Chapter 11 Reproductive system
Reproduction is the means by which a species is able to perpetuate itself. If animals lived for ever, there would be no need for another generation to take over from previous ones; in reality, all animals become old or ‘worn out’ and die and must be replaced if the species is not to become extinct.
All species of mammal have evolved separate sexes and they reproduce sexually. This is in contrast to less highly evolved species, which may reproduce asexually – producing offspring that are identical to the parent. Sexual reproduction involves the transfer of genetic material. After mating specialised germ cells – spermatozoa from the male and ova from the female – fuse to form a single-celled zygote. The zygote undergoes cell division to form the embryo. The offspring resulting from sexual reproduction are genetically different from each other and from their parents.
The reproductive system shares part of its structure with the urinary system and the combined systems may be referred to as the urogenital system.
Male reproductive system
The male dog is known as a dog; the male cat is known as a tomcat. The reproductive system of the dog and the tomcat are generally similar – any differences will be described where appropriate (Figs 11.1, 11.2). The parts of the male reproductive tract are:
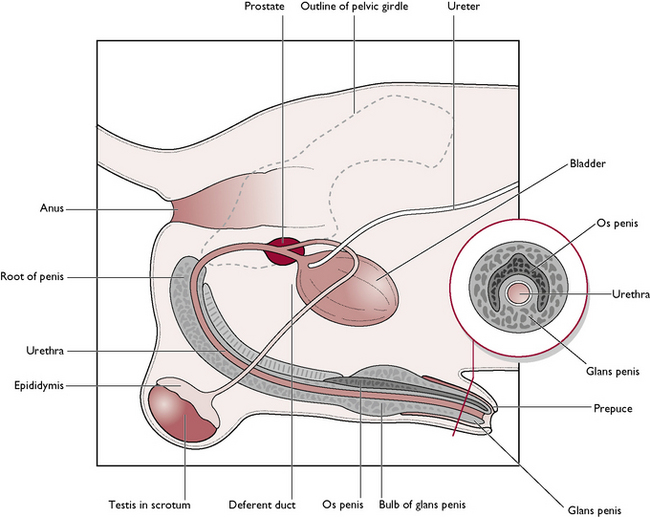
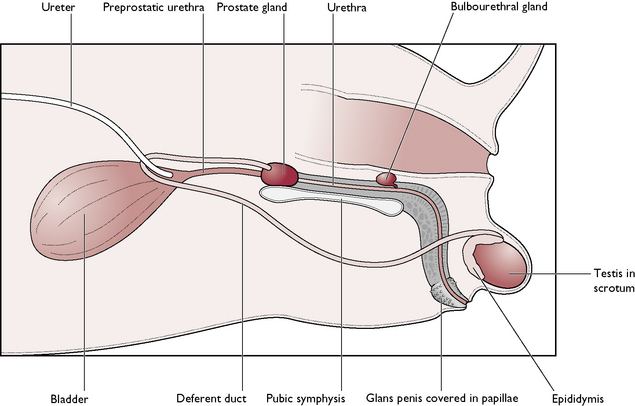
The testis
The testis is the male gonad, with functions as follows:
There is a pair of testes, which, in the adult animal, lie outside the body cavity in the scrotum. – a sac of relatively hairless and often pigmented skin. Spermatogenesis occurs most efficiently at temperatures below that of the core body temperature, so the testes are carried outside the body cavity in a cooler environment. In the dog, the scrotum lies between the hind limbs; in the cat it is attached to the perineum, ventral to the anus. Internally, the sac is divided into two, each part containing one testis; the left testis often hangs lower than the right one. Within the wall of the scrotum is the dartos muscle. In cold weather this contracts and thickens the scrotal skin, raising the temperature; in warm weather, the muscle relaxes and the scrotum becomes thinner and thus cooler. A constant temperature for spermatogenesis is therefore maintained.
Each testis is an oval-shaped structure wrapped in a double layer of peritoneum known as the tunica vaginalis (Fig. 11.3). The testicular tissue consists of numerous blind-ending tubules known as seminiferous tubules which are lined by two types of cells:
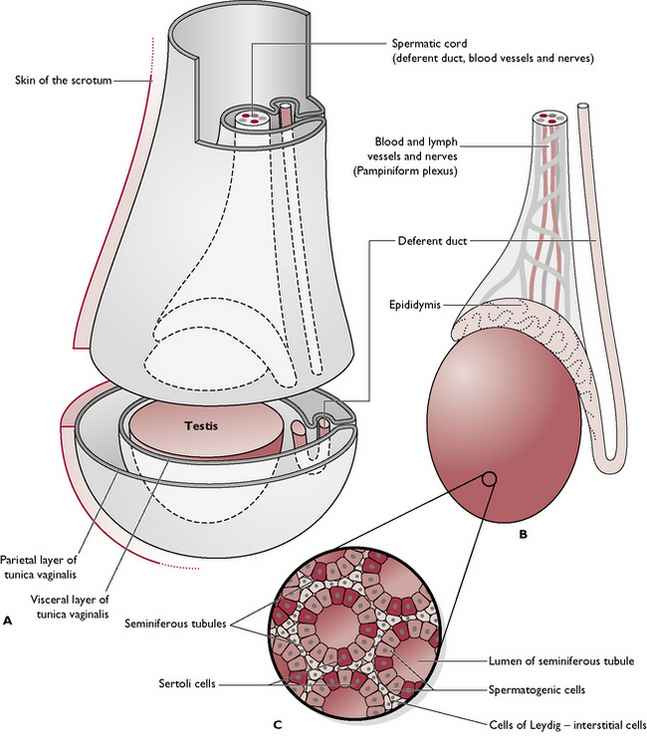
Fig. 11.3 A Testis within scrotum. B Scrotum removed. C Cross-section through the seminiferous tubules.
Lying between the tubules are the cells of Leydig or interstitial cells. They secrete testosterone and are under the control of interstitial cell stimulating hormone (ICSH) (see Ch. 6), produced by the anterior pituitary gland.
The coiled seminiferous tubules make up most of the testicular tissue and eventually combine to form slightly larger efferent ducts. These drain into the epididymis, lying along the dorsolateral border of the testis. The cauda epididymis or tail is attached to the caudal extremity of the testis and is the point at which the temperature of the testis is lowest. It is here that sperm are stored and undergo a period of maturation ready for fertilisation (Fig. 11.4).
