14 The Abdomen of the Dog and Cat
CONFORMATION AND SURFACE ANATOMY
The cranial boundary of the accessible abdominal wall is easily determined by palpation of the last rib and costal arch, but the caudal boundary is more difficult to discover because only the ventral part (pecten ossis pubis) of the bony ring about the pelvic inlet can be palpated between the thighs. The wings of the ilia, though prominent landmarks, rise above the level of the abdomen and pertain to the back. The thick muscles above the lumbar transverse processes are palpable but not the processes themselves; however, the tips of the spinous processes provide a guide to the identification of individual vertebrae.
The abdominal cavity is of course larger than these landmarks appear to indicate because the diaphragm bulges far into the rib cage at its cranial end. The organs in this intrathoracic part of the abdomen are protected by the ribs and are in part overlain by the caudal lobes of the lungs. The abdominal cavity is relatively less voluminous than in the large domestic species and has, by and large, the shape of a cone with a bulbous cranial base (Figure 14–1). Its longitudinal axis inclines cranioventrally at an angle that varies considerably; it is steepest in deep-chested breeds. Except in fat subjects and heavily pregnant or lactating bitches, the ventral abdominal wall rises from the sternum to the pecten in a straight or even slightly concave line. Dog fanciers use the expression “tucked-up” to describe animals with an especially shallow body depth at the loin. The skin fold that connects the flank with the stifle tends to obscure the shallowness of this part. Advancing pregnancy enlarges the abdomen in both depth and breadth and gives it a more cylindrical or even a barrel shape.
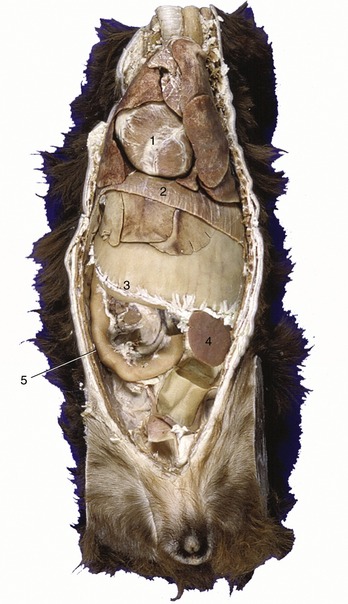
Figure 14–1 Ventral view of a canine trunk, demonstrating the so-called intrathoracic part of the abdomen. 1, Heart; 2, diaphragm; 3, distended stomach (with attachment of greater omentum); 4, spleen; 5, duodenum.
Superficial inguinal lymph nodes may be palpated in the groin, lateral to the bulbus glandis of the penis or in a comparable site in the bitch (Figure 14–2, B/6).
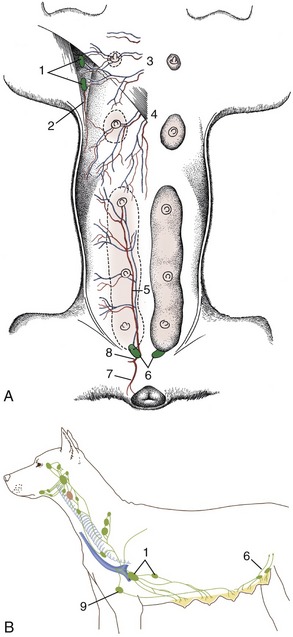
Figure 14–2 Blood vessels and lymphatics of the canine mammary glands. A, Ventral view of the mammary glands, blood vessels, and certain lymph nodes. B, Lateral view of regional lymph nodes. 1, Axillary and accessory axillary lymph nodes; 2, branch of lateral thoracic artery; 3, perforating branches of internal thoracic vessels; 4, branches of the cranial superficial epigastric vessels; 5, caudal superficial epigastric artery; 6, superficial inguinal lymph nodes; 7, ventral labial branch to vulva; 8, external pudendal artery; 9, sternal lymph node.
MAMMARY GLANDS
The mammary glands contribute to the contours during pregnancy and lactation. Dogs generally have five pairs of mammary glands, spread along the ventral aspect of the trunk (Figure 10–31, C, and Figure 10–32, B-C). The two cranial pairs are thoracic, the next two abdominal, and the caudal-most pair inguinal in position. A distinct midline separation is noted between the left and right mammary chains. Their pattern is often staggered, which is a favorable arrangement because it makes all teats equally accessible to the pups when the bitch suckles lying on her side. The glands are very small in the virgin (with the teats hidden by hair) but become very swollen, pendulous, and confluent with their ipsilateral neighbors toward parturition and during lactation. They regress greatly in the parous but nonpregnant and nonlactating bitch, although the teats remain enlarged in parous bitches, in which they are superimposed on the abdominal organs in ventrodorsal radiographs. The teats, which occur in rudimentary form in males, are bare and perforated at their tips by 10 or 12 fine openings through which milk is drawn.
The cat has four pairs of mammary glands of which the teat is situated approximately 3 cm from the midline. The nonlactating teat of the cat is about 1 mm high, perforated by four to eight openings, and enlarges almost tenfold in lactation.
In dogs and cats, the blood supply to the mammary glands varies in detail but mainly originates from the lateral and internal thoracic and the external pudendal arteries; some assistance may be provided by lesser vessels from other sources. In most cases, the cranial three (in cats two) glands are supplied craniolaterally by the lateral thoracic artery (from the axillary) and deeply by the cranial superficial epigastric artery and perforating branches of the intercostal arteries (both from the internal thoracic). The two caudal pairs receive blood from the caudal superficial epigastric artery (from the external pudendal) and deeply from branches of the cranial abdominal and deep circumflex iliac arteries. The veins are satellite. Both arteries and veins anastomose freely, forming arterial and venous plexuses (Figure 14–2, A), which may cross the midline.
Lymph from the cranial three (in cats two) glands goes to the axillary, accessory axillary, and sternal nodes, and that from the two caudal glands goes to the superficial inguinal (mammary) node, which is located dorsal to the caudal border of the inguinal mammary gland (or in the cat, also to the caudal epigastric nodes, which are small and located along the course of the caudal epigastric vessels). In the dog, the third pair usually drains toward the axillary lymph node but may also drain caudally. The pathways are erratic, and some lymph may cross the midline. It is said that in the cat the lymph vessels do not cross the midline nor penetrate the thoracic wall. The superficial inguinal nodes and caudal glands are related to the vaginal process, which is vulnerable during surgical removal of a diseased gland; injury to the process may cause inadvertent opening of the peritoneal cavity. In both species the superficial inguinal nodes drain the adjacent part of the abdominal wall in addition to the caudal mammary glands.
These details obtain importance from the prevalence of mammary tumors in both dogs and cats. In bitches they are the commonest of all tumors and show a disturbingly high (ca. 50%) incidence of malignancy. Although somewhat less common in cats, mammary tumors are even more likely to be malignant in this species.
THE ABDOMINAL WALL
(See also pp. 52–55.)
The ventrolateral abdominal wall is constructed according to the common pattern and has only a few features of distinction; because abdominal surgery is so frequently performed in dogs and cats, it is necessary to be familiar with the details and it may be prudent to review the description previously given. The principal distinctions concern the linea alba and rectus sheath, which are now described more fully because most abdominal incisions are median or paramedian. The description of the inguinal canal also bears recapitulation.
The linea alba is the fibrous seam in which the aponeuroses of the right and left oblique and transverse abdominal muscles come together. It extends from the xiphoid process to the pubis and includes the umbilicus at about the level of the third lumbar vertebra. The linea alba is about 1 cm* wide cranial to the umbilicus but gradually narrows behind this point and is reduced to a barely visible line in its caudal third (see Figure 2–26). Incisions through the linea alba spare the muscles, vessels, and nerves; there is the additional advantage that the parietal peritoneum does not retract from the edges of a median incision as happens elsewhere. The falciform ligament (see further on) and the median ligament of the bladder attach to the dorsal surface of the linea alba, cranial and caudal to the umbilicus, respectively. Umbilical hernias, often associated with an over-wide linea alba and hypoplastic rectus muscles, are common.
The rectus sheath is formed by the aponeuroses of the oblique and transverse abdominal muscles. In the dog and cat the aponeuroses of the external and internal oblique muscles pass ventrally (externally) to the rectus muscle over the length of the linea alba. However, the most cranial portion of the internal oblique muscle also detaches an additional lamina that passes dorsally (internally), on the deep surface of the rectus (Figure 2–26, A). The aponeurosis of the transverse abdominal muscle passes dorsally (internally) to the rectus muscle in the cranial half of the abdomen, but it changes position to the ventral surface in the caudal portion, leaving the dorsal surface of the rectus covered only by fascia and peritoneum (see Figure 2–23, B). The rectus muscle is adherent to its sheath only at the tendinous inscriptions.
