CHAPTER 42 The back
Most clinical disorders of the back present as low back pain with or without associated lower limb pain, so historically most attention has been paid to the anatomy of the lower (lumbosacral) back. In this Section, the term ‘the back’ will include the whole of the posterior aspect of the trunk and of the neck. The whole of this region has great clinical importance but its anatomy has often been neglected. Recent understanding of the detailed topography of the bony and soft-tissue elements of the lower back owes much to the work of Bogduk (2005).
The soft tissues of the back of the trunk and neck include the skin and subcutaneous fat, the underlying fascial layers and the musculature. The deep, ‘true’ or epaxial muscles lie within compartments in their own fascial ‘skeleton’. The bony framework to which the muscles and fasciae attach includes not only elements of the axial skeleton, i.e. the vertebral column and occiput, but also elements of the pectoral and pelvic girdles as well as the ribs. The occiput is described below, the scapula on page 793, the ribs on page 918 and the pelvis on page 1352.
SKIN
The skin of the back of the trunk is thick and highly protective, but has low discriminatory sensation. The superficial fascia is thick and fatty in most areas of the back. Its attachment to the deeper fascial layers is strong in the midline, especially in the neck, but becomes weaker more laterally. The skin of the back of the neck is thicker than that of the front of the neck, but thinner than that of the back of the trunk. The quantity, texture and distribution of hair vary with sex, race and the individual, though well-defined hair tracts have been delineated (Fig. 42.1).

Fig. 42.1 Hair tracts on the dorsal surface of the body.
(Redrawn by permission from Wood Jones F (ed) 1949 Buchanan’s Manual of Anatomy, 8th edn. London: Baillière Tindall and Cox.)
Lines of skin tension run horizontally in the cervical and lumbosacral regions but form segments of two adjacent circles in the thoracic region (Fig. 42.2).

Fig. 42.2 Lines of skin tension on the dorsum of the trunk and head.
(From Kraissl CL, Plast Reconstruct Surg 8: 1–28, 1951. By permission from Lippincott Williams and Wilkins.)
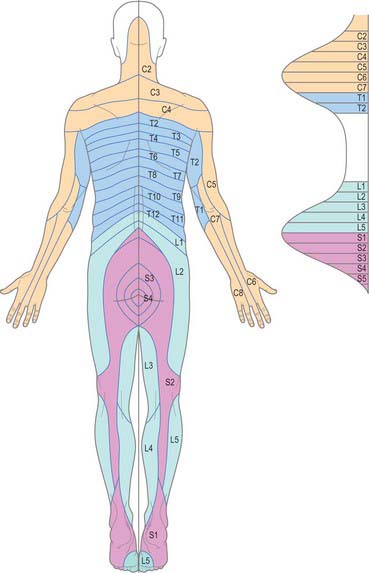
CUTANEOUS INNERVATION AND DERMATOMES
The skin of the back of the neck and trunk is innervated by the dorsal (posterior primary) rami of the spinal nerves (see Fig. 43.6 and pp. 755–756 where dorsal rami are covered in detail). In the cervical and upper thoracic regions (down to T6) skin is supplied by the medial branches of these rami, while in the lower thoracic, lumbar and sacral regions it is supplied by the lateral branches. The total area supplied by these dorsal rami is shown in Fig. 43.6. The spinal nerves involved include C2 to C5, T2 to L3, S2 to S4, and Co1. The pattern of their dermatomes is shown in Fig. 42.3. There is about half a segment of overlap between these cutaneous ‘strips’: the strips supplied by the dorsal rami do not correspond exactly to those served by ventral rami, and differ slightly in both width and position.
CUTANEOUS VASCULAR SUPPLY AND LYMPHATIC DRAINAGE
The skin of the back of the trunk receives its arterial blood supply mainly from musculocutaneous branches of posterior intercostal, lumbar and lateral sacral arteries (Fig. 42.4), which all accompany the cutaneous branches of their respective dorsal rami. In addition, there is a supply from the dominant vascular pedicles of the superficial (extrinsic) back muscles. The skin over the scapula is supplied by branches of the suprascapular, dorsal scapular and subscapular arteries. The skin of the back of the neck is supplied mainly from the occipital and deep cervical arteries. The superficial cervical or transverse cervical artery supplies the skin of the lower part of the back of the neck (see Cormack & Lamberty 1994 in the Bibliography).
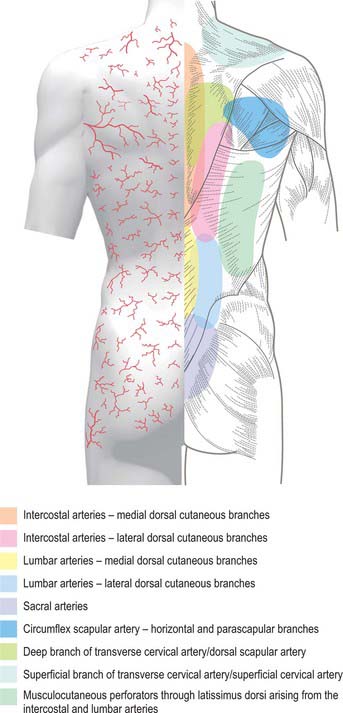
Fig. 42.4 Areas of cutaneous arterial supply on the dorsum of the trunk.
(Redrawn with permission from Cormack GC, Lamberty BGH 1994 The Arterial Anatomy of Skin Flaps. Edinburgh: Churchill Livingstone.)
Veins drain the skin of the back of the neck into tributaries of the occipital and deep cervical veins. The skin of the back of the trunk drains into the azygos system, via tributaries of the posterior intercostal and lumbar veins.
Lymph from the skin of the back of the neck drains into occipital, lateral deep cervical and axillary nodes (Fig. 42.5). From the back of the trunk, drainage is to the posterior (subscapular) axillary nodes and to the lateral superficial inguinal nodes.

FASCIAL LAYERS
The main fascial layers in the axial and paraxial regions of the trunk and neck are the thoracolumbar fascia, the deep cervical fascia and the prevertebral, endothoracic, retroperitoneal and posterior part of the pelvic fasciae (the latter four layers collectively form the continuous prevertebral plane). Other important structures with fascial components are the ligamentum nuchae and the aponeurosis of erector spinae (see below).
THORACOLUMBAR FASCIA
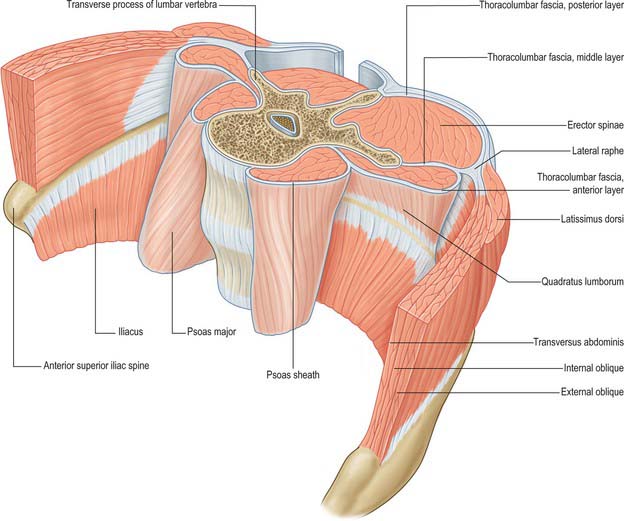
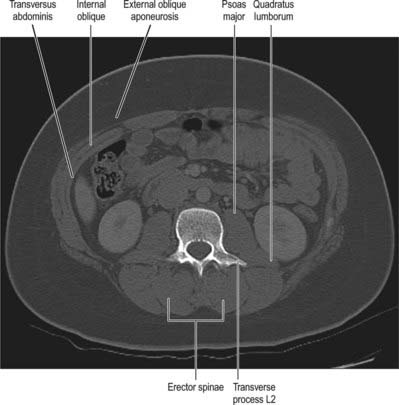
The thoracolumbar (lumbodorsal) fascia covers the deep muscles of the back and the trunk. Above, it passes anterior to serratus posterior superior and is continuous with the superficial lamina of the deep cervical fascia on the back of the neck. In the thoracic region the thoracolumbar fascia provides a thin fibrous covering for the extensor muscles of the vertebral column and separates them from the muscles connecting the vertebral column to the upper extremity. Medially it is attached to the spines of the thoracic vertebrae, and laterally to the angles of the ribs. In the lumbar region the thoracolumbar fascia is in three layers (Fig. 42.6, Fig. 42.7, Fig. 62.1). The posterior layer is attached to the spines of the lumbar and sacral vertebrae and to the supraspinous ligaments. The middle layer is attached medially to the tips of the lumbar transverse processes and the intertransverse ligaments, below to the iliac crest, and above to the lower border of the 12th rib and the lumbocostal ligament. The anterior layer covers quadratus lumborum and is attached medially to the anterior surfaces of the lumbar transverse processes behind the lateral part of psoas major; below, it is attached to the iliolumbar ligament and the adjoining part of the iliac crest; above, it forms the lateral arcuate ligament. The posterior and middle layers unite to form a tough raphe at the lateral margin of erector spinae, and at the lateral border of quadratus lumborum they are joined by the anterior layer to form the aponeurotic origin of transversus abdominis. At sacral levels, the posterior layer is attached to the posterior superior iliac spine and posterior iliac crest, and fuses with the underlying erector spinae aponeurosis. Bogduk (2005) describes two laminae in the posterior layer at lumbar levels, with varying orientation of the constituent collagen fibres relating to the biomechanical function of the fascia. The posterior and middle layers of the thoracolumbar fascia and the vertebral column together form an osteofascial compartment which encloses the erector spinae muscle group. The attachments of the fascia, especially those which give it continuity with the abdominal wall musculature, putatively give it an important role in lifting, though the exact details of this role remain controversial. The fascia may also play an important role in load transfer between the trunk and the limbs: its tension is affected by the actions of latissimus dorsi, gluteus maximus, and the hamstrings. An erector spinae compartment syndrome may be one cause of low back pain.
DEEP CERVICAL FASCIA
The investing layer of the deep cervical fascia forms the deep fascia of the posterior aspect of the neck (Fig. 28.4). It attaches in the midline to the external occipital protuberance, the ligamentum nuchae and the spine of the seventh cervical vertebra, and splits to enclose trapezius on each side. Inferiorly the posterior part of the investing layer attaches with trapezius to the spine and acromion of the scapula.
BONE
OCCIPITAL BONE
The occipital bone forms much of the back and base of the cranium (Fig. 42.8). It is trapezoid, internally concave and encloses the foramen magnum. It has four parts: basilar (basioccipital), which is the quadrilateral part in front of the foramen magnum; squamous, which is the expanded plate posterosuperior to the foramen; and two lateral (condylar or exoccipital), one on each side of the foramen magnum. The latter is anteromedian and wider behind, with an anteroposterior diameter greater than its transverse diameter.
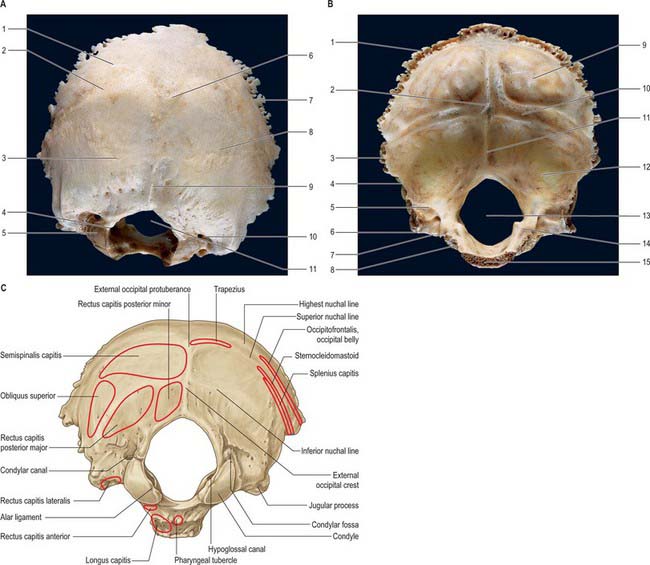
Fig. 42.8 Occipital bone. A, External surface; B, internal surface; C, muscle attachments. (A and B by permission from Berkovitz BKB, Moxham BJ 1994 Colour Atlas of the Skull. London: Mosby.) A: 1. Highest nuchal line. 2. Superior nuchal line. 3. Inferior nuchal line. 4. Hypoglossal canal. 5. Occipital condyle. 6. External occipital protuberance. 7. Lambdoid margin. 8. Squamous part. 9. External occipital crest. 10. Condylar canal. 11. Foramen magnum. B: 1. Lambdoid margin. 2. Internal occipital protuberance at ‘confluence of sinuses’. 3. Lateral angle. 4. Mastoid margin. 5. Groove for sigmoid sinus. 6. Jugular process. 7. Jugular notch. 8. Groove for inferior petrosal sinus. 9. Cerebral fossa. 10. Groove for transverse sinus. 11. Internal occipital crest. 12. Cerebellar fossa. 13. Foramen magnum. 14. Jugular tubercle. 15. Margin of basilar part for articulation with body of sphenoid.
The occipital bone provides attachment for a number of the muscles of the neck and upper part of the back (longus capitis; recti capitis anterior, lateralis, posterior major and minor; superior oblique; semispinalis capitis; splenius capitis; occipitofrontalis; trapezius; sternocleidomastoid), and articulates with the first cervical vertebra at the atlanto-occipital joints.
Squamous part
The squamous part is an expanded plate posterosuperior to the foramen magnum, convex externally and concave internally. On its external surface the external occipital protuberance lies midway between its summit and the foramen magnum. On each side, two curved lines extend laterally from this protuberance. The upper, faintly marked and often almost imperceptible, is the highest nuchal line, and the lower is the superior nuchal line. The epicranial aponeurosis is attached to the medial part of the highest nuchal line. The median external occipital crest is often faint; it descends from the external occipital protuberance to the foramen magnum. On each side an inferior nuchal line spreads laterally from the midpoint of the crest.
The internal surface of the squamous part is divided into four deep fossae by an irregular internal occipital protuberance and by ridged sagittal and horizontal extensions from the protuberance. The two superior fossae are triangular and adapted to the occipital poles of the cerebral hemispheres; the inferior fossae are quadrilateral and shaped to accommodate the cerebellar hemispheres. A wide groove with raised banks, the superior sagittal sulcus, ascends from the protuberance to the superior angle of the squamous part. The posterior part of the falx cerebri is attached to the margins of the sulcus. A prominent internal occipital crest descends from the protuberance and bifurcates near the foramen magnum, and provides an attachment for the falx cerebelli. The occipital sinus, sometimes double, lies in this attachment. A small vermian fossa may exist at the lower end of the internal occipital crest; when present, it is occupied by part of the inferior cerebellar vermis. On each side a wide sulcus for the transverse sinus extends laterally from the internal occipital protuberance; the tentorium cerebelli is attached to the margins of these sulci. The right sulcus is usually larger than the left, and passes into the sulcus for the superior sagittal sinus, whereas the left usually receives the straight sinus. The position of the confluence of the sinuses is indicated by a depression on one side of the protuberance.
The position of the fetal posterior fontanelle coincides with the junction between the superior angle of the squamous part of the occipital bone and the occipital angle of the parietal bone on either side. The lateral angles of the squamous part are marked internally by the ends of the transverse sulci and project between the parietal and temporal bones. The lambdoid borders extend from superior to lateral angles and are serrated for articulation with the occipital borders of the parietal bones at the lambdoid suture. The mastoid borders extend from the lateral angles to the jugular processes, articulating with the mastoid parts of the temporal bones. A variety of sutural bones (ossicles) may occur at or near the lambda, e.g. the ‘interparietal’ (Inca bone or ossicle of Goethe).
The occipital part of occipitofrontalis is attached to the lateral part of the highest nuchal line (Fig. 42.8C). Trapezius attaches to the medial third of the superior nuchal line and to the external occipital protuberance. Sternocleidomastoid attaches to the lateral half of the superior nuchal line, with splenius capitis just below the lateral third of that line. Semispinalis capitis is attached to the medial part of the area between the superior and inferior nuchal lines; obliquus capitis superior attaches to the lateral part of this area. Rectus capitis posterior major attaches to the lateral part of the inferior nuchal line and to the bone immediately below, and rectus capitis posterior minor attaches to the medial part of the inferior nuchal line and to the bone between that line and the foramen magnum.
Basilar part
The basilar part extends anterosuperiorly from the foramen magnum. In young skulls it presents a rough and uneven surface where it is joined to the body of the sphenoid by a growth cartilage (spheno-occipital synchondrosis). This plate has fully ossified by the 25th year, at which time the occipital and sphenoid bones are fused.
The inferior surface bears a small pharyngeal tubercle, about 1 cm in front of the foramen magnum, which gives attachment to the fibrous pharyngeal raphe. A small depression immediately anterior to the occipital condyle may occasionally be replaced by a small precondylar tubercle. The anterior atlanto-occipital membrane is attached to the anterior margin of the foramen magnum.
The superior surface has the form of a broad groove which slopes upwards and forwards from the foramen magnum, directly into the basilar part of the sphenoid: together these bones form the clivus. The lateral margins articulate below with the petrous part of the temporal bones.
Longus capitis is attached anterolateral to the pharyngeal tubercle, and rectus capitis anterior is attached to the small depression anterior to the occipital condyle (Fig. 42.8C).
Lateral (condylar) parts
The lateral (condylar) parts of the occipital bone flank the foramen magnum. On their inferior surfaces are occipital condyles for articulation with the superior articular facets of the atlas vertebra. The condyles are oval or reniform, their long axes converging anteromedially. The articular surfaces, wholly convex, face inferolaterally. They are occasionally constricted and a condyle may be in two parts (as may be the reciprocal surfaces of the atlas vertebra). A tubercle gives attachment to an alar ligament medial to each articular facet. The hypoglossal (anterior condylar) canal, which is situated anteriorly above each condyle, starts internally a little above the anterolateral part of the foramen magnum and continues anterolaterally. It may be partly or wholly divided by a spicule of bone and transmits the hypoglossal nerve and a meningeal branch of the ascending pharyngeal artery. A condylar fossa, behind each condyle, fits the posterior margin of the superior facet of the atlas vertebra in full extension of the skull. Its floor is sometimes perforated by a posterior condylar canal for a sigmoid emissary vein. A quadrilateral plate, the jugular process, projects laterally from the posterior half of each condyle, and contributes the posterior part of the jugular foramen. The jugular process is indented in front by a jugular notch, which may be partly divided by a small intrajugular process projecting anterolaterally. A paramastoid process sometimes projects downwards and may even articulate with the transverse process of the atlas vertebra. Laterally, the jugular process has a rough quadrilateral or triangular area that is joined to the jugular surface of the temporal bone by cartilage: it begins to ossify at around 25 years.
An oval jugular tubercle overlies the hypoglossal canal on the superior surface of the occipital condyle. Its posterior part often bears a shallow furrow for the glossopharyngeal, vagus and accessory nerves. A deep groove containing the end of the sigmoid sinus curves anteromedially around a hook-shaped process to end at the jugular notch. The posterior condylar canal opens into the posterior cranial fossa near the medial end of the groove.
Rectus capitis lateralis attaches to a roughened area on the inferior surface of the jugular process (Fig. 42.8C).
Ossification
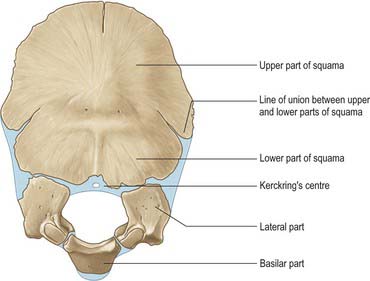
Above the highest nuchal lines the squamous part of the occipital bone is developed in a fibrous membrane and is ossified from two centres (one on each side) from about the second fetal month. This part of the occipital bone may remain separate as the interparietal bone. The remainder of the occipital bone is preformed in cartilage. Below the highest nuchal lines, the squamous part ossifies from two centres that appear in about the seventh week and soon unite. The two components of the squamous part unite in the third postnatal month, but the line of their union is recognizable at birth. The remainder of the cartilage of the occipital bone is ossified from five centres; two each for the lateral parts appear during the eighth week, and one for the basilar part appears around the sixth week.
At birth the occipital bone consists of four separate parts (Fig. 42.9), a basilar part, two lateral parts and a squamous part, all joined by cartilage and forming a ring around the foramen magnum. The squamous and lateral parts fuse together from the second year. The lateral parts fuse with the basilar part during years 3 and 4, but fusion may be delayed until the 7th year.
VERTEBRAL COLUMN
The vertebral column is a curved linkage of individual bones or vertebrae (Fig. 42.10). A continuous series of vertebral foramina runs through the articulated vertebrae posterior to their bodies, and collectively constitutes the vertebral canal, which transmits and protects the spinal cord and nerve roots, their coverings and vasculature (Fig. 42.11, 42.16). A series of paired lateral intervertebral foramina transmits the spinal nerves and their associated vessels between adjacent vertebrae. The linkages between the vertebrae include cartilaginous interbody joints and paired synovial facet (zygapophysial) joints, together with a complex of ligaments and overlying muscles and fasciae. The muscles directly concerned with vertebral movements and attached to the column lie mainly posteriorly. Several large muscles producing major spinal movements lie distant from the column and without direct attachment to it, e.g. the anterolateral abdominal wall musculature. Movements of the column and the muscles concerned are described on page 743. The column as a whole receives its vascular supply and innervation according to general anatomical principles which are considered below.
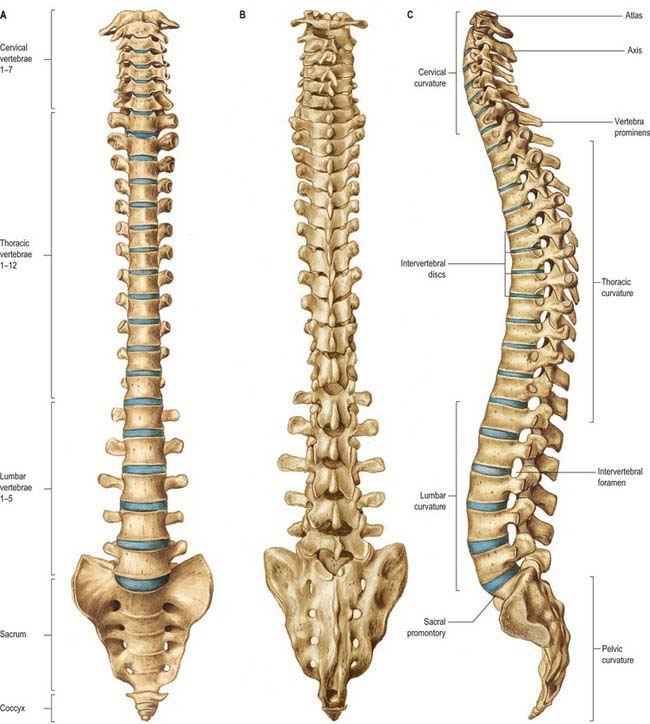
Fig. 42.10 The vertebral column: A, anterior aspect; B, posterior aspect; C, lateral aspect.
(From Sobotta 2006.)
Vertebral column morphology is influenced externally by mechanical and environmental factors and internally by genetic, metabolic and hormonal factors. These all affect its ability to react to the dynamic forces of everyday life, such as compression, traction and shear. These dynamic forces can vary in magnitude and are much influenced by occupation, locomotion and posture.
The adult vertebral column usually consists of 33 vertebral segments. Each presacral segment (except the first two cervical) is separated from its neighbour by a fibrocartilaginous intervertebral disc. The functions of the column are to support the trunk, to protect the spinal cord and nerves, and to provide attachments for muscles. It is also an important site of haemopoiesis throughout life. Its total length in males is about 70 cm and in females about 60 cm. The intervertebral discs contribute about a quarter of this length in young adults, though there is some diurnal variation in this contribution (p. 744). Approximately 8% of overall body length is accounted for by the cervical spine, 20% by the thoracic, 12% by the lumbar and 8% by the sacrococcygeal regions. Although the usual number of vertebrae is 7 cervical, 12 thoracic, 5 lumbar, 5 sacral and 4 coccygeal, this total is subject to frequent variability, and there have been reports of variation between 32 and 35 bones. The demarcation of groups by their morphological characteristics may be blurred: thus there may be thoracic costal facets on the seventh cervical vertebra, giving it the appearance of an extra thoracic vertebra; lumbar-like articular processes may be found on the lowest thoracic vertebra; the fifth lumbar vertebra may be wholly or partially incorporated into the sacrum. As a result of these changes in transition between vertebral types, there may be 23–25 mobile presacral vertebrae.
Anterior aspect
The anterior aspect of the column is formed by the anterior surfaces of the vertebral bodies and of the intervertebral discs (Fig. 42.10A). It has important anatomical relations at all levels, and should be considered in continuity. It forms part of several clinically significant junctional or transitional zones, including the prevertebral/retropharyngeal zone of the neck, the thoracic inlet, the diaphragm and the pelvic inlet. The anterior aspect of the column is covered centrally by the anterior longitudinal ligament, which forms a fascial plane with the prevertebral and endothoracic fascia and with the subperitoneal areolar tissue of the posterior abdominal wall. Infection and other pathological processes may spread along this fascial plane.
Lateral aspect
The lateral aspect of the vertebral column is arbitrarily separated from the posterior by articular processes in the cervical and lumbar regions and by transverse processes in the thoracic region (Fig. 42.10C). Anteriorly it is formed by the sides of vertebral bodies and intervertebral discs. The oval intervertebral foramina, behind the bodies and between the pedicles, are smallest at the cervical and upper thoracic levels, and increase progressively in size in the thoracic and upper lumbar regions. The lumbosacral (L5/S1) intervertebral foramen is the smallest of the lumbar foramina. The foramina permit communication between the lumen of the vertebral canal and the paravertebral soft tissues (a ‘paravertebral space’ is sometimes described), which may be important in the spread of tumours and other pathological processes. The lateral aspects of the column have important anatomical relations, some of which vary considerably between the two sides.
Posterior aspect
The posterior aspect of the column is formed by the posterior surfaces of the laminae and spinous processes, their associated ligaments, and the facet joints (Fig. 42.10B). It is covered by the deep muscles of the back.
Structural defects of the posterior bony elements
Deformity and bony deficiency may occur at several sites within the posterior elements. The laminae may be wholly or partially absent, or the spinous process alone may be affected, with no abnormalities in the overlying soft tissues (spina bifida occulta). A defect may occur in the part of the lamina between the superior and inferior articular processes (pars interarticularis): this condition is spondylolysis, and may be developmental or result from acute or fatigue fracture. If such defects are bilateral, the column becomes unstable at that level, and forward displacement of that part of the column above (cranial to) the defects may occur: this is spondylolisthesis. Abnormality of the laminar bone, or degenerative changes in the facet joints, may also lead to similar displacement in the absence of pars defects. The deformity of the vertebral canal resulting from severe spondylolisthesis may lead to neural damage. Much more rarely, bony defects may occur elsewhere in the posterior elements, e.g. in the pedicles.
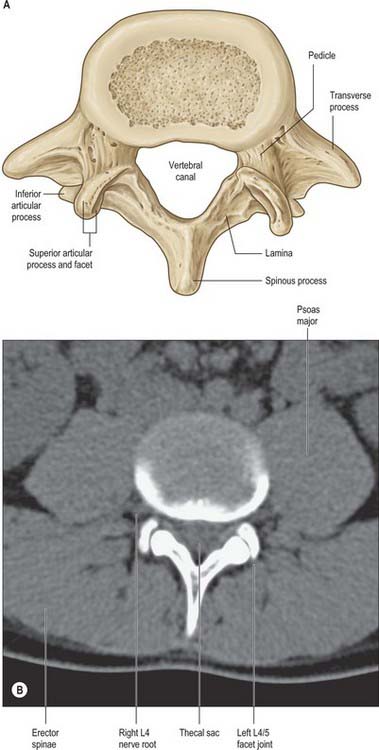
Detailed anatomical relations of all aspects of the vertebral column at the various levels are best appreciated by the study of horizontal (axial) sections and images (see Fig. 42.33B and Fig. 42.55).
