. It blends superiorly with the interpubic disc and extends laterally attached to the inferior pubic rami. Its inferior edge is separated from the anterior border of the perineal membrane by an opening which is traversed by the deep dorsal vein of the penis or clitoris.
The pubic symphysis is supplied by pubic branches of the obturator, superficial external pudendal and inferior epigastric arteries.
The pubic symphysis is innervated by branches from the iliohypogastric, ilioinguinal and pudendal nerves.
The interpubic disc and the superior and arcuate ligaments are the main stabilizing factors of the pubic symphysis.
Angulation, rotation and displacement are possible but slight and are likely during movement at the sacroiliac and hip joints. Excessive movement may occur as a sports injury. Some separation occurs late in gestation and during childbirth: on occasion this is considerable.
Anteriorly the pubic symphysis is related to superficial fascia and skin. Because of the obliquity of the joint, the proximal ends of the penile or clitoral shafts lie anterior to the lower half of the joint. Inferiorly the urethra lies about 2.5 cm away in the male, and somewhat closer in the female, as it passes through the perineal membrane. Closer to the joint, the deep dorsal vein of the penis or clitoris passes between the arcuate ligament and the anterior border of the perineal membrane. Posteriorly the upper part of the joint is separated from the inferolateral surfaces of the bladder by the retropubic fat pad. Inferiorly in the male the prostatic venous plexus separates the prostate from the lower part of the joint. The region is sometimes termed the retropubic space. These relationships explain why traumatic disruption of the anterior bony pelvis may be associated with serious urogenital injury.
SACROILIAC JOINT
The sacroiliac joint is a synovial articulation between the sacral and iliac auricular surfaces. Fibrous adhesions and gradual obliteration occur in both sexes, earlier in males, and after the menopause in females. Radiological evidence of obliteration in normal subjects is occasionally seen before 50 years, but is not uncommon thereafter: in old age the joint may be completely fibrosed and occasionally even ossified.
The surfaces are nearly flat in infants, whereas in adults they are irregular, often markedly so, and sometimes undulant. The curvatures and irregularities, greater in males, are reciprocal: they restrict movements and contribute to the considerable strength of the joint in transmitting weight from the vertebral column to the lower limbs. The sacral surface is covered by hyaline cartilage which is thicker anteriorly than posteriorly in adults. The thinner cartilage on the iliac surface is also hyaline in type, as confirmed by the presence of type II collagen.
The ligaments of the sacroiliac joint are the anterior, interosseous and posterior sacroiliac, iliolumbar, sacrotuberous and sacrospinus ligaments.
The anterior sacroiliac ligament (Fig. 80.8), an anteroinferior capsular thickening, is particularly welldeveloped near the arcuate line and the posterior inferior iliac spine, where it connects the third sacral segment to the lateral side of the preauricular sulcus. It is thin elsewhere.
Interosseous sacroiliac ligament
The interosseous sacroiliac ligament is the major bond between the bones, filling the irregular space posterosuperior to the joint. It is covered superficially by the posterior sacroiliac ligament. Its deeper part has superior and inferior bands passing from depressions posterior to the sacral auricular surface to those on the iliac tuberosity. These bands are covered by, and blend with, a more superficial fibrous sheet connecting the posterosuperior margin of a rough area posterior to the sacral auricular surface to the corresponding margins of the iliac tuberosity. This sheet is often partially divided into superior and inferior parts, the former uniting the superior articular process and lateral crest on the first two sacral segments to the neighbouring ilium as a short posterior iliac ligament (see Fig. 42.52).
The posterior sacroiliac ligament
The posterior sacroiliac ligament (see Fig. 42.52) overlies the interosseous ligament: the dorsal rami of the sacral spinal nerves and vessels intervene. It consists of several weak fasciculi connecting the intermediate and lateral sacral crests to the posterior superior iliac spine and posterior end of the internal lip of the iliac crest. Inferior fibres, from the third and fourth sacral segments, ascend to the posterior superior iliac spine and posterior end of the internal lip of the iliac crest: they may form a separate long posterior sacroiliac ligament. This ligament is continuous laterally with part of the sacrotuberous ligament and medially with the posterior lamina of the thoracolumbar fascia.
The sacrotuberous ligament (Fig. 80.8, see Fig. 42.52) is attached by its broad base to the posterior superior iliac spine., the posterior sacroiliac ligaments (with which it is partly blended), to the lower transverse sacral tubercles and the lateral margins of the lower sacrum and upper coccyx. Its oblique fibres descend laterally, converging to form a thick, narrow band that widens again below and is attached to the medial margin of the ischial tuberosity. It then spreads along the ischial ramus as the falciform process, whose concave edge blends with the fascial sheath of the internal pudendal vessels and pudendal nerve. The lowest fibres of gluteus maximus are attached to the posterior surface of the ligament; superficial fibres of the lower part of the ligament continue into the tendon of biceps femoris. The ligament is pierced by the coccygeal branches of the inferior gluteal artery, the perforating cutaneous nerve and filaments of the coccygeal plexus.
The thin, triangular sacrospinous ligament (Fig. 80.8) extends from the ischial spine to the lateral margins of the sacrum and coccyx anterior to the sacrotuberous ligament, with which it blends in part. Its anterior surface is in fact the coccygeus muscle, i.e. muscle and ligament are coextensive. The sacrospinous ligament is often regarded as a degenerate part of coccygeus.
The sacrotuberous and sacrospinous ligaments convert the sciatic notches into foramina (Fig. 80.8).
The greater sciatic foramen is bounded anterosuperiorly by the greater sciatic notch, posteriorly by the sacrotuberous ligament and inferiorly by the sacrospinous ligament and ischial spine. It is partly filled by the emerging piriformis, above which the superior gluteal vessels and nerve leave the pelvis. Below it, the inferior gluteal vessels and nerve, internal pudendal vessels and pudendal nerve, sciatic and posterior femoral cutaneous nerves and the nerves to obturator internus and quadratus femoris all leave the pelvis.
The lesser sciatic foramen is bounded anteriorly by the ischial body, superiorly by its spine and sacrospinous ligament, and posteriorly by the sacrotuberous ligament. It transmits the tendon of obturator internus, the nerve to obturator internus, and the internal pudendal vessels and pudendal nerve.
The arterial supply of the sacroiliac joint is derived from the iliolumbar, superior gluteal and superior lateral sacral arteries, with corresponding venous drainage. Lymphatic drainage follows the arteries, reaching the iliac and lumbar nodes.
Nerve fibres ramify within the joint capsule and adjoining ligaments, but their source is uncertain. It is thought that the joint probably receives branches from the anterior and posterior rami of the first two sacral spinal nerves, and from the superior gluteal nerve, and there may also be contributions from the obturator nerve and the lumbosacral trunk: it would appear that Hilton’s law is not readily applicable to this joint.
The sacro-iliac joint has been implicated as the source of pain in the lower back and buttocks in approximately 15% of the population. Diagnosing sacroiliac joint-mediated pain is difficult because the presenting complaints are similar to those of other causes of back pain. Patients with sacroiliac joint-mediated pain rarely report pain above L5; most localize their pain to the area around the posterior superior iliac spine.
The sacroiliac joint is one of the most stable joints in the body, and supports the weight of the trunk. The reciprocal irregularity of the joint surfaces allows very little movement. The tendency of the sacrum to be forced downwards by the trunk is resisted by the extremely strong posterior ligaments, while the iliolumbar ligaments help to resist displacement of the fifth lumbar vertebra over the sacrum. The sacrotuberous and sacrospinous ligaments oppose upward tilting of the lower part of the sacrum when downward thrust is applied at its upper end.
Primary movement of the sacroiliac joint is minimal. All muscles that cross the joint act on the lumbar spine or on the hip. Such movements as do occur are secondarily imposed on the joint as the pelvis moves. Data from living subjects are technically difficult to obtain, and those based on plain radiographs are unreliable. Studies using implanted tantalum spheres and biplanar radiography have shown mean rotational ranges of less than 2°. Even when there is recordable movement, the direction of movement is irregular. Biplanar radiography has also shown that the axes of movement of the sacroiliac joint during hip movement are oblique, and that the axes differ in flexion and extension.
During pregnancy the pelvic joints and ligaments loosen under the influence of the hormone relaxin. Movements in the joints increase. Relaxation renders the sacroiliac locking mechanism less effective, permitting greater rotation and perhaps allowing alterations in pelvic diameters at childbirth, although the effect is probably small. The impaired locking mechanism diverts the strain of weightbearing to the ligaments, with frequent sacroiliac strain after pregnancy.
The sacroiliac joints have many important anterior relations. The internal and external iliac veins join to form the common iliac veins immediately anteriorly, separating the joints from the bifurcations of the common iliac arteries and, more anteriorly, the ureters. The lumbosacral trunk and the obturator nerve cross the anterior aspect of the joint behind the vessels. Piriformis partly attaches to the anterior capsule, separating the joint from the upper part of the sacral plexus.