The inguinal canal is a potential space between the external and internal abdominal oblique muscles that extends between deep and superficial openings (rings). The deep ring leads from the canal into the abdominal cavity, and the superficial ring leads from the canal to the subcutaneous tissues of the groin. In both sexes the canal conveys the external pudendal vessels and the genitofemoral nerve; it also conveys the spermatic cord in the dog and tom and the vaginal process in the bitch and queen. These all emerge at the superficial inguinal ring, a nearly sagittal slit in the external abdominal oblique aponeurosis about 3 cm lateral to the linea alba, close to where this attaches on the pubis (see Figure 2–27, A/4′). Only the caudal end of the ring is palpable. The narrow strip of aponeurosis (Figure 2–27, A between 4′ and 6) lateral to the ring forms the only barrier between the structures issuing from the canal and the large femoral vessels and saphenous nerve as they enter the thigh through the vascular lacuna (Figure 2–27/6).
The deep inguinal ring is visible only from within the abdomen. It is bounded caudolaterally by the caudal border of the external abdominal oblique aponeurosis (inguinal ligament), cranially by the unattached border (caudal edge) of the internal abdominal oblique muscle, and medially by the rectus muscle (see Figure 2–27, B). None of these boundaries is palpable in the intact animal. The parietal peritoneum that covers the ring evaginates through the inguinal canal and, named as the vaginal tunic, accompanies the spermatic cord into the scrotum. In the bitch and queen it envelops the round ligament of the uterus and is known as the vaginal process; the process is not present in females of other domestic species and is the occasional recipient of herniated abdominal organs (p. 461).
BLOOD SUPPLY
The ventral abdominal wall is supplied by four paired arteries, entering from the sternal and pelvic region, respectively. The cranial superficial epigastric artery branches off the internal thoracic artery, and runs between the abdominal muscles and the skin. It supplies the region cranial to the level of the umbilicus (it is enlarged in the lactating bitch). The cranial epigastric artery runs deep to the rectus, between it and its sheath. The caudal superficial epigastric artery, a branch of the external pudendal, is distributed subcutaneously and also supplies the prepuce; the caudal epigastric artery arises from the pudendoepigastric trunk and passes forward, first along the lateral border and then on the deep surface of the rectus muscle (see Figures 14–3 and 2–26). Cranial and caudal sets of vessels anastomose (Figure 14–2).
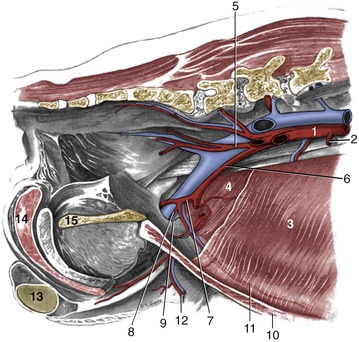
Figure 14–3 Abdominal wall and pelvic canal of the male dog, showing the breakup of the aorta; medial view. 1, Aorta; 2, caudal mesenteric a.; 3, transversus abdominis; 4, internal abdominal oblique m.; 5, internal iliac a.; 6, external iliac a.; 7, deep femoral a.; 8, pudendoepigastric trunk; 9, deep inguinal ring; 10, rectus abdominis m.; 11, caudal epigastric a.; 12, external pudendal a.; 13, left testis; 14, bulb of the penis; 15, pelvic symphysis.
The abdominal wall is most safely punctured (paracentesis) a short distance caudolateral to the umbilicus; this site avoids both the fat-filled falciform ligament and risk of injury to a full bladder. The falciform ligament, carrying the round ligament of the liver in its free border, is the remnant of the ventral mesogastrium that conveyed the umbilical vein from the umbilicus to the liver in the fetus. The part adjacent to the liver survives, if at all, as a simple peritoneal fold. The blood supply of the falciform ligament arises from along the length of the linea alba. The ligament commonly serves as a major fat storage depot and may become so thickened and enlarged that it complicates the opening and closure of a midline abdominal incision (Figure 14–11), especially in dogs. Part or all of this obstruction may be excised; care must be taken to place a ligature at the cranial end, before the ligament is totally removed.
GENERAL ASPECTS OF VISCERAL TOPOGRAPHY
Although the small intestine dominates the abdominal topography, it is not usually visible immediately when the cavity is opened because it is separated from the abdominal floor by the especially well-developed greater omentum (Figure 14–4). The organs that are usually exposed on removal of the abdominal floor are the ventral part of the spleen projecting beyond the left costal arch, a part of liver behind the xiphoid process, and the bladder directly before the pubis (see Figure 14–5 and Figure 14–6).
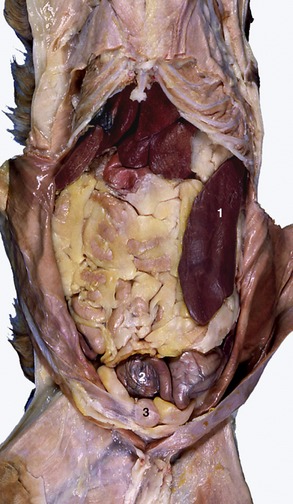
Figure 14–4 Ventral view of feline abdominal viscera; intestinal loops are concealed by fat-filled greater omentum. 1, Spleen; 2, part of gravid uterus with two ampullae; 3, bladder.
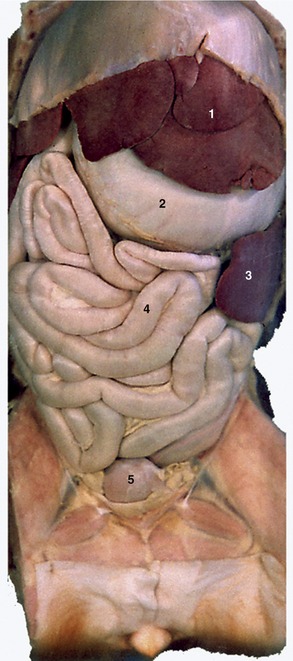
Figure 14–5 Abdominal viscera of the dog after removal of the greater omentum. 1, Liver; 2, stomach; 3, spleen; 4, small intestine; 5, bladder.
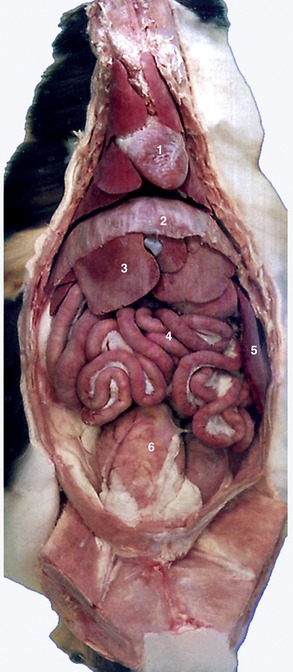
Figure 14–6 Ventral view of feline viscera after removal of omentum. 1, Heart; 2, diaphragm; 3, liver; 4, intestine; 5, spleen; 6, bladder.
The greater omentum is extremely well-developed and is folded on itself to form a flat sac with superficial and deep leaves that intervene between the intestinal mass and the abdominal floor (see Figure 3–33). The omental bursa exists as a potential space between the leaves. The opening to the omental bursa, the epiploic foramen, is a narrow passage that lies medial to the caudate process of the liver and is bounded dorsally by the caudal vena cava and ventrally by the portal vein.
Being the dorsal mesogastrium, the greater omentum attaches to the greater curvature of the embryonic stomach, as in other species. It arises from the roof of the abdominal cavity, near the caudal part of the liver and the celiac artery. Close to this attachment the left lobe of the pancreas is enclosed in the omentum. The dorsal attachment of the omentum runs between the esophageal hiatus and the epiploic foramen. At this point the greater omentum continues as mesoduodenum, in which the right lobe of the pancreas is situated. The omental bursa is connected to a hilus of the spleen by the gastrosplenic ligament. The omentum is connected to the descending colon caudally by the omental veil.
The superficial leaf (Figure 3–33/14) passes caudally from its attachment, in direct contact with the ventral abdominal wall to reach the bladder, where it is reflected dorsally to become the deep leaf (Figure 3–33/13). This runs forward between the superficial leaf and jejunal coils; at the cranial end of the jejunum it passes dorsally, against the caudal (visceral) surface of the stomach, to reach the left lobe of the pancreas, which it encloses and by means of which it gains the roof of the abdominal cavity. The right border of the omental sac is ventral to the descending duodenum; the left extends more dorsally to the level of the kidney and sublumbar muscles and is complicated by an attachment to a hilus of the spleen. The part of the omentum extending between the left crus of the diaphragm and the splenic hilus may be known as the phrenicosplenic ligament; the more generous part between the stomach and hilus forms the gastrosplenic ligament. As a further complication, a sagittal fold (omental veil) with a caudal free border connects the deep leaf with the left surface of the descending mesocolon.
