Chapter 14 Phytochemical variation within a species
As was indicated in Chapter 3, the species represents the unit of plant classification; it may constitute a homogeneous taxon of plants with little variation from one specimen to another or it may include various varieties or races which each have some distinctive feature(s). Thus with Datura stramonium four varieties are described: var. stramonium—white flowers, thorny capsules; var. inermis—white flowers, bald capsules; var. tatula—lilac flowers, thorny capsules and var. godronii—lilac flowers, bald capsules. Often such varieties represent single gene mutations (in the above case two independent genes are involved) and are morphologically recognizable. In other instances the mutation gives rise to a variant having a different secondary metabolite profile—not necessarily discernible in the morphological form; these are termed chemical races or chemodemes. The mutation may involve the presence or absence of a single component or, if acting at an early stage of the biosynthetic route, may involve a whole series of compounds.
Knowledge of the existence of such chemical races is not new and A. Tschirch (1856–1939) in his Handbuch der Pharmakognosie deals with the various ‘physiological forms’ of Vanilla planifolia, Cinchona spp., fennel, Piper nigrum and the balsam-containing trees. Also, 60 years ago A. R. Penfold and co-workers were describing chemical races of various Australian oil-producing species of Leptospermum, Boronia and Eucalyptus.
In addition to the above which involve submicroscopic point mutations associated with an alteration in the DNA chromosomal material, there are other genetic variations which may affect chemical constituents of the species. These include (a), the existence of polyploids in which chromosome sets become doubled, tripled, quadrupled etc., (b), the addition of one or a few chromosomes above the normal complement (extrachromosomal types), (c), gross structural changes to a chromosome, (d), artificially produced transgenic plants.
Two consequences of the above are, on the positive side, it provides the possibility for the selection and establishment of superior strains with respect to the chemical constituents as described in this chapter. The negative aspect concerns the arrival on the drug market of material which is low in active constituents. This, together with other factors, necessitates the need for strict quality control (Chapter 16) which is being vigorously addressed by various legislative bodies (Chapter 2). Transgenic crops present their own problems.
CHEMICAL RACES, CHEMOTYPES, CHEMODEMES
The plant kingdom has been subjected to an extensive, but not exhaustive, chemical investigation. Thousands of samples have been screened for substances of medicinal value or for suitable precursors of therapeutically active compounds. Many other plants have been studied chemically from the viewpoints of manural treatments, plant resistance and biosynthesis of active constituents. From such observations has emerged evidence for the existence of ‘chemical races’, ‘chemotypes’ or ‘chemodemes’. These are defined as chemically distinct populations within a species and have similar phenotypes but different genotypes and as such are identical in external appearance but differ in their chemical constituents.
Before the existence of a chemical race can be established, certain fundamental observations are necessary. A chemical analysis of a number of random samples of a particular species may show a variation between the samples but would be insufficient to demonstrate any genetical differences, since factors such as age, climate and soil can all exert profound effects on the result of the ultimate analysis. Samples of seed, or clones from different plants, must be raised together under uniform conditions, and to exclude hybrids, which do not breed true, cultivation for a number of generations is desirable. It may then be possible to demonstrate that differences occur in either the nature or quantity of a particular constituent and that these differences are of a hereditary nature.
Such observations necessitate numerous assays and precise horticultural work, to which must be added the difficulties of dealing with plants which may take years to mature. Furthermore, many of the more important vegetable drugs cannot be successfully cultivated in temperate climates, so that it is not surprising that the number of medicinal plants fully investigated under ideal conditions is still limited. However, with the world-wide increase in genetic studies on medicinal plants, progress is being made towards the isolation of those enzymes associated with the existence of specific chemical races; recently, for example, the cloning of an enzyme involved in ginkolide biosynthesis (K. SangMin, Phytochemistry, 2006, 67, 1435).
The clinical significance of chemical races is illustrated by Valerian; the plant normally contains both volatile oil and iridoid compounds, the latter with reported cytotoxic activity. As the sedative properties of the drug are ascribable to the valerenic acid and valerone constituents the cultivation of chemical races lacking the iridoids was introduced.
Fixed oils
Agriculturally, the cultivation of seed oil plants is second only in importance to that of cereals. Most of the fixed oil produced is used by the food industry but there are also important industrial and other, including pharmaceutical, uses. It is not surprising therefore that sustained breeding programmes for the improvement of yields and quality of oil have been in progress over many years. Normal rapeseed oil contains, as an acylglycerol, 20–40% of erucic acid, an acid having an extra long carbon chain (C22) and one double bond. Its presence in quantity renders the oil unsuitable for edible purposes but varieties are now extensively grown which contain no erucic acid. The value of the crop has been further enhanced by coupling low erucic acid content with one giving low glucosinolates in the protein meal thus improving the animal feed properties. However, erucic acid is industrially important for the manufacture of lubricants, artificial fibres and plasticizers, so that varieties of rape developed for their high erucic acid content are also important agricultural crops.
The production of oil from sunflower seed has been improved by varieties that yield linoleic acid-enriched oil and which are more convenient for harvesting by having a large single flower head and no side-shoots. Groundnuts, the source of Arachis Oil BP, exist as various strains with different relative proportions of fatty acids.
Safflower constitutes an important oil-seed crop and its genetic variability has facilitated the breeding of varieties with widely differing oil constitutions. High oleic varieties are used for oil for human consumption and high linoleic varieties are important for oils used as industrial coatings and lubricants.
The above examples involve plants with a short life-span so that breeding by classical methods is a relatively rapid procedure. However, this is not so with plants such as the coconut palm, olive and cocoa so that in these cases modern techniques involving gene transfer would have an obvious advantage for the introduction of new or modified oil characteristics.
Cyanogenetic glycosides
A well-known chemical race in the cyanogenetic series is the almond. There are many varieties of Prunus communis showing different morphological forms, with and without amygdalin, but some varieties have similar characters and differ only in the presence or absence of the glycoside. Also in this group the clovers, especially Trifolium repens, have been extensively studied and Linaria has been shown by Dillemann to produce a chemical race by introgressive hybridization. L. striata contains cyanogenetic glycosides, and, crossed with the non-active L. vulgaris, gives rise to hybrids which, on repeated back-crossing with L. vulgaris, give some plants difficult to distinguish from L. vulgaris but which contain the cyanogenetic principles of L. striata.