Accessory sacroiliac articulations are not uncommon. They develop behind the articular surface between the lateral sacral crest and posterior superior iliac spine and iliac tuberosity, and are acquired fibrocartilaginous joints resulting from the stresses of weightbearing. They have a joint capsule, are saddle-shaped, and may be single, double, unilateral or bilateral (Weisl 1954).
MUSCLES
MUSCLES OF THE ILIAC REGION
Although there is no ‘iliac region’ as such, this heading conveniently describes a group of three muscles that originate from the lumbar vertebral column (psoas major and minor) and the ilium (iliacus). Two (psoas major and iliacus) are attached together on the femur as flexors of the hip joint and are often considered as a functional unit, iliopsoas. Psoas minor only reaches the pubis, and acts on the spine and sacroiliac joint.
Psoas major
Psoas major is a long muscle that lies on either side of the lumbar vertebral column and the pelvic brim (Fig. 80.19). Its proximal attachments are complex. They include the anterior surfaces and lower borders of the transverse processes of all the lumbar vertebrae. There are five digitations, each from the bodies of two adjoining vertebrae and their intervertebral disc. The highest of these arises from the lower margin of the body of the 12th thoracic vertebra, the upper margin of the body of the first lumbar vertebra and the interposed thoracolumbar disc. The lowest arises from the adjacent margins of the bodies of the fourth and fifth lumbar vertebrae and the interposed disc. A series of tendinous arches extend across the narrow parts of the bodies of the lumbar vertebrae between the digitations already described. The lumbar arteries and veins, and filaments from the sympathetic trunk, pass medial to these arches. The upper four lumbar intervertebral foramina bear important relations to these attachments of the muscle. The foramina lie anterior to the transverse processes and posterior to the attachments to vertebral bodies, discs and tendinous arches. Thus, the roots of the lumbar plexus enter the muscle directly, the plexus is lodged within it, and its branches emerge from its borders and surfaces.
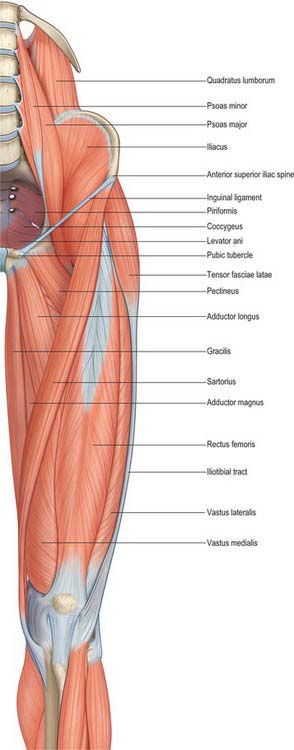
The muscle descends along the pelvic brim, continues posterior to the inguinal ligament and anterior to the capsule of the hip joint, and converges to a tendon which, having received on its lateral side nearly all the fibres of iliacus, becomes attached to the lesser trochanter of the femur. The large subtendinous iliac bursa, which occasionally communicates with the cavity of the hip joint, separates the tendon from the pubis and the capsule of the joint.
The complex vertebral attachments of psoas major sometimes display minor numerical variations.
The upper limit of psoas major is posterior to the diaphragm in the lowest part of the posterior mediastinum. It may be in contact with the posterior extremity of the pleural sac. In the abdomen its anterolateral surface is related to the medial arcuate ligament (a linear arched thickening in the psoas fascia), extraperitoneal tissue and peritoneum, the kidney, psoas minor, renal vessels, ureter, testicular or ovarian vessels and genitofemoral nerve. Anteriorly, the right psoas is overlapped by the inferior vena cava and crossed by the terminal ileum; the left psoas is crossed by the colon. Its posterior surface is related to the transverse processes of the lumbar vertebrae and the medial edge of quadratus lumborum. The lumbar plexus is embedded posteriorly in the substance of psoas major. Medially the muscle is related to the bodies of the lumbar vertebrae and lumbar vessels. Along its anteromedial margin it is in contact with the sympathetic trunk, aortic lymph nodes and, along the pelvic brim, with the external iliac artery. This margin is covered by the inferior vena cava on the right side, and lies posterior and lateral to the abdominal aorta on the left side. In the thigh, psoas is related anteriorly to the fascia lata and the femoral artery, and posteriorly to the capsule of the hip joint from which it is separated by a bursa. At its medial border, it is related to pectineus and the medial circumflex femoral artery, and to the femoral vein, which may overlap it slightly. At its lateral border it is related to the femoral nerve and iliacus. The femoral nerve descends at first through the fibres of psoas major, and then in the furrow between it and iliacus.
Branches of the lumbar plexus diverge from the abdominal part of psoas. Emerging from the lateral border, from above downwards, are the iliohypogastric, ilioinguinal and lateral femoral cutaneous and femoral nerves. Emerging from the anterolateral surface is the genitofemoral nerve. The obturator and accessory obturator nerves and the upper root of the lumbosacral trunk all emerge from the medial border.
Psoas major is supplied by a rich network of arteries derived from the lumbar, iliolumbar, obturator, external iliac and femoral arteries. In general terms, the upper part of the muscle is supplied by the lumbar arteries, the mid part by the anterior branch of the iliolumbar artery (the main artery to the muscle) with contributions from the deep circumflex and external iliac arteries, and the distal part by the femoral artery and its branches. The psoas sheath has an arterial supply independent from that of the muscle, though the same vessels contribute.
Psoas major is innervated by the ventral rami of the lumbar spinal nerves, mainly L1 and 2 with some contribution from L3.
Psoas minor
Psoas minor (Fig. 80.19) is absent in almost 40% of subjects. When present, it lies anterior to psoas major, entirely within the abdomen. It arises from the sides of the bodies of the twelfth thoracic and first lumbar vertebrae and from the disc between them. It ends in a long, flat tendon which is attached to the pecten pubis and ilio-pubic ramus and, laterally, to the iliac fascia.
Psoas minor lies on psoas major, and its proximal anterior relations are those of the anterior or anteromedial surface of that muscle.
Iliacus
Iliacus (Fig. 80.19) is a triangular sheet of muscle that arises from the superior two-thirds of the concavity of the iliac fossa, the inner lip of the iliac crest, the ventral sacroiliac and iliolumbar ligaments, and the upper surface of the lateral part of the sacrum (see Fig. 42.36). In front, it reaches as far as the anterior superior and anterior inferior iliac spines, and receives a few fibres from the upper part of the capsule of the hip joint. Most of its fibres converge into the lateral side of the strong tendon of psoas major, and the muscles then insert together into the lesser trochanter, but some fibres are attached directly to the femur 2.5 cm below and in front of the lesser trochanter.
In the abdomen, the anterior surface of iliacus is related to its fascia (which separates the muscle from extraperitoneal tissue and peritoneum), the lateral femoral cutaneous nerve, the caecum (on the right) and the iliac part of the descending colon (on the left). On its posterior surface is the iliac fossa; at its medial border, psoas major and the femoral nerve. In the thigh, its anterior surface is in contact with the fascia lata, rectus femoris, sartorius and profunda femoris artery, and its posterior surface is in contact with the capsule of the hip joint, from which it is partially separated by a bursa it shares with psoas major.
Iliacus is supplied by the same arterial network as psoas major, and there is mutual overlap of the arterial territories of each muscle. The main supply is from the iliac branches of the iliolumbar artery, with contributions from the deep circumflex iliac and obturator arteries, and branches of the femoral artery.
Psoas major, acting from above together with iliacus, flexes the thigh upon the pelvis. Electromyographic studies do not support the common view that psoas major acts as a medial rotator of the hip joint, but activity has been described in lateral rotation, particularly in the young. When psoas major and iliacus of both sides act from below, they contract powerfully to bend the trunk and pelvis forwards against resistance, as in raising the trunk from the recumbent to the sitting posture when undertaking a ‘sit-up’ exercise.
Electromyography does not support the commonly held belief that unilateral action of psoas causes lateral and forward flexion of the trunk to that side.
In symmetrical upright stance, iliopsoas has some action from below to maintain the vertebral column upright. Psoas major is active in balancing the trunk while sitting.
Both psoas and iliacus may be the sites of pathological collections of fluid. An abscess (typically tuberculous) of vertebral origin may track down through psoas and present as a mass in the thigh. Haematoma or infection within the iliacus fascia may present as a mass or as a flexion deformity of the hip.
Iliopsoas may be tested clinically by actively flexing the hip against resistance, in the supine position with hip and knee flexed.
MUSCLES OF THE GLUTEAL REGION
Tensor fasciae latae
Tensor fasciae latae (Fig. 80.19) arises from the anterior 5 cm of the outer lip of the iliac crest, from the lateral surface of the anterior superior iliac spine and part of the border of the notch below it, between gluteus medius and sartorius, and from the deep surface of the fascia lata. Proximal attachments may extend to the aponeurotic fascia superficial to gluteus medius. It descends between, and is attached to, the two layers of the iliotibial tract of the fascia lata and usually ends approximately one-third of the way down the thigh, although it may occasionally extend as far as the lateral femoral condyle.