The greater omentum always contains fat. This is first deposited along the small omental vessels, giving the structure a lacy appearance; however, in obese dogs (less so in cats) it forms a more or less continuous layer.
The lesser omentum is considerably wider than the short space it has to bridge between the lesser curvature of the stomach and the liver. It blends on the right with the mesoduodenum, the bile duct marking the boundary between the two. The papillary process of the liver is loosely enveloped by the lesser omentum. The portion of the lesser omentum between the liver and the duodenum is also called the hepatoduodenal ligament; between the liver and the stomach it is called the hepatogastric ligament.
THE SPLEEN
The form of the spleen (see also p. 264) is the same in dogs and cats; it is an elongated, roughly dumbbell-shaped organ that lies more or less vertically against the left abdominal wall (Figure 14–7, A/4). Its position is much influenced by the distention of the stomach (and by its own capacity to become engorged). The dorsal end reaches the left crus of the diaphragm, passing between the gastric fundus and the cranial pole of the left kidney under cover of (usually) the last two ribs. The larger ventral end may cross the ventral midline, to reach under the costal cartilages of the right side. It then provides a dense triangular shadow on the abdominal floor in lateral radiographs (Figure 14–8, A/3). A similar shadow between the stomach and left kidney may reveal the position of the organ in ventrodorsal films. In the cat, the ventral part of the spleen is always located outside the rib cage. The parietal surface makes contact (in dorsoventral sequence) with the diaphragm, costal arch, and abdominal muscles. The visceral surface is divided by a hilar ridge into a cranial strip related to the stomach and a caudal strip related to the left kidney and intestine.
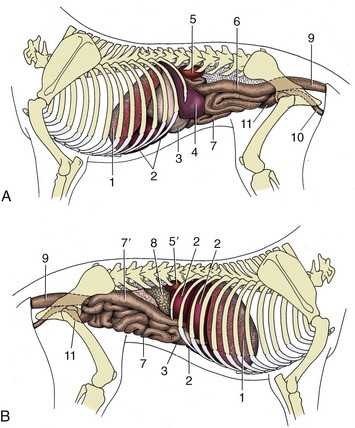
Figure 14–7 Visceral projections on the left (A) and right (B) canine abdominal walls. 1, Diaphragm; 2, liver; 3, stomach; 4, spleen; 5, 5′, left and right kidneys; 6, descending colon; 7, small intestine; 7′, descending duodenum; 8, pancreas; 9, rectum; 10, female urogenital tract; 11, bladder.
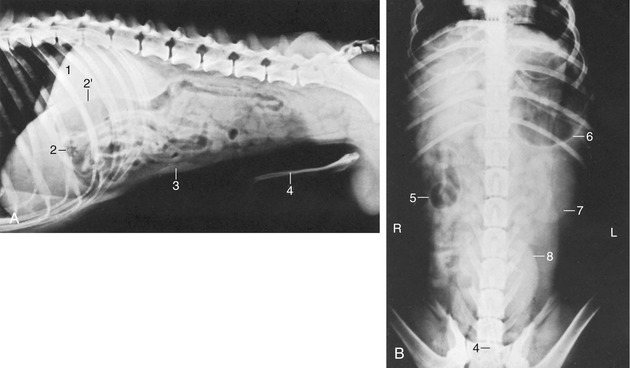
Figure 14–8 Lateral (A) and ventrodorsal (B) radiographic views of the canine abdomen. 1, Liver; 2, pyloric part of stomach; 2′, descending duodenum; 3, spleen; 4, os penis; 5, cecum; 6, fundus of stomach; 7, left kidney; 8, bladder.
The wide gastrosplenic ligament attaches the spleen to the greater curvature of the stomach. Although permitted considerable mobility, the spleen follows the movements of the stomach. When the stomach enlarges, the spleen is displaced caudally and ventrally, reaching the pelvic inlet; it may then be palpated through the abdominal wall.
Another restraining influence is provided by the tether of its blood vessels. The splenic artery and vein pass (as several divergent branches) to the dorsal end of the spleen. The splenic artery arises as a branch of the celiac artery, and before reaching the spleen, it gives off branches to the left limb of the pancreas. The left gastroepiploic vessels are detached about the middle of the hilus and cross to the greater curvature of the stomach within the gastrosplenic ligament (Figure 14–9/3,11). The splenic lymph nodes lie by the splenic vessels, a few centimeters distant from the organ. The spleen has efferent (which follow large arteries) but no afferent lymphatic vessels.
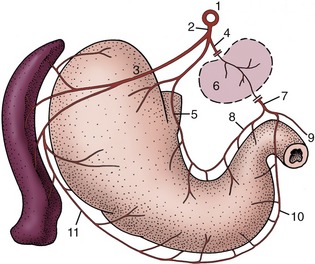
Figure 14–9 The blood supply of the stomach and spleen, caudal view; schematic. 1, Aorta; 2, celiac a.; 3, splenic a.; 4, hepatic a.; 5, left gastric a.; 6, indication of the liver; 7, gastroduodenal a.; 8, right gastric a.; 9, cranial pancreaticoduodenal a.; 10, right gastroepiploic a.; 11, left gastroepiploic a.
The spleen serves as an important blood reservoir in the dog and cat, and its size and weight therefore vary widely (Figure 14–6). The spleen in a resting dog or cat contracts and relaxes rhythmically because of the presence of many smooth muscle fibers throughout the organ. These fibers relax when anesthetics are used, which results in marked splenic enlargement, and contract because of stress or injection of catecholamines, expelling free blood cells and plasma from the red pulp. The spleen has no parasympathetic nerve supply.
Rupture of the spleen is not uncommon after traffic accidents, but fortunately the organ may be removed without risk to life. The relatively loose attachment of the spleen to the stomach facilitates access to the vascular supply at surgery (splenectomy*).
THE STOMACH
The dog has a simple stomach (see also pp. 123–129) that exhibits the idealized form described on page 125 only when moderately full. The fundus and body merge smoothly and are capable of great expansion, while the cylindrical and thicker walled pyloric part is less able to enlarge. The fundus projects dorsally to the left of the cardia, against the liver. The cardia is generally wide, and this may be related to the ease with which dogs vomit. The pylorus, on the other hand, is narrow, and pyloric stenosis is not uncommon in the young. When the organ is quite empty, the body also becomes more or less cylindrical, and the fundus then forms a bulbous dorsal enlargement. When the organ is greatly distended, all parts except the pyloric canal merge in a common sac. The capacity of the stomach ranges from 0.5 to 6.0 L with the average about 2.5 L; it is thus relatively large in relation to body size.
The position and relations obviously depend on the degree of fullness; the cardia provides a fixed point opposite the ninth intercostal space. The fundus and body lie mainly to the left of the median plane, in contact with the diaphragm and liver, respectively, but the ventral part of the body crosses to the right before being continued by the pyloric part, which also lies against the liver (Figure 14–11/3,4,7); indeed, its lesser curvature is bound to the porta of the liver by the lesser omentum. The greater curvature faces mainly to the left, toward the spleen, and ventrally, where it usually lies on the ventral fringe of the liver and on the falciform ligament (Figure 14–11/6); it reaches the abdominal floor only when the stomach is greatly distended, and in these circumstances it may be palpated through the abdominal wall. Otherwise, the stomach is out of reach and aligned with the ninth to twelfth ribs of the left side (or thereabouts; Figure 14–12, A). As the stomach expands, its ventral parts (mainly the body) move caudoventrally into broad contact with the abdominal floor and left costal arch, displacing the jejunum from contact with the liver in the process. Excessive distention, not uncommon in this greedy species, may carry the stomach to a level behind the umbilicus. Such gross enlargement also alters its cranial relationships, pushing the liver to the right and the diaphragm forward, reducing the thoracic cavity.
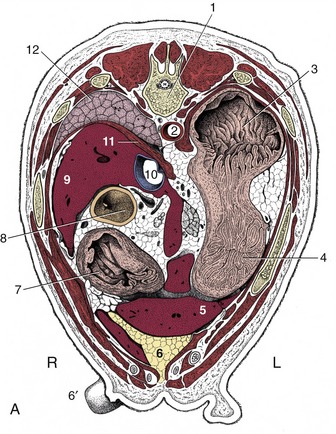
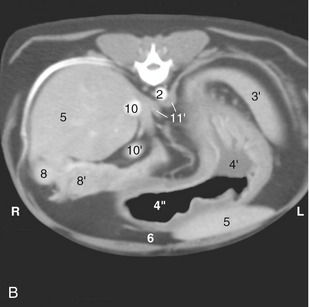
Figure 14–11 A, Transverse section of the canine trunk at the level of the twelfth thoracic vertebra. B, Corresponding computed tomographic (CT) image slightly more caudal than A; the dog was lying on its back during the CT procedure. 1, Twelfth thoracic vertebra; 2, aorta; 3, fundus of stomach; 3′, spleen; 4, body of stomach; 4′, with fluid; 4″, with gas; 5, liver; 6, fat-filled falciform ligament; 6′, teat; 7, pyloric part of stomach; 8, descending duodenum; 8′, right lobe of pancreas; 9, caudate process of liver; 10, caudal vena cava; 10′, portal vein; 11, diaphragm; 11′, crura of diaphragm; 12, right lung.