Alkaloids
The Duboisia species form an important commercial source of the tropane alkaloids and have been extensively studied by Australian workers. With both D. myoporoides and D. leichhardtii, trees from natural stands in various locations were examined and their progeny were raised side by side in experimental plantations. The trees produce hyoscine, hyoscyamine, norhyoscyamine, tigloidine and valeroidine, and the proportion of any one alkaloid to total alkaloid may vary greatly. It was shown not only that seasonal and environmental factors are involved in this variation, but also that within a species there exists a wide range of alkaloid genotypes. Other varieties containing nicotine and nornicotine were also reported. Interspecific hybrids between the two species were studied and four hybrid clones were selected for possible exploitation as high alkaloid yielding strains. Thus, in this genus we have the possibility of two distinct types of chemical race—different alkaloid types within a species and different alkaloid types among hybrid phenotypes.
An example of the improvement of the morphine content of opium poppies by genealogical selection is furnished by the work of Lecat. The original seed gave capsules having an average morphine content of 0.385%. From this heterogeneous population were selected six individuals whose capsules analysed about 0.7% morphine. The seeds of these plants formed the heads of the lines cultivated in successive years, during which the best plants were collected and all those containing less than 0.7% morphine were rejected. The harvest of 1955 gave capsules with an average morphine content of 0.765%, thus doubling the original morphine content of the population. Such a method of breeding does not produce a race of plants surpassing individual morphine contents from the original heterogeneous population; it merely produces a homogeneous race of the alkaloid-rich plants.
Phillipson and colleagues reported at least three different chemical races of Papaver fugax and P. armeniacum in which either (1) 1-benzyltetrahydroquinoline, proaporphine, aporphine, (2) morphinane or (3) rhoeadine types are the major alkaloids; there are at least two different chemical strains of P. tauricola containing either the first or third types of the above. Three different isoquinoline alkaloid chemotypes of Thalictrum minus have been reported from Bulgaria. Papaver bracteatum is a species exhibiting races with respect to thebaine.
From Claviceps purpurea a number of races have been isolated containing different groups of ergot alkaloids and these have obvious implications for the commercial production of alkaloids.
One fodder crop in which the presence of alkaloids is undesirable is lupin seed. Ordinary wild forms are bitter and contain alkaloids of the lupinane series but over the years a number of sweet forms have been developed for commercial purposes in Europe. The strains depend for their low alkaloid content on the presence of a particular recessive gene. However, as a number of such genes exist, cross-fertilization between two different sweet strains will again give bitter progeny. To avoid this happening, considerable care is necessary in regions where different strains are grown side by side. Other plants for which there is evidence of alkaloid varieties include Ephedra distachya and the Lycopodium species.
Chemical races appear to be lacking in the pharmaceutically important indole alkaloid-containing genera Strychnos, Rauwolfia and Catharanthus.
Anthraquinones
The purgative anthraquinone drugs owe their activity to complex mixtures of the 1,8-dihydroxy derivatives of anthranols, their glycosides and free anthraquinones. The relative proportions of the constituents of the mixture, which greatly influence the pharmacological activity, depend not only on time of collection, age of plant, drying conditions and geographical source, but also on genetical factors. In a programme involving Rheum palmatum, van Os produced races varying in their rhein/chrysophanol ratio and other hereditary strains for high- and low-yielding total anthraquinones. The analysis of individual Cassia angustifolia plants has indicated that selection of individuals for high sennoside B-yielding strains is a possibility.
Cardiac glycosides
With Digitalis purpurea the property of high glycoside content is hereditary. The proportion of glycosides derived from digitoxin and gitoxin is also very different in plants of different origin and remains so during subsequent cultivation under standardized conditions. The strains were distinguished chemically as digipurpurin, strospeside and digitoxin types. It now remains to prove that these characters are independent of the phenotype (i.e. that they are not inseparably associated with other characters of the parent plant). One race, ‘Cambridge’, which is relatively rich in digitoxin, is easy to distinguish; the other digitoxin race found in the Vosges differs little from the other selections. Variation in the proportion and quantity of glycosides in D. lanata has also been noted in mixed populations and, by the selfing of selected individuals, strains rich in a particular glycoside have been produced, the inherited character being strongly developed. Valuable physiological forms could thus be produced and Ligeti has recommended that these strains be designated by such names as ‘D. lanata Ehrh, chemo-varieties A and C’, depending on the respective predominance of lanatosides A and C. It appears that with such in-bred lines continuous selection is still required to prevent reversion to the normal character level of the species.
The great value of the radioimmunoassay (q.v.) for the rapid selection of high-yielding strains of Digitalis lanata has been demonstrated by Weiler and Westekemper. After two selection steps involving the analysis of over 10 000 individual plants, the average digoxin content of the plants could be raised two to threefold and several strains with average digoxin concentrations in the leaf of 0.6% were isolated. Individual plants were found with 0.9–1.0% digoxin content. As is usual with this type of selection, no plants better than the few best of the original selection were obtained.
Following intensive chemical investigation of the genus Strophanthus, Reichstein and his colleagues differentiated four chemical varieties of the polymorphous S. sarmentosus from different geographical sources. They are sarverogenin-, sarmentogenin- and sarmutogenin-producing types with glycosides of these, and a fourth form which has a low glycosidal content (Fig. 14.1). Although the locality of growth may produce quantitative differences in the constituents of the various races, the overall type is genetically controlled. Similar variation may exist among those plants that yield steroidal saponins, several thousand of which, from different localities have been screened for their sapogenin content.
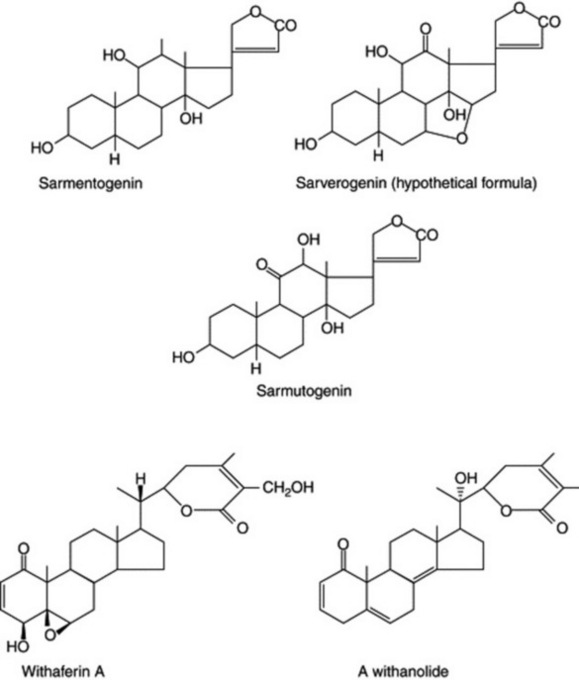
Withanolides
The plant Withania somnifera (Solanaceae), in addition to producing alkaloids, contains steroidal lactones. Investigations over the years, carried out on various sources of plant material, and concerning the non-alkaloidal constituents, had given differing results, which were explained by the work of Abraham et al. (1968) on Israeli plants. Three chemotypes were discovered among 24 populations of W. somnifera collected in various parts of the country. Chemotype I contained predominantly withaferin A (0.2% of the dry weight), which is the principle responsible for the plant’s bacteriostatic and antitumor properties. Chemotype II contains a compound of similar structure, and chemotype III a mixture of related compounds comprising a group of steroidal lactones—the withanolides (Fig. 14.1). The only morphological difference observed between the chemotypes was a difference in flowering time (12 days early) for chemotype III. Since then, other chemotypes of W. somnifera have been reported from India and South Africa.