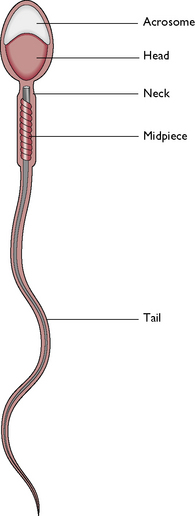
Fig. 11.4 A normal spermatozoon. The acrosome protects the head of the sperm and contains enzymes which aid penetration of the ovum. The head contains the haploid number of chromosomes. The midpiece contains enzymes and mitochondria to provide energy for movement. The tail produces a powerful propulsive force.
The blood supply to the testis is via the testicular artery. This leaves the aorta in the abdomen, just caudal to the renal artery. As it enters the scrotum, the testicular artery runs alongside the epididymis and then divides to form the convoluted pampiniform plexus. This complicated capillary network ensures that the blood is cooled before it enters the testicular tissue.
Testicular descent
In the early embryo, the undifferentiated gonads develop inside the abdomen close to the kidney. In the male, the gonad becomes the testis and a band of tissue known as the gubernaculum forms and runs from the caudal end of each testis to the inside of the developing scrotal sac. During late gestation, the testes are pulled caudally by the contraction of the gubernaculum and they migrate through the abdomen. The testes leave the abdominal cavity viathe inguinal canal – a channel between the fibres of the external abdominal oblique muscle in the groin or inguinal area (see Ch. 4). As each testis with its associated blood capillaries, nerve and deferent duct passes through the inguinal canal into the scrotum, it becomes wrapped in a double fold of peritoneum which forms the tunica vaginalis (Fig. 11.3).
The testes begin their descent into the scrotum during early neonatal life and should be palpable within the scrotum by 12 weeks of age in the puppy and 10–12 weeks in the kitten. Failure of the testes to descend is described as cryptorchidism; the testes may be retained in the abdomen or within the inguinal canal.
Deferent duct
The epididymis continues as the deferent duct (also called the vas deferens or ductus deferens; Figs 11.1, 11.2), which passes out of the scrotum into the abdominal cavity via the inguinal canal within the spermatic cord. The spermatic cord is wrapped in the tunica vaginalis and also contains the testicular artery and vein and the testicular nerve. Lying within the cord is a strip of muscle derived from the internal abdominal oblique muscle and known as the cremaster muscle. Contraction of this muscle raises the testis closer to the body in response to cold and works in conjunction with the Dartos muscle to maintain a constant temperature for the testes.
During ejaculation, the sperm and fluid produced in the seminiferous tubules are propelled along the epididymis and up the deferent duct, which joins the urethra. At this junction, the walls of the deferent ducts are thickened and glandular; the whole area is surrounded by the prostate gland (Figs 11.1, 11.2).
The penis
The functions of the penis are to:
The urethra runs through the centre of the penis and extends from the bladder to the tip of the penis. It is shared by both the reproductive and urinary systems (see Ch. 10). The penis of the dog and the cat are anatomically different.
The dog
The penis runs from the ischial arch of the pelvis, passes cranioventrally along the perineum and between the hind limbs (Fig. 11.5). The urethra lies in the centre and is surrounded by a layer of cavernous erectile tissue known as the corpus spongiosum penis. This is expanded proximally into the bulb of the penis and, towards the tip, as the glans penis. Surrounding this and serving to attach the penis to the ischial arch is a pair of erectile tissue crura (sing. crus), known as the corpus cavernosum penis. These form the root of the penis at its attachment to the ischial arch. The urethra runs in a groove between the two crura.
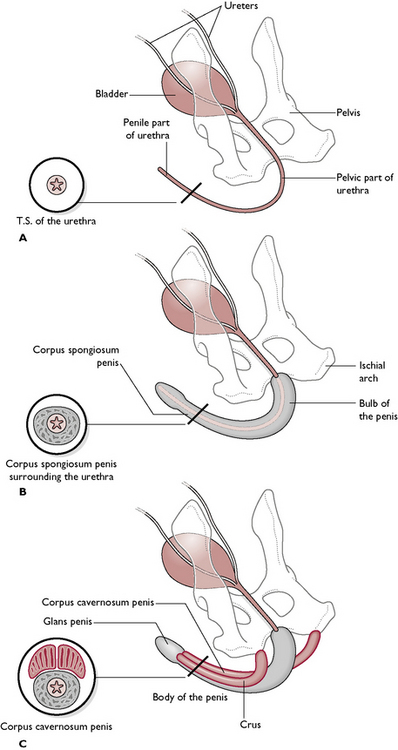
Fig. 11.5 Structure of the penis in three layers, with transverse sections through the urethra. A Shows the urethra only. B Shows the urethra with the layer of cavernous erectile tissue surrounding it: the corpus spongiosum penis. C Shows the corpus cavernosum penis, which forms the two crura and attaches the penis to the ischial arch.
Cavernous erectile tissue is made of connective tissue perforated by ‘caverns’ lined by endothelium. During sexual excitement these caverns fill with blood under pressure and the tissue becomes engorged and erect.
Within the tissue of the glans penis is a tunnel-shaped bone, the os penis, whose function is to aid entry of the penis into the vagina of the bitch during the early stages of mating when erection is only partially complete. The canine os penis lies dorsal to the urethra, which runs through the ‘tunnel’ in the bone. At this point the urethra cannot expand and this can be a common site for urethral blockage with urinary calculi.
Many dogs develop an infection of the lubricating glands of the prepuce. This is known as balanitis or balanoposthitis and it results in a greenish-yellow discharge. The condition is so common that it is considered almost normal and is usually only treated if the mucous membrane becomes ulcerated or smelly.
The distal part of the penis is contained within a sheath of hairy skin known as the prepuce. This is suspended from the ventral abdominal wall and covers and protects the penis. It is lined with mucous membrane and is well supplied with lubricating glands. During mating, the prepuce is pushed back to reveal the glans penis. Afterwards, the retractor penis muscle pulls the penis back into the prepuce.
The tomcat
The main parts of the penis are similar to those of the dog, except that the cat penis is shorter and points backwards – the external opening is ventral to the anus (Fig. 11.2). The glans penis is covered with tiny barbs, which elicit a pain reflex as the male withdraws from the female after mating. This stimulates the nerve pathway to the hypothalamus, resulting in ovulation approximately 36 hours later – known as induced ovulation. The os penis lies ventral to the urethra in the cat. During sexual excitement, the penis engorges and points cranioventrally so that the mating position in cats is similar to that seen in the dog.
Accessory glands
The function of the accessory glands is to secrete seminal fluids which:
There are two types of gland (Figs 11.1, 11.2):
Female reproductive system
The female dog is known as a bitch; the female cat is known as a queen. The parts of the female reproductive system are:
The reproductive tract of the bitch and queen are similar and vary only in size (Fig. 11.6). The tract is designed to carry several fetuses during a single pregnancy and is said to be bicornuate (two horns). The bitch and the queen bear litters of young: they are multiparous.