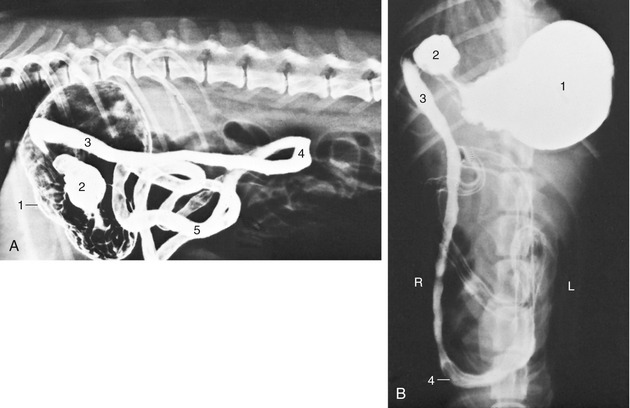
Figure 14–12 Lateral (A) and ventrodorsal (B) radiographic views of the canine abdomen after administration of a barium suspension. 1, Stomach; 2, pyloric part; 3, descending duodenum; 4, caudal flexure of duodenum; 5, jejunum.
Survey radiographs of the abdomen generally reveal few details of the stomach beyond the gas that naturally collects in the uppermost part of the organ—the fundus in the animal standing or in right lateral recumbency. This useful orientation feature is lost when the animal is placed in other positions. A more complete demonstration of the topography is obtained with the administration of a barium meal (Figure 14–12). The existence of the rugae may be revealed by defects in the outline of the contrast mass; the most satisfactory depiction is obtained after the evacuation of the bulk of the meal, when the residual agent clings to the mucosa and fills the spaces between adjacent rugae.
A number of structures join the stomach to neighboring parts. The fundus is directly bound to the left crus of the diaphragm (gastrophrenic ligament), while there are looser attachments between the cardia and the diaphragm, the lesser curvature and liver (lesser omentum), and the greater curvature and spleen (greater omentum). Except at these reflections, the stomach is completely covered with serosa.
The stomach receives blood from all three branches into which the celiac artery divides directly after leaving the aorta between the crura of the diaphragm. The branches to the stomach therefore approach from the right of the fundus and dorsal to the cardia (Figure 14–9). The splenic artery supplies short branches as it crosses the caudal surface of the fundus before reaching the spleen. A more substantial branch (left gastroepiploic artery; Figure 14–9/11) follows the greater curvature to an anastomosis with the right gastroepiploic artery (a branch of the hepatic artery). The left gastric artery (Figure 14–9/5) supplies the fundus, cardiac region, and a branch to the esophagus before following the lesser curvature to an anastomosis with the right gastric artery (Figure 14–9/8), a further branch of the hepatic artery. The arterial arcades that follow the curvatures supply fair-sized branches to adjacent parts of both surfaces. The arteries are mostly accompanied by satellite veins, which contribute as gastrosplenic and gastroduodenal veins to the portal vein. Gastric lymphatics drain into hepatic lymph nodes but may have passed the splenic and gastric nodes first. Large vessels are absent from the strips midway between the curvatures, which are therefore the preferred locations for incision. The parietal surface can be exposed and opened through a midline or paracostal incision (a common procedure for the recovery of foreign bodies), but the visceral surface is inaccessible unless the omental bursa is opened first (see p. 122).
Gastric volvulus is relatively common, especially in large deep-chested breeds. In this mishap the distended stomach rotates about the esophagus (usually in a clockwise direction as seen from behind, between 270° and 360°), and this closes the esophagus at the cardia. The pyloric end of the stomach, less firmly held in place by the lesser omentum and bile duct, moves ventrally and to the left, which stretches the cranial part of the duodenum across the ventral surface of the cardia. The ventral leaf of the greater omentum, still attached to the greater curvature of the stomach, covers the ventral aspect of the displaced stomach and is visible when entering the abdominal cavity at the time of surgery. The rotation compresses the veins, which causes congestion of the stomach and engorgement of the spleen. The position of the spleen varies depending on the degree of volvulus, and it may even rotate on its own pedicle. Counterclockwise rotation of the stomach is possible to a maximum of 90°; the pylorus and antrum move dorsally along the right abdominal wall, and in this case, there is no displacement of the omentum over the ventral surface of the stomach.
The stomach of the cat is more sharply flexed on itself, and the pyloric part reaches little, if at all, into the right half of the abdomen. Gross distention is also less common in cats, which tend to moderate their appetites better than dogs. The cat’s stomach is generally similar to that of the dog; its topography and that of the intestines are shown in the radiographs of Figures 14–13, 14–14, and 14–6). The rugae in the stomach as seen in contrast radiographs are conspicuously fewer and proportionately smaller in cats than in dogs. Pyloric stenosis caused by hypertrophy of the pyloric circular smooth muscle can be encountered in Siamese cats.
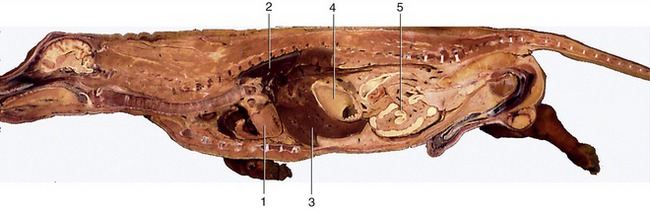
Figure 14–13 Median section of canine trunk, providing overview of viscera. 1, Heart; 2, lung; 3, liver; 4, stomach; 5, intestine.
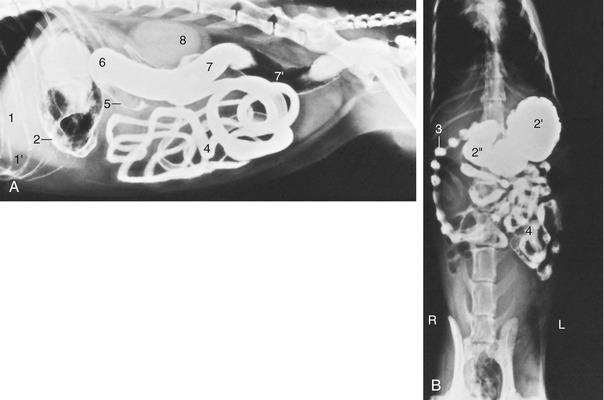
Figure 14–14 Lateral (A) and ventrodorsal (B) radiographic views of the feline abdomen after administration of a barium suspension. 1, Liver; 1′, fat-filled falciform ligament elevating the liver; 2, gas and barium in stomach; 2′, fundus; 2″, pyloric part of stomach; 3, descending duodenum—the striking “string-of-pearls” appearance (characteristic of cats) is due to segmental peristalsis; 4, jejunum; 5, ascending colon; 6, transverse colon; 7, descending colon; 7′, gas in descending colon; 8, kidneys (superimposed).
INTESTINES
(See also pp. 129–135.)
Because the general features of the intestinal tract have been described, it is now appropriate to concentrate on its relationships to other organs and to external landmarks and on its attachments and blood supply.
The small intestine is relatively short, perhaps three or four times the body length. Of this, the duodenum contributes, on average, only 25 cm. The short cranial part of the duodenum passes dorsally and to the right, against the visceral surface of the liver, roughly opposite the ninth intercostal space. It is continued caudally beyond the porta as the descending duodenum, which follows the right abdominal wall to reach a point somewhere between the fourth and sixth lumbar vertebrae (Figure 14–7, B/7′). In its passage it is related dorsally to the right lobe of the pancreas, ventrally to the jejunal mass, and medially to the ascending colon and cecum (Figure 14–15/5). The mesentery of the descending duodenum begins by being relatively long but shortens toward the caudal flexure, where the gut is closely anchored to the abdominal roof. An additional (duodenocolic) fold with a free caudal border attaches the duodenum to the descending mesocolon at this level. The ascending duodenum (Figure 14–15/6), which begins at the caudal flexure, is more tightly tethered than the preceding segment and runs forward, close to the midline, between the descending colon on the left and the root of the mesentery. It turns ventrally at the cranial limit of the root to be continued by the jejunum. Other relations of this part are the medial border of the left kidney dorsally and the jejunal mass ventrally (Figure 14–8, A, and Figure 14–12, B).