The muscle itself is supplied mainly by a large ascending branch of the lateral circumflex femoral artery. The tensor fasciae latae musculocutaneous flap is raised on this pedicle. The superior part of the muscle receives branches from the superior gluteal artery. The fascia surrounding the muscle is supplied on its superficial aspect by the superficial circumflex iliac artery and on its deep surface by the lateral circumflex femoral artery.
Tensor fasciae latae, acting through the iliotibial tract, extends the knee with lateral rotation of the leg; it may also assist in abduction and medial rotation of the thigh, though its role as an abductor is debatable. The muscle helps to maintain upright posture while minimizing energy expenditure on muscle activity: when the subject is standing it acts from below to steady the pelvis on the head of the femur and, through the iliotibial tract, to steady the condyles of the femur on the tibial condyles while the knee extensors are relaxed. The muscle aids gluteus medius in postural abduction at the hip. Postural control is its main function.
In the last 20° or so of extension the pull of the iliotibial tract is anterior to the flexion axis of the knee and so the tensor fasciae latae is a weak extensor. Flexion greater than 20° leads to the iliotibial tract passing posterior to the axis of flexion so that the muscle becomes a weak flexor.
Gluteus maximus
Gluteus maximus (Fig. 80.20) is the largest and most superficial muscle in the gluteal region. It is a broad, thick quadrilateral mass, which, with its overlying adipose fascia, forms the familiar prominence of the buttock. Gluteus maximus is thicker and more extensive in man than in any non-human primate, features that presumably correlate with the evolutionary transition to bipedality and a permanently upright posture. The muscle has a coarse fascicular architecture, with large bundles of fibres separated by fibrous septa. It arises from the posterior gluteal line of the ilium, and the rough area of bone, including the crest, immediately above and behind it; from the aponeurosis of erector spinae; the dorsal surface of the lower part of the sacrum and the side of the coccyx; the sacrotuberous ligament; and the fascia (gluteal aponeurosis) which covers gluteus medius. There may be additional slips from the lumbar aponeurosis or ischial tuberosity. The muscle may also be bilaminar. The fibres descend laterally; the upper part of the muscle, together with the superficial fibres of the lower part, ends in a thick tendinous lamina which passes lateral to the greater trochanter and is attached to the iliotibial tract of the fascia lata. The deeper fibres of the lower part of the muscle are attached to the gluteal tuberosity between vastus lateralis and adductor magnus.
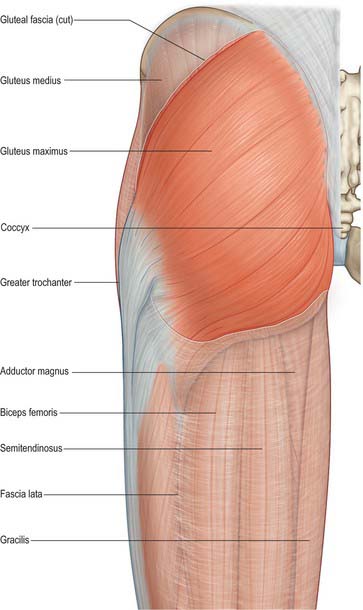
A thin fascia separates the superficial surface of gluteus maximus from the overlying thick adipose subcutaneous tissue. The deep surface of the muscle is related to the ilium, sacrum, coccyx, sacrotuberous ligament, part of gluteus medius, piriformis, the gemelli, obturator internus, quadratus femoris, the ischial tuberosity, greater trochanter, and the attachments of biceps femoris, semitendinosus, semimembranosus and adductor magnus to the ischial tuberosity. Three bursae lie deep to gluteus maximus: trochanteric, over the greater trochanter; gluteofemoral, between the tendon of gluteus maximus and that of vastus lateralis; and ischiofemoral, over the gluteal tuberosity, which is less commonly present.
The superficial division of the superior gluteal artery reaches the deep surface of the muscle between piriformis and gluteus medius. The inferior gluteal and internal pudendal vessels, the sciatic, pudendal and posterior femoral cutaneous nerves, muscular branches from the sacral femoral cutaneous nerves, and muscular branches from the sacral plexus all leave the pelvis below piriformis. The first perforating artery and the terminal branches of the medial circumflex femoral artery are also deep to the lower part of gluteus maximus. Its upper border is thin and overlies gluteus medius. Its prominent lower border is free and slopes downwards and laterally. It is crossed by the horizontal gluteal fold (the posterior flexure line of the hip joint), which marks the upper limit of the back of the thigh on the surface.
The dominant vascular pedicle is usually that of the inferior gluteal artery, which supplies approximately two-thirds of the muscle. The remainder is supplied mainly by the superior gluteal artery, though this may sometimes be the dominant vessel. The lateral and distal borders of the muscle receive a supply from the first profunda perforator and from the medial circumflex femoral artery. Minor branches may be derived from the lateral circumflex femoral, lateral sacral and internal pudendal arteries.
Gluteus maximus musculocutaneous flaps may be based on either of the gluteal vessels or on the first profunda perforator, depending on the site and size of the defect requiring coverage.
Acting from the pelvis, gluteus maximus can extend the flexed thigh and bring it into line with the trunk. Acting from its distal attachment, it may prevent the forward momentum of the trunk from producing flexion at the supporting hip during bipedal gait. The muscle is inactive during standing, when swaying forwards at the ankle joints, or when bending forwards at the hip joints to touch the toes. However, it acts with the hamstrings in raising the trunk after stooping, by rotating the pelvis backwards on the head of the femur. It is intermittently active in the walking cycle and in climbing stairs, and continuously active in strong lateral rotation of the thigh. Its upper fibres are active in powerful abduction of the thigh. It is a tensor of the fascia lata, and through the iliotibial tract it stabilizes the femur on the tibia when the knee extensor muscles are relaxed.
Gluteus medius
Gluteus medius is a broad, thick muscle (Fig. 80.20, Fig. 80.21A). It arises from the outer surface of the ilium between the iliac crest and posterior gluteal line above, and the anterior gluteal line below, and also from the strong fascia superficial to its upper part. The fibres converge to a flat tendon that attaches to a ridge that slants downwards and forwards on the lateral surface of the greater trochanter.
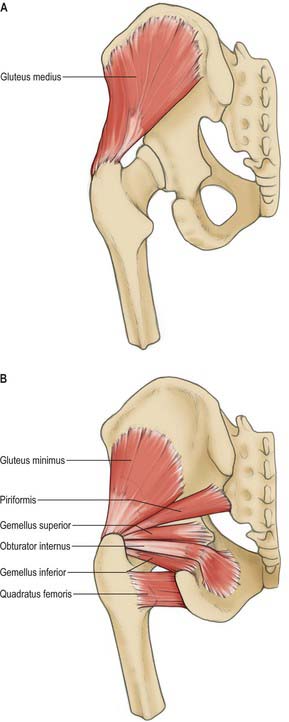
Fig. 80.21 A, Gluteus medius. B, Gluteus minimus and short external rotators of the hip.
(From Sobotta 2006.)
A deep slip of the muscle may be attached to the upper border of the trochanter. The posterior edge of gluteus medius sometimes blends with piriformis.
The posterior third of gluteus medius is covered by gluteus maximus, but it is superficial in its anterior two-thirds, where it is covered by a strong layer of deep fascia. Its deep surface is related to gluteus minimus. Branches of the deep divisions of the superior gluteal nerve and artery run between the medius and minimus muscles and are vulnerable during anterolateral and lateral approaches to the hip that involve splitting gluteus medius (see Fig. 80.22). Where the tendon glides on the anterosuperior part of the lateral surface of the trochanter, a bursa (trochanteric bursa of gluteus medius) separates it from the bone.
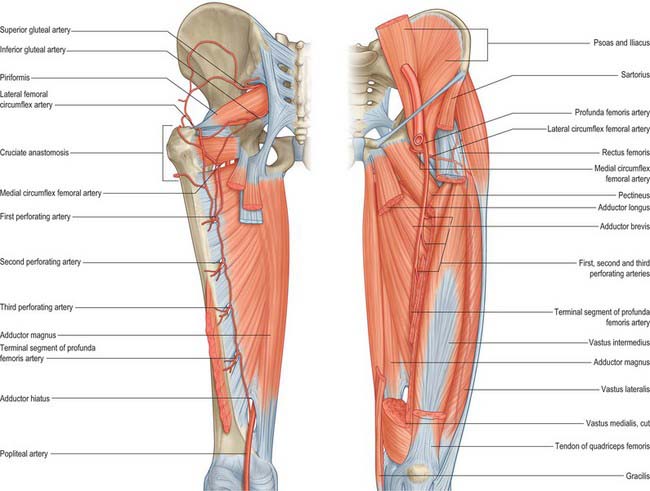
Gluteus minimus
Gluteus minimus lies deep to gluteus medius (Fig. 80.21B). The fan-shaped muscle arises from the outer surface of the ilium between the anterior and inferior gluteal lines and, behind, from the margin of the greater sciatic notch. The fibres converge below to the deep surface of an aponeurosis that ends in a tendon which is attached to an anterolateral ridge on the greater trochanter and contributes an expansion to the capsule of the hip joint.