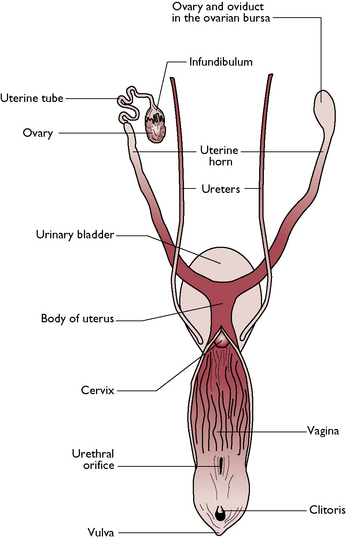
Fig. 11.6 Dorsal view of the reproductive system of the bitch.
(With permission from Colville T, Bassett JM 2001 Clinical anatomy and physiology for veterinary technicians. Mosby, St Louis, MO, p 330.)
The ovary
The ovary is the female gonad. The functions of the ovary are:
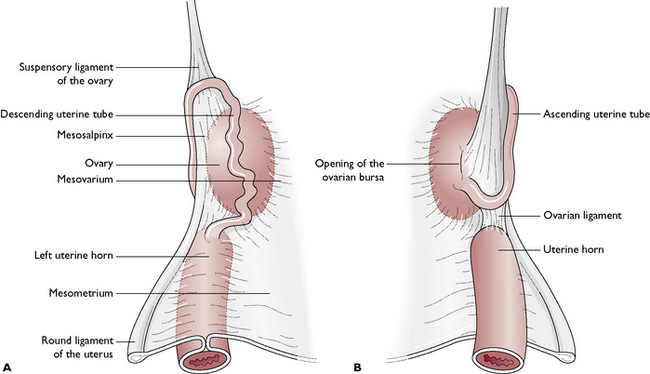
There is a pair of ovaries, one lying on each side of the dorsal abdominal cavity, caudal to the kidney (see Ch. 10, Fig. 10.2). The ovary is held close to the kidney by the ovarian (suspensory) ligament (Fig. 11.7). The ovary is suspended from the dorsal body wall by part of the visceral peritoneum called the mesovarium, which also encloses the infundibulum of the uterine tube. Part of the mesovarium forms a pocket-like structure known as the ovarian bursa, which completely covers the ovary. Within this is a small opening allowing ova to leave the ovary – this is a potential means of entry of infection into the peritoneal cavity.
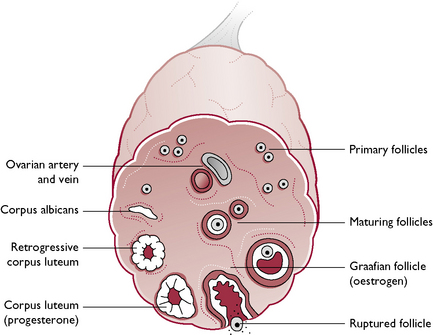
The tissue of the ovary consists of a framework of connective tissue, smooth muscle and blood capillaries, within which are a large number of germ cells and developing follicles (Fig. 11.8). In an immature animal, each ovary is oval with a smooth outline but, as sexual maturity approaches, the ovary becomes nodular as the follicles enlarge.
The uterine tube
This is also known as the oviduct or Fallopian tube (Fig. 11.6). The functions of the uterine tubes are:
Each uterine tube is a narrow convoluted structure lying close to the ovary (Fig. 11.7). The open end is funnel-shaped and known as the infundibulum. It is fringed with finger-like processes known as fimbriae, which spread over the surface of the ovary to capture ova as they are released. The ova pass down the lumen of the tube, which is lined with ciliated columnar epithelium. The cilia propel the ova along the tube towards the uterine horns. The uterine tube is suspended by part of the visceral peritoneum known as the mesosalpinx.
The uterus
The uterus is a Y-shaped structure lying in the midline of the dorsal abdomen (Fig. 11.6). During pregnancy, the weight of the conceptuses pulls the uterus ventrally and at full term it occupies the greater part of the abdomen. The function of the uterus is:
The uterus consists of two parts. A pair of uterine horns lead from the uterine tubes. Each horn is about five times the length of the uterine body and, during pregnancy, contains the developing embryos. The two horns join to form a short central body.
The cervix
The cervix is a short, thick-walled muscular sphincter that connects the uterine body with the vagina (Fig. 11.6). Running through the centre is a narrow cervical canal, which is normally tightly closed and relaxes only to allow the passage of sperm or fetuses. During pregnancy, the canal is blocked by a mucoid plug, which protects the conceptuses from infection. In the non-pregnant animal, the cervix lies in the pelvic cavity but during pregnancy the weight of the conceptuses pulls the cervix cranially and ventrally over the edge of the pelvic brim.
Ovariohysterectomy, more commonly called ‘spaying’, is performed by the veterinary surgeon to prevent unwanted pregnancies and oestrous cycles. The surgical procedure involves the complete removal of the reproductive tract from the ovaries to a point just cranial to the cervix. Ligation of the major blood vessels is essential, particularly if the procedure is performed during oestrus or pregnancy as the vessels may be extremely well developed.
The vagina and vestibule
The vagina and vestibule form a channel leading to the external opening of the reproductive tract – the vulva. The vagina leads from the cervix to the external urethral orifice – the point at which the urethra joins the reproductive tract. The vestibule leads from the external urethral orifice to the vulva and is shared by both the urinary and reproductive tracts.
The lumen is lined by stratified squamous epithelium, which undergoes hormonal changes during the oestrous cycle. The lining epithelium is folded longitudinally to allow widthways expansion during parturition and is surrounded by layers of smooth muscle. These are very strong and during canine mating they tighten on the penis of the male and maintain the ‘tie’.
The epithelial cells lining the vagina undergo hormonal changes during the oestrous cycle. These changes may be used as an aid to the correct timing of mating. The technique, known as vaginal exfoliative cytology, involves making smears from vaginal swabs taken every other day during pro-oestrus and early oestrus. The smears are then stained and examined microscopically looking for diagnostic changes in the cells that provide an indication of the stages of the cycle.
The vulva
The vulva marks the external opening of the urogenital tract. It consists of two parts:
The mammary glands
Although these are not strictly part of the reproductive tract, they are essential to reproduction in the mammal. The presence of mammary glands is the defining characteristic of the class Mammalia. All mammals feed their young on milk produced by the glands during a process known as lactation.
Mammary glands are modified cutaneous glands. In the dog and cat, they are present in both sexes but are rudimentary in the male. The glands lie externally on the ventral wall of the abdomen and thorax, on either side of the midline.