Steroidal alkaloids
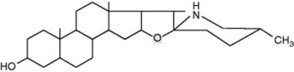
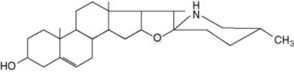
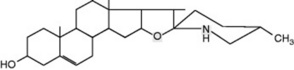
Solanum spp. (Solanaceae) contain steroidal glycosidic alkaloids some of which have been investigated as potential intermediates in corticosteroid synthesis. In S. dulcamara (the woody nightshade) Sander has distinguished a west European tomatidenol group and an east European soladulcidine-solasodine group (Table 14.1). Although polyploid forms do occur in the genus, these chemical varieties all had 2n = 24 chromosomes and were genetically stable. Subsequent work demonstrated that the different chemotypes can occur in the same locality. With the commercial species, about 3500 individual 6-month-old Solanum laciniatum and S. aviculare were analysed by radioimmunoassay (q.v.) and found to contain average leaf concentrations of 1.6–1.7% solasodine; from these a few individuals were selected for future breeding work.
Table 14.1 Chemical races of Solanum dulcamara.
| Aglycone | Sugars | Glycoside |
|---|---|---|
| Soladulicidine (25D) | Galatose (1 mol) | Soladulcidine-tetraoside |
| Glucose (2 mol) | ||
| Xylose (1 mol) | ||
 |
||
| Solasodine (25D) | Galactose (1 mol) | |
| Glucose (1 mol) | Solasonine | |
| Rhamnose (1 mol) | ||
 |
||
| Δ5-Tomatidenol (25L) | Galactose (1 mol) | |
| Glucose (1 mol) | α-Solamarine | |
| Rhamnose (1 mol) | ||
 |
||
Essential oils
The biochemical group of plants offering evidence of the largest number of chemical races is that containing volatile oils. Here, again, many of the differences within a species which have been reported may be due to factors other than genetic ones. Australia offers unique opportunities for the investigation of this problem as the flora is rich in oil-bearing plants. As an example, the common form of Eucalyptus dives contains piperitone as the chief constituent of the oil, but other races are known which produce principally phellandrene or cineole, while still others produce oils intermediate in composition.
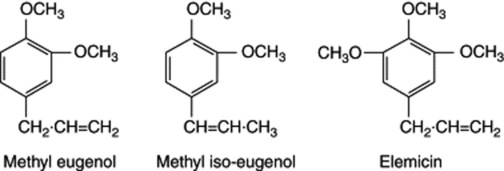
There are three races of Melaleuca bracteata producing volatile oils containing chiefly methyl eugenol, methyl iso-eugenol and elemicin, respectively. They can be transformed one into the other by simple chemical steps, which suggests that one of the compounds (e.g. methyl eugenol) occurs in all the races. With the appropriate enzyme, methyl iso-eugenol could be formed by a simple double-bond shift and elemicin by the addition of a hydroxyl group and subsequent methylation. Because of this a one-gene-one-enzyme hypothesis suggests itself and it would be possible to test this by breeding experiments.
In the American turpentine industry, breeding investigations have shown that oleoresin yields in pines are inherited. Two chemotypes differing in their Δ3-carene content of the oil have been recognized for Pinus sylvestris.
Plants of the Labiatae have long been cultivated for their volatile oils and many varieties of a single species may exist; these are not, however, necessarily true chemical races because morphological differences may also be involved. Chemical races in the genera Ocimum, Melissa, Micromeria and Thymus have been studied. As one example, Rovesti’s observations on the Ethiopian plant Ocimum menthaefolium are shown in Table 14.2. These forms occurred at different altitudes and exhibited a correlation between humidity of atmosphere and the constituents but cultivation of all four types at Asmara showed the chemical races to be stable and not transitory phenotypes varying with the environment. Similar wide variations of constituents of Melissa officinalis have been noted. Sixteen genotypes cultivated in the same field in former Yugoslavia for 2 years contained 0.046–0.246% oil. The constituents were the same for all oils but there was wide variation in relative amounts between the genotypes, namely citronellal 2.711–12.141%, linalool 1.501–6.380%, caryophyllene 1.210–19.073%, geranyl acetate + citronellol 9.710–26.913; (through Chem. Abs., 1991, 114, 58920). In Spain a 5-year selection and improvement programme for M. officinalis raised the essential oil content from 0.2–0.3% to more than 0.5% (T. Adzet et al., Planta Med., 1992, 58, 558).
Table 14.2 Oil composition of Ocimum menthaefolium.
| Race | Oil content | Composition of oil (%) |
|---|---|---|
| Var. camphorata | 0.285 | Camphor 23, cineole 13, pinene 10, estragol 40, sesquiterpene 10 |
| Var. estragolata | 0.248 | Estragol 73, linalool 9, anethole 5, sesquiterpene 5, limonene 1.8 |
| Var. anisata | 0.236 | Anethole 39, estragol 31, limonene 10, linalool 9, sesquiterpene 1.1 |
| Var. citrata | 0.212 | Citral 56, estragol 20, terpene alcohol 10, terpene (as limonene) 9, sesquiterpene 5 |
| Var. intermedia (between camphorata and estragolata) | 0.267 | Camphor 7, cineole 3, estragol 58 |
Chemotypes of Acorus calamus, sweet flag (Acoraceae), having differences in essential oil composition, have been DNA profiled.
Cinnamomum camphora (Lauraceae) exists as various chemical races which vary in their volatile oil composition, and a similar situation prevails for C. zeylanicum (C. verum). In the same family Ocotea pretissa gives oils of the sassafras type which may or may not contain camphor.