The muscle may divide into anterior and posterior parts. Separate slips may pass to piriformis, gemellus superior, or vastus lateralis.
Branches of the deep division of the superior gluteal artery and nerve run on the superficial surface of the muscle. The reflected tendon of rectus femoris and the capsule of the hip joint are deep to gluteus minimus. A bursa (trochanteric bursa of gluteus minimus) separates the tendon from the medial part of the anterior surface of the greater trochanter.
Gluteus minimus is supplied from both its surfaces, from the main trunk and the deep branch of the superior gluteal artery, with a contribution at its femoral attachment from the trochanteric anastomosis (Fig. 80.22).
Actions of gluteus medius and minimus
Both gluteus medius and minimus, acting from the pelvis, abduct the thigh, and their anterior fibres rotate it medially. Acting from the femur, they play an essential part in maintaining the trunk upright when the foot of the opposite side is raised from the ground in walking and running. In this phase the body weight tends to make the pelvis sag downwards on the unsupported side. This is counteracted by the gluteus medius and minimus of the supporting side, which, acting from below, exert such powerful traction on the hip bone that the pelvis is actually raised a little on the unsupported side. In symmetrical standing with the feet somewhat separated, the abductor muscles are usually ‘silent’ on electromyography, but with the feet placed parallel and close together they are active.
Testing
The supportive effect of the glutei (medius and minimus) on the pelvis when the contralateral foot is raised depends on the following conditions. The two muscles, and their innervation, must be functioning normally. The components of the hip joint, which forms the fulcrum, must be in their usual relation. The neck of the femur must be intact, with its normal angulation to the shaft.
When any one of these conditions is not fulfilled, e.g. in paralysis of the glutei, congenital dislocation of the hip, or coxa vara, the supporting mechanism is upset and the pelvis sinks on the unsupported side when the patient tries to stand on the affected limb. This is known clinically as Trendelenburg’s sign. Individuals with paralysis of gluteus medius and minimus have a characteristic lurching gait. Provided that these two muscles are intact, paralysis of other muscles acting on the hip joint produces remarkably little deficit in walking, or even in running.
Gluteus medius and minimus may be tested together by medial (internal) rotation of the thigh against resistance, in the supine position with hip and knee flexed. Both muscles may be tested together with tensor fasciae latae by abduction of the lower limb against resistance, in the supine position with the knee extended.
Piriformis
Piriformis (see Fig. 63.1, Fig. 80.21B) occupies a central position in the buttock, where it lies in the same plane as gluteus medius. It arises from the anterior surface of the sacrum by three digitations, which are attached to the portions of bone between the pelvic sacral foramina, and to the grooves leading from the foramina (see Fig. 42.36). It also arises from the gluteal surface of the ilium near the posterior inferior iliac spine, from the capsule of the adjacent sacroiliac joint, and sometimes from the upper part of the pelvic surface of the sacrotuberous ligament. The muscle passes out of the pelvis through the greater sciatic foramen, which it substantially fills. Here it constitutes an important surgical landmark in the identification of structures that emerge above and below it. It inserts into the medial side of the upper border of the greater trochanter of the femur via a rounded tendon that lies behind and above, but is often partially blended with, the common tendon of obturator internus and the gemelli. The muscle itself may be fused with gluteus medius.
Within the pelvis, the anterior surface of piriformis is related to the rectum (especially on the left), the sacral plexus of nerves and branches of the internal iliac vessels. The posterior surface lies against the sacrum. Outside the pelvis, its anterior surface is in contact with the posterior surface of the ischium and capsule of the hip joint and its posterior surface with gluteus maximus. Its upper border is in contact with gluteus medius and the superior gluteal vessels and nerve, its lower border with coccygeus and gemellus superior. The inferior gluteal and internal pudendal vessels, the sciatic, posterior femoral cutaneous and pudendal nerves, and muscular branches from the sacral plexus appear in the buttock in the interval between piriformis and gemellus superior. The relationship between piriformis and the sciatic nerve is variable. The undivided nerve may emerge above the muscle or through the muscle. The major divisions of the nerve may lie either side of the muscle, or (the most common variant) one division passes between the heads of a divided muscle and one division either above or below.
Piriformis can occasionally cause entrapment of the sciatic nerve in the buttock, giving rise to the so-called ‘piriformis syndrome’.
In the buttock, piriformis is supplied mainly from the superior gluteal artery, with contributions from the gemellar branches of the internal pudendal. There may be a separate branch from the inferior gluteal artery. In the pelvis the main supply is from the lateral sacral artery, with contributions from both gluteal vessels.
Obturator internus
Obturator internus (see Fig. 63.1, Fig. 80.21B) is situated partly within the true pelvis and partly posterior to the hip joint. It arises from the internal surface of the anterolateral wall of the lesser pelvic cavity. Its attachments, which almost surround the obturator foramen, are to the inferior ramus of the pubis, the ischial ramus, and the pelvic surface of the hip bone below and behind the pelvic brim, to the upper part of the greater sciatic foramen above and behind, to the obturator foramen below and in front (Fig. 80.5B). It also arises from the medial part of the pelvic surface of the obturator membrane, from the tendinous arch that completes the obturator canal, and, to a small extent, from the obturator fascia that covers the muscle. The fibres converge towards the lesser sciatic foramen and end in four or five tendinous bands on the deep surface of the muscle. These bands make a lateral right-angled turn around the grooved surface of the ischium between its spine and tuberosity. The grooved surface is covered with a smooth layer of hyaline cartilage and is separated from the tendon by a bursa: ridges on the surface correspond to furrows between the tendinous bands. These bands leave the pelvis through the lesser sciatic foramen and unite to form a single flattened tendon that passes horizontally across the capsule of the hip joint. The gemelli fuse with this tendon before it inserts into an anterior impression on the medial surface of the greater trochanter anterosuperior to the trochanteric fossa. A long, narrow bursa is usually interposed between the tendon and the capsule of the hip joint, and occasionally communicates with the bursa between the tendon and the ischium.
Within the pelvis, the anterolateral surface of the muscle is in contact with the obturator membrane and inner surface of the lateral wall of the pelvis. Its posteromedial surface is related to the obturator fascia, the origin of levator ani, and the sheath that surrounds the internal pudendal vessels and pudendal nerve, and forms the lateral wall of the ischiorectal fossa. Outside the pelvis, the muscle is covered by gluteus maximus, is crossed posteriorly by the sciatic nerve and passes behind the hip joint. As the tendon of obturator internus emerges from the lesser sciatic foramen it is overlapped both above and below by the two gemelli, which form a muscular canal for it. Near its termination the gemelli pass anterior to the tendon and form a groove in which it lies.
The main arterial supply of the extrapelvic part of obturator internus is from the gemellar branches of the internal pudendal artery. The intrapelvic part is supplied by the obturator artery, which gives branches to both surfaces of the muscle: it is also supplied by small branches from the internal pudendal artery.
Obturator externus
Obturator externus (Fig. 80.23) is a flat, triangular muscle covering the external surface of the anterior pelvic wall. It arises from the anteromedial two-thirds of the external surface of the obturator membrane, and from the adjacent bone of the pubic and ischial rami, extending for a short distance onto their pelvic surfaces between the margin of the obturator foramen and the obturator membrane. The whole muscle, and the tendon into which its fibres converge, spirals backwards, laterally and upwards, and thus crosses the inferior aspect and then the back of the neck of the femur and lower part of the capsule of the hip joint to end in the trochanteric fossa of the femur.
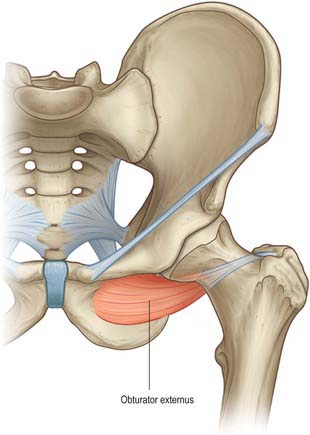
A bursa, which communicates with the hip joint, may be interposed between the tendon and the hip joint capsule and femoral neck. The obturator vessels lie between the muscle and the obturator membrane. The anterior branch of the obturator nerve reaches the thigh by passing in front of the muscle, and the posterior branch by piercing it.
Obturator externus receives a variable pattern of supply from the obturator and medial circumflex femoral arteries.
It has been suggested that the short muscles around the hip joint, pectineus, piriformis, obturators, gemelli and quadratus femoris, are more important as postural muscles than as prime movers, acting as adjustable ligaments to maintain the stability and integrity of the hip. However, these muscles are largely inaccessible to direct observation, and because of the hazards presented by their close relationship to important neurovascular structures there is a total lack of electromyographic data in man. In both bipedal walking and vertical climbing obturator externus is recruited during the early part of swing phase: in climbing it effects lateral rotation of the thigh, and in walking it probably counteracts the tendency to medial rotation produced by the anterior adductor muscles at this stage of the cycle. Obturator internus differs from externus in its pattern of use but its role in bipedal walking remains unclear. Its attachments suggest that it, like the gemelli, is a lateral rotator of the extended thigh and an abductor of the flexed thigh. These actions may be used to antagonize unwanted components of movement produced by the primary locomotor muscles.