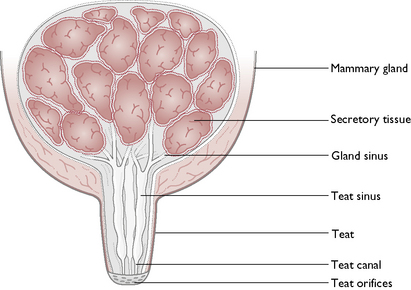
Each gland consists of glandular tissue embedded in connective tissue and lined by a secretory epithelium (Fig. 11.9). The milk produced drains through a network of sinuses that eventually form teat canals. These open on to the surface of each teat, known as a teat orifice. Each gland has one teat but each teat has several orifices.
Lactation
This is the production of milk and normally occurs during pregnancy. It is influenced by three hormones:
Composition of milk
This varies between different species and is an important consideration when feeding orphaned animals. The milk produced by the bitch and the queen is more concentrated and contains more protein and twice as much fat as cow’s milk. The average composition of milk is shown in Table 11.1.
Table 11.1 Average composition of milk
| Constituent | Quantity (%) |
|---|---|
| Water | 70–90 |
| Fat | 0–30 |
| Protein | 1–15 |
| Carbohydrate | 3–7 |
| Minerals | 1.5–1: calcium phosphate, magnesium, sodium, potassium and chlorideMilk is deficient in iron and copper; traces of iodine, cobalt, tin and silica are present |
| Vitamins | A, B2, B5, E, KMilk is low in vitamins C and D |
NB. Milk from a cat contains the amino acid taurine, as cats have a specific requirement for this.
The first milk secreted by the dam following parturition is known as colostrum. It is rich in maternal antibodies, which provide the neonate with immunity to diseases to which the dam has been exposed. It is essential that the neonate takes in colostrum within the first 24 hours of life. During this time, the protein antibodies can be absorbed by the small intestine without being digested. After 24 hours, normal protein digestion starts and the antibodies are broken down and destroyed. After a few days, production of colostrum stops and the composition of the milk remains constant.
The oestrous cycle
The oestrous cycle is the rhythmic cycle of events that occurs in sexually mature non-pregnant female mammals and includes limited periods of sexual receptivity known as oestrus. The function of the oestrous cycle is:
For the oestrous cycle to achieve the aim of a fertile mating, the timing of all interrelated components must coincide. The pattern and timing of the oestrous cycle varies between species – the cycle shown by the bitch is different from that shown by the queen.
Each cycle is divided into phases of varying lengths. These are:
During the oestrous cycle simultaneous changes occur in:
Ovary and reproductive tract
At birth, the ovary contains all the germ cells that animals will ever need; these act as a reservoir from which the primary follicles develop. At the onset of puberty or sexual maturity, serveral primary follicles develop to form ripe Graafian follicles. In multiparous species, there will be many follicles, divided between the two ovaries, but not necessarily equally. Each Graafian follicle consists of an ovum formed by the process of meiosis and containing the haploid number of chromosomes (see Ch. 1). This is suspended in fluid and surrounded by an outer layer of follicular cells. The Graafian follicle secretes the hormone oestrogen (Fig. 11.8).
When the follicle has reached full size, it ruptures to release the ovum – the process of ovulation. The ovum passes down the uterine tube and the remaining follicular tissue becomes reorganised to form the corpus luteum. The corpus luteum secretes the hormone progesterone.
Within the reproductive tract, the uterine walls become thickened and more glandular to create a suitable environment for implantation of the fertilised ova. The vaginal epithelium also changes and the bloodstained discharge seen in pro-oestrus in the bitch comes from the lining of the vagina.
Hormonal changes
Ovulation and the oestrous cycle are associated with a cycle of interrelated hormonal changes in the ovary and anterior pituitary gland (see Ch. 6, Fig. 6.4):
If the animal has not conceived, the corpus luteum regresses and the cycle begins again.
Pseudopregnancy (false pregnancy or pseudocyesis) is linked to the fact that the corpus luteum of the bitch remains in the ovary for about 6–7 weeks whether she is pregnant or not. Progesterone levels remain high and produce symptoms such as maternal behaviour, enlargement of mammary glands and lactation. Progesterone levels are normally high in all bitches but clinical signs only occur in certain bitches.
Behavioural changes and external signs
The bitch
Bitches usually have one or two oestrous cycles a year and during the period of oestrus they are described as being ‘in season’ or ‘on heat’. The bitch is monoestrous, i.e. during each period of ovarian activity there is only one period of oestrus, and there is no recognised breeding season – bitches may come into season at any time of the year. The bitch is a spontaneous ovulator, i.e. ovulation occurs without the stimulus of mating and takes place around the tenth day of the cycle (Fig. 11.10).
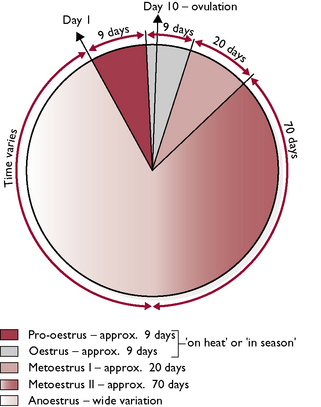
Bitches reach sexual maturity or puberty at about 6 months but there is wide variation between breeds – larger breeds mature significantly later than smaller breeds. Puberty is marked by the onset of the first season. The phases of the oestrous cycle are as follows:
 Metoestrus I: lasts for approximately 20 days, during which the vaginal discharge dries up, the swollen vulva shrinks and the bitch’s behaviour returns to normal. At the end of this phase, the bitch appears to be back to normal, but internally the corpus luteum in the ovary continues to secrete progesterone.
Metoestrus I: lasts for approximately 20 days, during which the vaginal discharge dries up, the swollen vulva shrinks and the bitch’s behaviour returns to normal. At the end of this phase, the bitch appears to be back to normal, but internally the corpus luteum in the ovary continues to secrete progesterone.The queen
The queen is seasonally polyoestrous. This means that she has several periods of oestrus or receptivity to the male during her breeding season, which is typically from January to September. Cats are ‘long day breeders’, stimulated to start cycling by the increasing hours of daylight that occur in the early spring. However, many domestic cats come into season all the year round as they live under the influence of central heating and electric light.
The queen is an induced ovulator, i.e. ovulation occurs in response to mating. The tip of the tomcat’s penis is covered in tiny barbs, which, when the penis is withdrawn at the end of mating, cause the queen a moment of pain. This starts a nerve pathway to the hypothalamus in the brain, and ultimately to the ovary, resulting in ovulation within 36 hours of mating.