Curvatures
Embryonic and fetal curvatures
The embryonic body appears flexed. It has primary thoracic and pelvic curves which are convex dorsally. Functional muscle development leads to the early appearance of secondary cervical and lumbar spinal curvatures in the sagittal plane. The cervical curvature appears at the end of the embryonic period, and reflects the development of function in the muscles responsible for head extension, an important component of the ‘grasp reflex’. Radiographic examination of human fetuses aged from 8 to 23 weeks shows that the secondary cervical curvature is almost always present. Lumbar flattening has also been identified as early as the eighth week. Ultrasound investigations support the role of movement in the development of these curvatures. The early appearance of the secondary curves is probably accentuated by postnatal muscular and nervous system development at a time when the vertebral column is highly flexible and is capable of assuming almost any curvature.
Neonatal curvatures
In the neonate the vertebral column has no fixed curvatures. It is particularly flexible and if dissected free from the body it can easily be bent (flexed or extended) into a perfect half circle. A slight sacral curvature can be seen which develops as the sacral vertebrae ossify and fuse. The thoracic part of the column is the first to develop a relatively fixed curvature, which is concave anteriorly. An infant can usually support its head at 3 or 4 months, sit upright at around 9 months, and will commence walking between 12 and 15 months. These functional changes exert a major influence on the development of the secondary curvatures in the vertebral column and changes in the proportional size of the vertebrae, in particular in the lumbar region. The secondary lumbar curvature becomes important in maintaining the centre of gravity of the trunk over the legs when walking starts, and thus changes in body proportions exert a major influence on the subsequent shape of curvatures in the vertebral column.
Adult curvatures
In adults, the cervical curve is a lordosis (convex forwards), and the least marked. It extends from the atlas to the second thoracic vertebra, with its apex between the fourth and fifth cervical vertebrae. Sexual dimorphism has been described in the cervical curvatures. The thoracic curve is a kyphosis (convex dorsally). It extends between the second and the 11th and 12th thoracic vertebrae, and its apex lies between the sixth and ninth thoracic vertebrae. This curvature is caused by the increased posterior height of the thoracic vertebral bodies. The lumbar curve is also a lordosis. It has a greater magnitude in females and extends from the 12th thoracic vertebra to the lumbosacral angle: there is an increased convexity of the last three segments as a result of the greater anterior height of the intervertebral discs and some posterior wedging of the vertebral bodies. Its apex is at the level of the third lumbar vertebra. The pelvic curve is concave anteroinferiorly and involves the sacrum and coccygeal vertebrae. It extends from the lumbosacral junction to the apex of the coccyx.
The presence of these curvatures means that the cross-sectional profile of the trunk changes with spinal level. The anteroposterior diameter of the thorax is much greater than that of the lower abdomen. In the normal vertebral column there are well-marked curvatures in the sagittal plane and no lateral curvatures other than in the upper thoracic region, where there is often a slight lateral curvature, convex to the right in right-handed persons, and to the left in the left-handed. Compensatory lateral curvature may also develop to cope with pelvic obliquity such as that imposed by inequality of leg length. The sagittal curvatures are present in the cervical, thoracic, lumbar and pelvic regions (Fig. 42.10). These curvatures have developed with rounding of the thorax and pelvis as an adaptation to bipedal gait.
Vertebral column in the elderly
In older people, age-related changes in the structure of bone lead to broadening and loss of height of the vertebral bodies. These changes are more severe in females. The bony changes in the vertebral column are accompanied by changes in the collagen content of the discs and by decline in the activity of the spinal muscles. This leads to progressive decline in vertebral column mobility, particularly in the lumbar spine. The development of a ‘dowager’s hump’ in the midthoracic region in females, caused by age-related osteoporosis, increases the thoracic kyphosis and cervical lordosis. Overall, these changes in the vertebral column lead directly to loss of total height in the individual.
In mid-lumbar vertebrae the width of the body increases with age. In men there is a relative decrease of posterior to anterior body height, while in both sexes anterior height decreases relative to width. Twomey et al (1983) observed a reduction in bone density of lumbar vertebral bodies with age, principally as a result of a reduction in transverse trabeculae (more marked in females as a result of postmenopausal osteoporosis), which was associated with increased diameter and increasing concavity in their juxtadiscal surfaces (end-plates).
Other changes affect the vertebral bodies. Osteophytes (bony spurs) may form from the compact cortical bone on the anterior and lateral surfaces of the bodies. Although individual variations occur, these changes appear in most individuals from 20 years onwards. They are most common on the anterior aspect of the body, and never involve the ring epiphysis. Osteophytic spurs are frequently asymptomatic, but may result in diminished movements within the spine.
Vascular supply and lymphatic drainage
Arteries
(See Crock 1996.)
The vertebral column, its contents and its associated soft tissues, all receive their arterial supply from derivatives of dorsal branches of the embryonic intersegmental somatic arteries (see Ch. 59). The named artery concerned depends on the level of the column. These intersegmental vessels persist in the thoracic and lumbar regions as the posterior intercostal and lumbar arteries. In the cervical and sacral regions, longitudinal anastomoses between the intersegmental vessels persist as longitudinal vessels which themselves give spinal branches to the vertebral column. In the neck the postcostal anastomosis becomes most of the vertebral artery, while the post-transverse anastomosis forms most of the deep cervical artery. The ascending cervical artery and the lateral sacral artery are persistent parts of the precostal anastomosis.
In the thorax and abdomen the primitive arterial pattern is retained by the paired branches of the descending aorta which supply the vertebral column (Fig. 42.12A). On each side, the main trunk of the artery (posterior intercostal or lumbar) passes around the vertebral body, giving off primary periosteal and equatorial branches to the body, and then a major dorsal branch. The latter gives off a spinal branch which enters the intervertebral foramen, before itself supplying the facet joints, the posterior surfaces of the laminae and the overlying muscles and skin. There is free anastomosis between these dorsal articular and soft-tissue branches, extending over several segments (Crock & Yoshizawa 1976; Boelderl et al 2002). At cervical and sacral levels the longitudinally running arteries described above have direct spinal branches. The spinal branches are the main arteries of supply to all bony elements of the vertebrae and to the dura and epidural tissues, and also contribute to the supply of the spinal cord and nerve roots via radicular branches. As they enter the vertebral canal the spinal arteries divide into postcentral, prelaminar and radicular branches. The postcentral branches, which are the main nutrient arteries to the vertebral bodies and to the periphery of the intervertebral discs, anastomose beneath the posterior longitudinal ligament with their fellows above and below as well as across the midline (Fig. 42.12A,B). This anastomosis also supplies the anterior epidural tissues and dura. The majority of the vertebral arch, the posterior epidural tissues and dura and the ligamentum flavum are supplied by the prelaminar branches and their anastomotic plexus on the posterior wall of the vertebral canal.
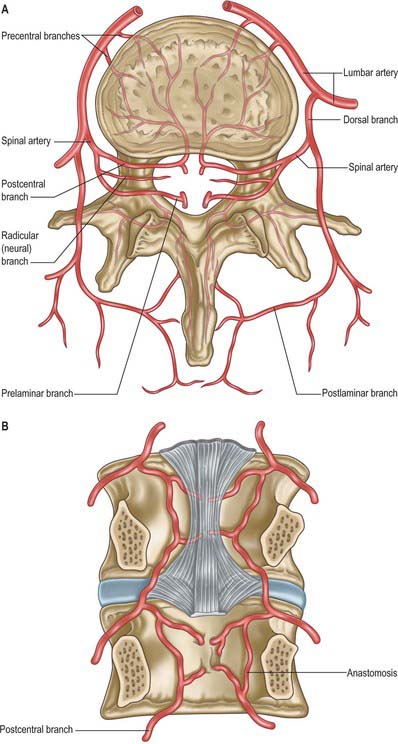
Veins
Veins of the vertebral column form intricate plexuses along the entire column, external and internal to the vertebral canal (Fig. 42.13). Both groups are devoid of valves, anastomose freely with each other, and join the intervertebral veins. Interconnections are widely established between these plexuses and longitudinal veins early in fetal life. When development is complete, the plexuses drain into the caval and azygos/ascending lumbar systems via named veins which accompany the arteries described above.
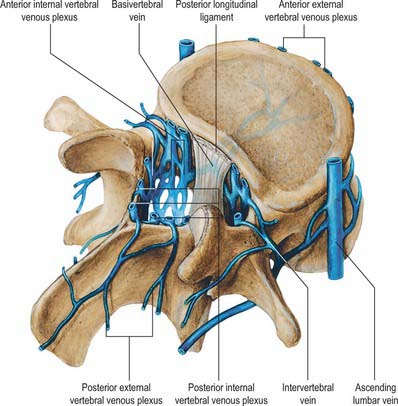
The veins also communicate with cranial dural venous sinuses and with the deep veins of the neck and pelvis. The venous complexes associated with the vertebral column can dilate considerably, and can form alternative routes of venous return in patients with major venous obstruction in the neck, chest or abdomen. The absence of valves allows pathways for the wide and sometimes paradoxical spread of malignant disease and sepsis. Pressure changes in the body cavities are transmitted to these venous plexuses and thus to the CSF, though the cord itself may be protected from such congestion by valves in the small veins which drain from the cord into the internal vertebral plexus.
External vertebral venous plexuses
The external vertebral venous plexuses are anterior and posterior. They anastomose freely, and are most developed in the cervical region. Anterior external plexuses are anterior to the vertebral bodies, communicate with basivertebral and intervertebral veins, and receive tributaries from vertebral bodies. Posterior external plexuses lie posterior to the vertebral laminae and around spines, transverse and articular processes. They anastomose with the internal plexuses and join the vertebral, posterior intercostal and lumbar veins.
Internal vertebral venous plexuses
The internal vertebral venous plexuses occur between the dura mater and vertebrae, and receive tributaries from the bones, red bone marrow and spinal cord. (For the venous drainage of the spinal cord see p. 757 and Fig. 43.11.)
They form a denser network than the external plexuses and are arranged vertically as four interconnecting longitudinal vessels, two anterior and two posterior.
The anterior internal plexuses are large plexiform veins on the posterior surfaces of the vertebral bodies and intervertebral discs. They flank the posterior longitudinal ligament, beneath which they are connected by transverse vessels into which the large basivertebral veins open. The posterior internal plexuses, on each side in front of the vertebral arches and ligamenta flava, anastomose with the posterior external plexuses via veins which pass through and between the ligaments. The internal plexuses interconnect by venous rings near each vertebra. Around the foramen magnum they form a dense network connecting with vertebral veins, occipital and sigmoid sinuses, the basilar plexus, the venous plexus of the hypoglossal canal, and the condylar emissary veins.
Basivertebral veins
The basivertebral veins are large and tortuous channels in bone, like those in the cranial diploë. They emerge from the posterior foramina of the vertebral bodies and drain into the anterior external vertebral plexuses through small openings in the vertebral bodies. Posteriorly they form one or two short trunks that open into the transverse vessels which unite the anterior internal vertebral plexuses. The basivertebral veins enlarge in advanced age.
Intervertebral veins
The intervertebral veins accompany the spinal nerves through intervertebral foramina, draining the spinal cord and internal and external vertebral plexuses, and ending in the vertebral, posterior intercostal, lumbar and lateral sacral veins. Upper posterior intercostal veins may drain into the caval system via brachiocephalic veins, whereas the lower intercostals drain into the azygos system. Lumbar veins are joined longitudinally in front of the transverse processes by the ascending lumbar veins, in which they may terminate. Alternatively, they may proceed around the vertebral bodies to drain into the inferior vena cava. Whether the basivertebral or intervertebral veins contain effective valves is uncertain but experimental evidence strongly suggests that their blood flow can be reversed (Batson 1957). This may explain how pelvic neoplasms, e.g. carcinoma of the prostate, may metastasize in vertebral bodies: the cells spread into the internal vertebral plexuses via their connections with the pelvic veins when blood flow is temporarily reversed by raised intra-abdominal pressure or postural alterations.
Lymphatic drainage
Little is known in detail about the lymphatic drainage of the vertebral column and its associated soft tissues. In general, deep lymphatic vessels tend to follow the arteries. The cervical vertebral column drains to deep cervical nodes, the thoracic to (posterior) intercostal nodes, and the lumbar column to lateral aortic and retro-aortic nodes. The pelvic part of the column drains to lateral sacral and internal iliac nodes.
Innervation
The innervation of the vertebral column and its associated soft tissues has been studied in greatest detail in the lumbar region (see below Fig. 42.44). The account given here relies particularly on the work of Bogduk, to whose textbook on the lumbosacral spine (Bogduk 2005) the interested reader is referred. See also the work of Groen et al (1990).
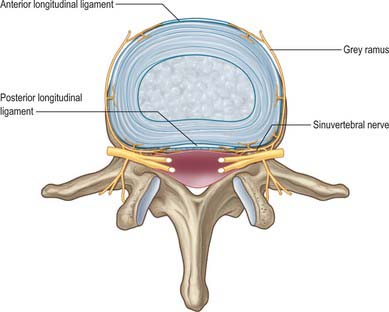
Fig. 42.44 The nerve supply of a lumbar intervertebral disc. Branches of the grey rami communicantes and the sinuvertebral nerves are shown entering the disc and the anterior and posterior longitudinal ligaments. Branches from the sinuvertebral nerves also supply the ventral aspect of the dural sac and the dural nerve-root sleeve.
Innervation is derived from the spinal nerves where they branch, in and just beyond the intervertebral foramina. There is an input from the sympathetic system via either grey rami communicantes or directly from thoracic sympathetic ganglia. The branches of the spinal nerve concerned are the dorsal ramus and the recurrent meningeal or sinuvertebral nerves (usually more than one at each level) (p. 753 and also Fig. 43.5). The dorsal ramus branches to supply the facet joints, periosteum of the posterior bony elements, overlying muscles and skin. The exact origin and branching pattern of the sinuvertebral nerves is controversial, but they may be best considered to be recurrent branches of the ventral rami. They receive the sympathetic input described above, then re-enter the intervertebral foramina to supply the structures that form the walls of the vertebral canal, the dura and epidural soft tissues. Their subsequent course is described on page 754.
VERTEBRAE: GENERAL FEATURES
A typical vertebra has a ventral body, a dorsal vertebral (neural) arch, extended by lever-like processes, and a vertebral foramen, which is occupied in life by the spinal cord, meninges and their vessels (Fig. 42.14).
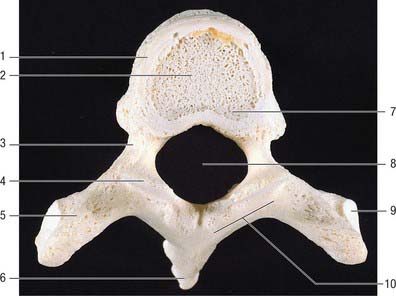
Fig. 42.14 Fourth thoracic vertebra, superior aspect. 1. Bone derived from anular epiphysis. 2. Vertebral body – bone derived from centrum. 3. Pedicle. 4. Superior articular facet. 5. Transverse process. 6. Spinous process. 7. Vertebral body – bone derived from neural arch. 8. Vertebral foramen. 9. Costal facet. 10. Lamina.
Opposed surfaces of adjacent bodies are bound together by intervertebral discs of fibrocartilage. The complete column of bodies and discs forms the strong but flexible central axis of the body and supports the full weight of the head and trunk. It also transmits even greater forces generated by muscles attached to it directly or indirectly. The foraminae form a vertebral canal for the spinal cord, and between adjoining neural arches, near their junctions with vertebral bodies, intervertebral foraminae transmit mixed spinal nerves, smaller recurrent nerves and blood and lymphatic vessels.
The cylindroid vertebral body varies in size, shape and proportions in different regions of the vertebral column. Its superior and inferior (discal) surfaces vary in shape from approximately flat (but not parallel) to sellar, with a raised peripheral smooth zone, formed from an ‘anular’ epiphysial disc, within which the surface is rough. These differences in texture reflect variations in the early structure of intervertebral discs. In the horizontal plane the profiles of most bodies are convex anteriorly, but concave posteriorly where they complete the vertebral foramen. Most sagittal profiles are concave anteriorly but flat posteriorly. Small vascular foramina appear on the front and sides, and posteriorly there are small arterial foramina and a large irregular orifice (sometimes double) for the exit of basivertebral veins (Fig. 42.15). The adult vertebral body is not coextensive with the developmental centrum but includes parts of the neural arch posterolaterally.
Viewed anteriorly there is a cephalocaudal increase in vertebral body width from the second cervical to the third lumbar vertebra, which is associated with an increased load-bearing function. The increase is linear in the neck but not in the thoracic and lumbar regions. There is some variation in size of the last two lumbar bodies, but thereafter width diminishes rapidly to the coccygeal apex. In the two lowest lumbar vertebrae there is an inverse relation between the areas of the upper and lower surfaces of the bodies and the size of the pedicles and transverse processes.
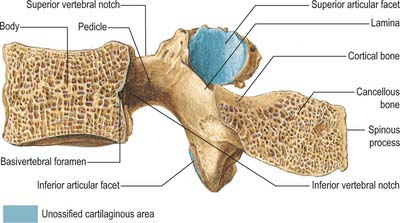
On each side the vertebral arch has a vertically narrower ventral part, the pedicle, and a broader lamina dorsally. Paired transverse, superior and inferior articular processes project from their junctions. There is a median dorsal spinous process.
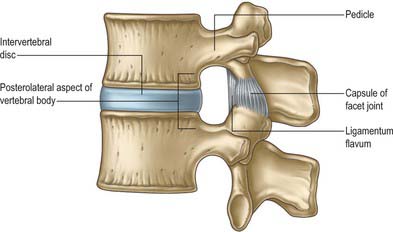
Pedicles are short, thick, rounded dorsal projections from the superior part of the body at the junction of its lateral and dorsal surfaces: the concavity formed by the curved superior border of the pedicle is shallower than the inferior one (Fig. 42.15). When vertebrae articulate by the intervertebral disc and facet joints, these adjacent vertebral notches contribute to an intervertebral foramen. The complete perimeter of an intervertebral foramen consists of the notches, the dorsolateral aspects of parts of adjacent vertebral bodies and the intervening disc, and the capsule of the synovial facet joint.
The laminae are directly continuous with the pedicles. They are vertically flattened and curve dorsomedially.
Lateral to the spinous processes, vertebral grooves contain the deep dorsal muscles. At cervical and lumbar levels these grooves are shallow and mainly formed by laminae. In the thoracic region they are deeper, broader and formed by the laminae and transverse processes. The laminae are broad for the first thoracic vertebra, narrow for the second to seventh, broaden again from the eighth to 11th, but become narrow thereafter down to the third lumbar vertebra.
The spinous process (vertebral spine) projects dorsally and often caudally from the junction of the laminae. Spines vary considerably in size, shape and direction. They lie approximately in the median plane and project posteriorly, although in some individuals a minor deflection of the processes to one side may be seen. The spines act as levers for muscles which control posture and active movements (flexion/extension, lateral flexion and rotation) of the vertebral column.
The paired superior and inferior articular processes (zygapophyses) arise from the vertebral arch at the pediculolaminar junctions. The superior processes project cranially, bearing dorsal facets which may also have a lateral or medial inclination, depending on level. Inferior processes run caudally with articular facets directed ventrally, again with a medial or lateral inclination which depends on vertebral level. Articular processes of adjoining vertebrae thus contribute to the synovial zygapophysial or facet joints, and form part of the posterior boundaries of the intervertebral foramina. These joints permit limited movement between vertebrae: mobility varies considerably with vertebral level.
Transverse processes project laterally from the pediculolaminar junctions as levers for muscles and ligaments, particularly those concerned in rotation and lateral flexion. In the cervical region, the transverse processes are anterior to the articular processes, lateral to the pedicles and between the intervertebral foramina. In the thoracic region, they are posterior to the pedicles, considerably behind those of the cervical and lumbar processes. In the lumbar region, the transverse processes are anterior to the articular processes, but posterior to the intervertebral foramina. There is considerable regional variation in the structure and length of the transverse processes. In the cervical region, the transverse process of the atlas is long and broad, which allows the rotator muscles maximum mechanical advantage. Breadth varies little from the second to the sixth cervical vertebra, but increases in the seventh. In thoracic vertebrae, the first is widest, and breadth decreases to the 12th, where the transverse elements are usually vestigial. The transverse processes become broader in the upper three lumbar vertebrae, and diminish in the fourth and fifth. The transverse process of the fifth lumbar vertebra is the most robust. It arises directly from the body and pedicle to allow for force transmission to the pelvis through the iliolumbar ligament.
The thoracic transverse processes articulate with ribs, but at other levels the mature transverse process is a composite of ‘true’ transverse process and an incorporated costal element. Costal elements develop as basic parts of neural arches in mammalian embryos, but become independent only as thoracic ribs. Elsewhere they remain less developed and fuse with the ‘transverse process’ of descriptive anatomy (Fig. 44.11).
Vertebrae are internally trabecular, and have an external shell of compact bone perforated by vascular foramina (Fig. 42.15). The shell is thin on the superior and inferior body surfaces but thicker in the arch and its processes. The trabecular interior contains red bone marrow and one or two large ventrodorsal canals which contain the basivertebral veins.
Sexual dimorphism in vertebrae has received little attention, but Taylor & Twomey (1984) have described radiological differences in adolescent humans and have reported that female vertebral bodies have a lower ratio of width to depth. Vertebral body diameter has also been used as a basis for sex prediction in the analysis of skeletal material (MacLaughlin & Oldale 1992).
Vertebral canal
The vertebral canal (Fig. 42.16) extends from the foramen magnum to the sacral hiatus, and follows the vertebral curves. In the cervical and lumbar regions, which exhibit free mobility, it is large and triangular, but in the thoracic region, where movement is less, it is small and circular. These differences are matched by variations in the diameter of the spinal cord and its enlargements. In the lumbar region, the vertebral canal decreases gradually in size between L1 and L5, with a greater relative width in the female.
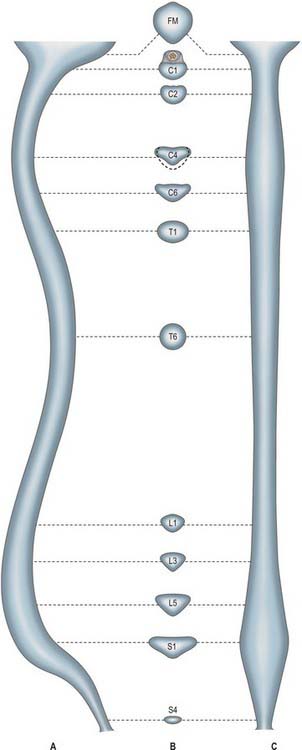
Fig. 42.16 The vertebral canal in section: A, sagittal; B, transverse (axial); C, coronal. FM: foramen magnum.
For clinical purposes it is useful to consider the vertebral canal as having three zones. These are a central zone, between the medial margins of the facet joints, and two lateral zones, beneath the facet joints and entering the intervertebral foramina. Each lateral zone, which passes into and just beyond the intervertebral foramen, can be further subdivided into subarticular (lateral recess), foraminal and extraforaminal regions (Macnab & McCulloch 1990). The lateral zone thus described forms the canal of the spinal nerve (the radicular or ‘root’ canal). The central zone of the canal is a little narrower than the radiological interpedicular distance if the lateral recess is considered to be part of the radicular canal rather than part of the central zone.
Spinal stenosis
Narrowing (stenosis) of the vertebral canal may occur at single or multiple spinal levels, and mainly affects the lumbar and cervical regions. Stenosis may affect the central canal and the ‘root canals’ either together or separately. There is a developmental form of the condition which mainly affects the central canal, but more commonly the stenosis is degenerative, and results from intervertebral disc narrowing and osteoarthritic changes in the facet joints. This latter combination is more likely to narrow the intervertebral foramen and the ‘root canal’, even though the sectional profile of the vertebral canal in affected lumbar vertebrae typically changes from the shape of a bell to that of a trefoil. The lumbosacral intervertebral foramen, which is normally the smallest in the region, is particularly liable to such stenosis. Severe spinal stenosis may compress the spinal cord and compromise its arterial supply. More localized ‘root canal’ stenosis will present with the clinical features of spinal nerve compression, but without the tension signs that characterize the stretching of nerve roots over a prolapsed disc. Ischaemia of the nerves and roots may provoke more damage than the actual physical compression of the neural tissue.
Intervertebral foramina
Intervertebral foramina (see also p. 712) are the principal routes of entry and exit to and from the vertebral canal, and are closely related to the main intervertebral articulations. (Minor routes occur between the median, often partly fused, margins of the ligamenta flava.) The same general arrangement applies throughout the vertebral column, between the axis and sacrum, although there are some quantitative and structural regional variations. Because of their construction, contents and susceptibilities to multiple disorders, the intervertebral foramina are loci of great biomechanical, functional and clinical significance. The specializations cranial to the axis and at sacral levels are described with the individual bones and articulations.
The boundaries of a generalized intervertebral foramen (Fig. 42.17) are anteriorly, from above downwards, the posterolateral aspect of the superior vertebral body, the posterolateral aspect of the intervertebral symphysis (including the disc), and a small (variable) posterolateral part of the body of the inferior vertebra; superiorly, the compact bone of the deep arched inferior vertebral notch of the vertebra above; inferiorly, the compact bone of the shallow superior vertebral notch of the vertebra below; and posteriorly a part of the ventral aspect of the fibrous capsule of the facet synovial joint. Cervical intervertebral foramina are distinct in having superior and inferior vertebral notches of almost equal depth which, in accord with the direction of the pedicles, face anterolaterally. External to them, and oriented in the same direction, is a transverse process. The thoracic and lumbar intervertebral foramina face laterally and their transverse processes are posterior. In addition, the anteroinferior boundaries of the first to tenth thoracic foramina are formed by the articulations of the head of a rib and the capsules of double synovial joints (with the demifacets on adjacent vertebrae and the intra-articular ligament between the costocapitular ridge and the intervertebral symphysis). Lumbar foramina lie between the two principal lines of vertebral attachment of psoas major. The walls of each foramen are covered throughout by fibrous tissue which is in turn periosteal (though the presence of a true periosteum lining the vertebral canal is controversial [Newell 1999]), perichondrial, anular and capsular. The more lateral parts of the foramina may be crossed at a variable level by narrow fibrous bands, the transforaminal ligaments (for detail of these ligaments see Bogduk 2005). The true foramen is the foraminal region of the canal of the spinal nerve (the radicular or ‘root’ canal). A foramen contains a segmental mixed spinal nerve and its sheaths, from two to four recurrent meningeal (sinuvertebral) nerves, variable numbers of spinal arteries, and plexiform venous connections between the internal and external vertebral venous plexuses. These structures, particularly the nerves, may be affected by trauma or one of the many disorders which may affect tissues bordering the foramen. In particular, nerve compression and irritation may be caused by intervertebral disc prolapse, or by bony entrapment as the size of the foramen decreases. This decrease may result from facet joint osteoarthritis, osteophyte formation, disc degeneration and degenerative spondylolisthesis, all of which may lead to lateral or foraminal spinal stenosis.