Chemical races appear to occur very widely in some Compositae. For Tanacetum vulgare (Tansy) ten different chemotypes have been reported for Finland with others for Piedmont (Italy) and Central Europe. Some principal components of these forms are thujone, isothujone, camphor, chrysanthenyl acetate and sabinene. Achillea millefolium (Yarrow) occurs as various chemical races and Artemisia dracunculus (Tarragon) has yielded, from plants originating mainly in France, an oil containing estragol, whereas from plants of Germany and Russia, sabinene, elemicin and trans-isoelemicin are the principal components. Wormwood BP/EP (A. absinthium) has a number of chemotypes with respect to its volatile oil content which may contain over 40% of any one of p-thujone, trans-sabinyl acetate cis-epoxyocimene or chrysanthenyl acetate. A clone of Artemisia annua giving a high yield of the important antimalarial artemisinin has been recorded (D. C. Jain et al., Phytochemistry, 1996, 43, 1993).
Further examples of chemical races among volatile oil-containing drugs of the Labiatae can be found in Chapter 22.
Miscellaneous
Other groups of active compounds which exist as chemical races are the phloroglucinol derivatives of Dryopteris, cannabinoids in cannabis, the bitter principles (e.g. amaragentin, sugars and volatile oil) of Gentiana lutea and the glycosides of Salix. The existence of two discrete chemotypes of Equisetum arvense with respect to flavonoid content is noted in the British Herbal Compendium Vol. 1. Artemisia annua plants raised in Holland from seeds obtained from a number of countries gave plants exhibiting distinct geographic chemotypes (T. E. Wallaart et al., Planta Medica, 2000, 66, 57).
These examples serve to show that the occurrence of chemical races in plants, whether they be of natural origin or produced by plant breeding, can offer considerable scope for the improvement of the therapeutic value of the drug either by adjustment of the individual constituents or by increase in the overall yield.
CHANGES IN CHROMOSOME NUMBER
Polyploidy
In some organisms the chromosomes can be grouped, not in pairs but in threes, fours or higher numbers; these are polyploid individuals—triploid, tetraploid, octaploid, etc. Such polyploids can be derived by the multiplication of the chromosomes of a single species (autoploids) or as a result of the multiplication of the chromosomes following hybridization between two species (alloploids). The latter case furnishes a mechanism whereby a hybrid of two species, itself infertile, may give rise to a constant fertile type by polyploid formation. In the new polyploid form, pairing of the chromosomes at meiosis is possible which was probably not so in the original hybrid. Types such as this arise naturally: Primula kewensis formed as above from the infertile hybrid P. verticillata × P. floribunda was first recorded in 1912; since then many new species have been synthesized as well as some Linnean species (e.g. the production of the hemp nettle, Galeopsis tetrahit Linn. from G. pubescens × G. speciosa). Of pharmacognostical interest is the stabilization of the (fertile) F1 hybrid of Datura ferox × D. stramonium by polyploid formation as indicated later in this chapter. In some species the somatic chromosome number varies irregularly within wide limits (e.g. the blue grass Poa pratensis from 20 to over 100). Such plants, termed aneuploids, do not breed true and often exhibit apogametic (asexual) reproduction.
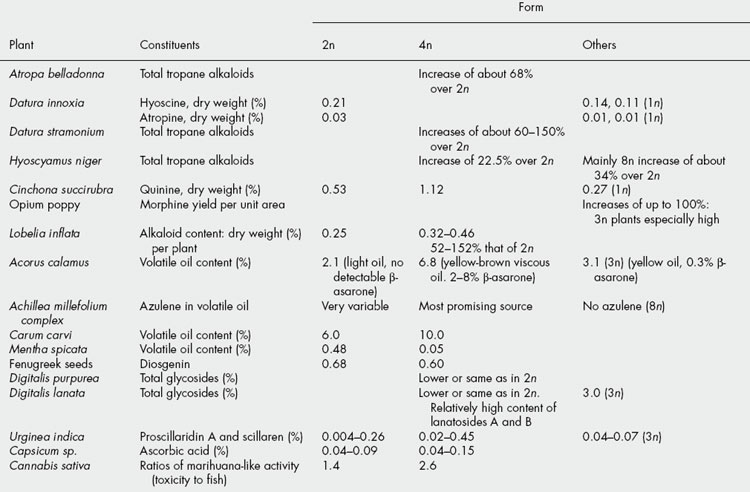
Among the natural polyploids of medicinal interest may be mentioned the mints and valerian; many of the former are alloploids. Valerian occurs across Europe in a variety of forms (2n, 4n and 8n); it is possible that the wide variability in pharmacological action of different samples of Valerianae Radix is associated with these different forms and a more rigid definition of the botanical source might be desirable. As indicated in Table 14.3, the oil compostion of Acorus calamus, the sweet flag, varies with the ploidy. For pharmaceutical purposes the 2n variety, containing no detectable toxic beta;-asarone is preferable.
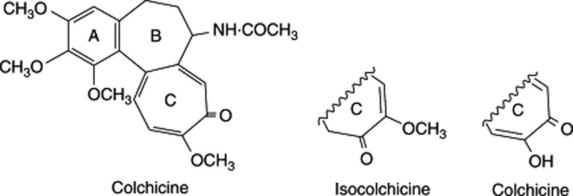
Polyploidy can be artificially induced in many plants by suitable treatment with the alkaloid colchicine. The cytological effect of colchicine on dividing cells was reported by Dustin, Havas and Lits in 1937 and in the same year used practically by Blakeslee in his Datura studies. In the presence of colchicine, chromosomes in a cell undergoing mitosis will continue to divide without the formation of a mitotic spindle figure. Sister cells therefore are not formed, and in the growing root tips of onion (2n = 16), a 72-h treatment with colchicine solution has given rise to cells containing as many as 256 chromosomes. This ‘C-mitotic’ activity of colchicine may arise from its interaction with the disulphide bonds of the spindle protein and by inhibition of the conversion of globular proteins to fibrous proteins. On cessation of treatment, the spindle figure again forms in the normal way.
C-mitotic activity is greatly influenced by modifications of the colchicine molecule. Thus, colchicine is 100 times more active than its isomer isocolchicine and colchiceine is virtually inactive.
Modification of substituents in other rings may not have such a marked effect on activity—colcemid, which possesses a methylamino substituent in place of the acetylamido group of colchicine, is reported to have effects the same as colchicine but with toxicity to animal cells.
Plant materials can be treated with colchicine in a number of ways. Seeds are frequently soaked in an aqueous solution of colchicine (0.2–2.0% solution for 1–4 days) before planting, and seedlings can be inverted onto filter paper soaked in the solution so that the growing points are not damaged. Alternatively, the soil around the roots of young seedlings can be moistened with the alkaloid solution. Young buds and shoots can be treated by immersion, and lanolin pastes and agar gels are useful for general application to tissues.