Young queens become sexually mature in the spring after they are born. This means that if born in September, they may be as young as 4 months at the time of the first season in January/February. The phases of the oestrous cycle (Fig. 11.11) are as follows:
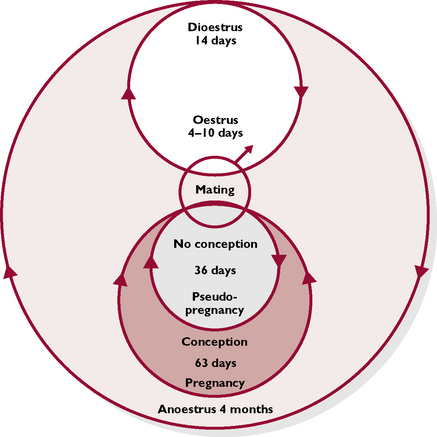
False pregnancies can occur in the cat but are very rare. If the queen is mated but does not conceive, a corpus luteum develops in the ovary and secretes progesterone, which switches off further follicular development. After about 36 days, the corpus luteum regresses and the cycles start again. There are rarely any clinical signs associated with this phenomenon in cats.
Embryonic and fetal development
The gestation period is the interval between fertilisation of the ovum and the birth of the offspring. In the bitch and the queen, the gestation period is approximately 63 days, although there is both individual and breed variation. During gestation, complex changes occur to the zygote that result in a viable neonate 9 weeks later.
Fertilisation and cell division
Each ovum released from the ovary during ovulation is covered in an outer protective layer of follicular cells known as the corona radiata and has an inner layer of glycoprotein known as the zona pellucida. The bitch and the queen are multiparous species, i.e. litter bearers, so they produce several ova at one time. The ova enter the infundibulum of the uterine tube and are transported down the uterine tube by muscular contractions and by the movements of the cilia of the epithelial lining.
After mating, sperm from the male travel up the female tract and fertilisation takes place within the upper part of the uterine tube. It is thought that sperm may live for as long as 7 days within the tract, relying on the correct environment within the uterine tube to survive. During this period the outer acrosome of the sperm (Fig. 11.4) releases enzymes, which are able to break down the zona pellucida of the ovum. Each ovum is then penetrated by one sperm, which results in a fertilisation reaction, preventing fertilisation by any other sperm (Fig. 11.12). The fertilised ovum is now referred to as a zygote and cell division begins within a few hours of fertilisation.
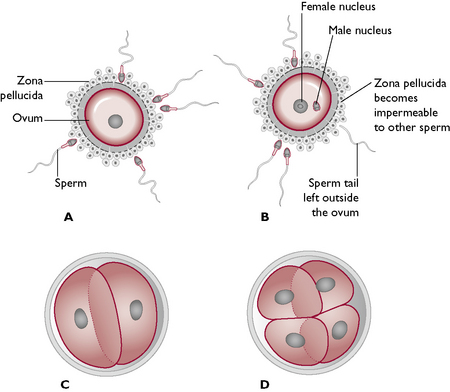
Fig. 11.12 Fertilisation of the ovum. A Spermatozoa approach ovum. B One sperm penetrates the ovum. C, D Cell division occurs by mitosis.
Cell division occurs by mitosis (see Ch. 1): one cell divides into two, two into four, and so on. The stage in which the cells are too numerous to count is called a morula. A fluid-filled cavity develops inside the morula and the structure becomes a blastocyst. During this time the ball of cells is free-floating, surviving on fluids within the lumen of the uterine tract, and it moves slowly down the tract towards the uterine horns.
Implantation
Once in the uterine horns, the blastocysts arrange themselves at equal distances along the horns and may even cross from one horn to another to achieve equal spacing – a process known as transuterine migration. They then attach to the wall of the horns by invading and partly destroying the hypertrophied endometrium, so that they are securely implanted. Implantation occurs between 14–20 days after ovulation in the bitch and 11–16 days after ovulation in the queen.
Development of the germ cell layers
Within the fluid-filled blastocyst, most of the cells come to lie on one side, forming the inner cell mass. A thinner layer of cells surrounding the fluid cavity form the trophoblast (Fig. 11.13). The inner cell mass becomes the embryo and, in the early stages, is a flat plate of cells. The trophoblast becomes the extra-embryonic membranes.
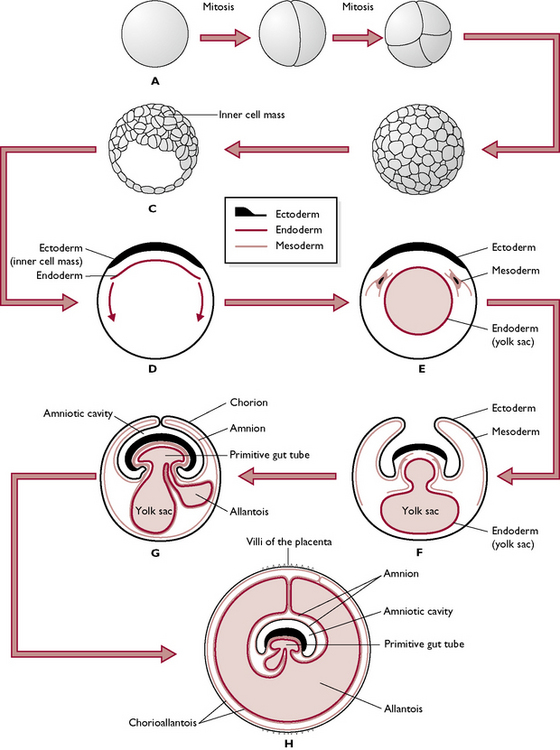
Fig. 11.13 Early embryonic development (see text also). A The fertilised cell divides by mitosis to form a ball of cells, the morula (B). The morula develops a cavity and is known as the trophoblast (C). A layer of endodermal cells starts to line the trophoblast (D) and forms the beginning of the yolk sac (E). Blocks of mesoderm start to form as the inner cell mass starts to be pinched off to form the embryo mesoderm (F). Another space forms from the primitive gut tube, this is the allantois (G) and the yolk sac begins to regress. Developing villi form a band around the extra-embryonic membranes (H).
The cells now begin to form three germ cell layers, which eventually differentiate into parts of the embryo and the different membranes:
An inner layer or endoderm – forms the lining of the digestive tract and other visceral systems.
Cells from the endoderm spread around to line the trophoblast and form the yolk sac. In mammals this does not contain yolk but in birds and reptiles the yolk sac is the source of nutrition for the embryo developing in the egg
The inner cell mass begins to curl around and enclose the endodermal and mesodermal cells, which form the internal organs, leaving the yolk sac and trophoblast to form the extra-embryonic membranes.