CERVICAL VERTEBRAE
The cervical vertebrae (Fig. 42.18, Fig. 42.19) are the smallest of the moveable vertebrae, and are characterized by a foramen in each transverse process. The first, second and seventh have special features and will be considered separately. The third, fourth and fifth cervical are almost identical, and the sixth, while typical in its general features, has minor distinguishing differences.
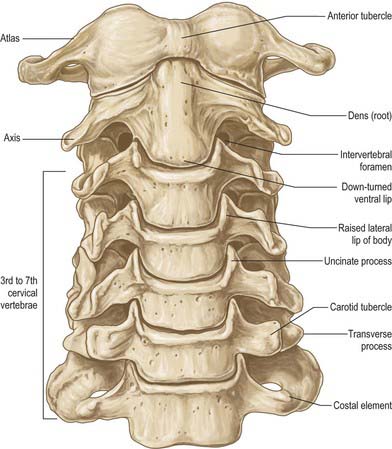
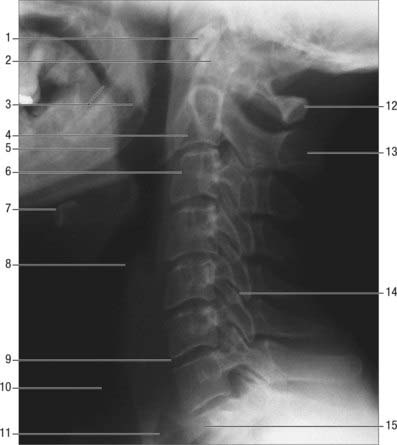
Fig. 42.19 Lateral radiograph of the cervical spine. 1. Anterior tubercle of atlas. 2. Dens of C2. 3. Soft palate. 4. Body of C2. 5. Pharyngeal part of tongue. 6. C3 characteristic cervical body. 7. Body of hyoid. 8. Epiglottis. 9. C6/7 intervertebral disc. 10. Air in trachea. 11. Air in oesophagus. 12. Posterior arch of atlas. 13. Spinous process of axis. 14. C5/6 facet joint. 15. Body of T1.
Typical cervical vertebra
A typical cervical vertebra (Fig. 42.20, Fig. 42.21) has a small, relatively broad vertebral body. The pedicles project posterolaterally and the longer laminae posteromedially, enclosing a large, roughly triangular vertebral foramen; the vertebral canal here accommodates the cervical enlargement of the spinal cord. The pedicles attach midway between the discal surfaces of the vertebral body, so the superior and inferior vertebral notches are of similar depth. The laminae are thin and slightly curved, with a thin superior and slightly thicker inferior border. The spinous process (‘spine’) is short and bifid, with two tubercles which are often unequal in size. The junction between lamina and pedicle bulges laterally between the superior and inferior articular processes to form an articular pillar (‘lateral mass’) on each side. The transverse process is morphologically composite around the foramen transversarium. Its dorsal and ventral bars terminate laterally as corresponding tubercles. The tubercles are connected, lateral to the foramen, by the costal (or intertubercular) lamella: these three elements represent morphologically the capitellum, tubercle and neck of a cervical costal element (p. 770, Fig. 44.11). The attachment of the dorsal bar to the pediculolaminar junction represents the morphological transverse process and the attachment of the ventral bar to the ventral body represents the capitellar process. In all but the seventh cervical vertebra, the foramen transversarium normally transmits the vertebral artery and vein and a branch from the cervicothoracic ganglion (vertebral nerve).
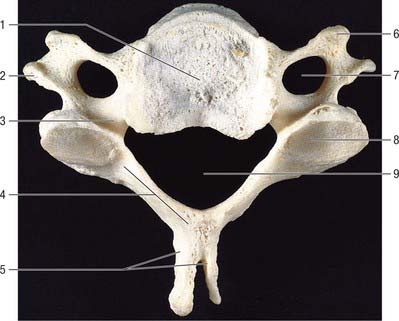
Fig. 42.20 Fourth cervical vertebra, superior aspect. 1. Body. 2. Posterior tubercle of transverse process. 3. Pedicle. 4. Lamina. 5. Bifid spinous process. 6. Anterior tubercle of transverse process. 7. Foramen transversarium. 8. Superior articular facet. 9. Vertebral foramen.
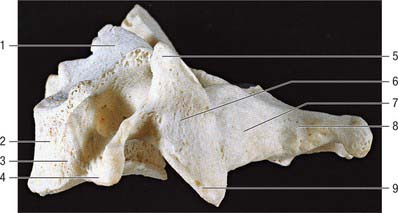
Fig. 42.21 Fourth cervical vertebra, lateral aspect. 1. Uncinate process. 2. Body. 3. Anterior tubercle of transverse process. 4. Posterior tubercle of transverse process. 5. Superior articular process. 6. Lateral mass. 7. Lamina. 8. Spinous process. 9. Inferior articular process.
The vertebral body has a convex anterior surface. The discal margin gives attachment to the anterior longitudinal ligament. The posterior surface is flat or minimally concave, and its discal margins give attachment to the posterior longitudinal ligament. The central area displays several vascular foramina, of which two are commonly relatively larger. These are the basivertebral foramina which transmit basivertebral veins to the anterior internal vertebral veins. The superior discal surface is saddle-shaped, formed by flange-like lips which arise from most of the lateral circumference of the upper margin of the vertebral body; these are sometimes referred to as uncinate or neurocentral lips or processes. The inferior discal surface is also concave: the concavity is produced mainly by a broad projection from the anterior margin which partly overlaps the anterior surface of the intervertebral disc. The discal surfaces of cervical vertebrae are so shaped in order to restrict both lateral and anteroposterior gliding movements during articulation. The paired ligamenta flava extend from the superior border of each lamina below to the roughened inferior half of the anterior surfaces of the lamina above. The superior part of the anterior surface of each lamina is smooth, like the immediately adjacent surfaces of the pedicles, which are usually in direct contact with the dura mater and cervical root sheaths to which they may become loosely attached. The spinous process of the sixth cervical vertebra is larger, and is often not bifid.
The superior articular facets, flat and ovoid, are directed superoposteriorly, whereas the corresponding inferior facets are directed mainly anteriorly, and lie nearer the coronal plane than the superior facets. The dorsal rami of the cervical spinal nerves curve posteriorly, close to the anterolateral aspects of the lateral masses, and may actually lie in shallow grooves, especially on the third and fourth pair. The dorsal root ganglion of each cervical spinal nerve lies between the superior and inferior vertebral notches of adjacent vertebrae. The large anterior ramus passes posterior to the vertebral artery, which lies on the concave upper surface of the costal lamella: the concavity of the lamellae increases from the fourth to the sixth vertebra. The fourth to sixth anterior tubercles are elongated and rough for muscle attachment. The sixth is the longest, the carotid tubercle of Chassaignac. The carotid artery can be forcibly compressed in the groove formed by the vertebral bodies and the larger anterior tubercles, especially the sixth. The posterior tubercles are rounded and more laterally placed than the anterior, and all but the sixth are also more caudal; the sixth is at about the same level as the anterior.
The ligamentum nuchae and numerous deep extensors, including semispinalis thoracis and cervicis, multifidus, spinales and interspinales, are all attached to the spinous processes. Tendinous slips of scalenus anterior, longus capitis and longus colli are attached to the fourth to sixth anterior tubercles. Splenius, longissimus and iliocostalis cervicis, levator scapulae and scalenus posterior and medius are all attached to the posterior tubercles. Shallow anterolateral depressions on the anterior surface of the body lodge the vertical parts of the longus colli.
Cervical vertebrae ossify according to the standard vertebral pattern described on page 768. Incomplete segmentation (‘block vertebra’) is common in the cervical spine and most commonly involves the axis and third cervical vertebra.
C1, atlas
The atlas, the first cervical vertebra (Fig. 42.22), supports the head. It is unique in that it fails to incorporate a centrum, whose expected position is occupied by the dens, a cranial protuberance from the axis. The atlas consists of two lateral masses connected by a short anterior and a longer posterior arch. The transverse ligament retains the dens against the anterior arch.
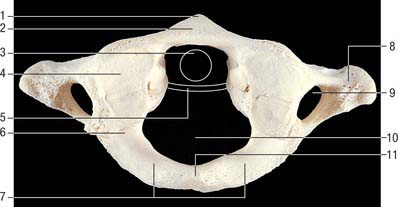
Fig. 42.22 First cervical vertebra (atlas), superior aspect. 1. Anterior tubercle. 2. Anterior arch. 3. Outline of dens. 4. Superior articular facet, on lateral mass (bipartite facet in this specimen). 5. Outline of transverse ligament. 6. Groove for vertebral artery and C1 (beneath bony overhang from lateral mass here). 7. Posterior arch. 8. Transverse process. 9. Foramen transversarium. 10. Vertebral foramen. 11. Posterior tubercle.
The transverse ligament divides the vertebral canal into two compartments. The anterior third (approximately) of the canal is occupied by the dens. The posterior compartment is occupied by the spinal cord and its coverings, and the cord itself takes up about half of this space (i.e. the cord, like the dens, occupies one third of the canal).
The anterior arch is slightly convex anteriorly, and carries a roughened anterior tubercle to which is attached the anterior longitudinal ligament (which is cylindrical at this level). Its upper and lower borders provide attachment for the anterior atlanto-occipital membrane and diverging lateral parts of the anterior longitudinal ligament. The posterior surface of the anterior arch carries a concave, almost circular, facet for the dens.
The lateral masses are ovoid, their long axes converging anteriorly. Each bears a kidney-shaped superior articular facet for the respective occipital condyle, which is sometimes completely divided into a larger anterior and a smaller posterior part (Lang 1986). The inferior articular facet of the lateral mass is almost circular and is flat or slightly concave. It is orientated more obliquely to the transverse plane than the superior facet, and faces more medially and very slightly backwards. On the medial surface of each lateral mass is a roughened area which bears vascular foramina and a tubercle for attachment of the transverse ligament. In adults the distance between these tubercles is shorter than the transverse ligament itself, with a mean value of approximately 16 mm.
The posterior arch forms three-fifths of the circumference of the atlantal ring. The superior surface bears a wide groove for the vertebral artery and venous plexus immediately behind, and is variably overhung by the lateral mass; the first cervical nerve intervenes. The flange-like superior border gives attachment to the posterior atlanto-occipital membrane, and the flatter inferior border to the highest pair of ligamenta flava. The posterior tubercle is a rudimentary spinous process, roughened for attachment of the ligamentum nuchae.
The transverse processes are longer than those of all cervical vertebrae except the seventh (Fig. 42.18). They act as strong levers for the muscles which make fine adjustments to keep the head balanced. Maximum atlantal width varies from 74–95 mm in males and 65–76 mm in females, and this affords a useful criterion for assessing sex in human remains. The apex of the transverse process, which is usually broad, flat and palpable between the mastoid process and ramus of the mandible, is homologous with the posterior tubercle of typical cervical vertebrae: the remaining part of the transverse process consists of the costal lamella. A small anterior tubercle is sometimes visible on the anterior aspect of the lateral mass. The costal lamella is sometimes deficient, which leaves the foramen transversarium open anteriorly.
The superior oblique parts of longus colli are attached on each side of the anterior tubercle. The anterior surface of the lateral mass gives attachment to rectus capitis anterior. Rectus capitis posterior minor is attached just lateral to the posterior tubercle. Rectus capitis lateralis is attached to the transverse process superiorly, and obliquus capitis superior is located more posteriorly. Obliquus capitis inferior is attached laterally on the apex, below which are slips of levator scapulae, splenius cervicis and scalenus medius.
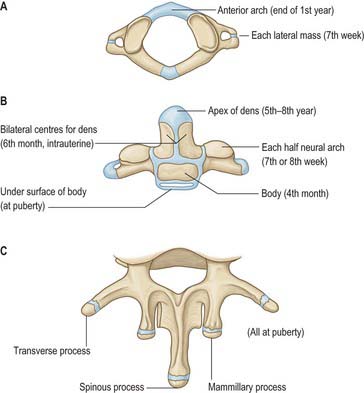
The atlas is commonly ossified from three centres (Fig. 42.23). One appears in each lateral mass at about the seventh week, gradually extending into the posterior arch where they unite between the third and fourth years, usually directly but occasionally through a separate centre. At birth, the anterior arch is fibrocartilaginous, and a separate centre appears about the end of the first year. This unites with the lateral masses between the sixth and eighth year, the lines of union extending across anterior parts of the superior articular facets. Occasionally the anterior arch is formed by the extension and ultimate union of centres in the lateral masses and sometimes from two lateral centres in the arch itself.
The central part of the posterior arch may be absent and replaced by fibrous tissue. Frequently bony spurs arise from the anterior and posterior margins of the groove for the vertebral artery. These are sometimes referred to as ponticles, and they occasionally convert the groove into a foramen. More often the foramen is incomplete superiorly. Rarely the atlas may be wholly or partially assimilated into (fused with) the occiput.
C2, axis
The axis, the second cervical vertebra (Fig. 42.24, Fig. 42.25), acts as an axle for rotation of the atlas and head around the strong dens (odontoid process), which projects cranially from the superior surface of the body.
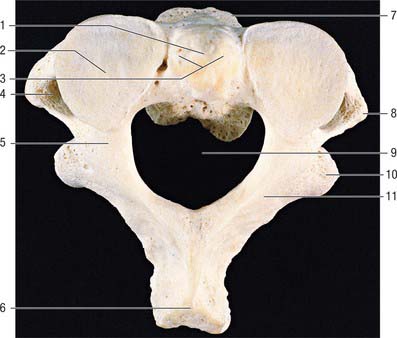
Fig. 42.24 Second cervical vertebra (axis), superior aspect. 1. Dens – attachment of apical ligament. 2. Superior articular facet on lateral mass. 3. Dens – attachments of alar ligaments. 4. Foramen transversarium. 5. Pedicle. 6. Spinous process. 7. Body. 8. Transverse process. 9. Vertebral foramen. 10. Inferior articular process. 11. Lamina.
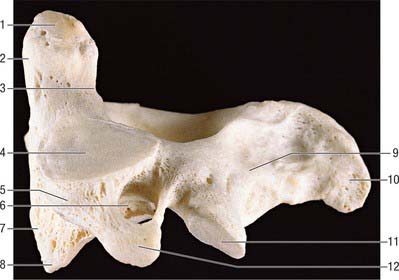
Fig. 42.25 Second cervical vertebra (axis), lateral aspect. 1. Dens – attachment of alar ligament. 2. Facet for anterior arch of atlas. 3. Groove for transverse ligament of atlas. 4. Superior articular facet. 5. Lateral mass. 6. Divergent foramen transversum. 7. Body. 8. Ventral lip of body. 9. Lamina. 10. Spinous process. 11. Inferior articular facet. 12. Transverse process.
The dens is conical in shape with a mean length of 15 mm in adults. It may be tilted a little, up to 14°, posteriorly, or, less often, anteriorly on the body of the axis: it may also tilt laterally up to 10°. The posterior surface bears a broad groove for the transverse ligament which is covered in cartilage. The apex is pointed, and from this point arises the apical ligament. The alar ligaments are attached to the somewhat flattened posterolateral surfaces above the groove for the transverse ligament. The anterior surface bears an ovoid articular facet for the anterior arch of the atlas, and the surface is pitted by many vascular foramina, which are most numerous near the apex.
The body consists of less compact bone than the dens. It is composite, and consists of the partly fused centra of the atlas and axis, and a rudimentary disc (synchondrosis) between them which usually remains detectable deep within the body of the axis throughout life. Large ovoid articular facets are present on either side of the dens at the junction of the body and neural arch: they are flat or slightly convex for articulation with the masses of the atlas. The facets lie in a plane anterior to the plane of the intercentral (Luschka) articulations, with which they are, in part, homologous. The somewhat triangular downward projecting anterior border gives attachment to the anterior longitudinal ligament. Posteriorly, the lower border receives the posterior longitudinal ligament and the membrana tectoria.
The pedicles are stout, and the superior surface carries part of the superior articular facet, which also projects laterally and downwards onto the transverse process. The anterolateral surface is deeply grooved by the vertebral artery, running beneath the thin lateral part of the inferior surface of the superior articular facet, which can become quite thin. The inferior surface of each pedicle bears a deep, smooth inferior intervertebral notch, in which the large root sheath of the third cervical nerve lies. The interarticular part of the pedicle is short and lies between the relatively small inferior posterior articular process (which is located at the pediculolaminar junction and bearing a small anteriorly facing facet) and the superior articular surface.
The transverse process is pointed, projects inferiorly and laterally, and arises from the pediculolaminar junction and the lateral aspect of the interarticular area of the pedicle. The rounded tip is homologous with the posterior tubercle of a typical cervical vertebrae. The foramen transversarium is directed laterally as the vertebral artery turns abruptly laterally under the superior articular facet. Small anterior tubercles may be present near the junction of the costal lamella with the body.
The laminae are thick, and give attachment to the ligamenta flava.
The spinous process is large, with a bifid tip and a broad base, which is concave inferiorly. The ligamentum nuchae is attached to the apical notch.
The anterior surface of the body carries a deep depression on each side for the attachment of the vertical part of longus colli. Levator scapulae, scalenus medius and splenius cervicis are all attached to the tips of the transverse processes and the intertransverse muscles are attached to their upper and lower surfaces. The lateral surfaces of the spinous process give origin to obliquus capitis inferior, and rectus posterior major is attached a little more posteriorly. The inferior concavity of the process receives semispinalis and spinalis cervicis, multifidus more deeply, and the interspinales near the apex.
Small branches arise mainly from the vertebral artery at the level of the intervertebral foramen for the third cervical nerve and form paired anterior and posterior longitudinal channels, branches of which enter the dens near the base and near the apex. The anterior channel also receives numerous twigs from nearby branches of the external carotid artery via branches to longus colli and the ligaments of the apex, hence avascular necrosis does not occur after fracture of the base of the dens.
The axis is ossified from five primary and two secondary centres (Fig. 42.23). The vertebral arch has two primary centres and the centrum one, as in a typical vertebra. The former appear about the seventh or eighth week, and that for the centrum about the fourth or fifth month. The dens is largely ossified from bilateral centres, appearing about the sixth month and joining before birth to form a conical mass, deeply cleft above by cartilage. This cuneiform cartilage forms the apex of the odontoid process. A centre appears in it which shows considerable individual variation in both time of appearance and time of fusion to the rest of the dens: it most often appears between five and eight years, but sometimes even later, fusing with the main mass about the 12th year. The cartilage was thought to be part of the cranial sclerotomal half of the first cervical segment or pro-atlas. It has also been suggested that the apical centre for the dens is itself derived from the pro-atlas, which may also contribute to lateral atlantal masses. The dens is separated from the body by a cartilaginous disc, the circumference of which ossifies while its centre remains cartilaginous until old age; possible rudiments of adjacent epiphyses of atlas and axis may occur in the disc. A thin epiphysial plate is formed inferior to the body around puberty.
Ossification may sometimes be incomplete. Thus the apical cuneiform centre may fail to fuse with the dens, or the dens itself may fail to fuse with the body, instead forming an os odontoideum. Some believe that this results from old unrecognized trauma rather than ossification failure. Interposition of the transverse ligament may prevent union of fractures through the base of the dens. Hypoplasia of the dens is usually accompanied by atlanto-occipital assimilation and basilar invagination. Abnormalities of the dens are common, and can result in atlanto-axial subluxation. In some skeletal dysplasias there is abnormal ossification in which the dens ossifies separately and much later than the atlantal centrum. This is probably a result of abnormal mobility in the cartilaginous anlage, and normal ossification may be restored if motion is prevented by surgical fusion.
C7, seventh cervical vertebra
The seventh cervical vertebra, the vertebra prominens (Fig. 42.26), has a long spinous process which is visible at the lower end of the nuchal furrow. It ends in a prominent tubercle for the attachment of the ligamentum nuchae, and the muscles detailed below. The thick and prominent transverse processes lie behind and lateral to the foramina transversaria. The latter transmit vertebral veins, but not the vertebral artery, and each is often divided by a bony spicule. The costal lamella is relatively thin and may be partly deficient. It is grooved superiorly for the anterior ramus of the seventh cervical nerve, and usually carries a small and inconspicuous anterior tubercle. The posterior tubercle is prominent. The suprapleural membrane is attached to the anterior border of the transverse process.
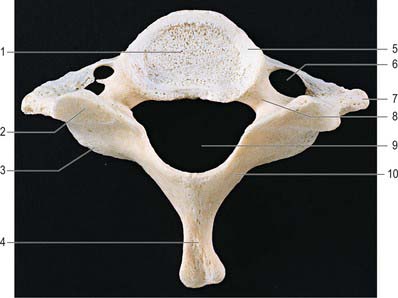
Fig. 42.26 Seventh cervical vertebra, superior aspect. 1. Body. 2. Superior articular facet. 3. Inferior articular process. 4. Spinous process. 5. Uncinate process. 6. Foramen transversarium (foramina are asymmetrical in this specimen). 7. Transverse process. 8. Pedicle. 9. Vertebral foramen. 10. Lamina.
The costal lamella of the transverse process may be separate as a cervical rib. The foramina transversaria may be asymmetrical: sometimes one is absent if the costal lamella is undeveloped.
Trapezius, spinalis capitis, semispinalis thoracis, multifidus and interspinales all attach to the tubercle of the spinous process. The anterior border of the transverse process receives the attachment of scalenus minimus (pleuralis), when present. The first pair of levatores costarum is attached to the transverse processes.
Ossific centres for the costal processes appear about the sixth month and join the body and transverse processes between the fifth and sixth years; they may remain separate and grow anterolaterally as cervical ribs. Separate ossific centres may, on occasion, also occur in the costal processes of the fourth to sixth cervical vertebrae.
THORACIC VERTEBRAE
Thoracic vertebrae in general and changes with descending level
All thoracic vertebral bodies display lateral costal facets and all but the lowest two or three transverse processes also have facets (Fig. 42.27). The facets articulate with the head of the rib (costocapitular facet) and its tubercle (costotubercular facet) respectively. The first and ninth to 12th vertebrae also have atypical features, but the remainder are very similar, except for relatively minor details.
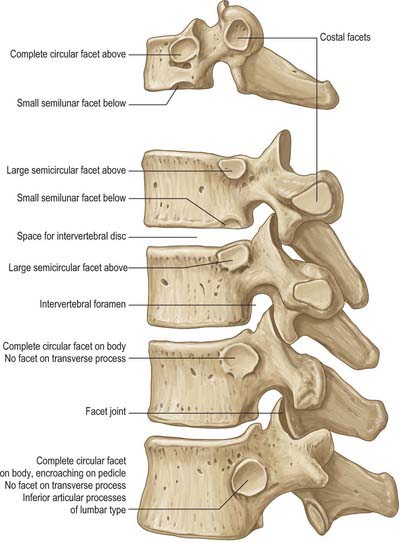
The body is typically a waisted cylinder (Fig. 42.28) except where the vertebral foramen encroaches, and transverse and anteroposterior dimensions are almost equal. On each side there are two costal facets (which are really demifacets): the superior and usually larger pair at the upper border are anterior to the pedicles, while the inferior pair at the lower border are anterior to the vertebral notches. The vertebral foramen (Fig. 42.14) is small and circular, so the pedicles do not diverge as they do in cervical vertebrae: the thoracic spinal cord is smaller and more circular than the cervical cord. The laminae are short, thick and broad, and overlap from above downwards. The spinous process slants downward. The thin and almost flat superior articular processes project from the pediculolaminar junctions and face posteriorly and a little superolaterally. The inferior processes project down from the laminae and their facets are directed forwards and a little superomedially. The large, club-like transverse processes also project from the pediculolaminar junctions. Each passes posterolaterally and bears, near its tip, anterior oval facets for articulation with the tubercle of the corresponding rib.
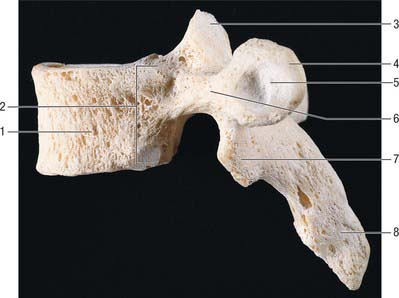
Fig. 42.28 Fourth thoracic vertebra, lateral aspect. 1. Body. 2. Costocapitular demifacets. 3. Superior articular facet. 4. Transverse process. 5. Costotubercular facet. 6. Pedicle. 7. Inferior articular process. 8. Spinous process.
The bodies of upper thoracic vertebrae gradually change from cervical to thoracic in type, and the lower change from thoracic to lumbar. The body of the first is typically cervical, its transverse diameter being almost twice the anteroposterior; the second retains a cervical shape, but its two diameters differ less. The third body is the smallest, and has a convex anterior aspect unlike the flattened first and second thoracic vertebrae. The remaining bodies increase in size and, because of its increased anteroposterior diameter, the fourth is typically ‘heart-shaped’. The fifth to eighth increase their anteroposterior dimension but change little transversely. These four, in transverse section, are asymmetrical, their left sides being flattened by pressure of the thoracic aorta. The rest increase more rapidly in all measurements, so that the 12th body resembles that of a typical lumbar vertebra. These modifications may contribute to the greater range of flexion-extension seen at the cervical and lumbar ends of the thoracic vertebral column.
The anterior and posterior longitudinal ligaments are attached to the borders of the bodies, and around the margins of the costal facets there are the capsular and radiate ligaments of the costovertebral joints. Thoracic pedicles show a successive caudal increase in thickness. The superior vertebral notch is recognizable only in the first thoracic vertebra, whereas the inferior notch is deep in all. Ligamenta flava are attached at the upper borders and lower anterior surfaces of the laminae.
Thoracic transverse processes shorten in caudal succession. In the upper five or six vertebrae the costal facets are concave and face anterolaterally, and at lower levels the facets are flatter and face superolaterally and slightly forwards. The costotransverse ligament is attached to the anterior surface medial to the facet; the lateral costotransverse ligament is attached to its tuberculated apex and the superior costotransverse ligament is attached to its lower border.
Thoracic spines overlap from the fifth to the eighth vertebra, whose spine is the longest and most oblique. Supraspinous and interspinous ligaments are attached to the spines.
A change in orientation of articular processes from thoracic to lumbar type usually occurs at the 11th thoracic vertebra, but sometimes at the 12th or 10th. In the transitional vertebra the superior articular processes are thoracic, and face posterolaterally, while the inferior are transversely convex and face anterolaterally. The transitional vertebra marks the site of a sudden change of mobility from predominantly rotational to predominantly flexion–extension.
Longus colli arises from the upper three thoracic vertebral bodies, lateral to the anterior longitudinal ligament, and psoas major and minor arise from the sides of the 12th near its lower border. Upper and lower borders of the transverse processes provide attachment for the intertransverse muscles or their fibrous vestiges. The posterior surfaces of the transverse processes provide attachment for the deep dorsal muscles, and levator costae is attached posteriorly on the apex. Trapezius, rhomboid major and minor, latissimus dorsi, serratus posterior superior and inferior and many deep dorsal muscles are attached to the spines. Rotatores attach to the posterior aspects of the laminae.
Thoracic vertebrae all ossify according to the standard vertebral pattern described on page 768.
T1
The first thoracic vertebra (Fig. 42.27) resembles a cervical vertebra in its body, both in shape and in the distinctive posterolateral ‘lipping’ which forms the anterior border of the superior vertebral notch. There are circular superior costal facets for articulation with the whole facet on the head of the first rib. The smaller, semilunar inferior facets articulate with a demifacet on the head of the second rib. The upper costal facet is often incomplete, in which case the first rib articulates with the seventh cervical vertebra and the intervening disc. A small, deep depression often occurs below the facet. The long, thick spine is horizontal and commonly as prominent as that of the seventh cervical vertebra.