Gemellus inferior and gemellus superior
Gemellus inferior arises from the upper part of the lateral surface of the ischial tuberosity, immediately below the groove for the tendon of obturator internus. It blends with the lower border of this tendon, and inserts with it into the medial surface of the greater trochanter.
Gemellus superior, the smaller of the two gemelli, arises from the dorsal surface of the ischial spine, blends with the upper border of the tendon of obturator internus, and inserts with it into the medial surface of the greater trochanter. It is sometimes absent.
The two gemelli can be regarded as accessory to obturator internus (see Fig. 80.21B).
The superior gemellus is supplied by the internal pudendal artery and its gemellar branches; by the inferior gluteal and sometimes also by the superior gluteal artery. The inferior gemellus is supplied mainly by the medial circumflex femoral artery.
Quadratus femoris
Quadratus femoris (see Fig. 80.21B) is a flat, quadrilateral muscle lying between gemellus inferior and the upper margin of adductor magnus, from which it is separated by the transverse branch of the medial circumflex femoral artery. It arises from the upper part of the external aspect of the ischial tuberosity and inserts into a small tubercle a little above the middle of the trochanteric crest of the femur and into the bone for a short distance below. It may be absent.
The muscle passes behind the hip joint and the neck of the femur, separated from them by the tendon of obturator externus and the ascending branch of the medial circumflex femoral artery (which may cause troublesome bleeding if injured during the posterior approach to the hip). The sciatic nerve crosses the muscle posteriorly. A bursa is often present between the muscle and the lesser trochanter.
MUSCLES OF THE THIGH
The presence and position of the femoral neck cause the femoral shaft to lie obliquely, which means that the anterior (extensor) muscle group, the quadriceps, runs obliquely distally and medially and so applies a pull to the patella which is both laterally and proximally directed. The adductor muscles occupy the region between the quadriceps and the medial margin of the thigh. They are attached distally to the posterior surface of the femur and lie more posteriorly than the quadriceps. The posterior (flexor) muscle group, the hamstrings, lie behind the adductors. The attachments of these muscles determine the nature and direction of displacement of femoral shaft fractures.
Anterior compartment
The muscles of the anterior compartment (Figs 80.2, 80.19) include sartorius and rectus femoris, which can act both at the hip and knee joints, and vasti medialis, lateralis, and intermedius, which act only at the knee. Articularis genu, a derivative of vastus intermedius, completes the group: it retracts the synovial capsule of the knee joint. Rectus femoris and the vasti extend the knee joint through a common tendon and hence are collectively termed the quadriceps femoris muscle. Adductor longus and pectineus are sometimes considered to be part of both the anterior and the adductor compartments.
Sartorius
Sartorius is a narrow strap muscle, and is the longest muscle in the body. It arises by tendinous fibres from the anterior superior iliac spine and the upper half of the notch below it. It crosses the thigh obliquely over to the medial side, then descends more vertically to the medial side of the knee. The muscle fibres terminate at this point and a thin, flattened tendon curves obliquely forwards and expands into a broad aponeurosis. The aponeurosis is attached to the proximal part of the medial surface of the tibia in front of gracilis and semitendinosus, together forming the ‘pes anserinus’ (Fig. 80.24). A slip from its upper margin blends with the capsule of the knee joint, and another from its lower margin merges with the superficial layer of the deep fascia on the medial side of the leg. This sheet of tissue passes inferiorly to lie superficial to and over the distal insertions of gracilis and semitendinosus. It has to be split by sharp dissection to reveal these two tendons if they need to be harvested, e.g. in cruciate ligament surgery (Fig. 80.24).
In some cases sartorius is absent, while in others the muscle may possess an extra head: when the latter occurs, the extra head is attached to the pectineal line or to the femoral sheath. Variations in the distal attachment of sartorius, in the vicinity of the knee, have been described.
The main arterial supply to sartorius is derived from multiple branches of the femoral system, and enters the medial half of the muscle from its deep surface. Vessels supplying the proximal third of sartorius may arise from the common femoral, the main trunk of the profunda, the artery of the quadriceps, the superficial femoral or the lateral circumflex femoral artery. There may be an additional proximal supply from the superficial circumflex iliac artery. Those supplying the middle third of the muscle arise from the superficial femoral artery. The distal vessels arise from the superficial femoral within the adductor canal and from the descending genicular artery.
Sartorius assists in flexing the leg at the knee, and the thigh on the pelvis, particularly when these two movements are combined. It also helps to abduct the thigh and to rotate it laterally. (Together with inversion of the foot, these movements bring the sole of the foot into direct view.) The fact that sartorius represents only 1% of the physiological cross-sectional area of all muscles which cross the hip or knee joint would appear to suggest that its role in walking is a minor one. In a subject ascending steps, electromyographic activity of sartorius increases during lateral rotation of the thigh at the end of the swing phase immediately preceding heel strike, which presumably decelerates the limb. Sartorius may therefore have a significant involvement in climbing.
Femoral triangle
The femoral triangle is a depressed, intermuscular space in the anteromedial aspect of the proximal thigh, lying immediately distal to the inguinal ligament. The latter constitutes the base of the femoral triangle’s inverted triangular outline. Its lateral boundary is the medial margin of sartorius. Its medial boundary is the medial margin of adductor longus. Its distal extremity, the apex, is where sartorius overlaps adductor longus. Its floor is provided laterally by iliacus and psoas major, medially by pectineus and adductor longus. Its roof is the overlying fascia lata. The femoral vessels, passing from midbase to apex, are in the deepest part of the triangle. Lying lateral to the artery and outside the femoral sheath is the femoral nerve, which, on entering the femoral triangle divides into multiple branches. The triangle also contains fat and lymph nodes.
Quadriceps femoris
Quadriceps femoris (Figs 80.2, 80.19), the great extensor muscle of the leg, covers almost all of the front and sides of the femur. It can be divided into four parts, each named individually. One, rectus femoris, arises from the ilium and travels straight down the middle of the thigh, its shape and path determining its name. The other three arise from the shaft of the femur and surround it (apart from the linea aspera) from the trochanters to the condyles: vastus lateralis is lateral to the femur, vastus medialis is medial to it, and vastus intermedius lies in front of the femur. Rectus femoris crosses both hip and knee joints, while the three vasti only cross the knee joint.
The tendons of the four components of quadriceps unite in the lower part of the thigh to form a single strong tendon attached to the base of the patella, and some fibres continue over it to blend with the patellar tendon. The patella is a sesamoid bone in the quadriceps tendon, and the patellar tendon, which extends from the patellar apex to the tubercle of the tibia, is the continuation of the main tendon. The medial and lateral patellar retinacula are expansions from its borders. The suprapatellar bursa (a synovial extension of the knee joint) lies between the femur and the suprapatellar part of the quadriceps tendon. The deep infrapatellar bursa lies between the patellar tendon and the proximal end of the tibia.
The arterial supply to the quadriceps group has been traditionally ascribed to a single branch of either the profunda femoris or of the lateral circumflex femoral, the ‘artery of the quadriceps’. This vessel, which may be large, occasionally arises directly from the femoral artery. However, according to Taylor & Razaboni (1994) this artery does not supply all four components of the quadriceps: vastus medialis is supplied directly from the (superficial) femoral artery. The supply of the individual muscle components is described below: their sheaths may have an additional and variable supply.
Quadriceps as a group shows little anatomical variation.
Rectus femoris
Rectus femoris is fusiform. Its superficial fibres are bipennate, the deep fibres parallel. It has a double origin on the ilium: a straight tendon arises from the anterior inferior iliac spine, and a thinner, flatter reflected tendon from a groove above the acetabulum and from the fibrous capsule of the hip joint. The two unite at an acute angle and spread into an aponeurosis that is prolonged downwards on the anterior surface of the muscle: the muscular fibres arise from this aponeurosis. The fibres end in a broad, thick aponeurosis that forms over the lower two-thirds of its posterior surface and gradually narrows into the thick, flat tendon by which it is attached to the base of the patella. This constitutes the superficial central part of the quadriceps tendon.
Rectus femoris may arise from the anterior superior iliac spine, and its reflected head may be absent.
Proximally the muscle is covered by tensor fasciae latae, iliacus and sartorius. The reflected head lies beneath gluteus minimus. The capsule of the hip joint, vastus intermedius, the anterior borders of vasti lateralis and medius, the lateral circumflex femoral artery and some branches of the femoral nerve all lie deep to rectus femoris.
There are two main pedicles, a superior and an inferior. The superior enters rectus femoris at the junction of its upper and middle thirds and arises from the ‘artery of the quadriceps’. The inferior and larger branch arises from the ‘artery of the quadriceps’ at about midthigh level and enters the muscle a few centimetres more distally. There are additional contributions from the lateral circumflex femoral and, less often, the superficial circumflex iliac arteries.