Development of the extra-embryonic membranes
The extra-embryonic membranes surround and protect the embryo but do not form the embryo itself:
Development of the embryo
As the inner cell mass grows, the cells begin to curve underneath and the head and trunk are formed. Inside, the main body cavity or coelom is formed and later it is divided into thorax and abdomen by the diaphragm. All the internal organs are developed by 35 days – this process is known as organogenesis. After this, the fetus goes through a period of rapid growth until it reaches its final size prior to birth.
The later stages of development in a medium-sized breed of dog are summarised in Table 11.2. Kittens are often slightly in advance of puppies of the same stage.
Table 11.2 The developmental stages of the canine embryo and fetus
| Timescale | Stage of development |
|---|---|
| 3 weeks | 5 mm long; fore and hindlimbs are small buds sticking out from the trunk; amnion is complete and the allantois is formed |
| 4 weeks | 20 mm long; limbs are small cylinders with evidence of a paw shape; eyes are pigmented; external ear has a ridge of visible skin |
| 5 weeks | 35 mm long; ear flap is distinct; eyelids partly cover the eyes; digits can be seen on the paws; external genitalia are near to final positions; tactile (sinus) hairs are present on the upper lip; formation of internal organs (organogenesis) is complete |
| 6 weeks | 60 mm long; prominent scrotal or vulval tissues; digits widely spread; eyelids are fused; hair follicles and tactile follicles present on the body; claws present; ossification of skeleton at 45 days |
| 7 weeks | 100 mm long; body hair and colour markings are developing |
| 8 weeks | 150 mm long; hair covering is complete; pads have developed |
| 9 weeks | Ready for birth |
The placenta
The placenta is a thickened vascular band that develops from the allantochorion around the centre of the conceptus. The allantochorion produces small, finger-like villi that burrow into the endometrium of the uterine horn. Blood capillaries covering the membrane extend into the villi and come into close apposition with the blood capillaries of the endometrium. These form the maternal and fetal parts of the placenta, which are not continuous with each other.
The villi develop into broad bands running around the ‘waist’ of the conceptus. Nutrients and oxygen pass from the dam across the placenta and into the fetus via the umbilical blood vessels; waste products pass in the opposite direction. The umbilical cord contains the umbilical artery and vein, the remnants of the yolk sac and the stalk of the allantois connected to the urachus (see Ch. 7).
The placenta of the dog and cat is restricted to one zone and is therefore described as being a zonary placenta (Fig. 11.14). Other species have more diffuse types of placenta. Between the placenta and the membranes, at the edge of the placenta, is an area where blood has escaped from broken blood capillaries and become trapped. This is the marginal haematoma and it stains the parturient discharges green in bitches and brown in queens. This discolouration is normal!
Fundamental genetics
Genetics is the science of inheritance and is the method by which characteristics or traits are passed from one generation to another. These characteristics may be visible, e.g. eye or coat colour but also may be the less obvious but vital characteristics such as blood grouping or metabolic pathways in the liver – the formation of every enzyme or protein in the body is controlled genetically. The information that determines these characteristics is carried by structures known as genes. Genes are arranged in long chains known as chromosomes, which are found within the nucleus of every cell in the body (see Ch. 1).
Chromosomes
Chromosomes are made of the protein deoxyribonucleic acid (DNA), which consists of two parallel unbranched strands twisting around each other in a double helix. The strands are formed by a series of four amino acids – adenine, thymine, guanine and cytosine – arranged in a varying sequence. The amino acids, which are also called bases or nucleotides, form cross-links with the opposite strand to create a ladder-like structure (Fig. 11.15). Each base only forms a cross-link with a specific other base, i.e. adenine only links with thymine and cytosine only links with guanine. These are then referred to as base pairs. The sequence of bases on one strand determines the sequence of bases on the opposite strand and this sequence or genetic code determines the makeup of each gene along the chromosome. The genetic code within each gene determines the type of protein formed by the cells.
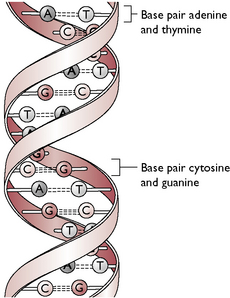
In a resting cell the helical structure of each chromosome is tightly coiled within the nucleus so that when the nucleus is seen under a light microscope it appears very dense. The chromosomes can be more easily differentiated when they uncoil prior to cell division by mitosis or meiosis (see Ch. 1).
Chromosomes are always considered in pairs – one of each pair is received from each parent. Each species of animal has a characteristic number of pairs of chromosomes within the nucleus of every cell (Table 11.3). The total complement of chromosomes is known as the diploid number. This becomes halved to the haploid number during meiosis. Thus each gamete (ovum or spermatozoon) formed by meiosis contains the haploid number. After fertilization, during which the nucleus of the ovum fuses with the nucleus of the sperm, the resulting zygote contains the diploid number of chromosomes and cell division to form the embryo begins.
Table 11.3 Characteristic number of chromosomes in the nucleus of a range of species
| Species | No. of pairs (diploid number) | Total number of chromosomes |
|---|---|---|
| Dog | 39 | 78 |
| Cat | 19 | 38 |
| Human | 23 | 46 |
| Horse | 32 | 64 |
| Sheep | 27 | 54 |
| Ox | 30 | 60 |
| Pig | 19 | 38 |
| Kangaroo | 6 | 12 |
Within the majority of chromosome pairs the chromosomes are identical to each other and they are described as being homologous (Fig. 11.16). Within each cell there is one pair of chromosomes that may not be homologous – these are the sex chromosomes. The other chromosomes in the cell are referred to as the autosomes and they influence all the characteristics that are not concerned with sex determination.

Fig. 11.16 Metaphase spread and karyotype of a dog. The dog has a total of 78 chromosomes in each cell. This chromosome spread is from a male. A female would have XX sex chromosomes (boxed) instead of the X and Y chromosomes.
(With permission from Long S 2006 Veterinary genetics and reproductive physiology. Butterworth-Heinemann, Edinburgh, p 27.)
Sex determination
Within each cell one pair of the total complement of chromosomes is the sex chromosomes, which are concerned with the determination of the sex of the animal. These are given the letters X and Y. The Y chromosome is small and carries the genes that determine male characteristics; there is very little room for any other genes. The X chromosome is larger and carries the genes that determine female characteristics, although it may carry other genes as well.
In the mammal the female has two X chromosomes while the male has an X and a Y, which means that the female has more space for genes than the male. To compensate for this, one of the X chromosomes of the female is inactivated and becomes contracted, forming a Barr body.