T9
The ninth thoracic vertebra (Fig. 42.27) is otherwise typical, but it often fails to articulate with the tenth ribs, in which case the inferior demifacets are absent.
T10
The 10th thoracic vertebra (Fig. 42.27) only articulates with the 10th pair of ribs, so that superior facets only appear on the body. These are usually large and semilunar, but are oval when the tenth ribs fail to articulate with the ninth vertebra and intervening disc. The transverse process may or may not bear a facet for the tubercle of the 10th rib.
T11
The 11th thoracic vertebra (Fig. 42.27) articulates only with the heads of the 11th ribs. The circular costal facets are close to the upper border of the body and extend onto the pedicles. The small transverse processes lack articular facets. The 11th and 12th thoracic spinous processes are triangular, with blunt apices, a horizontal lower and an oblique upper border.
T12
The 12th thoracic vertebra (Fig. 42.27) articulates with the heads of the 12th ribs by circular facets somewhat below the upper border, spreading on to the pedicles. The body is large and the vertebra has some lumbar features. The transverse process is replaced by three small tubercles: the superior is largest, projects upwards and corresponds to a lumbar mammillary process, though it does not lie as close to the superior articular process; the lateral tubercle is the homologue of a transverse process; the inferior is the homologue of a lumbar accessory process. The superior and inferior processes are surprisingly long in some specimens.
LUMBAR VERTEBRAE
Lumbar vertebrae in general
The five lumbar vertebrae are distinguished by their large size and absence of costal facets and transverse foramina. The body is wider transversely (Fig. 42.29, see Fig. 42.33B). The vertebral foramen is triangular, larger than at thoracic levels but smaller than at cervical levels. The pedicles are short. The spinous process is almost horizontal, quadrangular and thickened along its posterior and inferior borders. The superior articular processes bear vertical concave articular facets facing posteromedially, with a rough mammillary process on their posterior borders (Fig. 42.30A). The inferior articular processes have vertical convex articular facets which face anterolaterally. The transverse processes are thin and long, except on the more substantial fifth pair. A small accessory process marks the posteroinferior aspect of the root of each transverse process. The accessory and mammillary processes are linked by a fine ligament, the mammillo-accessory ligament, which is sometimes ossified, and beneath which runs the medial branch of the dorsal primary ramus of the spinal nerve (Bogduk 2005).
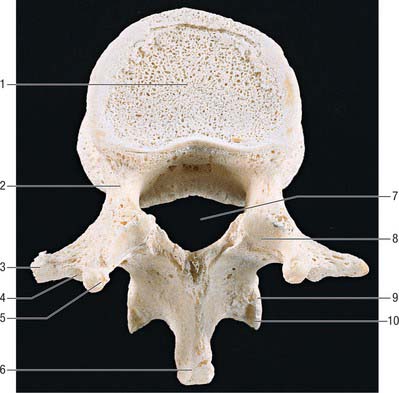
Fig. 42.29 First lumbar vertebra, superior aspect. 1. Body. 2. Pedicle. 3. Transverse process. 4. Accessory process. 5. Mammillary process. 6. Spinous process. 7. Vertebral foramen. 8. Superior articular facet. 9. Lamina. 10. Inferior articular facet.
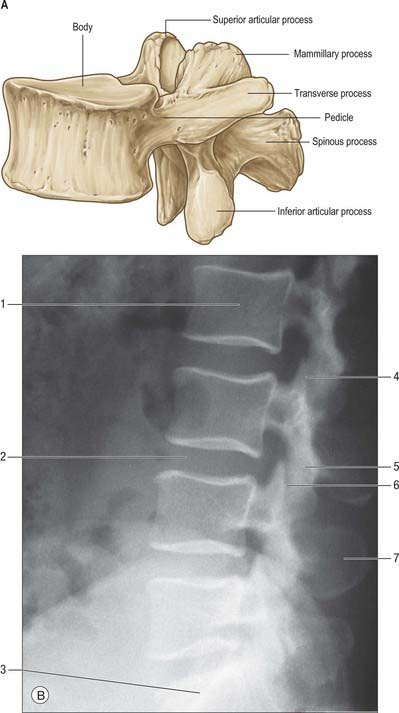
Fig. 42.30 Lumbar spine. A, Lumbar vertebra, lateral aspect. B, Lateral radiograph, 24-year-old male. 1. L2 vertebral body. 2. Site of L3–4 intervertebral disc. 3. S1 vertebral body. 4. Superior facet. 5. Inferior facet. 6. L3–4 facet joint. 7. L4 spinous process.
Strong paired pedicles arise posterolaterally from each body near its upper border. Superior vertebral notches are shallow and the inferior ones are deep. The laminae are broad and short, but do not overlap as much as those of the thoracic vertebrae. The fifth spine is the smallest, and its apex is often rounded and down-turned. Upper lumbar superior articular processes are further apart than inferior ones, but the difference is slight in the fourth and negligible in the fifth. The articular facets are reciprocally concave (superior) and convex (inferior), which allows flexion, extension, lateral bending and some degree of rotation. There are sex differences in the angle of inclination and depth of curvature of the articular facets. The facets are sometimes asymmetrical. Transverse processes, except the fifth, are anteroposteriorly compressed and project posterolaterally. The lower border of the fifth transverse process is angulated, passes laterally and then superolaterally to a blunt tip, and the whole process presents a greater upward inclination than the fourth. The angle on the inferior border may represent the tip of the costal element and the lateral end the tip of the true transverse process. The lumbar transverse processes increase in length from first to third and then shorten (Fig. 42.31). The fifth pair incline both upwards and posterolaterally. The costal element is incorporated in the mature transverse process.
Fig. 42.31 Anteroposterior radiograph of lumbosacral spine in a 22-year-old male. 1. L3 inferior articular process. 2. L4 superior articular process. 3. Right sacroiliac joint. 4. Ilium. 5. Psoas lateral border. 6. L3 transverse process. 7. Facet joint. 8. L4 pedicle. 9. L4 spinous process. 10. Anterior sacral foramen.
The first lumbar vertebral foramen contains the conus medullaris of the spinal cord, while lower foramina contain the cauda equina and spinal meninges (Fig. 42.11B, Fig. 42.32A). Variation occurs in the sagittal and coronal dimensions of the lumbar vertebral canal, both within and between normal populations.
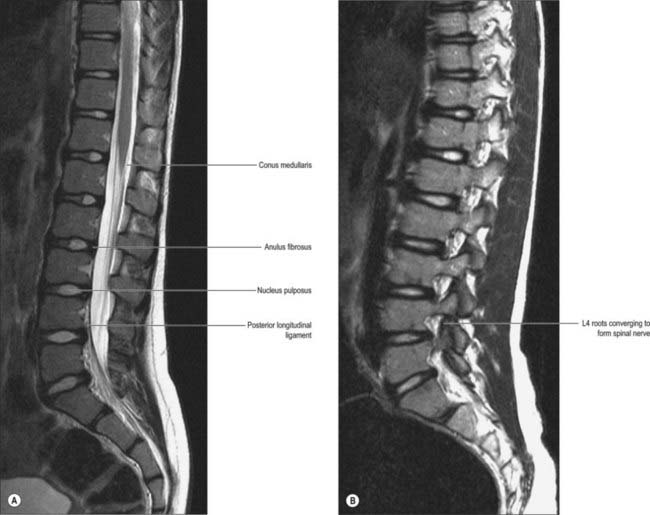
Muscle and fascial attachments (see also p. 708)
Upper and lower borders of lumbar bodies give attachment to the anterior and posterior longitudinal ligaments. The upper bodies (three on the right, two on the left) give attachments to the crura of the diaphragm lateral to the anterior longitudinal ligament. Posterolaterally, psoas major is attached to the upper and lower margins of all the lumbar bodies, and between them, tendinous arches carry its attachments across their concave sides (see Fig. 62.5). The posterior lamella of the thoracolumbar fascia, erectores spinae, spinales thoracis, multifidi, interspinal muscles and ligaments, and supraspinous ligaments are all attached to spinous processes. All lumbar transverse processes present a vertical ridge on the anterior surface, nearer the tip, which marks the attachment of the anterior layer of the thoracolumbar fascia, and separates the surface into medial and lateral areas for psoas major and quadratus lumborum respectively. The middle layer of the fascia is attached to the apices of the transverse processes; the medial and lateral arcuate ligaments attach to the vertical ridge on the anterior aspect of the first pair, and the iliolumbar ligament attaches to the apices of the fifth pair. Posteriorly the transverse processes are covered by deep dorsal muscles, and fibres of longissimus thoracis are attached to them and to their accessory processes. The ventral lateral intertransverse muscles are attached to their upper and lower borders, while the dorsal attach cranially to the accessory process and caudally to the upper border of the transverse process. The mammillary process, homologous with the superior tubercle of the 12th thoracic vertebra, gives attachment to multifidus and the medial intertransverse muscle. The latter also attaches to the accessory process, which is sometimes difficult to identify.
Lumbar vertebrae ossify according to the standard vertebral pattern described on page 768, but also have two additional centres for the mammillary processes. A pair of scale-like epiphyses usually appear on the tips of the costal elements of the fifth lumbar vertebra (Fig. 42.23).
L5
The fifth lumbar vertebra (Fig. 42.33) has a massive transverse process which is continuous with the whole of the pedicle and encroaching on the body. The body is usually the largest and markedly deeper anteriorly, so contributing to the lumbosacral angle.
Segmentation anomalies (sacralization) are considered below with the sacrum. The costal element of the first lumbar vertebra may form a short lumbar rib, which articulates with the transverse process, but not usually with the body, of the vertebra.
SACRUM
The sacrum is a large, triangular fusion of five vertebrae and forms the posterosuperior wall of the pelvic cavity, wedged between the two hip (innominate) bones. Its blunted, caudal apex articulates with the coccyx and its superior, wide base with the fifth lumbar vertebra at the lumbosacral angle. It is set obliquely and curved longitudinally, the dorsal surface is convex, and the pelvic surface is concave: this ventral curvature increases pelvic capacity. Between base and apex are dorsal, pelvic and lateral surfaces and a sacral canal. In childhood, individual sacral vertebrae are connected by cartilage, and the adult bone retains many vertebral features. The sacrum consists of trabecular bone enveloped by a shell of compact bone of varying thickness.
The base (Fig. 42.34) is the upper surface of the first sacral vertebra, the least modified from the typical vertebral plan. The body is large and wider transversely, and its anterior projecting edge is the sacral promontory. The vertebral foramen is triangular, its pedicles are short and diverge posterolaterally. The laminae are oblique, inclining down posteromedially to meet at a spinous tubercle. The superior articular processes project cranially, with concave articular facets directed posteromedially to articulate with the inferior articular processes of the fifth lumbar vertebra. The posterior part of each process projects backwards and its lateral aspect bears a rough area homologous with a lumbar mammillary process.
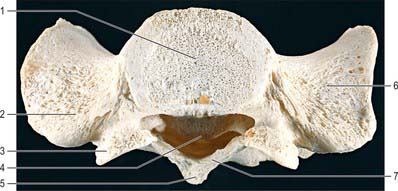
Fig. 42.34 Sacrum, superior aspect (base). 1. Body of S1. 2. Posterosuperior ala (transverse process element). 3. Superior articular process. 4. Sacral canal. 5. Spinous process of S1. 6. Anterosuperior ala (costal element). 7. Lamina.
The transverse process is much modified as a broad, sloping mass which projects laterally from the body, pedicle and superior articular process. It is formed by the fusion of the transverse process and the costal element to each other and to the rest of the vertebra, and forms the upper surface of the sacral lateral mass or ala.
Terminal fibres of the anterior and posterior longitudinal ligaments are attached to the ventral and dorsal surfaces of the first sacral body. Its upper laminar borders receive the lowest pair of ligamenta flava. The ala is smooth superiorly, concave medially and rough laterally, and covered almost entirely by psoas major. The smooth area is grooved obliquely by the lumbosacral trunk. The rough area is for the lower band of the iliolumbar ligament, which lies lateral to the fifth lumbar spinal nerve and to the anterior sacroiliac ligament.
The anteroinferior pelvic surface (Fig. 42.35) is vertically and transversely concave, but the second sacral body may produce a convexity. Four pairs of pelvic sacral foramina communicate with the sacral canal through intervertebral foramina, and transmit ventral rami of the upper four sacral spinal nerves. The large area between the right and left foramina, which is formed by the flat pelvic aspects of the sacral bodies, bears evidence of their fusion at four transverse ridges. The longitudinal bars between the foramina are costal elements, which fuse to the vertebrae. Lateral to the foramina the costal elements unite. Posteriorly they unite with the transverse processes to form the lateral part of the sacrum, which expands basally as the ala.
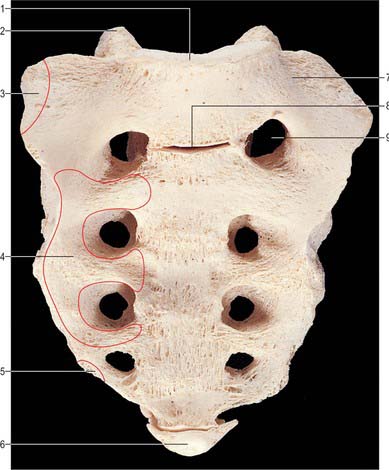
Fig. 42.35 Sacrum, anterior (pelvic) surface. 1. Upper border of body of S1 (sacral promontory). 2. Superior articular process of S1. 3. Attachment of iliacus. 4. Attachment of piriformis. 5. Attachment of coccygeus. 6. Coccyx. 7. Ala. 8. Incompletely fused S1–2 intervertebral joint. 9. First pelvic sacral foramen.
The first three sacral ventral rami emerge from the pelvic sacral foramina and pass anterior to piriformis. The sympathetic trunks descend in contact with bone, medial to the foramina, as do the median sacral vessels in the midline. Lateral to the foramina, lateral sacral vessels are related to bone. Ventral surfaces of the first, second, and part of the third sacral bodies are covered by parietal peritoneum and crossed obliquely, left of the midline, by the attachment of the sigmoid mesocolon. The rectum is in contact with the pelvic surfaces of the third to fifth sacral vertebrae and with the bifurcation of the superior rectal artery between the rectum and third sacral vertebra.
The posterosuperior aspect of the dorsal surface bears a raised, interrupted, median sacral crest with four (sometimes three) spinous tubercles which represent fused sacral spines (Fig. 42.36). Below the fourth (or third) tubercle there is an arched sacral hiatus in the posterior wall of the sacral canal. This hiatus is produced by the failure of the laminae of the fifth sacral vertebra to meet in the median plane, and as a result the posterior surface of the body of that vertebra is exposed on the dorsal surface of the sacrum. Flanking the median crest, the posterior surface is formed by fused laminae, and lateral to this are four pairs of dorsal sacral foramina. Like the pelvic foramina, they lead into the sacral canal through intervertebral foramina, and each transmits the dorsal ramus of a sacral spinal nerve. Medial to the foramina, and vertically below each articular process of the first sacral vertebra, is a row of four small tubercles, which collectively constitute the intermediate sacral crest. These are sometimes termed articular tubercles, and represent fused contiguous articular processes. The inferior articular processes of the fifth sacral vertebra are free and project downwards at the sides of the sacral hiatus as sacral cornua, connected to coccygeal cornua by intercornual ligaments. The interrupted roughened crest to the lateral side of the dorsal sacral foramina is the lateral sacral crest which is formed by fused transverse processes, whose apices appear as a row of transverse tubercles.
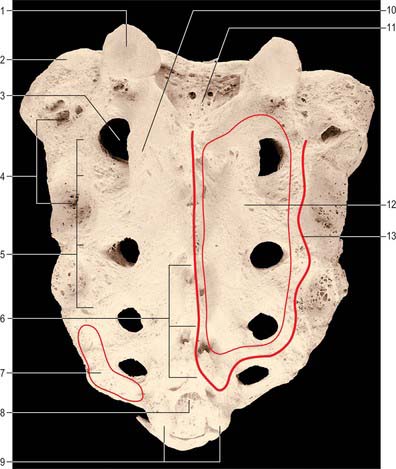
Fig. 42.36 Sacrum, posterior (dorsal) surface. 1. Superior articular facet of S1. 2. Ala. 3. First dorsal sacral foramen. 4. Attachments of interosseous sacroiliac ligaments. 5. Lateral crest and transverse tubercles. 6. Median crest and spinous processes. 7. Attachment of gluteus maximus. 8. Sacral hiatus. 9. Cornua. 10. Intermediate crest and articular tubercle (inferior articular process). 11. Posterior surface of body of S1 forming anterior wall of sacral canal. 12. Area of attachment of multifidus (bounded by thin line). 13. Attachment of erector spinae aponeurosis (thick line).
The upper three sacral spinal dorsal rami pierce multifidus as they emerge via dorsal foramina.
The lateral surface (Fig. 42.37A) is a fusion of transverse processes and costal elements. It is wide above, and rapidly narrows in its lower part. The broad upper part bears an auricular surface for articulation with the ilium, and the area posterior to this is rough and deeply pitted by the attachment of ligaments. The auricular surface, borne by costal elements, is like an inverted letter L. The shorter, cranial limb is restricted to the first sacral vertebra; the caudal limb descends to the middle of the third. Beyond this the lateral surface is non-articular and reduced in breadth. Caudally it curves medially to the body of the fifth sacral vertebra at the inferior lateral angle, beyond which the surface becomes a thin lateral border. A variable accessory sacral articular facet sometimes occurs, posterior to the auricular surface.
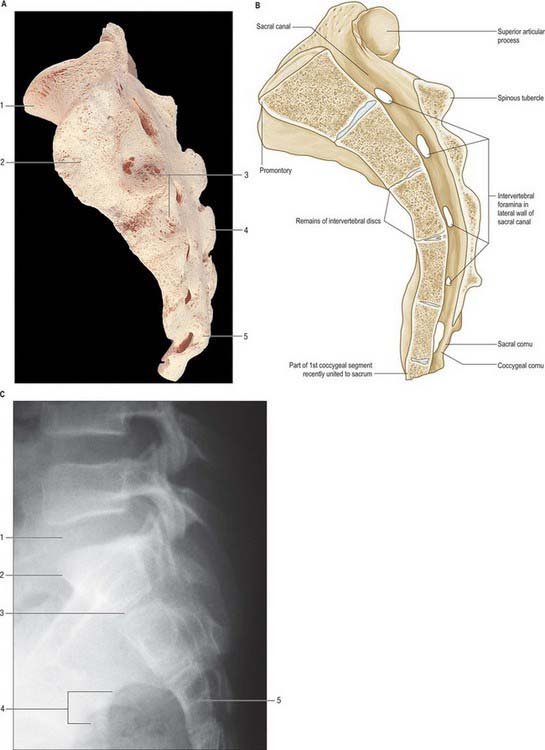
Fig. 42.37 A, Sacrum, lateral aspect: 1. Promontory. 2. Auricular (articular) surface. 3. Attachments of interosseous sacroiliac ligaments. 4. Spinous process. 5. Sacral cornu (left). B, Median sagittal section through the sacrum. C. Lateral radiograph of lumbosacral junction in a 14-year-old male 1. L5–S1 disc. 2. Sacral promontory. 3. S1–2 disc. 4. Greater sciatic notches. 5. S3.
The auricular surface is covered by hyaline cartilage, and formed entirely by costal elements. It shows cranial and caudal elevations and an intermediate depression, behind which a third elevation is visible in the elderly. The surface becomes more corrugated with age. The rough area behind the auricular surface shows two or three marked depressions for the attachment of strong interosseous sacroiliac ligaments. Below the auricular surface the sacrotuberous and sacrospinous ligaments are attached between gluteus maximus dorsally and coccygeus ventrally.
The apex is the inferior aspect of the fifth sacral vertebral body, and bears an oval facet for articulation with the coccyx.
The sacral canal (Fig. 42.37B) is formed by sacral vertebral foramina, and is triangular in section (Fig. 42.34). Its upper opening, seen on the basal surface, appears to be set obliquely. The inclination of the sacrum means that it is directed cranially in the standing position. Each lateral wall presents four intervertebral foramina, through which the canal is continuous with pelvic and dorsal sacral foramina. Its caudal opening is the sacral hiatus. The canal contains the cauda equina and the filum terminale, and the spinal meninges. Opposite the middle of the sacrum, the subarachnoid and subdural spaces close: the lower sacral spinal roots and filum terminale pierce the arachnoid and dura mater at that level. The filum terminale with its meningeal coverings emerges below the sacral hiatus and passes downwards across the dorsal surface of the fifth sacral vertebra and sacrococcygeal joint to reach the coccyx. The fifth sacral spinal nerves also emerge through the hiatus medial to the sacral cornua, and groove the lateral aspects of the fifth sacral vertebra.
The pelvic surface gives attachment to piriformis in its second to fourth segments, to iliacus superolaterally, and to coccygeus inferolaterally. The dorsal surface gives attachment to the aponeurosis of erector spinae along a U-shaped area of spinous and transverse tubercles, covering multifidus which occupies the enclosed area (Fig. 42.34). On the lateral border below the auricular surface, gluteus maximus is attached dorsal and coccygeus is attached ventral to the sacrotuberous and sacrospinous ligaments.
The sacrum resembles typical vertebrae in the ossification of its segments (Fig. 42.38). Primary centres for the centrum and each half vertebral arch appear between the 10th and 20th weeks. Primary centres for the costal elements of the upper three or more segments appear superolateral to the pelvic sacral foramina, between the sixth and eighth prenatal months. Each costal element unites with its half vertebral arch between the second and fifth years, and the conjoined element so formed unites anteriorly with the centrum and posteriorly with its opposite fellow at about the eighth year. Thereafter the upper and lower surfaces of each sacral body are covered by an epiphysial plate of hyaline cartilage which is separated from its neighbour by the fibrocartilaginous precursor of an intervertebral disc. Laterally, successive conjoined vertebral arches and costal elements are separated by hyaline cartilage; a cartilaginous epiphysis, sometimes divided into upper and lower parts, develops on each auricular and adjacent lateral surface. Soon after puberty the fused vertebral arches and costal elements of adjacent vertebrae begin to coalesce from below upwards. At the same time individual epiphysial centres develop for the upper and lower surfaces of bodies, spinous tubercles, transverse tubercles and costal elements.
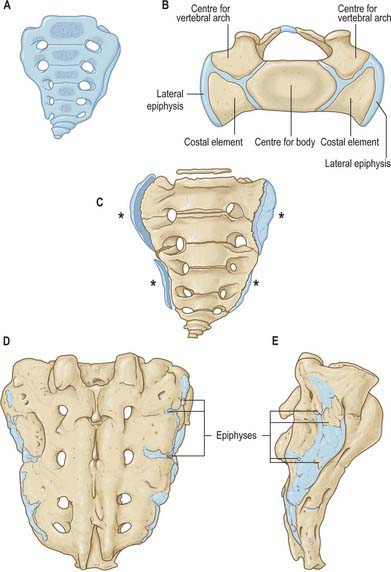
Fig. 42.38 Ossification of the sacrum and coccyx. A, At birth. B, The base of the sacrum of a child about four years old. C, At the 25th year: epiphysial plates for each lateral surface are marked by asterisks. D, E, The epiphyses of the costal and transverse process of the sacrum at the 18th year.
The costal epiphysial centres appear at the lateral extremities of the hyaline cartilages between adjacent costal elements; two anterior and two posterior centres appear in each of the intervals between the first, second and third sacral vertebrae. Ossification spreads from these into the auricular epiphysial plates. One costal epiphysial centre, placed anteriorly, occurs in each remaining interval and from them ossification spreads to the epiphysial plate covering the lower part of the lateral surface of the sacrum. Sacral bodies unite at their adjacent margins after the twentieth year, but the central and greater part of each intervertebral disc remains unossified up to or beyond middle life.
The sacrum may contain six vertebrae, by development of an additional sacral element or by incorporation of the fifth lumbar or first coccygeal vertebrae. Inclusion of the fifth lumbar vertebra (sacralization) is usually incomplete and limited to one side. In the most minor degree of the abnormality a fifth lumbar transverse process is large and articulates, sometimes by a synovial joint, with the sacrum at the posterolateral angle of its base. Reduction of sacral constituents is less common but lumbarization of the first sacral vertebra does occur: it remains partially or completely separate. The bodies of the first two sacral vertebrae may remain unfused when the lateral masses are fused. The dorsal wall of the sacral canal may be variably deficient, due to imperfect development of laminae and spines. Orientation of the superior sacral articular facets displays wide variation, as does the sagittal curvature of the sacrum. Asymmetry (facet tropism) of the superior facets alters the relation between the planes of the two lumbosacral facet joints.
COCCYX
The coccyx (Fig. 42.39) is a small triangular bone and is often asymmetrical in shape. It usually consists of four fused rudimentary vertebrae, although the number varies from three to five, and the first is sometimes separate. The bone is directed downwards and ventrally from the sacral apex: its pelvic surface is tilted upwards and forwards, its dorsum downwards and backwards. Orientation varies with mobility and between individuals.
The base or upper surface of the first coccygeal vertebral body has an oval, articular facet for the sacral apex. Posterolateral to this, two coccygeal cornua project upwards to articulate with sacral cornua: they are homologues of the pedicles and superior articular processes of other vertebrae. A rudimentary transverse process projects superolaterally from each side of the first coccygeal body and may articulate or fuse with the inferolateral sacral angle, completing the fifth sacral foramina.
The second to fourth coccygeal vertebrae diminish in size and are usually mere fused nodules. They represent rudimentary vertebral bodies, though the second may show traces of transverse processes and pedicles.
The gap between the fifth sacral body and the articulating cornua represents, on each side, an intervertebral foramen which transmits the fifth sacral spinal nerve. The dorsal ramus descends behind the rudimentary transverse process, and the ventral ramus passes anterolaterally between the transverse process and sacrum.
Muscle and ligament attachments
The lateral parts of the pelvic surface, including the rudimentary transverse processes, give attachment to the levatores ani and coccygei. The anterior sacrococcygeal ligament is attached to the front of the first and sometimes second coccygeal vertebral bodies (Fig. 80.8). The cornua give attachment to the intercornual ligaments. The lateral sacrococcygeal ligament connects the transverse process to the inferolateral sacral angle. Gluteus maximus is attached to the dorsal surface, and both levator ani and sphincter ani externus are attached to the tip of the bone. The median area gives attachment to the deep and superficial posterior sacrococcygeal ligaments, the superficial descending from the margins of the sacral hiatus and sometimes closing the sacral canal. The filum terminale, which is situated between the two ligaments, blends with them on the dorsum of the first coccygeal vertebra.
Each coccygeal segment is ossified from one primary centre. A centre in the first segment appears about birth and its cornua may soon ossify from separate centres. Remaining segments ossify at wide intervals up to the 20th year or later. Segments slowly unite: union between the first and second is frequently delayed until 30 years. The coccyx often fuses with the sacrum in later decades, especially in females.
LIGAMENTS OF THE VERTEBRAL COLUMN
ANTERIOR LONGITUDINAL LIGAMENT
The anterior longitudinal ligament (Fig. 42.40B) is a strong band extending along the anterior surfaces of the vertebral bodies. It is broader caudally, thicker and narrower in thoracic than in cervical and lumbar regions, and is also relatively thicker and narrower opposite vertebral bodies than at the levels of intervertebral symphyses. It extends from the basilar part of the occipital bone to the anterior tubercle of C1 and the front of the body of C2, then continues caudally to the front of the upper sacrum. Its longitudinal fibres are strongly adherent to the intervertebral discs, hyaline cartilage end-plates and margins of adjacent vertebral bodies, and are loosely attached at intermediate levels of the bodies, where the ligament fills their anterior concavities, flattening the vertebral profile. At these various levels ligamentous fibres blend with the subjacent periosteum, perichondrium and periphery of the anulus fibrosus. The anterior longitudinal ligament has several layers. The most superficial fibres are the longest and extend over three or four vertebrae, the intermediate extend between two or three, and the deepest from one body to the next. Laterally, short fibres connect adjacent vertebrae.