Newly formed polyploids usually require a number of generations to stabilize themselves and treatments of the above type often fail to give a uniform plant regarding chromosome number; such mixochimeric conditions may involve different chromosome numbers in the three germ layers of the plant.
Typical effects of polyploidy compared with the diploid state are larger flowers, pollen grains and stomata. The influence of polyploidy on the constituents of a number of drug plants is indicated in Table 14.3; some figures quoted are taken from extensive studies and are given as an approximate indication of the differences obtained. As can be seen, the effects of polyploidy are not generally predictable and each species must be examined individually. Care must be taken that the method used to express the results does not give a deceptive effect. Thus, with lobelia, tetraploid plants are smaller than diploid ones, so that, in spite of an increased percentage of alkaloid in 4n plants expressed on a dry weight basis, the total alkaloid content per plant may not exceed that of the 2n plants. A similar situation exists with tetraploid Artemisia annua with respect to artemisinin content (T. Wallaart et al., Planta Medica, 1999, 65, 728). With tetraploid caraway plants, notwithstanding a 13% smaller crop of fruits from one plant, the total volatile oil content of this plant was increased by 100%; the 4n caraway was also found to be perennial (2n = biennial) and to possess an increased frost resistance.
Berkov reported (Pharm. Biol., 2001, 39, 329) that for autotetraploids of Datura innoxia, D. stramonium and Hyoscyamus niger, the 4n seeds contained, respectively, 1.8, 1.65 and 1.96 times the alkaloid content of the 2n seeds. Also (S. Berkov and S. Philipov, Pharm. Biol., 2002, 40, 617), for D. stramonium roots, concentrations of the principal alkaloids together with the 13 minor ones were higher in the 4n, compared with the 2n, roots.
In some species polyploidy does not affect the relative proportions of the individual constituents—for example, solanaceous herbs produce increased quantities of tropane alkaloids in the 4n state and reduced amounts as haploids but the proportion of hyoscine to hyoscyamine remains unaltered; the proportion of carvone in oil of caraway derived from 4n plants is also unchanged. However, 4n Digitalis lanata is reported to contain a relatively high proportion of lanatosides A and B compared with the 2n form. Haploid D. lanata plants raised from androgenic cell cultures are reportedly smaller than the 2n form, have some morphologically abnormal flowers and show very variable cardenolide contents (B. Diettrich et al., Planta Medica, 2000, 66, 237). The sesquiterpene lactones of Ambrosia dumosa, family Compositae, exhibit marked differences between the diploid and polyploid forms.
V. Lebot and J. Levesque (Phytochemistry, 1996, 43, 397) record that some 100 tons of kava root (Piper methysticum) (q. v.) are imported annually into Europe. The plants are all sterile decaploids (2n = 10x = 130) and are raised by smallholders throughout the Pacific Islands. Unfortunately the yields of kavalactones from different sources vary enormously and there is a real need for clonal selection for genetic improvement. The above authors have examined by HPLC the chemical composition of 121 cultivars originating from 51 Pacific Islands.
Extrachromosomal types
Sometimes plants occur with one or more chromosomes extra to the somatic number and these are known as extrachromosomal types. They were first noticed by Blakeslee’s group in 1915, although their genetic constitution was not immediately apparent, when they sporadically appeared in pure line cultures of Datura stramonium. Such plants were later shown to possess 25 chromosomes in the somatic cell and with Datura (n = 12), twelve 2n + 1 types are possible, each one containing a different extra chromosome. The chromosomes were designated by numbering their halves (or ends), so that the largest chromosome is 1.2 and the smallest 23.24. All 12 types eventually appeared in Blakeslee’s cultures and were originally named according to some obvious characteristic of the plant (e.g. Globe, Rolled, Ilex, etc.) although the end-numbering system can also be used to identify them; thus, Globe = 2n + 21.22. Other 2n + 1 types are also produced and are termed secondaries, tertiaries and compensating. Secondary types have the extra chromosome made up of two identical halves of a chromosome (e.g. 2n + 1.1) and in tertiary types it is composed of two halves of different chromosomes. Compensating types lack one of the normal chromosomes, which is compensated for by two others each carrying a different half of the missing one (e.g. 2n − 1.2 + 1.9 + 2.5). At meiosis 2n + 1 types produce a mixture of n and n + 1 gametes and so do not breed true; they proved particularly useful to geneticists for gene location.
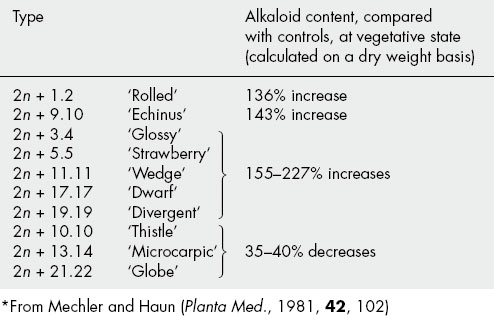
In 1963, Stary reported the analysis of the primary types Poinsettia (2n + 17.18) and Globe (2n + 21.22) and showed them to possess more hyoscine than hyoscyamine in the leaves of mature plants, whereas in diploid strains the reverse is true. Mechler and Haun have reported on the total alkaloid content of other 2n + 1 types, including some secondary types. Their abstracted results are given in Table 14.4.
ARTIFICIAL PRODUCTION OF MUTATIONS
The mutagenic properties of X-rays and radium emissions were exploited as early as 1921 by Blakeslee at the commencement of his classical studies on the genetics of the genus Datura. Since then, all types of ionizing radiation (α-particles, β-rays, γ-rays, thermal and fast neutrons) have been extensively studied in this respect and a number of new varieties of crop plants have been produced (barley, peas, soya beans, mustard and rape). In barley approximately one-fifth of all viable mutations produced by ionizing radiations are of the ‘erectoides’ (dense spike, stiff straw) type; in other crops increased yields, early maturity and mildew resistance have been achieved.
Subsequent to a small number of investigations on chemical mutagens dating from 1910, the avalanche of research on this subject started during and after World War II, following observations by Auerbach and Robson on the production of mutations in Drosophila (fruit fly) by mustard gas. Further stimulus was given to this work by the discovery that many chemical mutagens also possess carcinogenic or anticarcinogenic properties. These substances vary enormously in their molecular complexity and chemical properties, and it is only more recently that their mode of action has been elucidated.
Collectively, ionizing radiations and chemicals will produce a mutation spectrum which covers all of the groups listed earlier. The former, however, produce in the chromosomes aberrations of a more random nature than do chemicals, which often act principally at certain loci—particularly at those areas of the chromosome which stain differently at mitosis (heterochromatin). Also, the distribution of effects between nuclei is more random with X-rays than with chemicals.