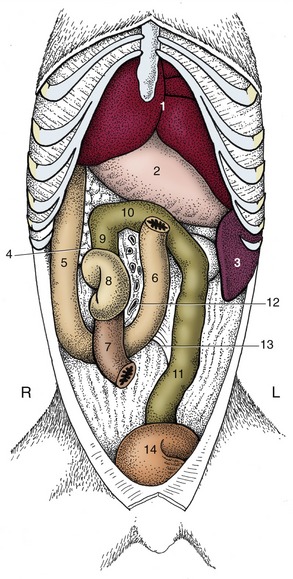
Figure 14–15 The canine duodenum, cecum, and colon in situ; ventral view. 1, Liver; 2, stomach; 3, spleen; 4, pancreas; 5, descending duodenum; 6, ascending duodenum; 7, ileum; 8, cecum; 9, 10, 11, ascending, transverse, and descending colon; 12, vessels in root of mesentery; 13, duodenocolic fold; 14, bladder.
The jejunum and short ileum form a mass occupying the ventral part of the abdomen between the stomach and the bladder (Figures 14–5, 14–6, 14–16, and 14–17). The coils of the jejunum are quite mobile, and at first sight their disposition appears to be haphazard; closer inspection shows that there is some pattern to the arrangement. The mainly sagittal coils of the proximal part lie largely cranial to the more transverse coils of the distal part (Figure 14–8, A). The suspending mesentery is relatively long and imposes little restraint, which allows the gut to slip freely over the floor in response to respiratory and other movements. This feature enables the surgeon to exteriorize much of the jejunum to improve the exposure of more dorsal organs. Dorsally, the jejunal mass extends to the descending duodenum on the right and the kidney and sublumbar muscles on the left. The jejunal coils are generally entirely related to the folded greater omentum ventrally; cranially only the deep leaf intervenes between them and the stomach. The ileum arises at the caudal end of the mass and passes forward and to the right to open into the ascending colon below the first or second lumbar vertebra.
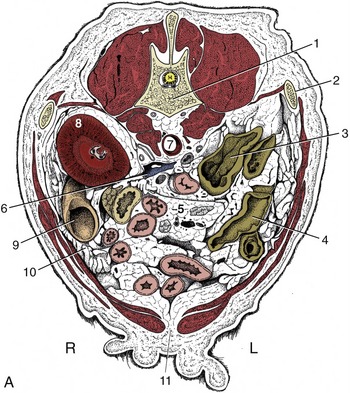
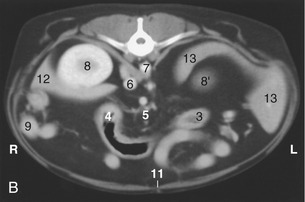
Figure 14–16 A, Transverse section of the canine abdomen at the level of the first lumbar vertebra. B, Corresponding computed tomographic (CT) image slightly more caudal than A; the dog was lying on its back during the CT procedure. 1, First lumbar vertebra; 2, last rib; 3, descending colon; 4, transverse colon; 5, lymph nodes and blood vessels in mesentery; ventral to them is the jejunum; 6, caudal vena cava; 7, aorta, between crura of diaphragm; 8, right kidney; 8′, cranial pole of left kidney; 9, descending duodenum and pancreas; 10, greater omentum; 11, linea alba; 12, liver; 13, spleen.
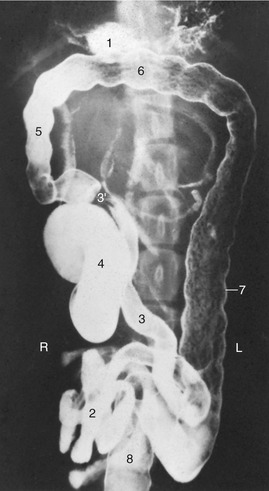
Figure 14–17 Ventrodorsal radiographic view of the canine abdomen after administration of a barium suspension. 1, Residue of barium in stomach; 2, jejunum; 3, ileum; 3′, ileocolic junction; 4, cecum; 5–7, ascending, transverse, and descending colon; 8, rectum.
Small patches of aggregate lymph nodules of varying sizes are present throughout the small intestine; the largest are said to be in the ileum.
In life the intestine is not uniformly full, and at any moment most parts are flattened and molded by the pressures of adjacent viscera. The lumen may be locally obliterated, and when a passage is retained, it is more often than not reduced to a narrow channel along one margin—a “keyhole” section. This explains the narrow streaks that are the common representation of the small intestine in radiographs obtained after the administration of a barium meal. Segmental and peristaltic movements continually alter the configuration in life. After the administration of a contrast medium, the duodenum of the cat often displays segmental contractions that are sufficiently pronounced to divide the gut content into a linear series of globular expansions separated by (more or less) empty regions; this creates the very striking “pearl necklace” effect (Figure 14–14, B). A similar appearance in other regions of the cat’s bowel, or in the duodenum of the dog, is probably evidence of abnormality.
The ileocecocolic junction is peculiar in that the ileum and colon are in line and form a continuous tube that is joined by the cecum to one side. (In the other species it is the cecum and colon that meet end-to-end.) The cecum is short, although it is of varying length, and twisted (Figure 14–15/8 and Figure 14–17/4). It is joined to the ileum by a short (ileocecal) fold and is oriented craniocaudally, although its rounded blind end may finally point in any direction. The cecum communicates with the ascending colon through the cecocolic orifice adjacent to the ileal orifice. The cecum lies to the right of the root of the mesentery and is related to the right kidney dorsally, the descending duodenum and pancreas laterally, and the jejunum ventrally. It lies below the second lumbar joint and thus is broadly level with the most caudal part of the costal arch. The cecum of the cat is small and comma-shaped. Surprisingly, it can be located on palpation by reference to the firm ileocecocolic junction at the level of the fourth lumbar vertebra. The firmness can be mistaken for a tumor or intussusception (Figure 14–19/4).
The colon, 65 cm long on average, is only slightly wider than the small intestine. It is easily recognized by its course cranial to the root of the mesentery and its nearly straight descent on the left toward the pelvis, which it enters dorsal to the bladder (and uterus) (see Figure 3–45 and Figure 14–17). The short ascending part lies to the right, between the descending duodenum and the root of the mesentery, and generally makes contact with the pyloric part of the stomach. Its narrow mesocolon permits it little mobility. The transverse colon runs from right to left, cranial to the root of the mesentery and ventral to the left lobe of the pancreas (see Figure 14–15). It is more loosely attached and sinks within the abdomen; usually it is the lowest part of the colon when depicted in lateral radiographs. The free attachment sometimes allows it to fold on itself to appear as no more than a flexure connecting the ascending with the descending colon. The descending colon is by far the longest segment. It passes caudally, to the left of the mesenteric root, to reach the pelvic cavity, where it continues as the rectum (Figure 14–7, A/6). It is related dorsally to the left kidney and sublumbar muscles and ventrally to the jejunal mass; it may lie against the left abdominal wall (Figures 14–13/4 and 14–18/4). The descending colon is the only segment of the large intestine of the dog that may easily be palpated. No part of the colon lies retroperitoneal.
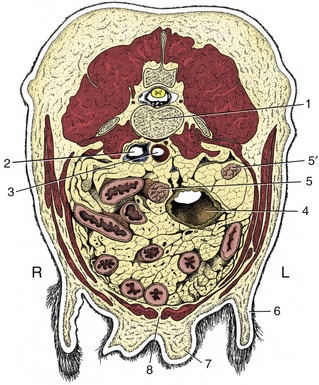
Figure 14–18 Transverse section of the canine abdomen at the level of the fourth or fifth lumbar vertebra. 1, Lumbar vertebra; 2, caudal vena cava; 3, aorta; 4, descending colon; 5, 5′, right and left uterine horns; 6, flank fold; 7, mammary gland; 8, linea alba.
The prominence of the cecum and colon in plain radiographs of the canine abdomen is determined by the amount of gas and the nature and volume of the digestive residues present (see Figure 14–17). The cecum almost always contains sufficient gas to provide a reminder of the twisted course of its lumen. This convenient identifying feature is not found in cats, in which the simpler conformation rarely allows gas to be retained; see Figure 14–19, which also depicts the cat’s colon.
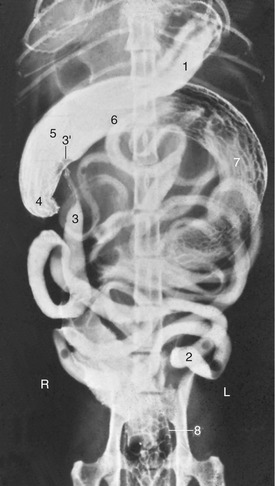
Figure 14–19 Ventrodorsal radiographic view of the feline abdomen after administration of a barium suspension. 1, Residue of barium in stomach; 2, jejunum; 3, ileum; 3′, ileal papilla; 4, cecum; 5–7, colon—the long descending part (7) has curved far to the right in this animal; 8, rectum.