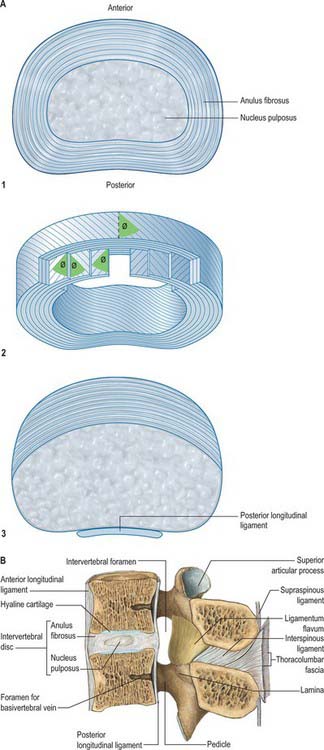
Fig. 42.40 A, The main structural features of an intervertebral disc. For clarity the number of fibrocartilaginous laminae has been greatly reduced. Note alternating obliquity of collagen fascicles in adjacent laminae. 1. Lumbar disc. 2. Detailed structure of anulus fibrosus. ø = approximately 65°. 3. Cervical disc. (After Bogduk.) B, Median sagittal section through upper lumbar vertebral column showing discs and ligaments.
(From Sobotta 2006.)
POSTERIOR LONGITUDINAL LIGAMENT
The posterior longitudinal ligament (Fig. 42.41) lies on the posterior surfaces of the vertebral bodies in the vertebral canal, attached between the body of C2 and the sacrum, and continuous with the membrana tectoria above. Its smooth glistening fibres, attached to intervertebral discs, hyaline cartilage end-plates and adjacent margins of vertebral bodies, are separated between attachments by basivertebral veins and the venous channels which drain them into anterior internal vertebral plexuses. At cervical and upper thoracic levels the ligament is broad and of uniform width, but in lower thoracic and lumbar regions it is denticulated, narrow over vertebral bodies and broad over discs. Its superficial fibres bridge three or four vertebrae, while deeper fibres extend between adjacent vertebrae as perivertebral ligaments, which are close to and, in adults, fused with, the anulus fibrosus of the intervertebral disc. The layers of the posterior longitudinal ligament and the relationship of the ligament to associated membranes in the epidural space are fully discussed by Loughenbury et al (2006).
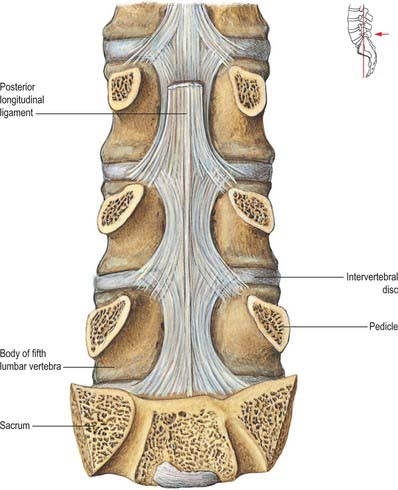
LIGAMENTA FLAVA
The ligamenta flava (Fig. 42.40B, Fig. 42.42) connect laminae of adjacent vertebrae in the vertebral canal. Their attachments extend from facet joint capsules to the point where laminae fuse to form spines. Here their posterior margins meet and are partially united; the intervals between them admit veins which connect the internal and posterior external vertebral venous plexuses. Their predominant tissue is yellow elastic tissue, whose almost perpendicular fibres descend from the lower anterior surface of one lamina to the posterior surface and upper margin of the lamina below. The anterior surface of the ligaments is covered by a fine, continuous smooth lining membrane (Newell 1999). The ligaments are thin, broad and long in the cervical region, thicker in the thoracic and thickest at lumbar levels. They arrest separation of the laminae in spinal flexion, preventing abrupt limitation, and also assist restoration to an erect posture after flexion, perhaps protecting discs from injury.
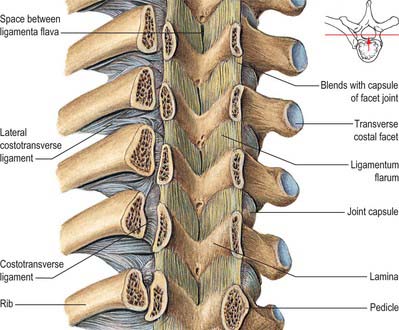
INTERSPINOUS LIGAMENTS
Interspinous ligaments (Fig. 42.40B) connect the facing edges of consecutive spinous processes, and extend ventrally as far as the ligamentum flavum and dorsally to the supraspinous ligament, when this ligament is present (see below). They differ structurally in thoracic, lumbar and cervical levels. The thoracic interspinous ligaments are narrow and elongated, whereas those at lumbar levels are thick and quadrilateral, and occur as closely applied pairs, the left and right ligaments being separated by a narrow or potential cleft. In the lumbar ligaments, collagen fibres run obliquely inferiorly and ventrally and only the deepest fibres are truly ligamentous. The more dorsal fibres are derived from tendons of longissimus thoracis that dip into the interspinous space to gain attachment to the superior edge of the spinous process rather than to its tip. Distinctive interspinous ligaments are not evident at cervical levels, where they are represented by the median septum of the ligamentum nuchae as it passes between the cervical spinous processes.
SUPRASPINOUS LIGAMENT
The supraspinous ligament (Fig. 42.40B) is a strong fibrous cord which connects the tips of spinous process from C7 to L3 or L4. It is regularly deficient. The most superficial fibres extend over three or four vertebrae, the deeper span two or three, and the deepest connect adjacent spines and are continuous with the interspinous ligament. Most of the ligament is formed by the tendons of muscles with posterior midline attachments, i.e. semispinalis, longissimus, trapezius and latissimus dorsi. Only the most superficial fibres lack any connection with muscle. Below L4 the ligament is replaced by the decussating fibres of latissimus dorsi.
INTERTRANSVERSE LIGAMENTS
Intertransverse ligaments run between adjacent transverse processes. At cervical levels they consist of a few, irregular fibres which are largely replaced by intertransverse muscles; in the thoracic region they are cords intimately blended with adjacent muscles; in the lumbar region they are thin and membranous. For the detailed anatomy of the lumbar intertransverse ligaments, see Bogduk (2005).
LIGAMENTOUS INSTABILITY
Damage to the ligaments controlling stability of the column may occur in the absence of evident bony pathology. This is particularly prevalent in inflammatory disease of the upper cervical spine, where rheumatoid arthritis may weaken or destroy the ligaments on which atlanto-axial stability depends. The transverse ligament is stronger than the dens, which therefore usually fractures before the ligament ruptures. The alar ligaments are weaker, and combined head flexion and rotation may avulse one or both alar ligaments: rupture of one side results in an increase of about a third in the range of rotation to the opposite side. Pathological softening of the transverse and adjacent ligaments or of the lateral atlanto-axial joints results in atlanto-axial subluxation, which may cause spinal cord injury. Ligamentous damage may also occur in spinal injuries, particularly at cervical levels.
Developmental laxity of ligaments may also lead to problems with instability, especially if there is an episode of trauma: this combination is probably responsible for atlanto-axial rotational instability. Laxity of cervical spinal ligaments may be a normal variant in children, and lead to diagnostic difficulties. In radiographs of the upper cervical spine in children aged less than 8, a deceptive appearance of subluxation (‘pseudosubluxation’) may result from a combination of ligamentous laxity and facet orientation. This usually occurs between C2 and C3, but may occasionally be seen at C3/4. Clinical and other radiological features should facilitate the correct diagnosis.
JOINTS
All vertebrae from C2 to S1 articulate by secondary cartilaginous joints (symphyses) between their bodies, synovial joints between their articular processes, and fibrous joints between their laminae, transverse and spinous processes. In the cervical region, from C3 to C7, joints have been described between the uncinate or neurocentral processes of the inferior vertebral body and the bevelled lateral border of the superior body at each level. These small unco-vertebral or neurocentral ‘joints’ are absent at birth, do not contain synovium, and are probably clefts in the intervertebral discs.
INTERVERTEBRAL JOINTS
Joints between the vertebral bodies
Joints between vertebral bodies are symphyses. Typical vertebral bodies are united by anterior and posterior longitudinal ligaments and by fibrocartilaginous intervertebral discs between sheets of hyaline cartilage (vertebral end-plates).
Articulating surfaces: intervertebral discs
The intervertebral discs are the chief bonds between the adjacent surfaces of vertebral bodies from C2 to the sacrum. Except at the sites of the uncovertebral (neurocentral) joints of Luschka, disc outlines correspond with the adjacent bodies. Their thickness varies in different regions and within individual discs. Each disc consists of an outer lamellated anulus fibrosus and an inner nucleus pulposus. (Fig. 42.40A1).
In cervical and lumbar regions the discs are thicker anteriorly, contributing to the anterior convexity of the vertebral column. In the thoracic region they are nearly uniform, and the anterior concavity is largely due to the vertebral bodies. Discs are thinnest in the upper thoracic region and thickest in the lumbar region. They adhere to thin layers of cartilage on the superior and inferior vertebral surfaces, the vertebral end-plates. The latter do not reach the periphery of the vertebral bodies but are encircled by ring apophyses. The end-plates contain both hyaline cartilage and fibrocartilage. The fibrocartilaginous component lies nearer to the disc, and is sometimes considered not to be part of the end-plate itself. The fibrocartilaginous components of the end-plates above and below the nucleus pulposus, together with the innermost lamellae of the anulus fibrosus, form a flattened sphere of collagen which surrounds and encloses the nucleus (Fig. 42.43). The overall proportion of fibrocartilage in the end-plate increases with age. While all discs are attached to the anterior and posterior longitudinal ligaments, discs in the thoracic region are additionally tied laterally, by intra-articular ligaments, to the heads of ribs articulating with adjacent vertebrae. Intervertebral discs form about a quarter of the length of the postaxial vertebral column: cervical and lumbar regions make a greater contribution than the thoracic and are thus more pliant.
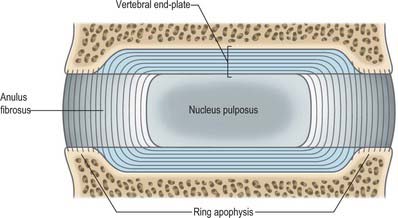
Fig. 42.43 Structure of the vertebral end-plate: the collagen fibres of the inner two-thirds of the anulus fibrosus sweep around into the vertebral end-plate and form its fibrocartilaginous component. The peripheral fibres of the anulus are anchored into the bone of the ring apophysis.
(By permission from Bogduk N 1997 Clinical Anatomy of the Lumbar Spine and Sacrum, 3rd edn. Edinburgh: Churchill Livingstone.)
The anulus fibrosus has a narrow outer collagenous zone and a wider inner fibrocartilaginous zone. Its lamellae, which are convex peripherally when seen in vertical section, are incomplete collars. The internal vertical concavity of the lamellae conforms to the surface profile of the nucleus pulposus. In all quadrants of the anulus, about half the lamellae are incomplete; the proportion increases in the posterolateral region. The exact nature of the interlamellar substance remains in some doubt. Posteriorly, lamellae join in a complex manner. Fibres in the rest of each lamella are parallel and run obliquely between vertebrae at about 65° to the vertical (Fig. 42.40A2). Fibres in successive lamellae cross each other obliquely in opposite directions, thus limiting rotation. The obliquity of fibres in deeper zones varies in different lamellae. Posterior fibres may sometimes be predominantly vertical, which possibly predisposes them to herniation.
This standard description of the anulus may not apply at all spinal levels: a recent cadaveric study indicates that the anulus is usually incomplete posteriorly in adult cervical discs (Mercer & Bogduk 1999) (Fig. 42.40A3).
The nucleus pulposus is better developed in cervical and lumbar regions and lies between the centre of the disc and its posterior surface. At birth it is large, soft, gelatinous and composed of mucoid material. It contains a few multinucleated notochordal cells and is invaded by cells and collagen fibres from the inner zone of the adjacent anulus fibrosus. Notochordal cells disappear in the first decade, and the mucoid material is gradually replaced by fibrocartilage, derived mainly from the anulus fibrosus and the plates of hyaline cartilage adjoining the vertebral bodies. The nucleus pulposus becomes less differentiated from the remainder of the disc as age progresses, and gradually becomes less hydrated and increasingly fibrous. The type II collagen of the nucleus becomes more like the type I of the anulus as its fibril diameter increases. The quantity of aggregated proteoglycans in the nucleus decreases, while the keratan sulphate/chondroitin sulphate ratio increases. As increased cross-linking occurs between collagen and the proteoglycans the discs lose their water-binding capacity, become stiffer and more liable to injury. Contrary to what was previously thought, it has now been shown that lumbar discs do not decrease in overall height as a part of normal ageing. The anulus gradually loses height as its radial bulge increases, but the nucleus retains height and may increase in convexity as it increasingly indents the end-plate. Loss of trunk height with age results from a decrease in vertebral body height (Bogduk 2005). When the disc is not loaded, pressure in the nucleus pulposus is low at all ages.
For a review of the structure and function of the human intervertebral disc see Adams et al (2006).
The ligaments associated with the joints between the vertebral bodies are described on page 728.
Small offshoots of spinal branches of arteries supplying the vertebral column (see p. 714) form an anastomosis on the outer surface of the anulus fibrosus and supply its most peripheral fibres. Normal discs are otherwise avascular and are dependent for their nutrition on diffusion from vertebral bone beneath adjacent end-plates and from the peripheral anulus. Vascular and avascular parts differ in their reaction to injury. Venous drainage is via the external and internal vertebral venous plexuses to the intervertebral veins and thence to the larger named veins which drain the vertebral column. Lymphatic drainage of the vertebral column is briefly considered above (see p. 715). Nothing specific is known about the lymphatic drainage of the disc.
The nerve supply of intervertebral discs has been studied in detail in the lumbar region (Fig. 42.44). The outer third of the anulus is innervated by the sinuvertebral nerves (see p. 754): each sinuvertebral nerve supplies both the disc at the level of its spinal nerve of origin and the disc one level above. The anterior part of the anulus is supplied by the sympathetic (grey ramus communicans) rather than by the mixed nerve. In damaged and degenerate discs the nerves may penetrate more centrally into the disc substance. The sinuvertebral nerves are condensations within extensive nerve plexuses which lie on the posterior longitudinal ligament. Similar plexuses have been demonstrated anteriorly, covering the anterior longitudinal ligament, and laterally in the fetus.
Relations and ‘at risk’ structures
Posterior, lateral and anterior relations of the intervertebral disc are important in the planning of interventional investigative and therapeutic procedures ranging from discography to open disc surgery. The posterolateral surface of the disc forms the anterior boundary of the intervertebral foramen on each side, and so is closely related to the spinal nerve and its accompanying vessels. More centrally the disc is related posteriorly to the dura mater covering the spinal cord and the cauda equina. Anterior relations of the discs vary considerably with vertebral level, but important ‘at risk’ structures include the pharynx and oesophagus, the descending aorta and the inferior vena cava. Laterally, relations change with level, but the parietal pleura in the thorax, and the sympathetic trunk and psoas muscles in the lumbar region, are important examples.
A prolapsed intervertebral disc most commonly affects the 20–55 year age group, and is most often seen at the L4/5 and lumbosacral levels. It may also affect the cervical discs, particularly at C5/6 and C6/7. The thoracic discs are rarely affected. Acute tearing or chronic degeneration of the posterior lamellae of the anulus fibrosus allows deformation and herniation of the disc contents. The disc most often prolapses just lateral to the posterior longitudinal ligament and can compress one or two spinal nerves unilaterally (Fig. 42.45). Much less commonly, the prolapse is central, in the midline posteriorly. The compression of neural structures may then be bilateral, affecting the cord itself or the whole cauda equina. If the damaged anulus ruptures completely, some of the nuclear tissue may escape into the vertebral and ‘root’ canals. This sequestrated material may migrate within the canals and cause nerve compression at spinal levels distant from that of the disc rupture. The disc material itself may have an irritative effect on the spinal nerve.
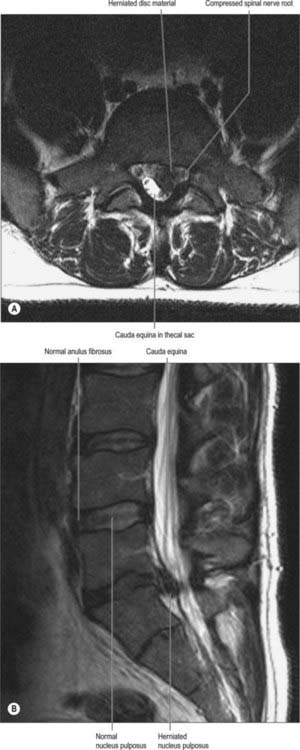
Regarding the anatomy of the vertebral canal and intervertebral foramen in relation to disc prolapse, it is important to understand that one or both of two spinal nerves and their roots may be affected by a single prolapse, depending upon the exact site of the prolapse in the horizontal plane. At the level of each disc and foramen, there are two spinal nerves (and their roots) to consider: these are the exiting nerve and the traversing nerve (Macnab & McCulloch 1990) (Fig. 42.46). The nerve usually affected at lumbar levels is the traversing nerve, which crosses the back of the disc on its way to become the exiting nerve at the level below. Thus a lumbosacral (i.e. L5/S1) disc prolapse usually compresses the S1 nerve. However, a prolapse may affect the exiting nerve at its own level. This is especially likely if the prolapse is in the extraforaminal zone of the ‘root’ canal, the so-called ‘far lateral’ prolapse. At cervical levels, because the roots and nerve leave the vertebral canal almost horizontally, the prolapse usually affects the exiting nerve. This nerve will still bear the number of the vertebra below the affected disc, because cervical nerves exit the canal above the pedicle of their numerically corresponding vertebra. Neurological presentation will include signs and symptoms of spinal nerve damage at the affected level. Thus pain and sensory loss will be dermatomal in distribution. Sensory changes usually precede motor loss.
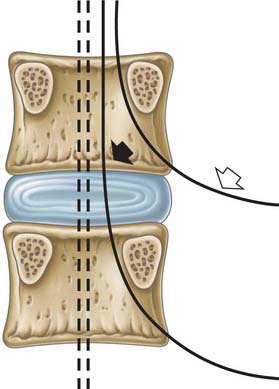
Fig. 42.46 Exiting and traversing nerve roots. The upper root (open arrow) is the exiting root at this level: the lower (closed arrow) is the traversing root here, which becomes the exiting root at the level below. The dotted roots are traversing roots of the lower segment.
Internal disruption of a lumbar intervertebral disc is more common than disc prolapse, and is now an increasingly recognized cause of back pain. Typically, the nucleus is decompressed and the inner lamellae of the anulus appear to collapse into it.
For more detail on disc pathology and its consequences, see Adams et al (2006).
Facet (zygapophysial) joints
Joints between the vertebral articular processes (zygapophyses) are synovial and have long been called zygapophysial joints by anatomists. However, in current clinical practice they are commonly called ‘facet joints’.
Detailed description of the anatomy of these joints is to be found in Bogduk (2005), where Bogduk explains why the term ‘facet joint’ is both incorrect and essentially ambiguous, on the grounds that facets are not restricted to zygapophysial articular processes and occur, for example, in both costovertebral and costotransverse joints.
Facet joints are of the simple (cervical and thoracic) or complex (lumbar) synovial variety: the articulating surfaces are covered in hyaline cartilage and are carried on mutually adapted articular processes. The size and shape of these processes vary with spinal level and are described with the individual vertebrae.
The fibrous capsule is thin and loose and attached peripherally to the articular facets of adjacent articular processes. The capsules are longer and looser in the cervical region. According to Bogduk, the anterior fibrous capsule is replaced entirely by the ligamentum flavum in the lumbar spine.
Bogduk describes two types of intra-articular structure in lumbar facet joints, namely subcapsular fat and ‘meniscoid’ structures. The latter structures may be collagenous, fibroadipose or purely adipose, and project into the crevices between non-congruent articular surfaces. They resemble inclusions seen in the small joints of the hand; their function is conjectural.
Ligaments which work in conjunction with, and modify the function of, the facet joints throughout the vertebral column are described on page 728.
The synovium is attached around the periphery of the articular cartilages and lines the fibrous capsule. In the lumbar region it is reflected over the intracapsular structures described above.
The posterior spinal branches of the arteries that supply the vertebral column form arterial anastomoses around the facet joints. Venous drainage is via the external and internal posterior vertebral venous plexuses to the intervertebral veins and thence to the larger named veins that drain the vertebral column. Lymphatic drainage follows the principles described for the vertebral column (p. 715).
The facet joints are profusely innervated by medial branches of the dorsal primary rami of the spinal nerves, which give articular branches to the joints above and below them.
Relations and ‘at risk’ structures
Anteriorly the capsules of the facet joints form the posterior boundaries of the intervertebral foramina. Posteriorly and laterally the joints are related to the deep muscles of the back, some of whose fibres attach to the capsules. The joints also lie in close relation to the medial branches of the dorsal rami of the spinal nerves and to their accompanying arteries and veins. Damage to the medial branches of the dorsal rami may denervate the deep back muscles. Access to the facet joints and their related nerves may be required in the diagnosis and treatment of spinal pain.
CRANIOVERTEBRAL JOINTS
The articulation between the cranium and vertebral column is specialized to provide a wider range of movement than those which occur in the rest of the axial skeleton. It consists of the occipital condyles, the atlas and the axis, and functions like a universal joint that permits horizontal and vertical scanning movements of the head, and is adapted for eye–head co-ordination.
Atlanto-occipital joints
The atlas articulates with the occipital bone of the skull by a pair of synovial joints. The bones are connected by articular capsules and by the anterior and posterior atlanto-occipital membranes (Fig. 42.47, Fig. 42.48).
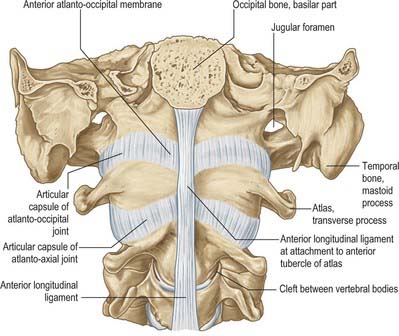
Fig. 42.47 Atlanto-occipital and atlanto-axial joints: anterior aspect. On each side a small cleft has been opened between the lateral part of the upper surface of the body of the third cervical vertebra and the bevelled, inferior surface of the body of the axis.
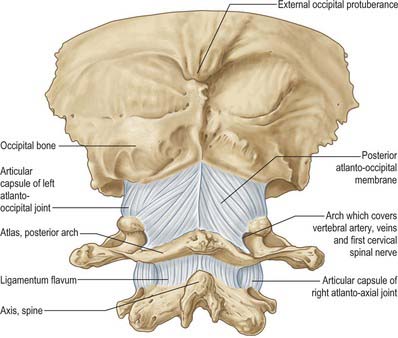
Each joint consists of two reciprocally curved articular surfaces, one on the occipital condyle and the other on the lateral mass of the atlas. The atlantal facets are concave and tilted medially.
The fibrous capsules surround the occipital condyles and superior atlantal articular facets. They are thicker posteriorly and laterally, where the capsule is sometimes deficient, and may communicate with the joint cavity between the dens and the transverse ligament of the atlas.
The ligaments are the anterior and posterior atlanto-occipital membranes.
Anterior atlanto-ocipital membrane
The anterior atlanto-occipital membrane (Fig. 42.47, Fig. 42.49) is a broad, dense fibrous structure which connects the anterior margin of the foramen magnum to the upper border of the anterior arch of the atlas. Laterally, it blends with the joint capsule, and medially it is strengthened by a median cord, which is the anterior longitudinal ligament stretching between the basilar occipital bone and anterior atlantal tubercle.
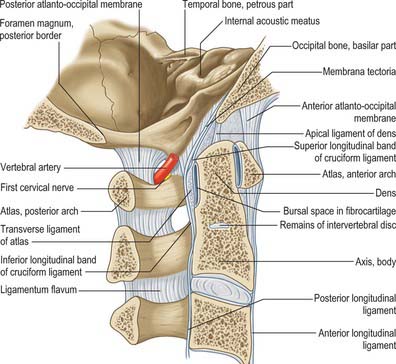
Posterior atlanto-occipital membrane
The posterior atlanto-occipital membrane (Fig. 42.48, Fig. 42.49) is broad, but relatively thin, and connects the posterior margin of the foramen magnum to the upper border of the posterior atlantal arch, blending laterally with the joint capsules. It arches over the grooves for the vertebral arteries, venous plexuses and first cervical nerve, and forms the ‘floor’ of the suboccipital triangle. The ligamentous border of this arch is sometimes ossified. The membrane is usually attached by a soft-tissue ‘bridge’, which may contain muscle or tendon fibres, to rectus capitis posterior minor, and is firmly attached anteriorly to the spinal dura in the same area (Zumpano et al 2006). The ligaments connecting the axis and the occipital bone are functionally involved with the posterior atlanto-occipital membrane.
The synovial cavities of one or both joints may communicate with that of the posterior component of the median atlanto-axial joint.
The arterial supply of this region is derived from an anastomosis between branches of the deep cervical, occipital and vertebral arteries.
The joints are innervated by branches of the ventral primary ramus of the first cervical spinal nerves.
Factors maintaining stability include the fibrous capsules, the atlanto-occipital membranes, the shape of the articular surfaces, the ligaments connecting the axis and the occipital bone, the ligamentum nuchae and the posterior neck muscles. The suboccipital muscles play an important proprioceptive and postural role.
The orientation and shape of the atlanto-occipital joints allow mainly for flexion-extension, which has a total range of about 15°. Axial rotation and lateral flexion have not been measured in living subjects, but a few degrees of motion in these directions can be produced by forceful movement in cadavers.
The following muscles produce these movements. For flexion: longus capitis and rectus capitis anterior; for extension: recti capitis posteriores major and minor, obliquus capitis superior, semispinalis capitis, splenius capitis and trapezius (cervical part); for lateral flexion: rectus capitis lateralis, semispinalis capitis, splenius capitis, sternocleidomastoid and trapezius (cervical part); and for rotation: obliquus capitis superior, rectus capitis posterior minor, splenius capitis and sternocleidomastoid.
Relations and ‘at risk’ structures
Posteriorly the joints are closely related to the vertebral arteries as they pass from the foramina transversaria into the foramen magnum. The dorsal primary ramus of the first cervical nerve and rectus capitis posterior major lie posteromedially and rectus capitis anterior lies anteriorly.
Atlanto-axial joints
The atlas articulates with the axis at three synovial joints. These are a pair between the lateral masses and a median complex between the dens of the axis and the anterior arch and transverse ligament of the atlas.
The articular surfaces of the joints between the lateral masses are often classified as planar. The bony articular surfaces are more complex in shape and are usually reciprocally concave in the coronal plane; the medial parts are somewhat convex in the sagittal plane (especially that of the axis). The cartilaginous articular surfaces are usually less concave. The median joint is a pivot between the dens and a ring formed by the anterior arch and transverse ligament of the atlas. A vertically ovoid facet on the anterior dens articulates with a facet on the posterior aspect of the anterior atlantal arch.
The fibrous capsules for the lateral joints are attached to the articular margins and are thin and loose. Each has a posteromedial accessory ligament attached below to the axial body near the base of its dens, and above to the lateral atlantal mass near the transverse ligament. The fibrous capsule for the median joint is also relatively weak and loose, especially superiorly.