Mutagenic agents act at various stages of nuclear organization. Thus, at that stage of the interphase (non-dividing) nucleus when DNA synthesis is taking place, aberrations involving chromatid exchanges and isochromatid breaks occur. These effects do not become immediately (0–8 h) manifest in the cell but appear as delayed effects 8–48 h after treatment. Ionizing radiations and most chemicals produce aberrations of this type. Clearly, breaks which occur in the interphase nucleus chromosomes before DNA synthesis occurs (chromosomes unsplit) would be of the chromosome type and these are induced by X-ray treatment and by a few chemicals (e.g. ethoxycaffeine and streptonigrin). Other mutations may be induced during the DNA post-synthetic stage of the interphase nucleus and during mitosis itself—as in the production of polyploids by colchicine and in the inducement of binucleate or polynucleate conditions due to inhibition of cell plate formation by cyclic organic compounds (e.g. halogenated derivatives of benzene and toluene, hydrazinotropone compounds, aminopyrine). Most mutagens produce more than one type of fragmentation or exchange effect (e.g. ethoxycaffeine and streptonigrin, besides producing chromosome exchanges, also induce sub-chromatid exchanges). A few of the many known mutagens are given in Table 14.5.
Table 14.5 Cytological action of some chemical mutagens.
| Group | Examples | Cytological effect |
|---|---|---|
| DNA precursors and related compounds | Adenine | Inhibition of early stages of DNA synthesis, purine formation |
| Deoxyadenosine | Inhibition of deoxyribonucleotide synthesis | |
| Ethoxycaffeine | Complex formation with inhibition of the DNA-polymerase reaction, degradation and denaturation of DNA | |
| DNA base analogues | Thymidine analogue 5-bromodeoxyuridine | Production of abnormal DNA |
| Antibiotics | Streptomycin | As ethoxycaffeine |
| Actinomycin | As ethoxycaffeine | |
| Alkylating agents | Nitrogen mustard | Production of abnormal DNA |
| Diepoxypropyl ether | ||
| Nitroso compounds and chelating agents | Many compounds (e.g. cupferron, 8-hydroxy-quinoline) | Possibly combine with some trace heavy metals (Fe, Cu) which may be bound with the DNA molecule |
| Miscellaneous | Inorganic cyanides | Inhibition of cytochrome oxidase with resultant peroxide formation |
| Maleic hydrazide | Reaction with sulphydryl groups | |
| Hydroxylamine | Combination with cytosine component of DNA |
Factors which may influence the effect of mutagenic treatment include oxygen tension within the tissues, temperature and pH. Chemical mutagens can be applied in a similar way to colchicine (q.v.). Seeds, whole plants, isolated organs, growing points, etc., are suitable for direct irradiation. In order to obtain single mutations in a plant, irradiation of pollen, which is subsequently used to fertilize a normal flower, is often advantageous. It is unlikely that a pollen grain will retain its viability if it undergoes more than one mutational change.
Among plants of medicinal interest, the production of polyploid forms has already been discussed. Blakeslee’s radiation work on Datura stramonium resulted in the production of many single gene mutation types (e.g. Zigzag, Quercina, Bunchy, Equisetum—names derived from some characteristic aspect of the plant). These mutants are not isolated individuals but are produced regularly by radiation treatment. Some forms such as ‘pale’ (chlorophyll-deficient) are more frequent than others. In many cases Blakeslee was able to map the position of the genes responsible for these effects. Other mutants obtained in these studies were of the extra-chromosomal type (q.v.).
Several workers have studied, without a full genetic analysis, alkaloid production in various species of Datura raised from irradiated seeds; types have been produced which show differences in the relative proportions of the alkaloids synthesized but no new alkaloids have been detected by this treatment.
By the irradiation of poppy seeds with 60Co a number of mutations have been produced, including ones producing plants with an increased morphine content; these increases were maintained in the X2 generation with an average morphine content of 0.52% compared with 0.32% for the controls.
As mentioned earlier, races of sweet lupins (almost free of bitterness) can be obtained by selection. More recently, bitter lupin seeds of an X-ray-induced early-maturing mutant of Lupinus digitatus were treated with ethylmethanesulphonate solution, and, of the 440 progeny, 11 were mutants which could be classed as sweet. Four of these were of normal vigour and near-normal fertility.
Breeding experiments have been performed with irradiated Mentha piperita in the USA in an endeavour to produce a dominant mutation (bud sport) for Verticillium (wilt) resistance, a disease to which mints are particularly prone; a successful strain, Todd’s Mitcham Peppermint, is now cultivated. A radiation-induced mutant of Scotch peppermint (Mentha × gracilis) has been shown to produce an oil typical of the ordinary peppermint (C-3 oxygenated monoterpenes) instead of the C-6 oxygenated monoterpenes characteristic of spearmint (see Fig. 22.4 for formulae). The results have given further insight into the biogenesis of these compounds (R. Croteau et al., Plant Physiol., 1991, 96, 744).
In India mutant strains of Capsicum annuum with increased yields (20–60%) of capsaicin have been isolated from M3 and M4 generations originating from seed treated with sodium azide and ethylmethanesulphonate.
In the future, haploid plants will undoubtedly find increasing use for the study of induced mutations; in many cases whole plants can now be regenerated from haploid tissue cultures of pollen. Such material has the advantage that induced recessive mutations, which in the diploid organism require subsequent breeding experiments for their study, are immediately apparent in the phenotype.
HYBRIDIZATION
In plant breeding hybridization forms a possible means of combining in a single variety the desirable characters of two or more lines, varieties or species, and occasionally of producing new and desirable characters not found in either parent. Hybridization, particularly between homozygous strains which have been inbred for a number of generations, introduces a degree of heterozygosity with resultant hybrid vigour (heterosis) often manifest in the dimensions and other characteristics of the plant. Several methods of breeding crops by the use of sexual hybridization are available and for these the reader is referred to standard works on the subject. Although this chapter is devoted principally to chemical variants of a particular species it is convenient to include here, in addition to intervarietal hybridization, interspecific hybridization in which hybrid vigour is also apparent.
The hybrid nature of a number of drugs (e.g. cinchona, q.v.) is well known. The commercial mints are hybrids and must therefore be propagated vegetatively, as the plants will not breed true and the progeny vary in their oil composition. Hegnauer considers that with the spearmint-type oil (high carvone content) genes of Mentha longifolia or of M. rotundifolia seem to be necessary. The cultivated peppermint (M. piperita) is probably a hybrid derived from M. aquatica and M. spicata and it is the former that contributes the menthofuran characteristics. Hybrids between various species of Mentha have been used to study the inheritance of a number of essential oil components, including menthol, carvone and pulegone.