Vastus medialis and vastus medialis obliquus
Vastus medialis arises from the lower part of the intertrochanteric line, spiral line, medial lip of the linea aspera, proximal part of the medial supracondylar line, the tendons of adductor longus and magnus, and the medial intermuscular septum. Its fibres pass downwards and forwards at an average angle of 15° to the long axis of the femur, most of them into an aponeurosis on the deep surface of the muscle which is attached to the medial border of the patella and quadriceps tendon. An expansion from this aponeurosis reinforces the capsule of the knee joint and is attached below to the medial condyle of the tibia.
The lowest fibres are nearly horizontal and form a bulge in the living subject, medial to the upper half of the patella. Some authors distinguish this part of the muscle as the vastus medialis obliquus, with fibres that originate largely from the tendon of adductor magnus and insert into the medial border of the patella. It plays an important role in the function of patellofemoral joint.
Vastus medialis is partly covered by rectus femoris and sartorius. In the central part of the thigh it forms the lateral wall of the adductor (Hunter’s) canal.
Vastus medialis is supplied by three branches of the (superficial) femoral artery. The superior and middle branches arise, sometimes from a common trunk, proximal to the adductor canal, while the inferior arises within the canal. There may also be minor branches from the profunda femoris and descending genicular arteries.
Vastus lateralis
Vastus lateralis is the largest component of quadriceps femoris. It arises by a broad aponeurosis from the upper part of the intertrochanteric line, the anterior and inferior borders of the greater trochanter, the lateral lip of the gluteal tuberosity, and the proximal half of the lateral lip of the linea aspera. This aponeurosis covers the proximal three-quarters of the muscle; many additional fibres arise from its deep surface. A few fibres also arise from the tendon of gluteus maximus and the lateral intermuscular septum between vastus lateralis and the short head of biceps femoris. The muscular mass thus formed is attached to a strong aponeurosis on the deep surface of the lower part of the muscle. This narrows to a flat tendon, which is attached to the base and lateral border of the patella and blends into the compound quadriceps femoris tendon. It contributes an expansion to the capsule of the knee joint which descends to the lateral condyle of the tibia and blends with the iliotibial tract.
Vastus lateralis is covered laterally by the fascia lata and the aponeurotic insertions of tensor fasciae latae and gluteus maximus. It is separated from vastus intermedius by branches of the femoral nerve and the lateral circumflex femoral artery. Posteriorly it is separated from biceps femoris by the lateral intermuscular septum.
Vastus intermedius
Vastus intermedius arises from the anterior and lateral surfaces of the upper two-thirds of the femoral shaft, and from the lower part of the lateral intermuscular septum. Its fibres end on the anterior surface of the muscle in an aponeurosis which forms the deep part of the quadriceps tendon and is attached to the lateral border of the patella and the lateral condyle of the tibia.
Vastus intermedius appears to be inseparable from vastus medialis. However, when rectus femoris is reflected a narrow cleft can be seen extending upwards from the medial border of the patella between the two muscles, sometimes as far as the lower part of the intertrochanteric line, proximal to which, the two muscles are frequently fused.
Vastus intermedius is almost completely covered by the other quadriceps components, except laterally where a small part becomes superficial behind vastus lateralis.
Vastus intermedius receives a lateral artery of supply, which arises from the ‘artery of the quadriceps’, and a medial artery that arises directly from the profunda. Arteries to the other quadriceps components may also contribute. There is an anastomotic network deep to the muscle: this supplies the suprapatellar pouch and articularis genu and may form a collateral pathway when the (superficial) femoral artery is blocked.
Articularis genu
Articularis genu is a small muscle, usually distinct from vastus intermedius but occasionally blending with it. It consists of several muscular bundles that arise from the anterior surface of the lower part of the femoral shaft and are attached to a proximal reflection of the synovial membrane of the knee joint. It is visible during knee arthroscopy when viewed looking superiorly from within the suprapatellar pouch.
Innervation of quadriceps group
Quadriceps femoris and articularis genu are supplied by the femoral nerve, L2, 3 and 4.
Actions of quadriceps group
Quadriceps femoris extends the knee. Rectus femoris helps to flex the thigh on the pelvis; if the thigh is fixed, it helps to flex the pelvis on the thigh. Rectus can flex the hip and extend the knee simultaneously. Electromyographic studies indicate that the three vasti are not equally active in different phases of extension or rotation. There is little or no activity in quadriceps during standing. Rectus femoris pulls the patella along the line of the mechanical axis of the lower limb (i.e. the line connecting the centres of the hip, knee, and ankle) because its attachment is anterior to the hip joint. The remaining quadriceps are attached to the shaft of the femur and so the pull is lateral as well as proximal. An extremely important dynamic function of vastus medialis obliquus is to counter this lateral vector on the patella during knee motion. Other static factors that help are the depth of the femoral trochlear groove, the buttressing effect of the lateral femoral condyle in the groove, and the restraining action of the medial patellofemoral ligament. Inadequacy of vastus medialis obliquus is a factor in producing patellar instability and pain. Strengthening of this muscle to improve patellar ‘tracking’ on the femur and stability is an important part of physiotherapy regimens for patellofemoral problems.
Articularis genu retracts the synovial suprapatellar bursa proximally during extension of the leg, presumably to prevent interposition of redundant synovial folds between patella and femur.
Adductor compartment
The muscles of the adductor compartment, gracilis, pectineus, adductor longus, adductor brevis, and adductor magnus (Fig. 80.25), have evolved, as their nerve supply suggests, both from flexor and extensor columns. All five muscles cross the hip joint, but only gracilis reaches beyond the knee. They are known collectively as the adductors of the thigh, although their actions are more complex than this, e.g. acting from below they have important roles in balancing the trunk on the lower limb during walking.
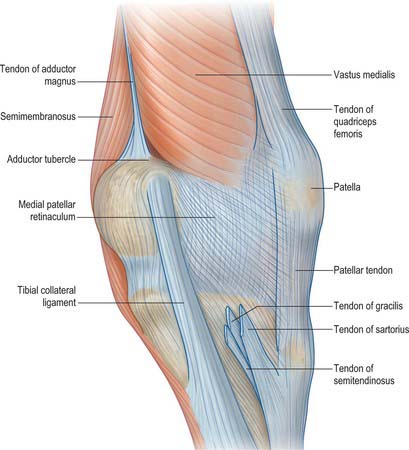
Fig. 80.25 Anteromedial aspect of the left knee region showing distal attachments of sartorius, gracilis, and semitendinosus.
Their blood supply is derived from the profunda, medial circumflex femoral, (superficial) femoral and obturator arteries. The profunda (or sometimes the first perforator) supplies a large branch, the ‘artery of the adductor muscles’.
Gracilis
Gracilis (Figs 80.2, 80.19) is the most superficial of the adductor group. It is thin and flat, broad above, narrow and tapering below. It arises by a thin aponeurosis from the medial margins of the lower half of the body of the pubis, the whole of the inferior pubic ramus, and the adjoining part of the ischial ramus. The fibres descend vertically into a rounded tendon, often harvested as a knee ligament graft, which passes across the medial condyle of the femur posterior to the tendon of sartorius. It then curves around the medial condyle of the tibia, where it fans out and is attached to the upper part of the medial surface of the tibia, just below the condyle, forming part of the pes anserinus. A few fibres from the lower part of the tendon continue into the deep fascia of the lower leg. Often there is a slip that blends with the tendon of the medial head of gastrocnemius. Unless divided this can lead to problems during surgical harvesting of the gracilis tendon.
The muscle is covered medially by the fascia lata throughout most of its length. Deep to gracilis lie adductor brevis and adductor magnus. The tibial attachment is immediately proximal to that of semitendinosus, and its upper edge is overlapped by the tendon of sartorius, with which it is partly blended. It is separated from the medial collateral ligament of the knee joint by the tibial intertendinous bursa.
The arterial supply to gracilis enters via its lateral surface. The main pedicle arises from the ‘artery to the adductors’ of the profunda and enters at the junction of the upper and middle thirds of the muscle. The gracilis musculocutaneous flap is based on this pedicle. A less important artery enters the distal third of the muscle from the femoral artery. There is a minor proximal supply from the medial circumflex femoral.
Adductor longus
Adductor longus (Figs 80.2, 80.26), the most anterior of the three adductors, is a large, fan-shaped muscle that lies in the same plane as pectineus. It arises by a narrow tendon with a flattened (sometimes C-shaped) cross-section, which is attached to the front of the pubis in the angle between the crest and the symphysis. It expands into a broad fleshy belly which descends posterolaterally and inserts by an aponeurosis into the linea aspera in the middle third of the femur, between vastus medialis and adductors magnus and brevis, usually blending with all of them. Its proximal attachment is vulnerable to overload from sporting activity: this is one cause of sport-related groin pain.