Anteriorly, the vertebral bodies are connected by the anterior longitudinal ligament (Fig. 42.47, Fig. 42.49): here a strong, thickened band attaches above to the lower border of the anterior tubercle of the anterior arch of the atlas and below to the front of the axial body. Posteriorly the vertebral arches are joined by the ligamenta flava (Fig. 42.48, Fig. 42.49) which are attached to the lower border of the atlantal arch above, and to the upper borders of the axial laminae. At this level these ligaments form a thin membrane, pierced laterally by the second cervical nerves.
Transverse atlantal and cruciform ligaments
The transverse atlantal ligament (Fig. 42.50) is a broad, strong band which arches across the atlantal ring behind the dens: its length varies about a mean of 20 mm. It is attached laterally to a small but prominent tubercle on the medial side of each atlantal lateral mass, and broadens medially where it is covered anteriorly by a thin layer of articular cartilage. It consists almost entirely of collagen fibres, which, in the central part of the ligament, cross one another at an angle to form an interlacing mesh. From its upper margin a strong median longitudinal band arises which inserts into the basilar part of the occipital bone between the apical ligament of the dens and membrana tectoria, and from its inferior surface a weaker and less consistent longitudinal band passes to the posterior surface of the axis. These transverse and longitudinal components together constitute the cruciform ligament. The transverse ligament divides the ring of the atlas into unequal parts (Fig. 42.22). The posterior two-thirds surrounds the spinal cord and meninges, the anterior third contains the dens, which the transverse ligament retains in position even when all other ligaments are divided.
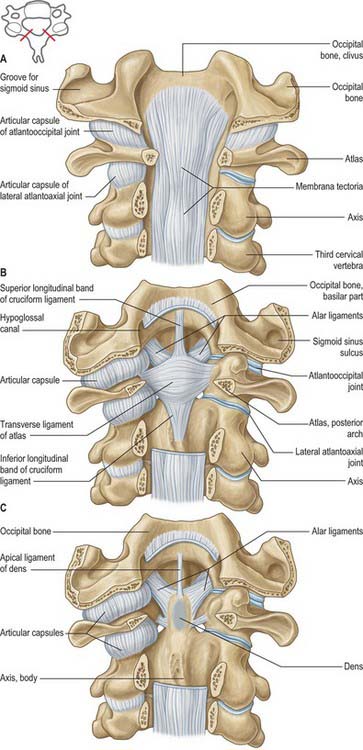
Ligaments connecting axis and occipital bone
Ligaments connecting the axis and the occipital bone consist of the membrana tectoria, the paired alar ligaments, the median apical ligament, and the longitudinal components of the cruciform ligament (Fig. 42.49, Fig. 42.50).
Inside the vertebral canal, the membrana tectoria is a broad strong band representing the upward continuation of the posterior longitudinal ligament. Its superficial and deep laminae are both attached to the posterior surface of the axial body. The superficial lamina expands as it ascends to the upper surface of the basilar occipital bone, and attaches above the foramen magnum, where it blends with the cranial dura mater. The deep lamina consists of a strong median band which ascends to the foramen magnum, and two lateral bands which pass to, and blend with, the capsules of the atlanto-occipital joints as they reach the foramen magnum. The membrane is separated from the cruciform ligament of the atlas by a thin layer of loose areolar tissue, and sometimes by a bursa.
The alar ligaments are thick cords, about 11 mm long, which pass horizontally and laterally from the longitudinally ovoid flattenings on the posterolateral aspect of the apex of the dens to the roughened areas on the medial side of the occipital condyles. In most individuals there is also an anteroinferior band, approximately 3 mm long, which inserts into the lateral mass of the atlas in front of the transverse ligament. Fibres occasionally pass from the dens to the anterior arch of the atlas. In addition, in some 10% of cases a continuous transverse band of fibres, the transverse occipital ligament, passes between the occipital condyles immediately above the transverse ligament. The ligaments consist mainly of collagen fibres arranged in parallel. The main function of the alar ligaments is now considered to be limitation of atlanto-axial rotation, the left becoming taut on rotation to the right and vice versa. The slightly downward movement of the atlas during rotation helps permit a wider range of movement by reducing tension in the alar ligaments, and in the capsules and accessory ligaments of the lateral atlanto-occipital joint.
The apical ligament of the dens fans out from the apex of the dens into the anterior margin of the foramen magnum between the alar ligaments, and represents the cranial continuation of the notochord and its sheath. It is separated for most of its extent from the anterior atlanto-occipital membrane and cruciform ligament by pads of fatty tissue, though it blends with their attachments at the foramen magnum, and with the alar ligaments at the apex of the dens.
The ligamentum nuchae and the anterior longitudinal ligament also connect cervical vertebrae with the cranium.
The synovial membranes of the lateral joints have no special features. The median joint has two synovial cavities which sometimes communicate. The synovial cavity of the posterior component of the median joint complex is larger, lying between the horizontally orientated ovoid facet, on the posterior surface of the dens and the cartilaginous anterior surface of the transverse ligament: communication often exists with one or both of the atlanto-occipital joint cavities.
The arterial supply of this region is derived from an anastomosis between branches of the deep cervical, occipital and vertebral arteries.
The joints are innervated by branches of the ventral primary ramus of the second cervical spinal nerve.
The most important factors maintaining stability are the ligaments, of which the transverse atlantal ligament is the strongest. The alar ligaments are weaker. Other ligaments connecting the axis and the occipital bone, the fibrous capsules, the ligamentum nuchae and the posterior neck muscles also contribute to stability: the suboccipital muscles play an important proprioceptive and postural role.
Movement is simultaneous at all three joints, and consists almost exclusively of rotation around the axis. The shape of the articular surfaces is such that, when rotation occurs, the axis ascends slightly into the atlantal ring, limiting stretch on the lateral atlanto-axial joint capsules. Rotation is limited mainly by the alar ligaments, with a minor contribution from the accessory atlanto-axial ligament. The normal range of atlanto-axial rotation is about 40°.
The muscles that produce atlanto-axial rotation act on the cranium, transverse processes of the atlas and spinous process of the axis. They are mainly obliquus capitis inferior, rectus capitis posterior major and splenius capitis of one side, and the contralateral sternocleidomastoid.
Relations and ‘at risk’ structures
The most important ‘at risk’ relation is the spinal cord, lying posterior to the median atlanto-axial joint. Anteriorly the atlanto-axial articulations, capsules and ligaments are separated from the buccopharyngeal fascia and superior constrictor by longus capitis and longus colli, the prevertebral fascia and the retropharyngeal (potential) space.
Variants of craniovertebral joints
Anomalies of the craniovertebral joints are sometimes seen. Many are minor segmentation anomalies, and may be incidental radiological or osteological findings. More severe anomalies may lead to craniocervical instability (Piper & Traynelis 1998).
LUMBOSACRAL JUNCTION
Articulations between the fifth lumbar and first sacral vertebrae resemble those between other vertebrae. The bodies are united by a symphysis which includes a large intervertebral disc. The latter is deeper anteriorly at the lumbosacral angle. The synovial facet joints are separated by a wider interval than those above.
The reciprocally curved surfaces of the facet joints show considerable individual variation in alignment and shape. Asymmetry (facet tropism) is not unusual.
The major ligament associated with the lumbosacral junction is the iliolumbar ligament (Fig. 80.8, Fig. 42.51).
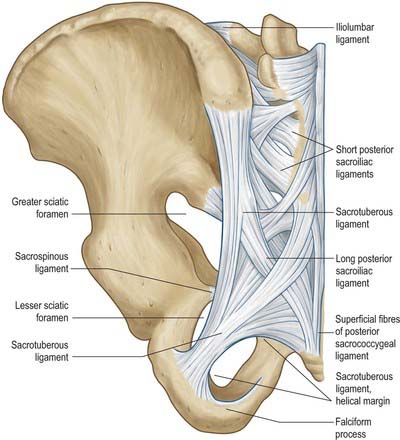
Fig. 42.51 Joints and ligaments on the posterior aspect of the left half of the pelvis and the fifth lumbar vertebra.
The iliolumbar ligament is attached to the tip and anteroinferior aspect of the fifth lumbar transverse process, and sometimes has a weak attachment to the fourth transverse process. It radiates laterally and is attached to the pelvis by two main bands. A lower band passes from the inferior aspect of the fifth lumbar transverse process and the body of the fifth lumbar vertebra across the anterior sacroiliac ligament to reach the posterior margin of the iliac fossa. An upper band, part of the attachment of quadratus lumborum, passes to the iliac crest anterior to the sacroiliac joint, and is continuous above with the anterior layer of the thoracolumbar fascia. The lower ligament has a more vertical component that reaches the posterior iliopectineal line: this component is a lateral relation of the L5 ventral ramus. A posterior component of the iliolumbar ligament passes behind quadratus lumborum to attach to the ilium.
In neonates and children the iliolumbar ‘ligament’ is muscular: the muscle is gradually replaced by ligament up to the fifth decade of life.
The vascular supply of the lumbosacral junction is derived mainly from the iliolumbar and superior lateral sacral arteries.
The lumbosacral junction is innervated by branches derived from the fourth and fifth lumbar spinal nerves: the pattern of distribution is as described on page 707.
Relations and ‘at risk’ structures
The lumbosacral disc is related anteriorly to the common iliac veins, the median sacral vessels, and the superior hypogastric plexus of nerves. The sympathetic trunks cross it anterolaterally, while the obturator nerves and lumbosacral trunks pass close laterally. The relations of the lumbosacral facet joints are similar to those of the lumbar facet joints (see above).
SACROCOCCYGEAL JUNCTION
The sacrococcygeal joint is usually a symphysis between the sacral apex and coccygeal base, united by a thin fibrocartilaginous disc which is somewhat thicker in front and behind than laterally. Its surfaces carry hyaline cartilage which varies from thin veils to small islands. Occasionally the coccyx is more mobile and the joint is synovial.
The anterior sacrococcygeal ligament consists of irregular fibres that descend on the pelvic surfaces of both sacrum and coccyx, and is attached in the same way as the anterior longitudinal ligament. The flat superior posterior sacrococcygeal ligament passes from the margin of the sacral hiatus to the dorsal coccygeal surface, roofing the lower sacral canal (Fig. 42.51). The deep posterior sacrococcygeal ligament passes from the back of the fifth sacral vertebral body to the dorsum of the coccyx and corresponds to the posterior longitudinal ligament. On each side, the lateral sacrococcygeal ligaments connect the coccygeal transverse processes to the inferolateral sacral angles, completing foramina for the fifth sacral spinal nerves. Similarly, the intercornual ligaments connect the sacral and coccygeal cornua, and a fasciculus connects the sacral cornua to the coccygeal transverse processes.
INTERCOCCYGEAL JOINTS
In the young the intercoccygeal joints are symphyses, with thin discs of fibrocartilage between coccygeal segments. Segments are also connected by extensions of the anterior and posterior sacrococcygeal ligaments. In adult males all segments unite comparatively early, but in females union is later. In advanced age the sacrococcygeal joint becomes obliterated. Occasionally the joint between the first and second segments is synovial. The apex of the terminal segment is connected to overlying skin by white fibrous tissue.
MUSCLES
The musculature of the back is arranged in a series of layers, of which only the deeper are true, intrinsic, back muscles (Fig. 42.52). The true back muscles are characterized by their position and by their innervation by branches of the posterior (dorsal) rami of the spinal nerves. The true back muscles below the neck lie deep to the posterior layer of the thoracolumbar fascia. In the lumbar region, where the layers of the thoracolumbar fascia are well-defined, they occupy the compartment between its posterior and middle layers.
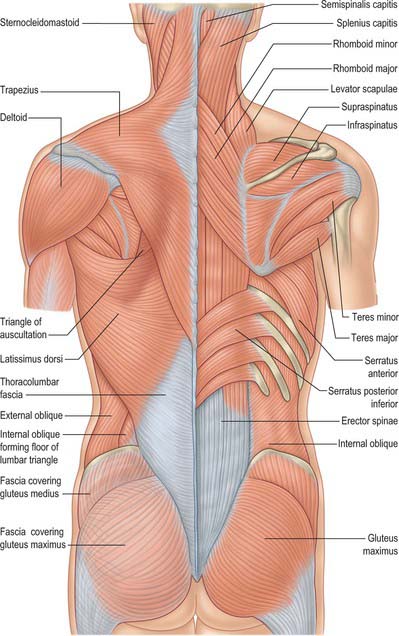
Fig. 42.52 Superficial muscles of the back of the neck and trunk. On the left only the skin, superficial and deep fasciae (other than gluteofemoral) have been removed; on the right, sternocleidomastoid, trapezius, latissimus dorsi, deltoid and external oblique have been dissected away.
Lying superficial to the true, intrinsic muscles are the extrinsic, ‘immigrant’ muscles (Fig. 42.52). The most superficial of these run between the upper limb and the axial skeleton, and consist of trapezius, latissimus dorsi, levator scapulae and the rhomboid muscles. Beneath this layer lie the serratus posterior group, superior and inferior, which are variably developed but usually thin muscles whose function may be respiratory or possibly proprioceptive. All the extrinsic muscles are innervated by ventral rami.
Trapezius, latissimus dorsi, levator scapulae, rhomboid major and rhomboid minor are described on pages 809–811; serratus posterior superior and inferior are described on pages 925–926. The muscles of the posterior abdominal wall are described on page 1070.
The intrinsic muscles are also arranged in layers. The more superficial layers contain the splenius muscles in the neck and upper thorax, and the erector spinae group in the trunk as a whole. The deeper layers include the spinotransverse group, which is itself layered into semi-spinalis, multifidus and the rotatores, and the suboccipital muscles. Deepest of all lie the interspinal and intertransverse muscles. The latter group constitute a mixture of dorsal and ventral spinal muscles. The lumbar intertransversarii mediales, thoracic intertransversarii and medial parts of cervical posterior intertransversarii are innervated by dorsal rami, but the others are supplied by ventral rami.
Muscle attachments and fibre direction
Although developmentally the muscles of the back extend caudally as the vertebral column elongates, drawing their nerve supply with them, they have traditionally been described as if their fibres ran caudocranially, so that the ‘origins’ of the muscles lay distal to their ‘insertions’.
It has been shown (Bogduk 2005) that in the lumbar spine it is more appropriate morphologically and biomechanically to view the muscles in the reverse direction – from above downwards. It is reasonable to argue that this view may also be applied to the cervical and thoracic muscles, and so for uniformity in this section, all muscles have been described in this way.
Ligamentum nuchae
The ligamentum nuchae is not a ligament of the neck, for it does not connect adjacent bones and lacks the internal structure typical of a ligament. It is a unique arrangement of tendons and fascia between the posterior muscles of the neck. It consists of a dorsal raphe and a median septal portion. The dorsal raphe lies superficially along the posterior midline of the neck. It is attached to the external occipital protruberance superiorly, and the tip of the C7 spinous process inferiorly. In its superior half it consists of the aggregated tendons of the most medial fibres of the cervical trapezius. Because of their longitudinal arrangement, these tendons have been described as forming the funicular portion of the ligamentum nuchae (Mercer & Bogduk 2003). In its inferior half the funicular portion is joined by the obliquely orientated tendons of splenius capitis and rhomboid minor. Across the midline the tendons of splenius capitis are continuous with those of rhomboid minor on the other side, and they interweave with the tendons of the reciprocal sets of muscles to produce the raphenous structure of the ligamentum nuchae. From the ventral surface of the dorsal raphe, a median fascial septum extends deeply towards the vertebral column and separates semispinalis capitis from its opposite partner. A lateral expansion of the septum extends laterally ventral to semispinalis capitis, separating it from multifidus and semispinalis cervicis. The median continuation of the septum reaches the tips of the cervical spinous processes, and extends into the cervical interspinous spaces as far as the ligamentum flavum. At upper cervical levels the septum is attached to the external occipital crest and the posterior tubercle of the atlas, and it blends with the posterior atlanto-occipital and posterior atlanto-axial membranes. Passing through these membranes it is attached to the posterior surface of the dura mater (Dean & Mitchell 2002). Only the dorsal raphe affords attachment to muscles: no muscles arise from the median septum.
Splenius capitis
Splenius capitis arises from the mastoid process and the rough surface on the occipital bone just below the lateral third of the superior nuchal line. Its fibres pass downwards and medially to reach the midline. The lower fibres insert into the tips of the spinous processes of the seventh cervical and upper three or four thoracic vertebrae and the intervening supraspinous ligaments. The tendons of the upper fibres interlace in the midline with those of the opposite side in the dorsal raphe of the ligamentum nuchae in the lower half of the cervical region (Fig. 42.53).
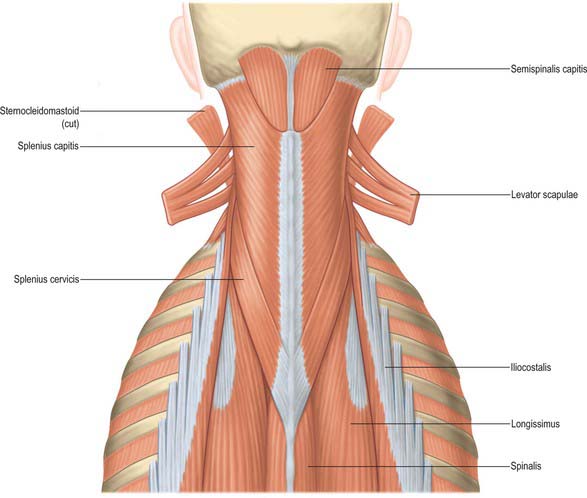
Splenius capitis lies deep to trapezius and covers semispinalis capitis and longissimus capitis. The upper part of the muscle lies beneath sternocleidomastoid. Between sternocleidomastoid and trapezius it forms part of the floor of the posterior triangle of the neck, above and behind levator scapulae.
Splenius cervicis
Splenius cervicis is confluent with splenius capitis, but covers more caudal regions of the neck and thoracic region (Fig. 42.53). It arises from the transverse process of the atlas, the tip of the transverse process of the axis and the posterior tubercle of the third cervical vertebra. Its fibres pass downwards and medially, wrapping around the other posterior intrinsic neck muscles, to insert into the third to sixth thoracic spinous processes.
Splenius cervicis lies deep to serratus posterior superior, the rhomboids and trapezius. It covers the upper parts of the erector spinae and the lower semispinalis muscles.
Erector spinae
Erector spinae (Fig. 42.6, Fig. 42.7, Fig. 42.54, Fig. 42.55) is a large musculotendinous mass which differs in size and composition at different vertebral levels. It consists of fascicles that assume systematic attachments to homologous parts of the skull, the cervical, thoracic, and lumbar vertebrae, the sacrum, and the ilium. Individual muscles are defined by the attachments of their fascicles and the regions that they span. There are three muscles each with three regional parts:
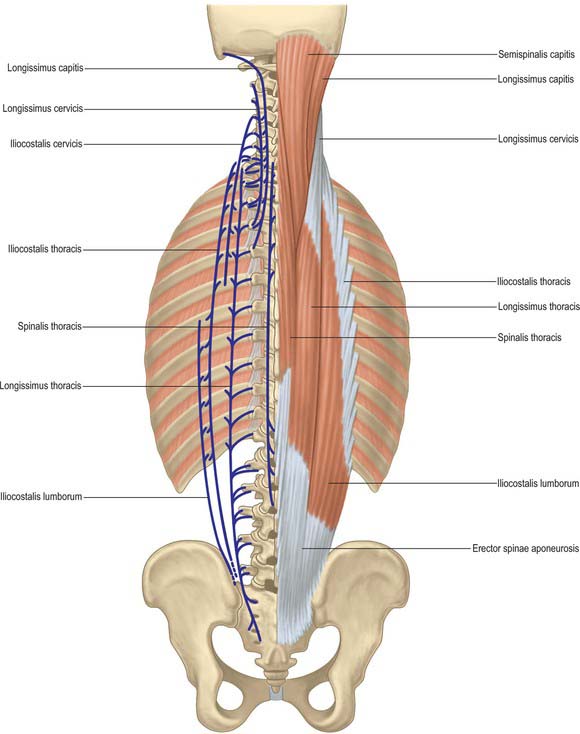
| Spinalis | Longissimus | Iliocostalis |
|---|---|---|
| Spinalis thoracis | Longissimus capitis | Iliocostalis cervicis |
| Spinalis cervicis | Longissimus cervicis | Iliocostalis thoracis |
| Spinalis capitis | Longissimus thoracis | Iliocostalis lumborum |
Spinalis thoracis is the most medial portion of erector spinae in the thoracic region. It consists of fascicles that arise from the spinous processes of the upper thoracic vertebra and insert into the spinous processes of the 11th and 12th thoracic and first two lumbar vertebrae. The number of fascicles and their segmental attachments vary. The fascicles are arranged in an overlapping series of increasingly longer, flat arcs. The shortest fascicles have the lowest origin and highest insertion, and the longest fascicles have the highest origin and lowest insertion. Laterally the muscle blends intimately with longissimus thoracis, and is considered by some to be a component of that muscle.
Spinalis is irregularly and poorly developed in other regions. When present, spinalis cervicis consists of paramedian fibres that arise variously from the spinous processes of the axis and the third and fourth cervical vertebrae, and insert into the lower part of the ligamentum nuchae and the spine of the seventh cervical vertebra (sometimes also to the first and second thoracic vertebrae). Spinalis capitis is represented by occasional fibres of semispinalis capitis that insert into the spines of the seventh cervical and first thoracic vertebrae instead of reaching thoracic transverse processes.
Longissimus is the central component of erector spinae. It assumes a series of attachments at sites that are homologous to the junction of the transverse and costal elements of the segment. These sites are represented at thoracic levels by the tip of a transverse process and the immediately adjacent posterior surface of the rib; at cervical levels by the transverse process and posterior tubercle; and at lumbar levels by the accessory process (the transverse element) and medial half of the transverse process (the costal element).
Longissimus capitis is a narrow flat band of muscle that arises from the posterior edge of the mastoid process, under cover of splenius capitis and sternocleidomastoid. It descends across the lateral surface of semispinalis capitis and inserts by a series of tendons into the transverse processes of the lower three or four cervical and upper four or so thoracic vertebrae.
Longissimus cervicis is a long thin muscle that arises by tendons from the posterior tubercles of the transverse processes of the second to sixth cervical vertebra. It descends into the thoracic region, between the tendons of longissimus capitis and longissimus thoracis, to insert by tendons into the transverse processes of the upper four or five thoracic vertebrae.
Longissimus thoracis is the largest component of erector spinae. It consists of many small fascicles that are aggregated in a particular manner to produce a very long, and in some places thick, muscle. It has lumbar and thoracic parts.
The lumbar part is formed by fleshy bundles that arise from the accessory process and the medial half or so of the posterior surface of the transverse process of each of the five lumbar vertebrae. The fascicles pass inferiorly and slightly laterally. Those from the first four lumbar vertebrae converge onto a common flat tendon that covers the lateral surface of the muscle, and separates it from the lumbar fibres of iliocostalis, for which reason it is called the lumbar intermuscular aponeurosis. The aponeurosis commences in the mid lumbar region, with a broad irregular base, and inferiorly it tapers to a truncated point that inserts into the medial surface of the ilium just dorsal to the ala of the sacrum. The fascicle from the first lumbar vertebra attaches rostrally and dorsally to the aponeurosis. Successive fascicles attach to it more ventrally and caudally. The fascicle from the fifth lumbar vertebra inserts separately, deep to the intermuscular aponeurosis, into the ventromedial aspect of the ilium and the upper fibres of the dorsal sacroiliac ligament. Medially the lumbar fibres of longissimus are separated from the multifidus by a wide cleavage plane filled with fat and veins.
The thoracic part consists of fascicles with small, fusiform muscle bellies that have short rostral tendons and long caudal tendons. The muscle bellies are arranged in a tiered fashion across the length of the posterior thoracic wall, with the highest lying medially and the lowest lying laterally. The upper four fascicles arise from the tips of the first four thoracic transverse processes. The succeeding fascicles have bifid tendons that arise from the transverse process and the adjacent rib at each of the lower eight thoracic segments. The long caudal tendons of the thoracic fascicles of longissimus are aggregated in parallel to form a wide aponeurosis, that allows them to assume a variety of caudal insertions. The tendons of the uppermost fascicles insert into the lumbar spinous processes and their supraspinous ligament. Those from the first thoracic segment reach the L1–2 level, and those from the sixth thoracic segment reach the L5 level. The fascicles from the seventh to ninth thoracic segments reach the median sacral crest, and those from the tenth and 11th thoracic segments attach transversely across the posterior surface of the third segment of the sacrum. The fascicle from the 12th thoracic segment reaches the sacrum and dorsal segment of the iliac crest just below where the intermuscular aponeurosis of the lumbar fibres of longissimus inserts into the ilium. The aponeurosis covers the multifidus and the lumbar fibres of longissimus; it extends from the midline as far laterally as the dorsal edge of the lumbar intermuscular aponeurosis, with which it fuses. Within the lumbar intermuscular aponeurosis the lumbar fibres of longissimus, from L1 to L5, complete the systematic progression of attachments exhibited by the thoracic fibres.
Iliocostalis is the most lateral component of the erector spinae. Its fascicles are attached to sites that are homologous with the ribs.
Iliocostalis cervicis consists of slender fascicles that arise by long tendons from the posterior tubercles of the fourth, fifth, and sixth cervical vertebra. They descend over the posterior thorax to insert into the third to sixth ribs at their angles.
Iliocostalis thoracis is a narrow, fusiform muscle. The fibres arise from the back of the transverse process of the seventh cervical vertebra and the superior borders of the angles of the upper six ribs: they lie lateral to iliocostalis cervicis, and insert into the upper borders of the angles of the lower six ribs.
Iliocostalis lumborum consists of lumbar and thoracic parts. The lumbar part is formed by fleshy fascicles that arise from the tips of the first four lumbar transverse processes and the posterior surface of the middle layer of thoracolumbar fascia lateral to these tips. These fascicles descend to the ilium in a laminated fashion, such that those from higher levels cover those from lower levels. They insert into the medial end of the iliac crest and the dorsal segment of the iliac crest, with the fascicle from L4 assuming the most ventral and lateral attachment, and the fascicle from L1 assuming the most dorsal and medial attachment. The attachment to the ilium is largely by fleshy fibres but the more superficial fibres are aponeurotic. The thoracic part consists of eight or nine small fascicles that respectively arise from the lower eight or nine ribs at their angles, lateral to the iliocostalis thoracis. Each fascicle arises from its rib by a ribbon-like tendon. The tendons are longer at high levels but become progressively shorter at lower levels. The muscle bellies of the fascicles are uniform in length, and each gives rise to a caudal tendon. These tendons are aggregated to form a dorsal aponeurosis that covers the lumbar part of iliocostalis lumborum and inserts in a linear fashion into the medial end of the iliac crest and its dorsal segment. Along this line the fascicles are represented serially, such that the fascicle from the 12th rib attaches most laterally and that from the four or fifth rib attaches most medial and inferiorly.
Together, the dorsal aponeuroses of the thoracic fibres of longissimus and the thoracic fibres of iliocostalis lumborum form a wide sheet of parallel tendons known as the erector spinae aponeurosis. It is attached to the lumbar spinous processes and supraspinous ligaments, the median sacral crest, the third sacral segment, the dorsal segment of the iliac crest, and the medial end of the iliac crest, and covers multifidus, and the lumbar fibres of both longissimus and iliocostalis. Significantly, the erector spinae aponeurosis is formed exclusively by the tendons of the thoracic fibres of longissimus thoracis and iliocostalis lumborum; it does not give rise to the lumbar fibres of these muscles, which are attached independently to the ilium. Some of the more superficial fibres of multifidus may insert into the deep surface of the erector aponeurosis over the sacrum, but otherwise the substantive insertion of multifidus is into the sacrum. A portion of the uppermost fibres of gluteus maximus arise from the dorsal surface of the inferolateral corner of the erector spinae aponeurosis. The lumbar intermuscular aponeurosis is a ventral extension of the erector spinae aponeurosis, separating the lumbar fibres of longissimus from those of iliocostalis.