The blood supply of the intestines comes mainly from the cranial and caudal mesenteric arteries; in addition, a part of the duodenum is supplied by the cranial pancreaticoduodenal branch of the gastroduodenal artery (from the celiac artery). The details are shown in Figure 14–20. The descending colon and rectum receive blood from the caudal mesenteric artery that, in both cats and dogs, branches off the aorta near the fifth lumbar vertebrae. The veins form the portal vein, with the exception of those from the caudal rectum that are directed toward the caudal vena cava.
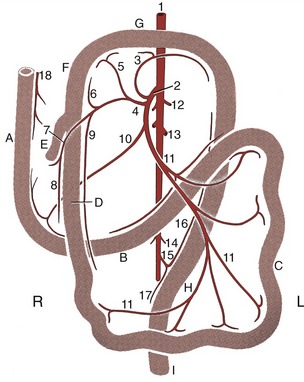
Figure 14–20 The blood supply of the intestinal tract, ventral view; schematic. A, Descending duodenum; B, ascending duodenum; C, jejunum; D, ileum; E, cecum; F, ascending colon; G, transverse colon; H, descending colon; I, rectum. 1, Abdominal aorta; 2, cranial mesenteric a.; 3, middle colic a.; 4, ileocolic a.; 5, right colic a.; 6, colic branch of ileocolic a.; 7, cecal a.; 8, antimesenteric ileal branch; 9, mesenteric ileal branch; 10, caudal pancreaticoduodenal a.; 11, jejunal aa.; 12, phrenicoabdominal aa.; 13, renal aa.; 14, testicular (ovarian) aa.; 15, caudal mesenteric a.; 16, left colic a.; 17, cranial rectal a.; 18, cranial pancreaticoduodenal a.
Several colic lymph nodes lie within the curvature of the ascending and transverse colon. The more prominent jejunal nodes lie high in the root of the mesentery; one, surprisingly large (perhaps 10 cm in the beagle), accompanies the jejunal arteries (Figure 14–16/5). Several smaller caudal mesenteric nodes lie within the descending mesocolon, scattered about the branches of the caudal mesenteric artery.
THE LIVER
The liver (see also pp. 135–139) is relatively large, weighing about 450 g on average, and accounts for 3% to 4% of the body weight. It is almost entirely intrathoracic, occupying a central position with only a slight bias to the right side (Figure 14–7/2 and Figure 14–10). The modest asymmetry is caused by the enlargement of the caudate process beneath the last ribs, where it makes contact with the right kidney (Figure 14–11/9). The ventral border extends across the costal arches and would be palpable were it not for the fat within the falciform ligament and the taut rectus muscles. Even so, it may be appreciated when significantly enlarged. The liver in dogs and cats is deeply divided by fissures extending from the ventral margin; the pattern, the relative extents, and the names of the lobes may be obtained from Figure 3–53.
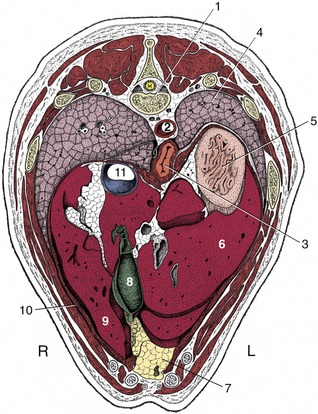
Figure 14–10 Transverse section of the canine trunk at the level of the eleventh thoracic vertebra. 1, Eleventh thoracic vertebra; 2, aorta; 3, esophagus; 4, left lung; 5, fundus of stomach; 6, left lateral lobe of liver; 7, fat-filled falciform ligament; 8, gallbladder; 9, right medial lobe of liver; 10, diaphragm; 11, caudal vena cava.
The cranial surface conforms to the curvature of the diaphragm with which it is in extensive contact and to which it is secured by the caudal vena cava embedded in the dorsal border. The attachment to the tendinous center of the diaphragm is completed by right and left coronary ligaments caudolateral to the vein. Most of the liver can therefore be retracted at operation to expose the diaphragm. The gallbladder is sunk deeply between the lobes, just to the right of the median plane opposite the eighth intercostal space; it usually makes contact with the diaphragm and always appears at the visceral surface, although it is too short to reach the ventral border (Figure 14–10/8).
The visceral surface, though concave, is made irregular by various visceral impressions. The largest of these is made by the body of the stomach to the left of the median plane; the pyloric part and duodenum produce a narrower impression leading away to the right (Figure 14–11/7). The other prominent impression, involving the right lateral lobe and caudate process, is made by the right kidney. Other organs that may touch the liver, especially when the stomach is empty, leave no mark, except the pancreas, which attaches near the porta.
The attachments on the visceral surface are larger but looser and are part (as earlier mentioned) of the lesser omentum. The hepatogastric ligament contains the bile duct, as well as the hepatic artery, the portal vein, and lymphatic vessels and nerves. Once the hepatic ducts receive the cystic duct from the gallbladder, the duct is known as the common bile duct (ductus choledochus). In dogs, it runs from the hilus to the duodenum. Its terminal portion continues for some 2 cm within the duodenal wall before opening by the side of the pancreatic duct on the major duodenal papilla, a small elevation 2 to 3 mm high, caudally directed and located about 3 to 6 cm from the pylorus in both dogs and cats.
Biopsy samples of liver tissue may be obtained by puncture caudal to the xiphoid process; the instrument is directed toward the large left lobe to avoid the gallbladder (see Figure 14–5).
In survey radiographs of the abdomen the liver appears as a large, uniformly dense shadow from which its size, relative to the species norm, may be crudely assessed. When such an assessment is made, it is necessary to be mindful that the liver is more or less completely “intrathoracic” in large, deep-chested breeds, whereas a more appreciable portion projects beyond the costal arch in dogs of less extreme conformation. Dorsal displacement of the liver, away from the abdominal floor, may be encountered in cats that are overindulged; it is due to the deposition of excessive fat within the falciform ligament.
THE PANCREAS
The slender pancreas (see also pp. 139–140) consists of two limbs or lobes that diverge from the vicinity of the pylorus. The left lobe is directed caudomedially and crosses the median plane behind the stomach to end against the left kidney (see Figure 3–56/5). It divides the branches of the celiac artery from those of the cranial mesenteric and is enclosed within the deep leaf of the greater omentum where this passes dorsal to the transverse colon. Its dorsal surface is crossed by the portal vein, where it makes contact with the hilus of the liver to the right of the median plane.
The longer right lobe is directed caudodorsally and follows the dorsal surface of the descending duodenum within the mesoduodenum. It is related dorsally to the visceral surface of the liver and, behind this, to the ventral surface of the kidney (Figure 14–16/9). It lies lateral to the ascending colon and dorsal to the small intestine.
Two secretory ducts open into the duodenum where the two lobes diverge. The smaller and inconstant pancreatic duct joins the bile duct just before this opens on the major duodenal papilla, 3 to 6 cm distal to the pylorus. The accessory pancreatic duct, the main channel, opens on the minor duodenal papilla 3 to 5 cm farther down the gut. Both papillae can be detected with the unaided eye. The duct systems of the two lobes communicate internally. In the cat the main duct is the pancreatic duct; in a minority of cats (around 20%) an accessory duct can also be found. When present, it opens onto the minor duodenal papilla, as in dogs, some 2 cm distal to the major papilla.
The major part of the pancreas is supplied by two of the three branches of the celiac artery; only the caudal part of the right limb receives blood from the cranial mesenteric artery. The left lobe is entered by branches of the splenic artery; branches from the hepatic artery supply the body of the pancreas (gastroduodenal artery) and the cranial half of the right lobe (cranial pancreaticoduodenal artery). Duodenal branches are given off from this vessel and course through the pancreatic tissue to supply the gut itself. Anastomoses between these various vessels occur within the gland. Lymphatics are abundant and drain into the duodenal lymph node if present or into the mesenteric lymph nodes.
One of the most encountered problems in the pancreas of the dog is the presence of an insulin-producing tumor, an insulinoma. Thorough inspection for metastases must be performed in the liver, the duodenum, the mesentery, and the hepatic, splenic, gastric, duodenal, and cranial mesenteric lymph nodes. Resection of the part of the pancreas is difficult because the blood supply is shared with the duodenum and the spleen. Removal of the spleen is indicated when the splenic artery cannot be preserved.