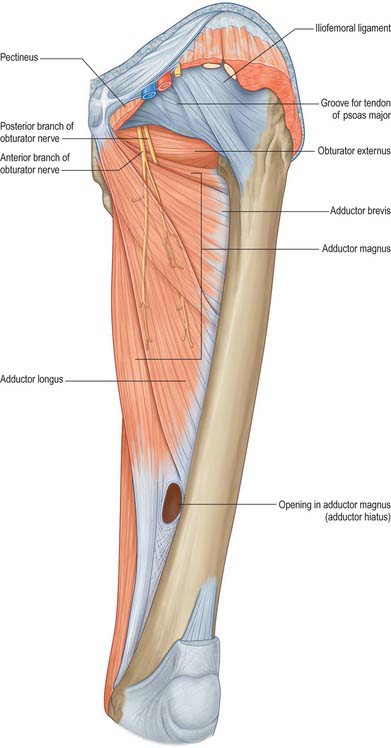
Anterior to adductor longus are the spermatic cord, fascia lata (which separates it from the long saphenous vein) and, near its attachment, the femoral artery and vein and sartorius. Posterior to it are adductor brevis and adductor magnus, the anterior branch of the obturator nerve and, near its attachment, the profunda femoris vessels. Lateral and medial to adductor longus are, respectively, pectineus, and gracilis.
The main supply to adductor longus is to the central part of the muscle from the ‘artery to the adductors’ of the profunda. There is an additional proximal supply from the medial circumflex femoral artery, and a more distal supply from the femoral artery and sometimes the descending genicular artery.
Adductor brevis
Adductor brevis (Figs 80.2A, 80.26) lies posterior to pectineus and adductor longus. It arises by a narrow attachment from the external aspect of the body and inferior ramus of the pubis, between gracilis and obturator externus. Like adductor longus it is somewhat triangular, and expands as it descends posterolaterally to insert via an aponeurosis into the femur, along a line from the lesser trochanter to the linea aspera, and on the upper part of the linea immediately behind pectineus and the upper part of adductor longus.
Adductor brevis often has two or three separate parts, or may be integrated into adductor magnus.
Anteriorly lie pectineus, adductor longus, the profunda femoris artery, and the anterior branch of the obturator nerve; posteriorly are adductor magnus and the posterior branch of the obturator nerve. The upper border of adductor brevis is related to the medial circumflex femoral artery, obturator externus, and the conjoined tendon of psoas major and iliacus. Its lower border is related to gracilis and adductor magnus. The second, or first and second perforating arteries pierce it near its femoral attachment.
The vascular supply to adductor brevis is variable. Usually the main supply is directly from the profunda femoris distally and from the ‘artery to the adductors’ more proximally. There is an additional proximal supply from the medial circumflex femoral artery. The deep surface receives branches from the obturator artery.
Adductor magnus
Adductor magnus (see Figs 80.2, 80.26), a massive triangular muscle, arises from a small part of the inferior ramus of the pubis, from the conjoined ischial ramus, and from the inferolateral aspect of the ischial tuberosity. The short, horizontal fibres from the pubic ramus are inserted into the medial margin of the gluteal tuberosity of the femur, medial to gluteus maximus; this part of the muscle, in a plane anterior to the rest, is sometimes called adductor minimus. The fibres from the ischial ramus fan out downwards and laterally, to insert via a broad aponeurosis into the linea aspera and the proximal part of the medial supracondylar line. The medial part of the muscle, composed mainly of fibres from the ischial tuberosity, is a thick mass which descends almost vertically, and ends in the lower third of the thigh in a rounded tendon, which can be palpated proximal to its attachment to the adductor tubercle on the medial condyle of the femur. The tendon is connected by a fibrous expansion to the medial supracondylar line.
The long, linear attachment of the muscle is interrupted by a series of osseo-aponeurotic openings, bridged by tendinous arches attached to the bone. The upper four are small and transmit the perforating branches and the termination of the profunda femoris artery. The lowest is large and allows the femoral vessels to cross to the popliteal fossa.
The vertical, ischiocondylar part of the muscle varies in its degree of separation from the rest. The upper border of adductor magnus may fuse with quadratus femoris.
Anteriorly lie pectineus, adductor brevis and adductor longus, the femoral and profunda vessels, and the posterior branch of the obturator nerve. A bursa separates the proximal part of the muscle from the lesser trochanter of the femur. Posteriorly are the sciatic nerve, gluteus maximus, biceps femoris, semitendinosus and semimembranosus. The superior border is parallel with quadratus femoris, and the transverse branch of the medial circumflex femoral artery passes between the muscles. The medial border is related to gracilis, sartorius and the fascia lata.
As expected from its position, adductor magnus is supplied from both its anterior and posterior aspects. The contribution from the anterior compartment is the more important. Here the obturator, profunda femoris and (superficial) femoral arteries contribute; the main supply is directly from the distal part of the profunda. Distally there may be contributions from the femoral and descending genicular arteries. Posteriorly there are branches from the medial circumflex femoral, the first and second perforating and the popliteal vessels.
Adductor magnus is composite and is doubly innervated by the obturator nerve and the tibial division of the sciatic nerve (L2, 3 and 4); the latter supplies the ischiocondylar part. Both nerves are derived from anterior divisions in the lumbosacral plexus, indicating a primitive flexor origin for both parts of the muscle.
Actions of the adductors
Extensive or forcible adduction of the femur is not often required. Although the adductors can act in this way, they more commonly act as synergists in the complex patterns of gait activity, and to some degree as controllers of posture. They are active during flexion and extension of the knee. Magnus and longus are probably medial rotators of the thigh. The adductors are inactive during adduction of the abducted thigh in the erect posture (when gravity assists), but active in other postures, such as the supine position, or during adduction of the flexed thigh when standing. They are also active during flexion (longus) and extension (magnus) of the thigh at the hip joint. In symmetrical easy standing their activity is minimal.
Testing of the adductors
The adductors are usually tested as a group by adduction of the thigh against resistance, in the supine position with the knee extended. The tendon of adductor magnus can be felt just proximal to the adductor tubercle on the medial condyle of the femur. Clinical testing of the other actions mentioned above is not feasible for the individual muscles.
Pectineus
Pectineus (Fig. 80.19) is a flat, quadrangular muscle in the femoral triangle. It may also be considered as part of the anterior compartment of the thigh. It arises from the pecten pubis, from the bone in front of it between the iliopubic ramus and the pubic tubercle, and from the fascia on its own anterior surface. The fibres descend, initially posteromedially and then posterolaterally to be attached along a line from the lesser trochanter to the linea aspera.
Pectineus may be bilaminar, in which case the two layers receive separate nerve supplies. Proximally it may be partially or wholly attached to the capsule of the hip joint.
Pectineus is related anteriorly to the deep lamina of the fascia lata, which separates it from the femoral vessels and long saphenous vein; posteriorly to the capsule of the hip joint, adductor brevis, obturator externus and the anterior branch of the obturator nerve; laterally to psoas major and the medial circumflex femoral vessels; and medially to the lateral margin of adductor longus.
The main arterial supply to pectineus is derived from the medial circumflex femoral artery, which enters the superficial surface of the muscle. There may be a branch from the (common) femoral more proximally, and a deep supply from the obturator artery.
Pectineus is innervated by the femoral nerve, L2 and 3, and accessory obturator nerve, L3, when present. Occasionally it receives a branch from the obturator nerve. The muscle may be incompletely divided into dorsal and ventral layers, which are supplied respectively by the obturator and femoral (or rarely accessory obturator) nerves.
Adductor canal
The adductor canal (Hunter’s canal; subsartorial canal) is a trough-shaped intermuscular tunnel occupying the distal two-thirds of the medial aspect of the thigh (Fig. 80.2). It starts at the apex of the femoral triangle and extends distally as far as the distal attachment of the tendon of adductor magnus. It is triangular in section, and is bounded anterolaterally by vastus medialis, posteromedially by adductor longus, and distal to adductor longus, by adductor magnus. Its anteromedial boundary (often referred to as the roof) is a strong and dense fascia that extends from the medial surface of vastus medialis to the medial edge of the adductors longus and magnus, overlapping in its stride, the femoral vessels in the adductor canal. This fascia, on account of being overlain by sartorius, is termed the subsartorial fascia.
The adductor canal contains the femoral artery and vein, the descending genicular and muscular branches of the femoral artery and their corresponding venous tributaries, the saphenous nerve, and the nerve to vastus medialis (until it enters its muscle). The femoral vessels pass from the adductor canal into the popliteal fossa via the adductor hiatus, an opening in the tendon of adductor magnus adjacent to the femoral shaft, two-thirds of the way down the adductor canal.
Posterior compartment
The posterior femoral muscles, biceps femoris, semitendinosus, and semimembranosus, are colloquially termed the ‘hamstrings’. They cross both hip and knee joints, and integrate extension at the hip with flexion at the knee. As the muscles span the back of the knee, they form the proximal lateral and medial margins of the popliteal fossa. The actions of these muscles and their clinical anatomy will be considered as a group after they have been described individually.