Erector spinae is covered in the lumbar and thoracic regions by the thoracolumbar fascia, and by serratus posterior inferior below and the rhomboids and splenii above. In the lumbar region it lies in the compartment between the posterior and middle layers of the thoracolumbar fascia.
Erector spinae is innervated by the lateral branches of the dorsal rami of the cervical, thoracic, and lumbar spinal nerves. At lumbar levels, lateral branches innervate iliocostalis and intermediate branches innervate longissimus.
The thoracic and lumbar components of erector spinae are powerful extensors of the vertebral column. Acting concentrically and bilaterally they can extend the thoracic and lumbar spines whereas acting unilaterally they can laterally flex the trunk. However, more commonly, erectores spinae act eccentrically.
From the upright posture, the trunk can flex forwards under the influence of gravity. This movement is initiated by flexor muscles, such as rectus abdominis, in order to bring the centre of gravity of the trunk forwards. Thereafter, erectores spinae control the descent of the thorax under gravity. When the trunk is fully flexed, many parts of erector spinae cease to contract and become electromyographically silent. In this position, flexion is limited by passive tension in the back muscles, and tension in the thoracolumbar fascia, the posterior spinal ligaments, and the intervertebral discs. Similarly, lateral flexion under gravity is controlled by the contralateral erector spinae, with input from the abdominal oblique muscles.
The function of the cervical and capital components of erector spinae has not been determined. These are small muscles with very little force capacity, and are poorly orientated to exercise extension or to control flexion of the head or cervical spine. As putative extensors they are dwarfed by semispinalis and splenius. Axial rotation of the head draws longissimus capitis around the perimeter of the cervical spine, orientating it perhaps so that it is able to restore the head to neutral from the rotated position.
Spinotransverse group
The spinotransverse muscle group consists of muscles where the fascicles span between a spinous process and the transverse elements of vertebrae at various levels below. The muscles are grouped according to the length of their fascicles and the region that they cover.
| Rotatores | Multifidus | Semispinalis |
|---|---|---|
| Rotatores thoracis | Multifidus | Semispinalis cervicis |
| Rotatores cervicis | Semispinalis thoracis | |
| Rotatores capitis | Semispinalis capitis |
Rotatores have the deepest and shortest fascicles, and span one and two segments, whereas the fascicles of multifidus span two, three, four or five segments, and those of semispinalis span about six segments.
Rotatores thoracis (Fig. 42.56) consist of eleven pairs of small quadrilateral muscles. The first pair lies between the first and second thoracic vertebrae, and the last between the 11th and 12th thoracic vertebrae. One or more may be absent from the upper or lower ends of the series. In each segment, a rotator brevis connects the lower border and lateral surface of the lamina above to the upper, posterior part of the transverse process of the vertebra immediately below; a rotator longus connects the base of the spinous process above to the transverse process two levels below. Rotatores are not obviously present at lumbar and cervical levels, where they may be represented by some of the deeper fascicles of multifidus.
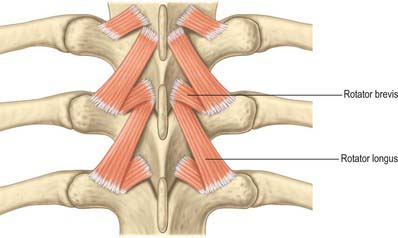
At cervical, thoracic, and upper lumbar levels, multifidus lies lateral to the spinous processes and covers the laminae of the underlying vertebrae, but at lumbosacral levels it expands to cover the posterior surface of the sacrum (Fig. 42.57). At each segmental level, multifidus is formed by several fascicles that arise from the caudal edge of the lateral surface of the spinous process and from the caudal end of its tip. They radiate caudally to insert into the transverse elements of vertebrae two, three, four and five levels below (Macintosh et al 1986). These sites are represented at cervical levels by the superior articular processes, at thoracic levels by the posterior surface of each transverse process near its base, and at lumbar levels by the mammillary processes. Fascicles that extend beyond the fifth lumbar vertebra insert into the dorsal surface of the sacrum. The longest fascicles from the first and second lumbar vertebrae insert into the dorsal segment of the iliac crest. From each spinous process the shortest fascicles pass inferiorly and laterally to their insertion; the longer fascicles assume a progressively steeper course and are arranged progressively more medially. The fascicles from a given segment are flanked and overlapped dorsolaterally by fascicles from successively higher segments, an arrangement that endows the intact muscle with a laminated structure.
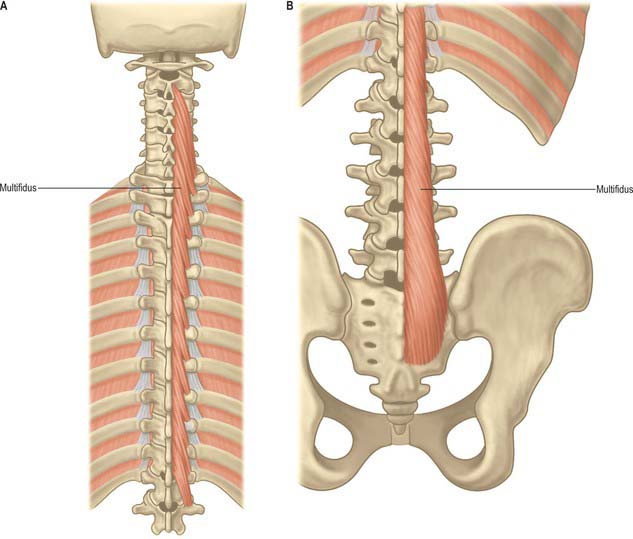
The semispinalis muscles are formed by the longest fascicles of the spinotransverse group. Three divisions are defined essentially by their origins (Fig. 42.58).
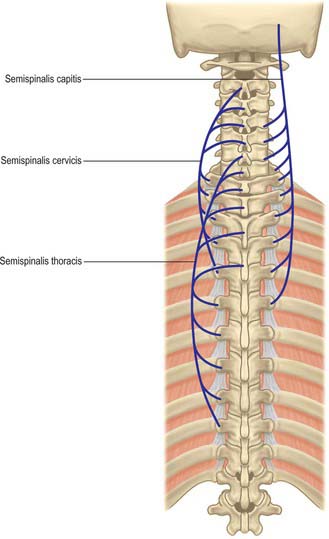
Semispinalis cervicis arises from the spinous processes of the second to fifth cervical vertebrae. Its fascicles span about six segments and cover the cervical and thoracic multifidus. They insert by fleshy or tendinous fibres into the posterior surfaces of the upper five or six thoracic transverse processes. The fascicle from the axis is the largest, and is composed chiefly of muscle.
Semispinalis thoracis consists of thin, fleshy fascicles that have long tendons at both ends. They arise from the lower two cervical and the upper four thoracic spinous processes, and insert into the transverse processes of the sixth to tenth thoracic vertebrae. They cover the thoracic multifidus.
Semispinalis capitis covers semispinalis cervicis. It arises from the medial part of the area between the superior and inferior nuchal lines of the occipital bone and forms a thick muscle bundle in the suboccipital region. Individual fascicles pass inferiorly, laterally and ventrally from the muscle, ending as flat tendons that insert successively into the superior articular processes of the lower four cervical vertebrae and the tips of the transverse processes of the upper six or seven thoracic vertebrae. The medial part of the muscle in the cervical region is formed by the thoracic fascicles, which are more or less distinct from the rest, and has been called biventer cervicis because it is traversed by an incomplete tendinous intersection.
Semispinalis is not formed in the lumbar region, but vestiges of its caudal tendons are represented as the mammillo-accessory ligaments of the lumbar vertebrae.
At cervical, thoracic, and lumbar levels components of erector spinae lie lateral to the spinotransverse group of muscles. They are covered at cervical levels by splenius, at thoracic levels by spinalis thoracis, and at lumbar levels by the erector spinae aponeurosis. In the neck, semispinalis capitis lies mainly deep to splenius and trapezius, but a small portion may be exposed to form the uppermost part of the floor of the posterior triangle of the neck.
Rotatores, multifidus, semispinalis thoracis, and semispinalis cervicis are all innervated by the medial branches of the dorsal rami of the appropriate spinal nerves. Semispinalis capitis is innervated by descending branches of the greater occipital nerve (C2) and the third cervical nerve (C3).
All the spinotransverse muscles are extensors. They extend the vertebrae from which they arise, or the head in the case of semi-spinalis capitis. The predominantly longitudinal orientation of their fascicles precludes any substantive action as rotators. Although rotatores have been presumed to rotate the thoracic vertebrae, this action has not been validated. Axial rotation has been expressly refuted as an action of the lumbar multifidus.
Interspinales
Interspinales are short paired muscular fasciculi attached above and below to the apices of the spines of contiguous vertebrae, one on either side of the interspinous ligament. They are most distinct in the cervical region, where they consist of six pairs, the first between the axis and third vertebra, and the last between the seventh cervical and first thoracic vertebrae. In the thoracic region they occur between the first and second vertebrae (sometimes between the second and third), and the 11th and 12th vertebrae. In the lumbar region there are four pairs between the five lumbar vertebrae. A pair is occasionally found between the last thoracic and first lumbar vertebrae, and another between the fifth lumbar vertebra and the sacrum. Sometimes cervical interspinales span more than two vertebrae.
Intertransversarii
Intertransversarii are small muscles between the transverse processes of the vertebrae. They are best developed in the cervical region, where they consist of posterior and anterior sets of muscles separated by the ventral rami of spinal nerves. Posterior intertransverse muscles are divisible into medial and lateral slips. Each medial slip, the intertransverse muscle ‘proper’, is often further subdivided into medial and lateral parts by the passage through it of the dorsal ramus of a spinal nerve. There are seven pairs of these muscles, the highest between the atlas and axis, and the lowest between the seventh cervical vertebra and the first thoracic vertebra; the anterior muscles between atlas and axis are often absent. In the thoracic region, intertransversarii consist of single muscles that span between the transverse processes of the last three thoracic and the first lumbar vertebrae. In the lumbar region they again consist of two sets of muscles. One set, intertransversarii mediales, connects the accessory process of one vertebra with the mammillary process of the next. The other set, intertransversarii laterales, can be divided into ventral and dorsal parts: the ventral parts connect the transverse processes of the lumbar vertebrae, and the dorsal parts connect the accessory processes to the transverse processes of succeeding vertebrae.
Different parts of the intertransversarii in the cervical and lumbar regions are homologous with certain muscles found in the thoracic region. The homologies are determined according to whether the muscle attaches between consecutive transverse elements of the vertebrae, or between transverse and costal elements, or between consecutive costal elements. They are reinforced by the nerve supply of the muscle, and by the relationship between the muscles and certain nerves.
The medial slips of the ‘proper’ posterior intertransverse muscles of the neck connect consecutive transverse elements, as do the intertransversarii mediales of the lumbar region. These muscles are homologous with the thoracic intertransverse muscles and ligaments. All are supplied by dorsal rami. The lateral slips of the ‘proper’ posterior intertransverse muscles connect a transverse element to a costal element, as do the intertransversarii laterales dorsales. They are homologous with levatores costarum, and are supplied by the ventral rami of spinal nerves. At each level the medial branch of a dorsal ramus separates the true intertransverse muscle from levator costae or its equivalent. The anterior intertransverse muscles of the neck and lateral parts of the posterior intertransversarii connect the costal processes of consecutive vertebrae, as do the intertransversarii laterales ventrales of the lumbar region: they are homologous with the intercostal muscles, and are supplied by ventral rami.
Actions of the short muscles of the back
The functions of interspinales and intertransversarii have not been established. They are very small muscles, and are unlikely to be able to generate enough force to be prime movers of the vertebrae. However, they are richly endowed with muscle spindles: the density approaches that found in the lumbricals of the hand and in the extraocular muscles. It therefore seems reasonable to assume that the short muscles of the back may serve a proprioceptive function in controlling the position of the vertebral column and its movements.
Vascular supply of the true back muscles
The true (intrinsic) muscles of the back receive their blood supply from: the vertebral artery; deep cervical artery; superficial and deep descending branches of the occipital artery; deep branch of the transverse cervical artery (when present); superior intercostal artery, via dorsal branches of the upper two posterior intercostal arteries; posterior intercostal arteries of the lower nine spaces via dorsal branches; dorsal branches of the subcostal arteries; dorsal branches of the lumbar arteries; dorsal branch of arteria lumbalis ima; dorsal branches of the lateral sacral arteries.
The detailed pattern of the arterial supply of the deep muscles of the back has been described by Michel Salmon (Taylor & Razaboni 1994). These muscles are supplied by dorsal branches of the posterior intercostal and lumbar arteries. In the thoracic and upper lumbar regions, where the components of erector spinae run in well-defined longitudinal columns, arterial trunks from these dorsal branches run in the sulci between the columns and between erector spinae and multifidus, giving off branches to supply the muscles. This vascular pattern is less regular in the lumbar region.
Suboccipital muscles
The suboccipital muscles (Fig. 42.59) are four small muscles that connect the occipital bone, atlas and axis posteriorly. They lie inferior to the anterior part of the occipital bone, where three of the muscles form the boundaries of the suboccipital triangle: rectus capitis posterior major lies above and medially; obliquus capitis superior lies above and laterally, and obliquus capitis inferior lies below and laterally. With the head in the anatomical position the suboccipital triangle lies almost in the horizontal plane.
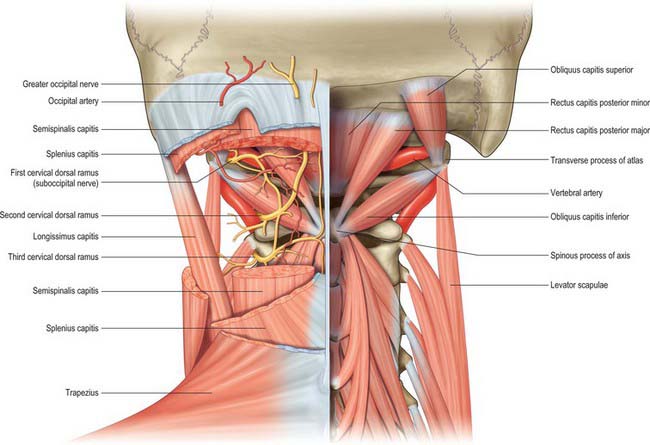
Fig. 42.59 Posterior view of the suboccipital triangles.
(From Drake, Vogl, Mitchell, Tibbitts and Richardson 2008.)
Rectus capitis posterior major
Rectus capitis posterior major is attached by a pointed tendon to the spine of the axis, becomes broader as it ascends, and is attached to the lateral part of the inferior nuchal line and the occipital bone immediately below it. As the muscles of the two sides pass upwards and laterally, they leave between them a triangular space in which parts of recti capitis posteriores minores are visible.
Rectus capitis posterior minor
Rectus capitis posterior minor is attached by a narrow pointed tendon to the tubercle on the posterior arch of the atlas. As it ascends it broadens before attaching to the medial part of the inferior nuchal line and to the occipital bone between the inferior nuchal line and the foramen magnum. Either muscle may be doubled longitudinally. There is usually a soft-tissue attachment to the posterior atlanto-occipital membrane, which itself is firmly attached anteriorly to the spinal dura in the same area (Zumpano et al 2006).
Obliquus capitis inferior, the larger of the two oblique muscles, passes laterally and slightly upwards from the lateral surface of the spine and the adjacent upper part of the lamina of the axis to the inferoposterior aspect of the transverse process of the atlas.
Obliquus capitis superior is attached by tendinous fibres to the upper surface of the transverse process of the atlas. It expands in width as it ascends dorsally, and is attached to the occipital bone between the superior and inferior nuchal lines, lateral to semispinalis capitis and overlapping the insertion of rectus capitis posterior major.
Relations of the suboccipital triangle
Medially the suboccipital triangle is covered by a layer of dense adipose tissue, deep to semispinalis capitis. Laterally it lies under longissimus capitis and sometimes splenius capitis, both of which overlap obliquus capitis superior. The ‘floor’ of the triangle is formed by the posterior atlanto-occipital membrane and the posterior arch of the atlas. The vertebral artery and the dorsal ramus of the first cervical nerve lie in a groove on the upper surface of the posterior arch of the atlas.
The suboccipital muscles receive their blood supply from the vertebral artery and deep descending branches of the occipital artery.
Actions of the suboccipital muscles
The suboccipital muscles are involved in extension of the head at the atlanto-occipital joints and rotation of the head and atlas on the axis. Obliquus capitis superior and the two recti are probably more important as postural muscles than as prime movers, but this is difficult to confirm by direct observation. Rectus capitis posterior major extends the head and, acting with obliquus capitis inferior, rotates the face towards the ipsilateral side. Rectus capitis posterior minor extends the head. Obliquus capitis superior extends the head and laterally flexes it to the ipsilateral side.
MOVEMENTS OF THE VERTEBRAL COLUMN
Spinal movements between individual vertebrae cannot be measured accurately by skin-surface techniques: only biplanar radiography is sufficient for this purpose. The best values for the normal adult lumbar spine are from Pearcy (1984a,b). Movements of the entire lumbar spine can be measured using skin-surface techniques, but the results are of limited clinical use because there is high inter- and intra-observer variation.
The intervertebral discs are the principal sites of vertebral column movement. Conversely, at most levels, the extent to which the discs may be deformed is also the limiting factor for motion. Bony deformation in the subchondral bone and articular cartilage may contribute to movement. Regional variations in mobility of the spine depend on the geometry, orientation and properties of the facet joints and related ligamentous complexes. Physiological intervertebral movements usually combine tilting (bending) and gliding (shear), so the instantaneous centre of rotation moves continually during the movement. During flexion and extension of the lumbar vertebrae, the centre of rotation usually lies near the centre of the intervertebral disc when viewed in the transverse plane, close to the superior end-plate of the vertebra below.
The oblique and ovoid articular surfaces of the facet joints ensure that spinal movements in different planes are usually ‘coupled’ to a certain extent. For example, lateral flexion would cause impingement of the articular surfaces on that side, leading to a posteriorly directed force on the upper vertebra that would act to rotate it about its long axis. Physiologically, coupled movements are variable. Although movements between individual vertebrae are small, their summation gives a large total range to the vertebral column in flexion, extension, lateral flexion, and axial rotation. Each pair of vertebrae with its interposed disc and ligaments is termed a motion segment or functional spinal unit.
In flexion, the anterior longitudinal ligament becomes relaxed as the anterior parts of the intervertebral discs are compressed. At its limit, the posterior longitudinal ligament, ligamenta flava, interspinous and supraspinous ligaments and the posterior fibres of the intervertebral discs are all tensed, the interlaminar intervals widen, the inferior articular processes glide on the superior processes of subjacent vertebrae, and their capsules become taut. Tension of extensor muscles is also important in limiting flexion, e.g. when carrying a load on the shoulders. Flexion is effectively absent in the thoracic region. In forward flexion of the lumbar spine, the muscles normally protect the osteoligamentous spine from injury. However, the margin of safety can be compromised during repetitive or sustained bending by a failure of the spinal reflexes. Once the muscle protection is lost, flexion injury affects first the interspinous ligaments and then the capsules of the facet joints. The ligamentum flavum has such a high content of elastin that it is always under tension, and can be stretched by 80% without damage. This ligament probably functions to provide a constant smooth lining to the vertebral canal which is never overstretched in flexion, and which never goes slack in extension.
In extension, the opposite events occur, and there is compression of posterior discal fibres. Extension is limited by tension of the anterior longitudinal ligament, anterior discal fibres and impaction of spines or articular processes. It is marked in cervical and lumbar regions, and much less at thoracic levels, partly because the discs are thinner, but also because of the presence of the ribs and chest musculature. In full extension, the axis of movement passes posterior to the disc, moves forwards as the column straightens and passes into flexion, and reaches the centre of the intervertebral disc in full flexion.
In lateral flexion, which is always combined with ‘coupled’ axial rotation, intervertebral discs are laterally compressed and contralaterally tensed and lengthened, and motion is limited by tension of antagonist muscles and ligaments. Lateral movements occur in all parts of the column but are greatest in cervical and lumbar regions.
Axial rotation involves twisting of vertebrae relative to each other with accompanying torsional deformation of intervening discs. About 50% of cervical rotation occurs at the atlanto-axial joint. Elsewhere in the column, although movement is slight between individual vertebrae, the range summates to become considerable for the column as a whole. In the post-cervical column, the effective range of rotation is greatest at the thoracolumbar junction. There is very little rotation in the remainder of the lumbar region.
In the cervical region the upward inclination of the superior articular facets allows free flexion and extension. The latter is usually greater, and is checked above by locking of the posterior edges of the superior facets of C1 in the occipital condylar fossae, and below by slipping of the inferior processes of C7 into grooves inferoposterior to the first thoracic superior articular processes. Flexion stops where the cervical convexity is straightened, checked by apposition of the projecting lower lips of vertebral bodies on subjacent bodies. Cervical lateral flexion and rotation are always coupled, and the superomedial inclination of the superior articular facets imparts rotation during lateral flexion.
Cervical motion can be considered to involve the upper (i.e. the atlanto-occipital and atlanto-axial complexes) and the lower cervical spine (C3–7). Two physiological movements take place at the atlanto-occipital joints, those of flexion-extension and lateral flexion. The atlanto-axial joints allow flexion-extension and rotation. Some studies have suggested that maximum flexion-extension occurs between the occiput and C1; however, Frobin et al (2002) noted between 12.6° and 14.5° at this level, which is less than at some of the other cervical levels. Global cervical flexion ranges from 45° to 58° depending on the method of assessment, age and sex: older subjects and females exhibit less motion (Ordway et al 1997; Trott et al 1996). At an intersegmental level, motion increases from the second cervical level, peaks at the mid cervical level (14–17° recorded at C4/5), before reducing at the junction of the cervical and thoracic spine (9.8–11.5° noted at C6/7) (Frobin et al 2002). Global ranges of lateral flexion range from 32° to 47°, with a gradual reduction in range with age and sex, while rotational movements range from 63° to 78°. Intersegmental ranges of motion vary from 4.7° to 6° for lateral flexion between C2 and C7, and 2–12° for rotation (White & Panjabi 1990).
In the thoracic region, especially superiorly, all movements are limited, reducing interference with respiration. Lack of upward inclination of the superior articular facets prohibits much flexion, and extension is checked by contact of the inferior articular margins with the laminae and of adjacent spines. Thoracic rotation is freer, though limited by the ribs at upper levels. Its axis is in the vertebral bodies in the midthoracic region, and in front of them elsewhere, so that rotation involves some lateral displacement. The direction of articular facets would allow free lateral flexion, but this is limited in the upper thoracic region by the resistance of the ribs and sternum. Rotation is usually combined with slight lateral flexion to the same side.
Movement in the thoracic spine is frequently regionalized to upper, mid- and lower thoracic. In the upper thoracic, flexion ranges from 7.8° to 9.5°, increasing to 10–11.4° in the mid-thoracic and 12.5° to 12.8° in the lower thoracic (Willems et al 1996). Extension is more consistent, ranging from 7.1° to 9.7° throughout the thoracic spine. Lateral flexion is greater in the lower thoracic spine, ranging from 5.6° to 6.2° in the upper thoracic, 7.9° to 8.1° in the mid-thoracic and 11.9° to 13.2° in the lower thoracic. However, rotation is greatest in the mid-thoracic region, being 11.8° to 15.9° in the upper thoracic, 21.5° to 25.3° in the mid-thoracic, and 8.3° to 11.8° in the lower thoracic.
Flexion movements are generally greater than extension or lateral flexion in the lumbar region. Axial rotation occurs about a centre of rotation in the posterior anulus, and is limited by bony contact in the facet joints after only 1–2° of movement. Functional transition between thoracic and lumbar regions is usually between the 11th and 12th thoracic vertebrae, where the facet joints usually fit so tightly that slight compression locks them, and prevents all movements but flexion.
During flexion of the lumbar spine there is an unfolding or straightening of the lumbar lordosis. Thus in full flexion the lumbar spine assumes a straight alignment or is curved slightly forwards. Normal ranges of global lumbar flexion range from 58° to 72° in under 40-year-olds and 40° to 60° in the over forties: females exhibit a reduced range compared with males (McGregor et al 1995; Dvorák et al 1995). At an intersegmental level, the L3/4 junction and L4/5 junction exhibit the greatest mobility, approximately 12° and 13° respectively, while at the lowest level (L5/S1) there is only 9°, and at the upper lumbar levels only 8° and 10° respectively (Pearcy et al 1984). Movements into extension are the converse of those seen in flexion. Normal ranges of global extension range from 25° to 30° in under 40-year-olds and 15° to 20° in those over forty. At an intersegmental level, L5/S1 and L1/2 exhibit the greatest mobility at approximately 5°, while the remaining levels exhibit less than 5° of extension. Ranges of lateral flexion and rotation in the lumbar spine are reduced compared to other regions of the spine. Global lateral flexion ranges from 20° to 35°, and rotation ranges from 25° to 40°: the ranges of both movements are reduced with age. Assessment of intersegmental rotation and lateral flexion has proven difficult because of the limitations of measurement techniques.
ROLE OF MUSCULATURE
Although muscles will move the spinal column, the majority of muscular activity is involved in providing stability to maintain posture and to provide a firm platform for limb function. Hence the concept of ‘core stability’ in modern rehabilitation programmes, especially in sports-related problems.
It is important to recognize the way in which the muscles of the back work in conjunction with those of the abdominal wall, particularly the oblique and transversus muscles, and with those of the lower limbs. The erector spinae group and internal oblique and transversus abdominis are anatomically and functionally connected by the thoracolumbar fascia (which encloses the former, and into which the latter are inserted). This fascia, together with collagenous tissue within the back muscles, plays an important role in resisting forward bending of the trunk, and during manual handling. The fascia is tensioned primarily by flexing the trunk, although this tension may be enhanced slightly by the lateral pull of the abdominal muscles. It is functionally advantageous to generate tension in the fascia and muscle sheaths, because the elastic strain energy stored in these stretched tissues can be used to help bring the trunk to an upright position and so reduce the metabolic cost of the movement (Adams et al 2002). The thoracolumbar fascia may also have an important function in transferring load between the trunk and the lower limbs: tension in the fascia can be increased by the actions of gluteus maximus and the hamstrings as well as by trunk flexion.
Muscles producing vertebral movements
The spinal column is moved both directly by muscles attached to it, and indirectly by muscles attached to other bones. Gravity always plays a part: movements with gravity usually involve the eccentric contraction of antagonists. Flexion is effected by longus capitis and longus colli, scaleni, sternocleidomastoid and rectus abdominis of both sides, aided in the lumbar region by the abdominal obliques; extension by the erector spinae complex and the transversospinalis group, splenius, semispinalis capitis and trapezius of both sides, together with the sub-occipital muscles; lateral flexion by ipsilateral longissimus, iliocostalis, abdominal obliques and flexors, and quadratus lumborum; rotation by sternocleidomastoid, splenius cervicis, abdominal obliques, rotatores and multifidus.
FACTORS INVOLVED IN STABILITY
The vertebral column is remarkable in that it combines mobility, stability and load-bearing capacity and also protects its contained neural structures, irrespective of its position. Much of the stability of the vertebral column depends on dynamic muscular control, but there are also bony and ligamentous ‘static’ stabilizers. There is considerable variation between segments of the column in terms of stability and mobility: the most mobile levels are the least stable. The latter are those in which the ratio of intervertebral disc height to vertebral body height is highest. Stability may be compromised by damage to any of these structures.