THE ADRENAL GLANDS AND KIDNEYS
The yellowish-white adrenal glands (see also pp. 221–222 and 174–181) (Figure 14–21/7,7′) of the dog are dorsoventrally flattened, about 2 to 3 cm long and 1 cm wide. Each occupies the retroperitoneal space medial to the kidney, cranial to the renal vessels, and dorsolateral to the aorta (the left one) or the caudal vena cava (the right one). The capsule of the right adrenal gland may be continuous with the tunica externa of the vena cava. The right adrenal gland is located ventral to the transverse process of the last thoracic vertebra, with its cranial two thirds covered by the caudate process of the liver. The left adrenal gland, which has a somewhat dorsoventrally flattened oval cranial portion and a cylindrical caudal projection, is positioned ventral to the transverse process of the second lumbar vertebra, just caudal to the origin of the cranial mesenteric artery and adjacent to the origin of the phrenicoabdominal artery. This paired artery courses on the dorsal surface of both left and right glands. The ventral surfaces are crossed and indented by the phrenicoabdominal veins; on the left, this surface is also related to the pancreas.
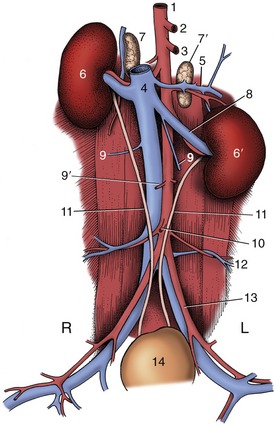
Figure 14–21 The canine urinary organs and adjacent blood vessels in situ. 1, Aorta; 2, celiac a.; 3, cranial mesenteric a.; 4, caudal vena cava; 5, phrenicoabdominal vessels; 6, 6′, right and left kidneys; 7, 7′, right and left adrenal glands; 8, left renal vessels; 9, ovarian vv.; 9′, ovarian aa.; 10, caudal mesenteric a.; 11, ureters; 12, deep circumflex iliac vessels; 13, external iliac vessels; 14, bladder.
The glands are diffusely supplied by branches from adjacent vessels: the aorta and the renal, phrenicoabdominal, lumbar, and cranial mesenteric arteries. The right adrenal vein directly supplies the vena cava, while the left adrenal vein supplies the left renal vein.
The nerve supply is derived from a dense network on the dorsal surface of the glands that appears continuous with the nearby celiac and mesenteric plexuses. The fibers that actually enter the glands are preganglionic and are provided by the splanchnic nerves that enter the abdominal cavity close by.
In cats the adrenal glands are shorter and similar to oval disks. The adrenal glands of older cats are occasionally calcified and are then visible on radiographs. The topography is the same in both species.
THE KIDNEYS
The account of the kidneys presented here concentrates on their positions and relations. Other aspects of their anatomy are considered in Chapter 15.
The kidneys in the dog are bean-shaped and retroperitoneally positioned against the sublumbar muscles. The right kidney is usually said to lie below the first three lumbar vertebrae, and the left one is said to lie below the second to fourth (Figure 14–22); however, this may specify their positions too definitely and they may be found a full vertebral length more caudally. The right kidney is more restricted by being deeply recessed within the liver and is related medially to the right adrenal gland and caudal vena cava, laterally to the last rib and abdominal wall, and ventrally to the liver and pancreas (Figure 14–23). The left kidney is related cranially to the spleen (or stomach when enlarged), medially to the left adrenal gland and aorta, laterally to the abdominal wall, and ventrally to the descending colon.
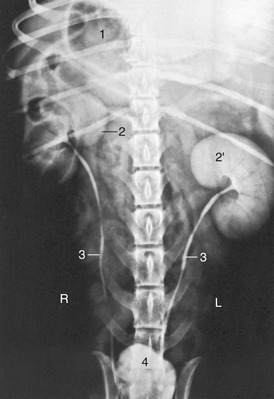
Figure 14–22 Urogram of a dog. 1, Gas in stomach; 2, 2′, right and left kidneys; 3, ureters; 4, bladder.
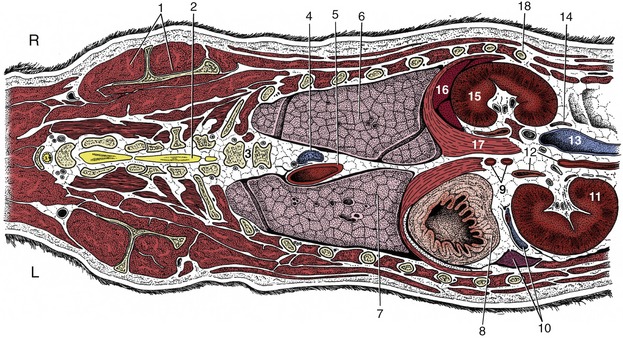
Figure 14–23 Dorsal section of the canine trunk at the level of the kidneys. 1, Supraspinatus and scapula; 2, spinal cord; 3, sixth and seventh thoracic vertebrae; 4, right azygous vein; 5, thoracic aorta; 6, 7, right and left lungs; 8, fundus of stomach; 9, celiac and cranial mesenteric arteries; 10, splenic vessels and spleen; 11, left kidney; 12, left adrenal gland and abdominal aorta; 13, caudal vena cava; 14, right ureter; 15, right kidney (the right adrenal gland is shown medial to the cranial pole); 16, liver; 17, right crus of diaphragm; 18, last rib.
The cat’s kidneys are relatively large and are given a distinctive appearance by capsular veins converging over the surface toward the hilus (Figure 14–24). They are more mobile than the kidneys of the dog (see Figures 14–13 and 14–14), especially the left one, which can be displaced cranially or caudally from its usual position below the second to fifth lumbar vertebrae; it has been taken for a pathological swelling. In cats, both kidneys are readily palpable.
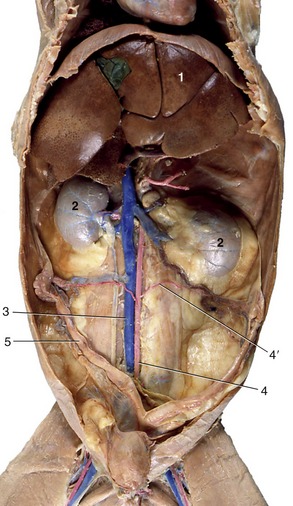
THE MAJOR VESSELS
The abdominal aorta and caudal vena cava run the length of the abdomen, partly recessed between the right and left sublumbar muscles.
The abdominal aorta gives rise to paired dorsal lumbar arteries; the last pair originates from the median sacral artery. Near the second lumbar vertebra in the dog, the caudal phrenic artery and the cranial abdominal artery branch off as a common trunk, which is also the origin of the adrenal arteries. In the cat the caudal phrenic artery originates from the celiac artery as a single artery. The deep iliac circumflex artery branches off the aorta near the sixth vertebra but may split off the external iliac artery in the dog.
The ventral branches of the aorta are the celiac artery, the cranial and caudal mesenteric, the renal arteries, and the ovarian/testicular arteries; sometimes a paired adrenal artery also branches off the aorta. The celiac artery branches off directly after the passage of the aorta through the diaphragm and divides into the hepatic, splenic, and left gastric arteries. The hepatic artery courses to the right of the midline before dividing into three or five branches, which supply the individual liver lobes. After giving off the hepatic branches, the hepatic artery bifurcates into the right gastric and gastroduodenal arteries. The gastroduodenal artery in turn divides into the right gastroepiploic and pancreaticoduodenal arteries.
The cranial mesenteric artery originates one vertebra behind the celiac artery and forms the base of the mesentery. It gives rise to the ileocolic, pancreaticoduodenal, and jejunal arteries in the dog and cat. The renal arteries branch off in the dog ventral to the first and second and in the cat ventral to the third and fourth lumbar vertebrae, and directly caudal to these vessels the genital arteries split off. Ventral to the fifth lumbar vertebrae is the origin of the caudal mesenteric artery, and one to two vertebral bodies more caudal the external iliac arteries split off to supply the hindlimbs. The abdominal aorta terminates opposite the seventh lumbar vertebra by bifurcating into right and left internal iliac and middle sacral arteries (see Figure 14–3). It lies in the furrow formed by the left and right iliopsoas muscles. In both companion species, but especially in the cat, the terminal segment of the aorta is commonly the location of a large thrombus, often known as a “saddle” thrombus from its disposition across the division, which may partially or wholly block the three terminal branches. The origin of the thrombus, the degree of obstruction it causes, and the rate at which it developed determine the severity of the clinical signs, which may include complete paralysis of the hindlimbs.
The portal vein results from the confluence of the cranial mesenteric, caudal mesenteric, and gastrosplenic veins. In dogs, the portal vein is additionally fed by the gastroduodenal vein, which originates from the merger of the right gastric, right gastroepiploic, and cranial pancreaticoduodenal veins. It has been reported that the contributions to the portal vein in cats are variable and cannot be described based on a common pattern.
Venography of the portal vein (Figure 7–44) is occasionally employed to ascertain the existence (and condition) of portosystemic connections. A small intestinal tributary is chosen for the injection. The shunts most commonly revealed connect the portal system with both the caudal caval tributaries at the abdominal roof and the azygous vein within the thorax.