Hybrids of species producing tropane alkaloids have received considerable study. In the genus Datura the effect of hybridization on chemical constituents is illustrated by the cross D. ferox × D. stramonium. The aerial organs of the latter normally contain hyoscyamine and hyoscine (2:1 ratio) at the flowering period; and those of the former, hyoscine with some meteloidine. The F1 of the cross consists of plants larger than either of the parents, and containing hyoscine as the principal alkaloid with only small amounts of other bases. In the F2, segregation occurs as regards both morphological characters and alkaloid constituents. With D. leichhardtii and D. innoxia the former plant produces hyoscyamine and hyoscine (2:1) and the latter species usually mainly hyoscine but sometimes, according to conditions of growth, appreciable quantities of hyoscyamine. In this instance the F1 hybrid contains a hyoscyamine:hyoscine ratio intermediate between that of the two parents. Various hybrids of tree daturas have also given favourable alkaloid yields and have been subjected to field trials in Ecuador. These plants are self-sterile but can be propagated by cloning (Chapter 13).
In experiments performed in Australia on hybrids of Duboisia leichhardtii and D. myoporoides no uniform dominance of either hyoscine or hyoscamine in the F1 was obtained, although the ratio generally favoured hyoscyamine even in geographical areas conducive to the production of hyoscine. Investigations of this type on Duboisia are complicated by the occurrence of chemical races and by the marked susceptibility of alkaloid content to environment. However, single desirable hybrids can be vegetatively propagated and have now become established as commercial crops (q.v.). Experiments in Japan on artificial crosses of Duboisia produced some F1 hybrids which contained more than twice the amount of alkaloids (hyoscine and hyoscyamine) than that contained by the parents. Greatest increases were found when D. myoporoides was the female parent. Naturally occurring intergeneric hybrids involving Duboisia spp. and other Anthocercideae have been analysed (El-Imam et al., Int. J. Pharmacog., 1991, 29, 263) and the alkaloid metabolism in callus and regenerating shoot cultures of the commercial hybrid D. leichhardtii × D. myoporoides studied (Gritsanapan and Griffin, Phytochemistry, 1992, 31, 3069).
Nicotiana tabacum (Solanaceae), as now cultivated, must have been derived from at least two different parent species, and ‘synthetic’ tobaccos can be prepared by using suspected species as parents. Although it has not been possible to produce in this way species exactly comparable to N. tabacum, such synthetic plants are most useful for the study of alkaloid inheritance characteristics. This is important in the commercial production of tobacco, in which both the quantity and the nature of the alkaloid produced are important. Demethylation of nicotine may take place in the leaves of some species, and by hybrid studies this reaction has been shown to be due, in the groups of plants studied, to either one pair of dominant factors or two pairs of dominant and independent factors.
The inheritance of the opium alkaloids (morphine, codeine, thebaine, narcotine and papaverine) has been studied in the cross Papaver somniferum × P. setigerum. A heterotic increase in codeine and thebaine was found in different F1 plants, and in the F2 plants, with the exception of codeine, some increase in alkaloid content was noted. An absence of narcotine was generally dominant over its presence (Khanna et al., Planta Med., 1986, p. 157). A continuation of this work to the F8 generation (Shukla et al., Int. J. Pharmacognosy, 1995, 33, 228) resulted in a population that was completely diploid but which showed considerable diversity with regard to the opium contents of morphine, narcotine and papaverine. The pattern of alkaloids was closer to that of P. somniferum than to that of P. setigerum with morphine contents ranging from 8.0 to 30.0%. The authors envisaged that a suitable breeding programme could result in opium with a higher level of morphine than that normally encountered. F1 hybrids of P. bracteatum and P. orientale contained a lower thebaine content and higher oripavine content than in either parent, a result which provided genetic evidence for the biosynthetic linkage between these alkaloids (Fig. 26.15).
In a continuation of the Israeli work on withanolides Eastwood et al. reported that a cross of a South African chemotype of Withania somnifera with the Israeli chemotype II produced three new withanolides not detected in either parent. The outstanding feature was the presence of a new oxidizing system in the hybrid which apparently oxidizes ring A to a diketone in the presence of either the saturated or the unsaturated lactone (Fig. 14.2). A further notable feature was the isolation of a compound having a saturated C2–C3 link, a feature which had only been noticed in a minor constituent of chemotype I. In Datura, too, hybrids may contain different withanolides from those of either parent and involve different oxidation states of ring C.
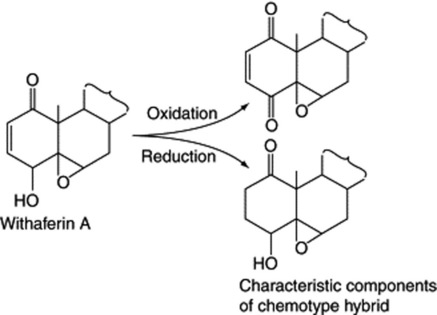
Fig. 14.2 Withanolide oxidations and reductions characteristic of a Withania somnifera chemotype hybrid (for complete structure of withaferin A see ‘Withanolides’).
Solasodine occurs in Solanum incanum up to the extent of 1.8% whereas S. melongena contains only traces of steroidal alkaloids. Work in Jammu, India, has shown the F1 hybrid of these species to be much more prolific in fruit-bearing than either of its parents, with a content of 0.5% solasodine. The diversity of glycoalkaloids found in Solanum species is, in part, the result of the independent inheritance of the aglycone and glycosidic moieties and explains the occurrence of a new alkaloid, sisuine, found in the hybrid of S. acaule and S. ajanhuiri. This group of hybrid clones is cultivated by the Aymara people of Western Bolivia.
In 1961, Calcandi and coworkers demonstrated (Table 14.6) that in the hybridization of Digitalis species, lanatoside formation is dominant to purpurea glycoside formation; in 1984 these observations were largely confirmed for some crosses and extended with more detailed analysis by Wichtl and Mangkudidjojo (Pharm. Zeit., 1984, 129, 686). For the glycoside content of other Digitalis F1 hybrids including those of D. ferruginea and D. cariensis, see Fingerhut et al., Planta Med., 1991, 57 (Suppl. No. 2), A70; A71.

Preliminary studies by Cornish et al. in 1983 indicated that foenugreek seed, a potential source of diosgenin (Chapter 23), is capable of genetic improvement regarding the monohydroxysapogenin yield by hybridization of various races of Trigonella foenum-graecum.