Trauma may affect any vertebral region. Levels of specialized mobility (e.g. atlanto-axial joint) and the junctions of mobile and relatively fixed regions (e.g. cervicothoracic, thoracolumbar) are particularly vulnerable to severe structural damage, often accompanied by spinal cord and nerve injury. Injuries of the vertebral column may be purely soft-tissue (ligaments, joint capsules and muscles) or may affect bony structures. Pure ligamentous/capsular injuries leading to instability may be particularly difficult to diagnose in the absence of gross radiological signs. In the cervical spine, subluxation and dislocation of the facet joints commonly occur without bony injury because of the orientation of the articular facets.
Chronic infections of many types (e.g. tuberculosis) may involve the vertebrae and lead to their deformity and collapse, affect their mechanical properties and compromise their neuroprotective function. Acute infections, spreading locally or via the bloodstream, may lead to the collection of pus within the vertebral canal causing spinal cord compression (epidural abscess).
The integrity of the vertebrae may also be affected by malignant disease, most commonly metastatic. Vertebrae have a copious blood supply throughout life, and many of the common cancers (e.g. breast, bronchus) spread via the arterial system. Cancers of the haemopoietic system (e.g. multiple myeloma) also commonly affect the vertebrae. Prostatic carcinoma has a predilection to metastasize to the vertebral column, often using the venous (Batson’s plexus) rather than the arterial route. Metastatic deposits may occur within the epidural space, compressing the contents of the dural sac at multiple levels.
Systemic inflammatory diseases may cause both deformity and instability of the vertebral column. Rheumatoid arthritis inflames facet joints and weakens ligaments, leading to instability, especially in the cervical spine. Ankylosing spondylitis and other seronegative arthritides affect joints and ligamentous attachments (entheses), leading to ectopic ossification of collagenous structures, fusion (ankylosis) of interbody and facet joints, and loss of the normal spinal curvatures. Widespread new bone formation at and around the joints of the column occurs in DISH (diffuse idiopathic skeletal hyperostosis). Such conditions would seem to increase stability of the column, at the expense of its mobility and function, but an ankylosed spine is very liable to fracture, with an associated risk of neural damage.
Full stability and load-bearing capacity both require intact vertebral bodies and intervertebral discs. Earlier views regarding the relative importance of the disc-body complex and the posterior elements have proved somewhat simplistic. Clinical observation led to the ‘three-column concept’ of spinal stability (Denis 1983), in which the column is divided into three longitudinal parts rather than two. The anterior column is formed by the anterior longitudinal ligament, the anterior half of the vertebral body and the anterior anulus fibrosus. The middle column is made up of the posterior longitudinal ligament, the posterior half of the vertebral body and pedicles and the posterior anulus fibrosus. The posterior column consists of the neural arch and facet joints and the posterior ligamentous complex. Failure of any column can lead to some instability, the status of the middle column being the most important. The more columns that are affected, the worse is the instability: an injury to two columns is usually unstable.
The elastic deformability of intervertebral discs permits tilting and axial rotation between vertebral bodies, and also helps to reduce vertical accelerations of the head. The main shock-absorbing mechanism of the column stems from the spinal curves, which increase and decrease slightly during locomotion against the restraining tension of the trunk muscles. The elastic strain energy in the stretched tendons of the muscle is responsible for shock absorption.
Both body height and spinal stability are subject to a marked diurnal variation. Body height is affected by changes from recumbency to the upright posture. These diurnal variations appear to be due to changes that occur within the cervical, thoracic and lumbar regions of the spine. Investigations using stereophotogrammetry have demonstrated that 40% of diurnal changes occur in the thoracic spine and affect the degree of kyphosis, and a further 40% in the lumbar spine, but do not affect the lordosis (Wing et al 1992). The greatest change in vertebral column length is found in adolescents and young adults. The height loss occurs within 3 hours of rising in the morning, with an overall loss of about 15 mm.
Although the curvatures within the vertebral column contribute to the changes in height, changes within the intervertebral disc contribute both to observed height loss and to variation in stability. Magnetic resonance imaging investigations reveal a dynamic movement of fluid into and out of an intervertebral disc and adjacent vertebral body over a 24-hour period. Body position affects the movement of fluid. In the early morning, the discs are swollen with water, the intervertebral ligaments and the anulus fibrosus are taut, and the intrinsic bending stiffness and stability of the osteoligamentous spine are relatively high. After several hours of normal activity, the discs lose approximately 20% of their water and height. This change makes the ligaments slack and greatly reduces the bending stiffness of the spine, so that relatively more of the stability of the spine must then be provided by the musculature. The diurnal expulsion of water from intervertebral discs also affects the distribution of compressive loading in the spine. As the day progresses, the hydrostatic pressure in the nucleus pulposus falls, and stress concentrations arise in the anulus fibrosus and facet joints.
All ligaments of the column, as well as the facet joint capsules, are important in the maintenance of stability. The anterior longitudinal ligament is very strong, and resists translational displacement (shear) of the vertebrae as well as extension. All the ligaments of the posterior complex resist flexion and rotation, and their integrity determines the range of movements allowed. These ligaments can support the whole column when the muscles are inactive, e.g. in quiet standing. At the limit of lumbar flexion the column is supported mainly by the thoracolumbar fascia and by collagenous tissue within the electrically silent muscles of the back.
Movements are both determined and constrained by the shape and orientation of the facet joints, whose articular surfaces stabilize the column primarily by resisting horizontal gliding (shear) movements and axial rotation. In the most mobile regions the joint surfaces are flatter and more horizontally placed, as will become apparent if a typical cervical facet joint is compared with a typical lumbar joint.
Certain regions of the vertebral column are further stabilized by additional extraspinal factors. The thoracic spine is stabilized by its position as an integral part of the thoracic cage and by its strong ligamentous linkages with the ribs. The sacrum is effectively a virtually fixed integral element of the bony pelvis.
The contribution to stability conferred by the musculature has been grossly underrated. The whole vertebral column is stabilized by the ‘guy-rope’ or staying effect of the long muscles which attach it to the girdles, the head and the appendicular skeleton. This effect is particularly marked for erector spinae, which controls global posture and movement. The small and deep muscles of the back are best able to resist shear movements between vertebrae because only they have sufficient angulation to the long axis of the vertebral column to do this effectively. The deep muscles can also fine-tune intervertebral movements.
For most back problems in clinical practice, especially chronic low back pain, enhancing muscle strength, stamina, and coordination with the many other muscle groups that contribute to stability, e.g. pelvic girdle muscles, is the most appropriate and effective therapeutic avenue. Only a minority of cases benefit from surgery. Furthermore, neglecting the musculature may explain the relatively high failure rates from surgery.
Mechanical injury to the vertebral column may be produced by flexion, extension, distraction, rotation, shear or compression: movements that cause damage are commonly combined, e.g. flexion, axial rotation and compression.
POSTURE AND ERGONOMICS
Posture is a descriptive term for the relative position of the body segments during rest or activity. The maintenance of good posture is a compromise between minimizing the load on the spine and minimizing the muscle work required.
The well-balanced erect body has a line of gravity which extends from the level of the external auditory meatus, through the dens of the axis just anterior to the body of the second thoracic vertebra, through the centre of the body of the 12th thoracic vertebra, and through the rear of the body of the fifth lumbar vertebra to lie anterior to the sacrum. The position of the line of gravity may move anteriorly with locomotion, and may vary between individuals.
The normal curvature of the cervical spine is a lordosis. However, as a result of pain, injury or poor ergonomics, this curve can become exaggerated to give a ‘protruding chin’ stance, i.e. hyperlordosis in the lower cervical spine.
The thoracic spine is held convex posteriorly, and this posture primarily results from the structure of the underlying vertebrae. However, this curve or kyphosis can become exaggerated to give the impression of a rounded back. Poor posture and ergonomics can lead to this exaggerated curvature but other important causes include tuberculosis, a wedge or compression fracture of a vertebral body, Scheuermann’s osteochondritis, ankylosing spondylitis, osteoporosis, and metastatic carcinoma.
The lumbar spine is held in a lordosis. The degree of this lordosis is determined by the lumbosacral angle and is normally 30–45°. The muscles responsible for this posture include erector spinae, rectus abdominus, the internal and external obliques, psoas major, iliacus, the gluteal and hamstring muscles. The lordosis can be increased (as a result of weak abdominal muscles and tight hamstring muscles), decreased, flattened (common in people either with acute or chronic low back pain), or reversed.
A common postural deviation seen throughout the spine is scoliosis or lateral curvature of the spine. It can be structural, compensatory or protective. In structural scoliosis, the lateral curvature is associated with vertebral rotation, and both the curve and the rotation become more accentuated on forward flexion. Such a scoliosis is common in adolescent girls and its cause is unknown. It may also be secondary to an underlying disorder, e.g. muscular dystrophy, spinal muscular atrophy or spina bifida. A compensatory scoliosis occurs when the pelvis is tilted laterally, e.g. as a result of unequal leg length or of a fixed abduction or adduction deformity at the hip joint. Usually there is no intrinsic abnormality of the spine itself and the scoliosis disappears when the pelvic tilt is corrected. A sciatic or antalgic scoliosis is a temporary deformity produced by the protective action of muscles in certain painful conditions of the spine.
Ergonomics has been defined as ‘the way humans work’, and it permits an appreciation of the effects of tasks and the work environment on underlying postural biomechanics. Nachemson (1975) showed that discs were loaded maximally in sitting and in lifting in a forward leaning position, so sitting posture and lifting have received considerable ergonomic attention.
In sitting the goal has been to determine the seat type and reclining angle associated with lowest disc pressure and the least paraspinal muscle activity. When sitting with the hips and knees flexed to 90° the pelvis rotates posteriorly, flattening the lumbar lordosis and consequently increasing the load on the intervertebral discs. Thus it is now advised that in sitting the angle between trunk and thigh should be between 105 and 135°, with the sacrum tilted at 16° and the fourth and fifth lumbar vertebrae supported.
In lifting heavy weights there is considerable initial compression of lumbar intervertebral discs, and large increases in thoracic and intra-abdominal pressure. The compressive force acting on the spine is shared between the vertebral bodies and the neural arch. In the lumbar spine, the neural arch typically resists 20% of this force once the disc height has been reduced by diurnal fluid expulsion, and when the spine is positioned upright. However, age-related narrowing of the disc can cause more than 50% of the compressive force to be resisted by the neural arch, which may explain why osteoarthritis of the facet joints commonly follows disc degeneration.
When lifting, manual handling advisors emphasize the importance of leg lifting as opposed to back lifting. Loads should also be kept close to the body to reduce the lever arm of the load. The use of deep inspiration to raise intra-abdominal pressure while lifting has also been advised, as this is believed to offer further support to the lumbar spine. The spine is at risk when lifting is combined with twisting, lateral bending, and asymmetric postures. However, heavy lifting remains one of the key work related risk factors for the spine together with whole body vibration, prolonged sitting, twisting and bending.
SURFACE ANATOMY
SKELETAL LANDMARKS
Vertebral spines
In the midline a median furrow runs from the external occipital protuberance above to the natal cleft below (Fig. 42.60, Fig. 42.61, Fig. 42.62). The furrow is most shallow in the lower cervical region and is deepest in the midlumbar zone. Inferiorly, it widens out into a flattened, triangular area, the apex of which lies at the start of the natal cleft and corresponds to the third sacral spine. Palpation of the median furrow reveals the sagittal curves of the spine: the cervical curve is convex forwards (lordosis) and extends from the first cervical to the second thoracic vertebra; the thoracic curvature is concave forwards (kyphosis) and extends from the second to the 12th thoracic vertebra; the lumbar curvature is convex forwards and extends from the 12th thoracic vertebra to the lumbosacral prominence. The external occipital protuberance is subcutaneous and can be felt and often seen (Fig. 42.61), and it can be palpated without difficulty when it is approached from below. The inion is the point situated on this protuberance in the median plane. The tips of the spines of the cervical vertebrae are obscured by the overlying ligamentum nuchae. The tubercle on the posterior arch of the atlas is impalpable; the first bony prominence which is encountered when the finger is drawn downwards in the midline from the external occipital protuberance is the spine of the second cervical vertebra. The ligamentum nuchae terminates inferiorly at the spine of the seventh cervical vertebra, which is the highest, and sometimes the only, visible projection in this region (vertebra prominens). Immediately below this the spine of the first thoracic vertebra is palpable; it is usually more prominent than the seventh cervical vertebra. The spine of the second thoracic vertebra can also often be felt. The third thoracic spinous process is level with the spine of the scapula (Fig. 42.60), and the seventh with the inferior scapular angle when the arm is by the side. The identification of the remaining thoracic spines is not easy, even in a thin subject when the trunk is fully flexed, because of the manner in which they overlap one another in the midthoracic region. In the upper and lower thoracic regions, the tips of the thoracic spines lie opposite the upper part of the body of the immediately subjacent vertebra. In the midthoracic region, they lie opposite the lower part of the vertebra below. The tip of the spine of each lumbar vertebra can usually be palpated without difficulty, especially if the trunk is flexed. Each lies opposite the inferior part of its own body. The body of the fourth lumbar vertebra is level with the summits of the iliac crests, so the fourth lumbar spine overlies the L4/5 interspace (a point useful in lumbar puncture). The second sacral spine is level with the posterior superior iliac spines. Sacral and coccygeal landmarks are described below.
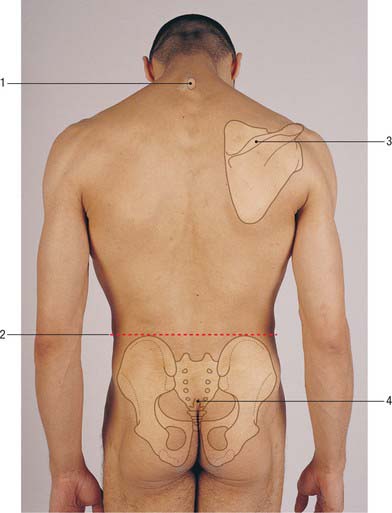
Fig. 42.60 Back view of trunk. 1. Vertebra prominens (spine of C7) 2. Supracristal plane. 3. Spine of scapula. 4. Position of sacral hiatus.
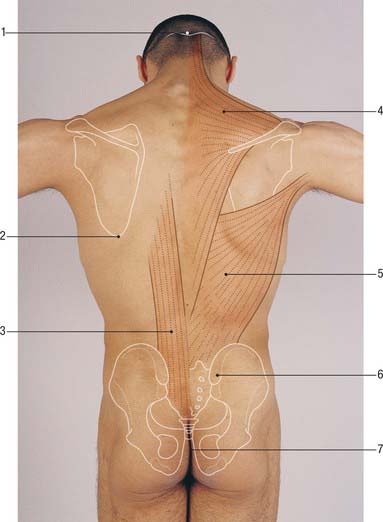
Fig. 42.61 Back view of trunk, arms abducted. 1. External occipital protuberance. 2. Inferior angle of scapula. 3. Erector spinae. 4. Trapezius. 5. Latissimus dorsi. 6. Posterior superior iliac spine underlying sacral dimple. 7. Natal cleft.
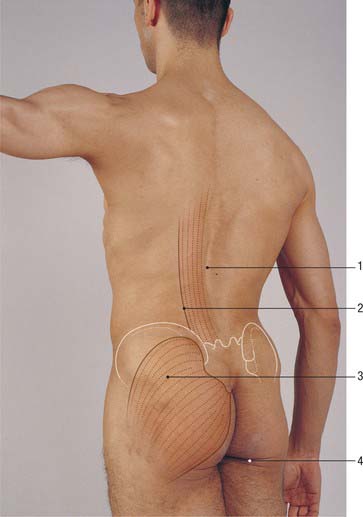
Fig. 42.62 Back of trunk, oblique view. 1. Median furrow. 2. Lateral border of erector spinae. 3. Gluteus maximus. 4. Gluteal fold.
Note that when the subject lies in the lateral position (on the side), the median soft-tissue furrow may not coincide with the median plane, especially in the lumbar region in the obese. Careful palpation may be necessary to identify the spines, a point which is of particular importance during lumbar or epidural puncture in this position.
Spinal levels of viscera
The palpable vertebral spines can be used as landmarks for the levels of the viscera. Some of the more important are shown in Table 42.1 (minor differences between the two sides are ignored).
Table 42.1 Vertebral spines as landmarks for the viscera.
| Spine | Viscera |
|---|---|
| C5 | Cricoid cartilage, start of oesophagus |
| C7 | Apex of lung |
| T3 | Aorta reaches spine. Tracheal bifurcation |
| T4 | Aortic arch ends. Upper border of heart |
| T8 | Lower border of heart. Central tendon of diaphragm |
| T10 | Lower border of lung. Cardia of stomach. Upper border of kidney |
| T12 | Lowest level of pleura. Pylorus |
| L1 | Hilum of kidney. Renal arteries. Superior mesenteric artery |
| L2 | Spinal cord terminates. Pancreas. Duodenojejunal flexure |
| L3 | Lower border of kidney |
| L4 | Bifurcation of aorta |
| L5 | Inferior vena cava begins |
Scapular landmarks
The shape of the back in the upper thoracic region is determined largely by the scapula and the muscles which attach to it, especially trapezius. The relative prominence of the scapula and its muscles depends upon the state of contraction of the latter. Bony scapular landmarks are most evident when the arms hang by the sides.
The scapula overlies the second to seventh ribs. Its superior angle is palpable beneath trapezius, and its inferior angle is just covered by latissimus dorsi: these angles are joined by the medial border of the scapula which runs vertically (Fig. 42.60, Fig. 42.61). The scapular spine runs subcutaneously and is easily palpable from its root medially to the acromion process laterally.
The root of the spine of the scapula lies opposite the third thoracic spine and the inferior angle lies opposite the seventh thoracic spine when the arm is by the side.
Posterior pelvic and sacrococcygeal bony landmarks
At the lower part of the back the iliac crest can be palpated throughout its whole length, and can be traced backwards and upwards from the anterior superior iliac spine to its highest point, and then downwards and medially to the posterior superior iliac spine, which is overlain by a dimple in the skin (Fig. 42.61). A line joining these sacral dimples passes through the second sacral spine and the body of the second sacral vertebra, and is a useful landmark for the lower end of the adult dural sac. There may also be a less prominent pair of dimples at the level of the L4/5 intervertebral disc. The posterior superior iliac spine lies over the centre of the sacroiliac joint. The supracristal plane joins the highest points of the iliac crest on each side. It passes through the body of the fourth lumbar vertebra and has been used as a clinical landmark when performing a lumbar puncture (p. 760) though difficulty in its definition reduces its reliability in this procedure. The tip of the coccyx can be felt deeply near the centre of the natal cleft. As the examining finger passes cranially, the sacral cornua can be felt on either side: these demarcate the sacral hiatus, and form the landmark for performing a caudal anaesthetic block.
MUSCULOTENDINOUS LANDMARKS
In the upper and middle cervical region the median furrow lies between the cylindrical prominences formed mainly by the semispinalis muscles, which are accentuated by neck extension against resistance. In the thoracic and lumbar regions, a broad elevation produced by the erector spinae muscle group extends for about one hand’s breadth on either side of the median furrow and is present between the iliac crest and the 12th rib (Fig. 42.62). The lateral border of this elevation then crosses the ribs at their angles, passing medially as it ascends. The muscle group can be demonstrated by extending the back against resistance.
Trapezius is a flat, triangular muscle that covers the back of the neck and shoulder (Fig. 42.61). Together, the two trapezius muscles resemble a trapezium or quadrilateral in which two of the angles correspond to the shoulders, a third to the occipital protuberance and the fourth to the spine of the 12th thoracic vertebra. The two muscles cover the back of the neck and shoulders like a monk’s cowl, hence the ancient name of the trapezius was musculus cucullaris. The upper part of trapezius is demonstrated by elevation of the shoulders, or by extension and lateral flexion of the neck, against resistance. The lower fibres are best seen when the subject pushes both hands hard against a wall with the elbows extended. The anterosuperior border of the muscle forms the posterior boundary of the posterior triangle of the neck and can be seen in muscular subjects, especially during elevation of the shoulder against resistance.
In a well-muscled subject the outline of latissimus dorsi can easily be traced, particularly if the arm is adducted against resistance. The triangle of auscultation (Fig. 42.52) lies between the upper border of latissimus dorsi, the lower lateral border of trapezius and the medial border of the scapula.
The lumbar triangle (Fig. 42.52), one of the sites of the rare primary lumbar hernia, lies inferiorly, between the lowermost outer border of latissimus dorsi, the posterior free border of external oblique, and the iliac crest.
CLINICAL EXAMINATION
Clinical examination of the back of the trunk and neck best follows the order of inspection, palpation and movement. The examination will be determined by the circumstances of presentation and by the history, and may include musculoskeletal, neurological and vascular observations. Information relevant to the neurological and vascular examination of the skin and material relating to spinal movements and deeper innervation are found above. Palpation of the region involves careful assessment of the bony and musculotendinous landmarks described above, looking in particular for asymmetry, deformity and tenderness. Note that, apart from the spines, most of the bony elements of the vertebrae and almost all of the intervertebral joints are not palpable from behind. In regions of lordosis (sagittal plane curves of the spine with anterior convexity, i.e. midcervical and mid- and lower lumbar), parts of the vertebral column can often be palpated anteriorly with care in well-relaxed, thin subjects.
Adams MA, Bogduk N, Burton K, Dolan P. The Biomechanics of Back Pain, 2nd edn. Edinburgh: Churchill Livingstone, 2006.
A comprehensive and detailed source of information on the functional anatomy, tissue biology and biomechanics of the lumbar spine..
Batson OV. The vertebral vein system. Am J Roentgenol. 1957;78:195-212.
A pioneering study of the venous plexuses of the vertebral column which has become the standard of reference in its field..
Boelderl A, Daniaux H, Kathrein A, Maurer H. Danger of damaging the medial branches of the posterior rami of spinal nerves during a dorsomedian approach to the spine. Clin Anat. 2002;15:77-81.
Detailed descriptions of the vascular supply and innervation of the posterior elements of the thoracolumbar spine and the overlying muscles..
Bogduk N. Clinical Anatomy of the Lumbar Spine and Sacrum, 4th edn. Edinburgh: Elsevier Churchill Livingstone, 2005.
The most thorough text currently available on the topographical and functional anatomy of the lumbosacral spine, with over 1000 references. The book incorporates biomechanical and physiological information which is related to the clinical problem of low back pain..
Crock HV. Atlas of Vascular Anatomy of the Skeleton and Spinal Cord. London: Martin Dunitz, 1996.
Crock HV, Yoshizawa H. The blood supply of the lumbar vertebral column. Clin Orthop. 1976;115:6-21.
Dean NA, Mitchell BS. Anatomic relation between the nuchal ligament (ligamentum nuchae) and the spinal dura mater in the craniocervical region. Clin Anat. 2002;15:182-185.
Describes continuity in the midline between the spinal dura and the ligamentum nuchae in human cadavers..
Denis F. The three column spine and its significance in the classification of acute thoracolumbar spinal injuries. Spine. 1983;8:817-831.
Seminal paper for the understanding and classification of spinal instability..
Dvorák J, Vajda EG, Grob D, Panjabi MM. Normal motion of the lumbar spine related to age and gender. Eur Spine J. 1995;4:18-23.
Frobin W, Leivseth G, Biggeman M, Brinckmann P. Sagittal plane segmental motion of the cervical spine. A new precision measurement protocol and normal motion data of healthy adults. Clin Biomech. 2002;17:21-31.
Groen G, Baljet B, Drukker J. The nerves and nerve plexuses of the human vertebral column. Amer J Anat. 1990;188:282-296.
An acetylcholinesterase whole-mount study of human fetal material giving detail of the perivertebral nerve plexuses and of the sinuvertebral nerves..
Lang J. Craniocervical region, osteology and articulations. Neuro Orthop. 1986;1:67-92.
Loughenbury PR, Wadhwani S, Soames RW. The posterior longitudinal ligament and peridural (epidural) membrane. Clin Anat. 2006;19:487-492.
Macintosh JE, Valencia F, Bogduk N, Munro RR. The morphology of the lumbar multifidus muscles. Clin Biomech. 1986;1:196-204.
MacLaughlin SM, Oldale KNM. Vertebral body diameters and sex prediction. Ann Hum Biol. 1992;19:285-293.
Describes the archaeological and forensic examination of skeletal material..
Macnab I, McCulloch J. Backache, 2nd edn. Baltimore: Williams and Wilkins, 1990. Chapter 1.
The functional anatomy of the lumbar spine, described as a basis for the clinical management of low back pain..
McGregor AH, McCarthy ID, Hughes SPF. Motion characteristics of the lumbar spine in the normal population. Spine. 1995;20(22):2421-2428.
Mercer S, Bogduk N. The ligaments and anulus fibrosus of human adult cervical intervertebral discs. Spine. 1999;24:619-628.
A human cadaveric microdissection study showing that the cervical anulus fibrosus is an anterior crescent rather than a uniformly circumferential structure..
Mercer SR, Bogduk N. Clinical anatomy of ligamentum nuchae. Clin Anat. 2003;16:484-493.
Nachemson A. Towards a better understanding of low-back pain: a review of the mechanics of the lumbar disc. Rheumatol Rehab. 1975;14:129-143.
Newell RLM. The spinal epidural space. Clin Anat. 1999;12:375-379.
Review of the morphological, developmental and topographical aspects of the spinal epidural space..
Ordway NR, Seymour R, Donelson RG, Hojnowski L, Lee E, Edwards T. Cervical sagittal range of motion using three methods. Spine. 1997;22:501-508.
Pearcy M, Protek I, Shepherd J. Three-dimensional X-ray analysis of normal movement in the lumbar spine. Spine. 1984;9:294-297.
Pearcy M, Tibrewal SB. Axial rotation and lateral bending in the normal lumbar spine. Spine. 1984;9:582-587.
Piper JG, Traynelis VC. Congenital malformations of the craniovertebral junction. In: Dickman CA, Spetzler RF, Sonntag VKH. Surgery of the Craniovertebral Junction. New York: Thieme, 1998.
Taylor GI, Razaboni RM. Michel Salmon: Anatomic Studies, Book 1. Arteries of the Muscles of the Extremities and Trunk. St Louis: Quality Medical Publishing, 1994.
A translated, updated and edited version of a classic French text first published in 1933. Now a major source-book in plastic surgery..
Taylor JR, Twomey LT. Sexual dimorphism in human vertebral shape. J Anat. 1984;138:281-286.
Trott PH, Pearcy MJ, Ruston SA, Fulton I, Brien C. Three-dimensional analysis of active cervical motion: the effect of age and gender. Clin Biomech. 1996;11:201-206.
Twomey L, Taylor J, Furniss B. Age changes in the bone density and structure of the lumbar vertebral column. J Anat. 1983;136:15-25.
White AA, Panjabi MM. Clinical Biomechanics of the Spine, 2nd edn. Philadelphia: JB Lippincott, 1990.
Willems JM, Jull GA, Ng JK-F. An in vivo study of the primary and coupled rotations of the thoracic spine. Clin Biomech. 1996;11:311-316.
Wing P, Tsang I, Gagnon F, Susak L, Gagnon R. Diurnal changes in the profile shape and range of motion of the back. Spine. 1992;17:761-765.
Zumpano MP, Hartwell S, Jagos CS. Soft tissue connection between rectus capitis posterior minor and the posterior atlanto-occipital membrane. Clin Anat. 2006;19:522-527.