LYMPHATIC STRUCTURES
The lymph nodes of the abdomen can be divided into a parietal and visceral group. The lumbar aortic lymph nodes are, when present, located along the aorta and vena cava. They supply the cisterna chyli or the caudal lumbar aortic nodes. The paired hypogastric lymph nodes are small and are located in the angle of the internal iliac and median sacral artery, ventral to the body of the seventh lumbar vertebra. They receive lymph from the thigh, the pelvic viscera, the tail, and a portion of the lumbar region and have efferent vessels to the cisterna chyli. The sacral lymph nodes are positioned ventral to the body of the sacrum but are often not present. They receive afferent vessels from the adjacent musculature and send off efferent vessels to the hypogastric nodes. The deep inguinal or iliofemoral lymph nodes can be found on the ventral surface of the tendon of the psoas minor at its insertion and receive lymph from the pelvic limb. The medial iliac lymph nodes lie between the deep circumflex iliac and the external iliac artery, ventral to the bodies of the fifth and sixth lumbar vertebrae and can be 4 cm long in the dog. They receive lymph from all parts of the dorsal half of the abdomen, the pelvis, and the pelvic limb, including that from the genital system and the caudal part of the digestive and urinary system. They also receive lymph from the deep and superficial inguinal, the left colic, sacral, and hypogastric lymph nodes and supply the cisterna chyli. The lymph nodes at the breakup of the aorta can be palpated rectally in larger individuals (Figure 14–25/3).
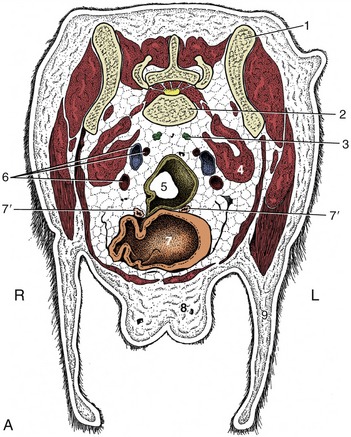
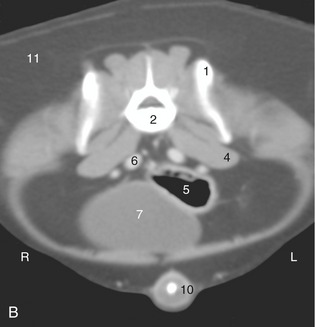
Figure 14–25 A, Transverse section of the canine abdomen at the level of the seventh lumbar vertebra. B, Corresponding computed tomographic image at about the same level. 1, Wing of ilium; 2, seventh lumbar vertebra; 3, sacral lymph nodes; 4, iliopsoas; 5, descending colon; 6, internal iliac artery (most dorsal), external iliac vein, and external iliac artery; 7, bladder; 7′, uterine horns; 8, mammary gland; 9, flank fold; 10, penis with os penis; 11, fat.
The visceral lymph nodes are those directly related to the abdominal organs. The gastric lymph node in the lesser omentum near the pylorus is very small and receives lymph from the esophagus, the stomach, the liver, the diaphragm, the mediastinum, and the peritoneum and sends its efferent vessels to the left hepatic or the splenic lymph nodes. The pancreaticoduodenal lymph node is also a small one and receives lymph from the duodenum, the pancreas, and the omentum and sends its vessels to right hepatic or right colic lymph nodes. The hepatic lymph nodes are situated on each side of the portal vein, at 1 to 2 cm from the hilus of the liver. They receive lymph from the stomach, the duodenum, the pancreas, and of course the liver. The three to five splenic lymph nodes along the course of the splenic artery can be 4 cm long in the dog and receive afferent vessels from the esophagus, the stomach, the pancreas, the spleen, the liver, the omentum, and the diaphragm. The cranial mesenteric lymph nodes are the largest nodes of the abdomen, can be found along the root of the mesojejunum, and receive lymph from the jejunum, the ileum, and the pancreas. The colic lymph nodes in the mesocolon receive afferent vessels from the ileum, the cecum, and colon.
The cisterna chyli is an elongated saccular reservoir receiving lymph from the lumbar and mesenteric lymphatic trunks. The cisterna chyli in the dog is located ventral to the first four lumbar vertebrae and dorsal, at the right side, to the aorta and is related to the crura of the diaphragm. The cisterna chyli in cats has a large saccular part dorsal to the aorta and a plexiform part ventral to the aorta and the last thoracic and first three lumbar vertebrae and is also closely associated with the diaphragmatic crura.
PALPATION
Abdominal palpation is an important diagnostic tool in the examination of companion animals. The system of reference to abdominal regions that is preferred by clinicians divides the abdomen into 18 compartments. Epigastrium, mesogastrium, and hypogastrium are visualised as being defined by two transverse planes: (1) the cranial one situated just caudal to the last rib, (2) the caudal one situated just cranial to the thigh musculature. The depth of the abdomen, between the lumbar muscles and the abdominal floor is then visualized as divided into three, more or less equal, dorsal middle and ventral parts, yielding nine compartments to each side of the median plan. Palpation of these compartments is performed in a systematic way, generally commencing with the dorsal epigastrium, continuing ventrally, and proceeding from superficial (muscle tension, overfilled intestines) to deep. Palpation is usually performed with the subject standing and with the converging extended fingers of the examiner’s hands placed over the flanks. For some purposes it is helpful to have the cranial part of the body raised, which allows the intrathoracic abdominal organs to slide caudally, and for other purposes it is helpful to have the subject laterally recumbent or supine. A one-handed approach, with the converged fingers opposed to the thumb, is useful with cats and small dogs. Whatever the technique, it is important to allay anxiety so that the animal relaxes its abdominal muscles; the recti then form a thick ventral median band, which may be initially disconcerting.
A number of abdominal organs may be identified and assessed by palpation through the abdominal wall. The procedure is most rewarding in cats and small dogs and least rewarding in large, well-muscled or obese dogs. The normal liver projects only slightly, and variably, behind the costal arches and is difficult if not impossible to recognize when the bilateral approach is used. Greater success may be obtained if the fingertips are insinuated deep to the costal arch, which is a liberty permitted only when the flank muscles are fully relaxed. It may then be possible to identify the sharp free margin and narrow adjoining strip of the liver. Homogenous enlargement of the liver can first be palpated in the ventral epigastrium (on superficial palpation); with ongoing enlargement the liver is also palpable in the medial epigastrium, especially when one places the fingers within the costal arch. The liver can be more easily reached at the left than at the right side. The empty stomach is tucked under the ribs, out of reach on the left side, but when full of ingesta or distended with gas, it projects behind the costal cartilages. It is more easily found in narrow, deep-chested dogs than in those in which the trunk is barrel-shaped. The stomach, when empty, does not contact the abdominal wall, but when moderately filled, it lies against it ventrally and to the left. The completely filled stomach, especially in pups, lies largely in contact with the ventral body wall, toward a transverse plane just caudal to the umbilicus. The spleen occupies the same region against the left flank, but because its usual consistency is soft and deformable, it is not easily appreciated unless considerably enlarged and firmed. Normally the spleen is located to the left in the epigastrium, close to the major curvature of the stomach (completely within the costal arch in the dog). In the case of enlargement the spleen moves ventrally and caudally; splenomegaly can be felt in the ventral and medial mesogastrium.
Success in locating the kidneys is rather unpredictable in the dog. Most often, only the caudal pole of the left kidney is within reach, and it may be identified by its firm, rounded contours. The right kidney is commonly inaccessible. In some dogs, generally of the larger breeds, the left kidney is pendulous and “floats” at a more ventral level than usual; this is the normal condition in the cat, in which both kidneys can generally be found and may be steadied through the abdominal wall for biopsy puncture. The entire surface of a “floating” kidney, including the depression at the dorsally facing hilus, may be examined. The left kidney contacts the dorsal part of the left lateral abdominal wall.
The fluctuating intestinal mass occupies a large part of the abdomen, extending from the roof to the floor and from one flank to the other. Identification of most individual parts is problematical. The descending duodenum may sometimes be identified on the right side if the fingers are first pressed against the abdominal roof and then drawn laterally. There is no difficulty in finding the jejunum, whose coils may be made to slip between the hands. In the dog the only part of the large intestine that may be sought with confidence is the descending colon on the left side. It is most readily identified when occupied by a column of hard or granular feces. The ascending colon and cecum may sometimes be identified, most readily when gas-distended, but the transverse colon is too deeply tucked under the ribs to be within reach. All parts of the large intestine are more readily found in cats, in which a useful guide to the positions of the cecum and ascending colon is provided by the firmness at the ileocecocolic junction. The lymph nodes associated with the intestine evade detection unless enlarged.