The male mammal is responsible for determining the sex of his offspring. During meiosis (see Ch. 1) within the ovary or testis four daughter cells are produced – these are either sperm or ova, known as the germ cells. The germ cells will be identical to each other but different from the parent cells. All the pairs of chromosomes split up and each one of a pair goes into the newly formed germ cells. Thus the sperm may contain either X or Y chromosomes while the ova will only contain X chromosomes. During fertilization the sperm cell fuses with the ovum to produce a zygote which will be either XY or XX, i.e. male or female.
This results in two male offspring (XY) and two female (XX) offspring, i.e. there is a 50% chance of producing either sex at each mating.
Genes
Genes are the units of inheritance. They are short sections of the base pairs making up the strands of DNA that are the chromosomes. The sequence of base pairs within the gene provides the code for each characteristic in the cell and hence the entire individual. Each gene influences one characteristic, although each characteristic may be influenced by several genes. These are known as polygenic characteristics.
In order to be able to describe genes and the way in which they function it is necessary to learn a new set of terms (Table 11.4). A particular gene will always be found at the same site on a particular chromosome – this site is referred to as its locus (pl. loci). The process of gene mapping is aimed at identifying the locus of every gene on every chromosome in the cell. As chromosomes are thought of in homologous pairs it follows that there will be another gene at the identical locus – this gene is referred to as its allele.
Table 11.4 Genetic terminology
| Term | Definition |
|---|---|
| Diploid number | Number of chromosomes found in a normal cell |
| Haploid number | Number of chromosomes found in a germ cell (ovum or sperm). This is half the diploid number and results from meiosis |
| Gamete | Either the ovum or the spermatozoon. Contains the haploid number of chromosomes |
| Zygote | Results from fusion of the ovum with the sperm and contains the diploid number of chromosomes |
| Mitosis | Method of cell division occurring in somatic cells, i.e. all the cells of the body except the germ cells |
| Meiosis | Method of cell division occurring in the germ cells |
| Locus | The position of a gene on a chromosome |
| Allele | A gene on the same locus of a pair of homologous chromosomes |
| Homozygous gene | Identical genes on the same locus |
| Heterozygous gene | Non-identical genes on the same locus |
| Dominant gene | A gene that overrides its allele |
| Recessive gene | A gene whose characteristic is not expressed when paired with a dominant gene but will be expressed with an identical recessive gene |
| Phenotype | The outward appearance of an individual |
| Genotype | The genetic makeup of an individual |
Genes at the same locus affect the same characteristic but they need not produce the same effect. For example both genes in a pair may affect eye colour but one may instruct the cells to make blue eye pigment while the other produces brown pigment. Genes at the same locus that produce the same effect, e.g. both brown eyes, are said to be homozygous; genes at the same locus that produce a different effect, e.g. one blue eyes and one brown eyes, are said to be heterozygous.
When a gene produces a characteristic effect it is said to be expressed. If only one of a pair of heterozygous genes is expressed the gene is described as being dominant and the gene that is not expressed is described as being recessive. Dominant genes override the effect of recessive genes and the recessive genes are only expressed when there is no dominant gene to override it, i.e. when they are homozygous. Occasionally there are cases where alleles giving different instructions for the same characteristic are both expressed, e.g. blood groups in humans. Such genes are described as being co-dominant.
The majority of genes act independently of each other but some genes can suppress the activity of genes at the same loci. These genes are said to exhibit epistasis. Epistasis is seen in albino animals and occurs when the albino gene suppresses all other genes associated with coat colour, resulting in a colourless or white coat.
An animal may have an external appearance that is different from what would be predicted by its genetic makeup. The external appearance of an animal is known as the phenotype. The genetic makeup is known as the genotype and this can only be accurately appreciated by examination of the DNA. The phenotype of an animal is influenced not only by the genes but by also by environmental factors such as diet, disease and exercise. This can be expressed as ‘phenotype = genotype + environment’. An example of this might be a fat whippet – this breed is naturally (genetically) very thin but can become fat (phenotype) by overeating (environment).
Mutations
It is important to understand that genes cannot normally be altered and will pass unchanged from generation to generation. The only reason that they may or may not be expressed in the phenotype is because they are dominant or recessive when mixed with the genes from another germ cell, i.e. after division by meiosis followed by fertilization. Occasionally a genetic change or mutation does occur but this is usually a random chance mutation or the result of chemical change or damage, e.g. by radiation. These changes will then pass into the next generation. Mutations may result in a harmful condition that could kill the affected individual, e.g. development of cancer cells, and then the gene will die out on the death of the affected individual. However it might produce a characteristic that gives the animal an advantage that could enhance its chances of survival, e.g. a change of coat colour, which might provide camouflage. The affected animal would then reproduce and pass its beneficial gene on to the next generation.
Genetic nomenclature
The language of genetics can be expressed using a system of letters, which can then be used to predict the outcome of mating or crossing one animal with a particular genotype to another animal with a different genotype.
Mendel’s laws of inheritance
For many years scientists were unaware of the method by which characteristics were passed from one generation to another, although they recognised that children bore a certain resemblance to their parents. In the mid-19th century an Austrian monk called Gregor Mendel (1822–1884), while working in the gardens of his monastery, developed an interest in plant and animal breeding. From his observations over many years he developed his laws of inheritance, which led him to be able to predict the outcome of various crosses.
Specialized types of gene
Most genes follow Mendel’s laws and are passed on predictably; however, there are some types of gene that may have effects that are less predictable.
Sex-linked genes
Genes that are located on one of the sex chromosomes, i.e. X or Y, are therefore most commonly associated with one or other of the sexes. Most sex linked genes are found on the X chromosome as it is larger than the Y chromosome and has more space to carry genes. Sex-linked genes are usually recessive and examples include the genes for haemophilia, colour-blindness and orange coat colour in cats. These characteristics are almost always associated with males, although they may occasionally affect females.
The mechanism of inheritance is:
Sex-limited genes
These are genes that influence a characteristic that is seen in only one sex. These genes are switched on by reproductive hormones produced by the animal at the onset of puberty, i.e. oestrogen or testosterone. Examples of sex-limited genes include breast size in women and milk yield in cows.
Lethal genes
These are genes that are incompatible with life. An animal that expresses a lethal gene will die from its effects sooner or later. They are usually recessive genes so affected individuals must be homozygous for the gene. There are three types of lethal gene:
Selective breeding
Selective breeding is the term used to describe the selection of certain individuals as stud animals for breeding rather than allowing all animals to breed randomly. In theory this means that beneficial or desirable characteristics will be passed on to the next generation while harmful or deleterious characteristics will be bred out, but in fact if selective breeding is carried out incorrectly it can create even more problems. Using one’s knowledge of genetics it is possible to predict the outcome of crossing one animal with another and so create an individual that is a perfect representation of its breed and is not afflicted by any inherited conditions.