Pyrethrum hybrids have been used in Kenya for pyrethrin production (see Chapter 40); these hybrids are produced either by crossing two clones assumed to be self-sterile or by planting a number of desirable clones together and bulking the seed. The aim is to obtain progeny which, owing to hybrid vigour, will increase pyrethrin yield per acre. Unfortunately, it appears that increased production of pyrethrins is not always associated with hybrid vigour. Research at the Pyrethrum Marketing Board, Kenya, indicated that clonal selection, based on vigour and individual pyrethrin content of hybrids raised from seed, followed by vegetative propagation, would give better results.
TRANSGENIC MEDICINAL PLANTS
Until recently the transfer of genetic material from one plant to another was dependent on hybridization as has been discussed above and its application is limited to those relatively few species which are intercompatible and give fertile F1 hybrids.
Developments in molecular biology and chemical genetics have provided new vistas for genetics and it is now possible to explain, in terms of the DNA molecule, chromosome duplication and the structure of the gene itself—two mysteries involving the nature of life which intrigued and baffled scientists for many years. From such fundamental studies the science of genetic engineering explosively emerged in 1973, when it became possible, by the formation of recombinant DNA, to transfer certain DNA sequences from the chromosomes of one organism to those of another. Since DNA assemblages represent the genes or hereditary characters of an organism, this led the way for the artificial transfer of a particular character from one organism to another.
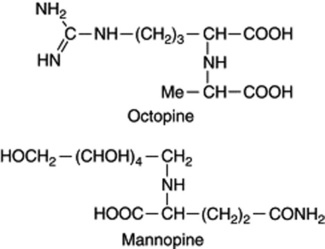
Nature itself does effect genetic transfers between very dissimilar organisms as evidenced by the well-known crown gall and hairy root diseases of plants. Both are caused by bacterial infection of the plant and involve, respectively, the soil organisms Agrobacterium tumefaciens and A. rhizogenes. Crown gall disease is essentially a plant cancer and hairy root disease is also a proliferation manifestation in which there is an excessive development of the root system. A further characteristic associated with both diseases is the production, by the host, of a group of substances called opines, the actual compounds synthesized being specific for the particular parasite involved. Many opines are dicarboxylic acids, the majority being formed by condensation of an amino acid (lysine, arginine, ornithine, histidine) with either pyruvate or α-ketoglutarate; others involve simple sugars such as mannose and sucrose, see Fig. 14.3. (For a review on opines (129 refs) see Y. Dessaux et al., Phytochemistry, 1993, 34, 31.) Another important feature concerning these diseases, first noted in the late 1960s, was that Agrobacterium strains would degrade only those opines for whose occurrence in the host they were responsible. Thus opines, of no value to the plant cell, constitute an exclusive food substrate for the particular bacterial strain. When, however, the bacterium is eliminated from the host plant (e.g. by treatment with an antibiotic) the characteristic morphological features and the production of opines continue to be maintained. This is because genetic material (T-DNA) of the bacterium carrying the genes which express the disease has been transported to, and integrated into, the plant DNA. These genes are carried on large plasmids which have been named Ti (Tumour-inducing) and Ri (Root-inducing) plasmids. Such transgenic plant material is of particular medicinal and phytochemical interest when grown as transformed cell cultures or hairy root cultures in studies involving the production of secondary metabolites and has already been discussed in some detail in Chapter 13.
A further development has been to utilise Agrobacterium in a vector system for transferring genes from one species of higher plant to another. Such transfers usually involve just a single enzyme system; multiple enzyme systems involved in secondary metabolism and originating from different chromosomes obviously present great problems. Of pharmacognostical significance is the work of Hashimoto’s group at Kyoto University on Atropa belladonna: the conversion ofhyoscyamine to hyoscine (Chapter 26) in the plant involves the enzyme hyoscyamine-6-hydroxylase and in the normal belladonna plant the gene is not strongly expressed and plants are hyoscyamine dominant. Conversely, although Hyoscyamus niger contains less alkaloid (% dry weight) than belladonna it contains a higher proportion of hyoscine. The hydroxylase gene of H. niger was placed under the control of the cauliflower mosaic virus 35S promoter and introduced to A. belladonna roots by a binary vector system using Agrobacterium rhizogenes (Phytochemistry, 1993, 32, 713). The belladonna hairy roots produced showed increased amounts, and increased enzyme activities, of the hydroxylase and contained up to fivefold higher concentrations of hyoscine than the wild-type hairy roots. A second paper (Proc. Natl. Acad. Sci. USA, 1992, 89, 11799) reported that whole plants regenerated from the transformed roots showed, in the first transformant and its selfed progeny, an alkaloid composition of the leaves and stems that was almost exclusively hyoscine. Such a plant, if developed commercially, would be a valuable temperate-region source of hyoscine.
In the same area K. Jouhikainen et al. (Planta, 1999, 208, 545) have demonstrated enhancement of scopolamine production in Hyoscyamus muticus hairy root cultures by introducing the above 35S-h6h transgene that codes for the enzyme hyoscyamine-6β-hydroxylase (EC 1.14.11.11). H. muticus is a species producing up to 6% dryweight of tropane alkaloids, normally hyoscyamine; the introduction of the above transgene moved alkaloid production towards scopolamine (but not entirely) and indicated the potential for greater conversion by the use of more efficient promoter systems.
With belladonna, the first successful application of transferring an agronomically useful trait to a medicinal plant has been reported (K. Saito et al., Plant Cell Rep., 1992, 11, 219). It involved the production of transgenic fertile A. belladonna plants integrated with a herbicide-resistant bar gene by means of Agrobacterium–Ri vector. The regenerated plants were produced via hairy roots and the bar trait was transferred to progeny, which showed resistance to bialaphos and phosphinothricin.
An example of the possible potential for increasing alkaloid yield in Cinchona is illustrated by the successful integration of two genes from Catharanthus roseus into the hairy roots of Cinchona officinalis (A.-Geerlings et al., Plant Cell Reports, 1999, 19, 191). These two genes, responsible for the respective production of the enzymes tryptophan decarboxylase and strictosodine synthase, are crucial to the synthesis of both the cinchona and catharanthus alkaloids (Chapter 26). As a result of the transfer both the quinine and quinidine levels were raised above the normal for C. officinalis hairy roots; in this instance however, after one year, the roots had lost their capacity to produce alkaloids.
Transgenic plants of Panax ginseng which develop abundant roots have been produced and could therefore be of commercial significance (D.-C. Yang and Y.-E. Choi, Plant Cell Reports, 2000, 19, 491).