The muscles of the posterior compartment (see Figs 80.2, 80.22) receive their blood supply from the perforating branches of the profunda femoris, most importantly through the first perforator. This vessel has important anastomoses with the inferior gluteal artery (on or within semitendinosus) and with the medial circumflex femoral artery, while the third perforator anastomoses with the medial superior genicular artery within the short head of biceps. This anastomotic chain forms an important collateral arterial pathway when the femoral artery is blocked.
Semitendinosus
Semitendinosus (see Fig. 80.2B), notable for the length of its tendon, lies posteromedial in the thigh. It arises from an inferomedial impression on the upper area of the ischial tuberosity (Fig. 80.5), by a tendon it shares with the long head of biceps femoris, and from an aponeurosis connecting the adjacent surfaces of the two muscles for 7.5 cm from their common origin. The belly is fusiform and ends a little below midthigh in a long, rounded tendon that runs on the posterior surface of semimembranosus. The tendon curves around the medial condyle of the tibia, passes over the medial collateral ligament of the knee joint (from which it is separated by the bursa of the pes anserinus), and inserts into the upper part of the medial surface of the tibia behind the attachment of sartorius and distal to that of gracilis (Fig. 80.24). At its termination it is united with the tendon of gracilis and gives off a prolongation to the deep fascia of the leg and to the medial head of gastrocnemius. A tendinous intersection is usually present near the midpoint of the muscle, which may also receive a muscular slip from the long head of biceps femoris. These connections with the medial head of gastrocnemius and biceps can cause difficulty when harvesting the tendon surgically for a graft.
Semitendinosus lies on semimembranosus throughout its length. The relations of the distal part of the muscle are described above and with the pes anserinus.
The two main arteries of supply to semitendinosus are superior and inferior. The superior is derived from either the medial circumflex femoral artery or from the first perforating artery. The inferior and larger branch arises from the first perforator distal to the origin of the superior branch. There is an accessory supply at the attachments of the muscle: at the ischial attachment this is derived from the inferior gluteal artery, and at the tibial attachment from the medial inferior genicular artery.
Semimembranosus
Semimembranosus (see Fig. 80.2B), so named because of the flattened form of its upper attachment, is posteromedial in the thigh. It arises by a long, flat tendon from a superolateral impression on the ischial tuberosity (Fig. 80.5). Inferomedially the tendinous fibres intermingle to some extent with those of biceps femoris and semitendinosus. The tendon receives, from the ischial tuberosity and ramus, two fibrous expansions that flank adductor magnus. It then broadens and descends deep to semitendinosus and the long head of biceps femoris. Muscle fibres arise from the tendon at about midthigh and converge to a second aponeurosis on the posterior aspect of the lower part of the muscle, which tapers to the heavy, rounded tendon of the distal attachment. The tendon divides at the level of the knee into five components. The main one is attached to a tubercle (sometimes called the tuberculum tendinis) on the posterior aspect of the medial tibial condyle. The others are: a series of slips to the medial margin of the tibia, immediately behind the medial collateral ligament; a thin fibrous expansion to the fascia over popliteus; a cord-like tendon to the inferior lip and adjacent part of the groove on the back of the medial tibial condyle, deep to the medial collateral ligament; and a strong expansion which passes obliquely upwards to the femoral intercondylar line and lateral femoral condyle and forms much of the oblique popliteal ligament of the knee joint.
Semimembranosus varies considerably in size, and may be absent. It may be double, and arise mainly from the sacrotuberous ligament. Slips to the femur or to adductor magnus may occur.
Semimembranosus overlaps the popliteal vessels and is itself partly overlapped by semitendinosus throughout its extent (see Fig. 80.2). Its deep surface lies on adductor magnus. The sciatic nerve lies laterally and surprisingly close to the surface. The distal end of the muscle partially overlies the medial head of gastrocnemius before passing anteromedially to it. An important bursa lies between the semimembranosus tendon and gastrocnemius, and often communicates with the knee joint and with a smaller bursa between the tendon and the posterior tibial margin.
Semimembranosus is supplied from the perforating arteries, usually from all, though predominantly from the first. Sometimes the predominant artery arises from the fourth perforator. A branch of the femoral or popliteal artery supplies the distal part of the muscle, and there may be a contribution from the inferior gluteal artery at the proximal attachment.
Biceps femoris
Biceps femoris (see Fig. 80.2) occupies a posterolateral position in the thigh. It has two proximal attachments. One, the long head, arises from an inferomedial impression on the upper area of the ischial tuberosity (Fig. 80.5), via a tendon which it shares with semitendinosus, and from the lower part of the sacrotuberous ligament. The other, the short head, arises from the lateral lip of the linea aspera, between adductor magnus and vastus lateralis. This attachment extends proximally almost to gluteus maximus and distally along the lateral supracondylar line to within 5 cm of the lateral femoral condyle, and from the lateral intermuscular septum. The long head forms a fusiform belly that descends laterally across the sciatic nerve. The fibres end in an aponeurosis which covers the posterior surface of the muscle. This aponeurosis receives on its deep surface the fibres of the short head, and gradually narrows to a tendon (the lateral hamstring). The main part of the tendon splits round the fibular collateral ligament and is attached to the head of the fibula. The remainder splits into three laminae. The intermediate lamina fuses with the fibular collateral ligament, while the others pass superficial and deep to the ligament to attach to the lateral condyle of the tibia.
The short head may be absent. Additional slips may arise from the ischial tuberosity, linea aspera, or medial supracondylar line.
Proximally biceps femoris is covered by gluteus maximus, but elsewhere it lies relatively superficially. Deep to it lie semimembranosus proximally, and the sciatic nerve, adductor magnus and the lateral head of gastrocnemius more distally. Semitendinosus and semimembranosus lie medially. The common fibular nerve descends along the medial border of the tendon and separates it distally from the lateral head of gastrocnemius. As a guide during surgery, it is safest to find a nerve proximally and dissect it free distally. The common fibular nerve is found emerging posterior to the biceps tendon, which is thus a useful guide to locating the nerve and thereby, to avoiding injury to the nerve. For example, when the lateral meniscus is sutured arthroscopically, care must be taken that all needle passages are anterior to biceps. The nerve is adherent to the biceps tendon: if part of the fibular head or the biceps attachment, usually with the insertion of the lateral collateral ligament, is avulsed, then the tendon will exert proximal traction on the nerve. This may contribute to common fibular nerve traction injury. Also, the abnormal position of the nerve may render it vulnerable to inadvertent injury during surgical exploration. A proximal fibular fracture may result in the nerve being trapped in the fracture line. A bursa may lie between the tendon and the fibular collateral ligament.
The long head of biceps femoris is supplied by the first and second perforating arteries, with accessory supplies at the ischial attachment from the inferior gluteal and medial circumflex femoral arteries and in the distal quarter from the lateral superior genicular artery. The short head is supplied superiorly by the second or third perforating artery and inferiorly by the lateral superior genicular artery.
Actions of posterior femoral muscles
Acting from above, the posterior femoral muscles flex the knee. Acting from below, they extend the hip joint, pulling the trunk upright from a stooping posture against the influence of gravity, biceps being the main agent. When the knee is semi-flexed, biceps femoris can act as a lateral rotator and semimembranosus and semitendinosus as medial rotators of the lower leg on the thigh at the knee. When the hip is extended, biceps is a lateral rotator and semimembranosus and semitendinosus are medial rotators of the thigh. As is the case with quadriceps femoris, the adductors and gluteus maximus, the hamstrings are quiescent in easy symmetrical standing. However, any action that takes the centre of gravity in front of a transverse axis through the hip joints, e.g. forward reaching, forward sway at the ankle joints, or forward bending at the hips, is immediately accompanied by strong contraction of the hamstrings. (This is in marked contrast to gluteus maximus, which contracts only when there is a call for powerful extension at the hip joint.)
When the knee is flexed against resistance, the tendon of biceps can be felt lateral to the popliteal fossa. Medial to the fossa, the tendons of gracilis (which is the more medial) and semitendinosus stand out sharply. The semimembranosus tendon is just palpable in the interval between them (and also by deep pressure from a ‘pincer’ grip beyond their margins). There is some evidence that semimembranosus, semitendinosus and biceps femoris, although they cross both hip and knee joints, may produce movement at one of these joints without resisting antagonists at the other. Usually, however, each of these muscles contracts as a whole, and whether or not movement takes place at hip or knee is determined by other muscles which act as fixators of these joints.
Testing
The posterior femoral muscles are tested clinically by active knee flexion against resistance in the supine or prone position. In the prone position the individual hamstring tendons can be identified more easily.
When relaxed, these muscles show considerable variation in length: in some individuals the muscles are so short as to impose a serious limitation on flexion of the trunk at the hip joints when the knees are kept extended. Movements such as stooping must then be accomplished by flexing the vertebral column or squatting. Perhaps the need for more stress on the lumbar spine accounts for the occurrence of hamstring tightness in adolescents who develop spondylolysis. These muscles are prone to tearing, which may be related to the relative complexity of muscles that cross two joints, or to the fact that they are subject to large forces including eccentric contractions.