Monohybrid crosses
Matings in which only one characteristic is identified and followed through the generations are described as monohybrid crosses.
For example: spot colour in Dalmatian dogs. There are two variations in the spot colour – black, which is more common, and liver. The gene for black spots is dominant while the gene for liver spots is recessive.
Let the gene for black spots be represented as the letter B
Let the gene for liver spots be represented as the letter b
1. Mate a homozygous black-spotted dog to a homozygous liver-spotted bitch
Using a grid known as a Punnett square we can work out the genotypes for the first filial generation, referred to as the F1 generation.
| Genes from the male (in the sperm) | ||
| B | B | |
| b | Bb | Bb |
| b | Bb | Bb |
| Genes from the female (in the ova) | ||
The F1 generation will consist entirely of puppies with the Bb genotype and their phenotype will be black-spotted.
2. Mate a heterozygous black-spotted dog to a heterozygous black-spotted bitch
| B | b | |
| B | BB | Bb |
| b | Bb | bb |
The genotypes will be 1BB:2Bb:1bb
The phenotypes will be three black-spotted puppies to one liver-spotted puppy.
There is a 1 in 4 chance of producing the double-recessive individual when you use a monohybrid cross. When crossing two animals that are heterozygous for any characteristic you will always get the same ratio of genotypes of 1:2:1. Statistically if you use a large sample of animals you are more likely to achieve this ratio. In a litter of four puppies you might not breed the liver-spotted puppy, however if you produced 100 puppies you improve the chances of breeding a liver-spotted puppy significantly.
3. Mate a homozygous liver-spotted dog to a heterozygous black-spotted bitch
| b | b | |
| B | Bb | Bb |
| b | bb | bb |
The phenotypes will be two black-spotted puppies to two liver-spotted puppies – there is an equal chance of producing black- or liver-spotted puppies.
Back-crossing to the recessive
This is a technique using test matings to ascertain whether an animal is carrying a particular recessive gene. This may be a gene that causes a harmful condition such as the eye disease progressive retinal atrophy or, in more minor cases, an unwanted coat colour. A recessive gene will only be expressed if an animal is homozygous for that gene. If the animal is heterozygous, the gene will not be expressed but the animal will carry the gene and may pass it on to its offspring.
For example: progressive retinal atrophy (PRA) is a condition seen in breeds such as Labradors, Golden Retrievers, Collies and Spaniels. It causes gradual deterioration of the retinal blood vessels and results in blindness by the time the dog is about 18 months old. It is important to identify carriers of the gene so that they do not pass it on to their puppies.
Let the gene for normal sight be S
The suspect bitch under investigation could have the genotype Ss or SS
1. Suppose the suspect bitch is Ss
Mate her to a dog who is known to have PRA and will therefore have the genotype ss.
| s | s | |
| S | Ss | Ss |
| s | ss | ss |
The puppies will be 2Ss, i.e. carriers of the gene but unaffected: 2ss, i.e. affected by the condition.
2. Suppose the suspect bitch is SS
Mate her to the known affected dog ss.
| s | s | |
| S | Ss | Ss |
| S | Ss | Ss |
All the puppies will have the genotype Ss, i.e. they will be unaffected by PRA but as they will be carriers of the gene they would have to be neutered and not used for breeding.
In this example if a bitch produces puppies that are not affected you can be sure that she is homozygous for normal sight; if she produces puppies that are affected by PRA she must be heterozygous. If the heterozygous bitch was mated to an unidentified heterozygous dog the F1 generation would produce genotypes in the ratio of 1SS:2Ss:1ss, i.e. there is a 1 in 4 chance of producing an affected puppy.
It is essential to identify carriers of harmful genes but in practice test matings are expensive and take time. Analysis of pedigrees may be helpful, assuming they are correct, but nowadays DNA analysis may be much quicker and much more accurate.
Dihybrid crosses
A dihybrid cross is one in which two characteristics are identified. Each gene obeys Mendel’s second law of independent assortment and will move independently. It may be paired with any other gene, resulting in a wide variation of genotypes and phenotypes. This can appear very complicated but, as in the previous monohybrid cross, if you cross two animals that are heterozygous for both characteristics you end up with a predictable ratio of genotypes, in this case of 9:3:3:1 (compared to 1:2:1 for a monohybrid cross) and the chances of producing an animal that is recessive for both characteristics is 15:1 (compared to 3:1 for a monohybrid cross).
If you expand this idea to crossing individuals with three characteristics it is known as a trihybrid cross. When you cross two animals that are heterozygous for all three characteristics the predicted ratios of the genotypes are 27:9:9:9:3:3:3:1 and the chances of producing the one individual that is homozygous for all three recessive characteristics is 63:1.
This theory is extremely important when attempting to breed the perfect example of a particular species or breed or just a perfectly healthy animal. The aim is to achieve homozygosity for beneficial characteristics without achieving homozygosity for harmful characteristics. It is important to understand the reducing chances of producing an individual that is homozygous for large numbers of characteristics.
Breeding strategies
Hundreds of years of selective breeding of cats and dogs have led to the development of distinct breeds. A breed may be defined as ‘a group of animals of the same species linked by a set of common characteristics’. When an animal of one breed is crossed with another animal of another breed the result is a crossbreed or mongrel, which is a normal fertile animal that is capable of reproducing and passing on the characteristics of both breeds to the next generation but will bear very little resemblance to its parents. The advantage of crossing two animals of the same breed is that you can predict what the offspring will look like. When crossing two mongrels the assortment of the genes may be completely random and the outcome unpredictable. The skill of breeding lies in understanding Mendel’s laws and in selecting the correct animals to be mated to each other. Random selection could result in a Crufts champion but it would be very, very lucky.
The aim of selective breeding is to achieve homozygosity for all the genes which control the desirable characteristics.
For example: Supposing an animal that is homozygous for gene Z is mated to an animal that is heterozygous for gene Z
Then the F1 generation will be:
| Z | Z | |
| Z | ZZ | ZZ |
| z | Zz | Zz |
If one of the homozygous individuals is then crossed back to one of the parents, i.e. ZZ × ZZ, then the offspring of the F2 generation will all be homozygous. The parents of the F2 generation are described as breeding ‘true’ for that characteristic and the outcome is predictable. When homozygosity is achieved the characteristic is described as being fixed in the breed. In attempting to reach this situation breeders will use a range of breeding strategies.
Congenital and inherited defects
Selective breeding aims to fix desirable characteristics but it can also result in the fixing of harmful genes. These are associated with poor health and in some cases eventual death. Animals may be born with a variety of harmful conditions but these need not necessarily be caused genetically.
Examples of inherited defects in dogs and cats include: