Chapter 15 Musculoskeletal System
Skeletal System
As the notochord and neural tube form, the intraembryonic mesoderm lateral to these structures thickens to form two longitudinal columns of paraxial mesoderm (Fig. 15-1A and B). Toward the end of the third week, these columns become segmented into blocks, that is, somites (Fig. 15-1C). Each somite differentiates into two parts (Fig. 15-1D and E):
• The ventromedial part is the sclerotome; its cells form the vertebrae and ribs.
• The dorsolateral part is the dermomyotome; cells from its myotome region form myoblasts (primordial muscle cells), whereas those from its dermatome region form the dermis (fibroblasts).
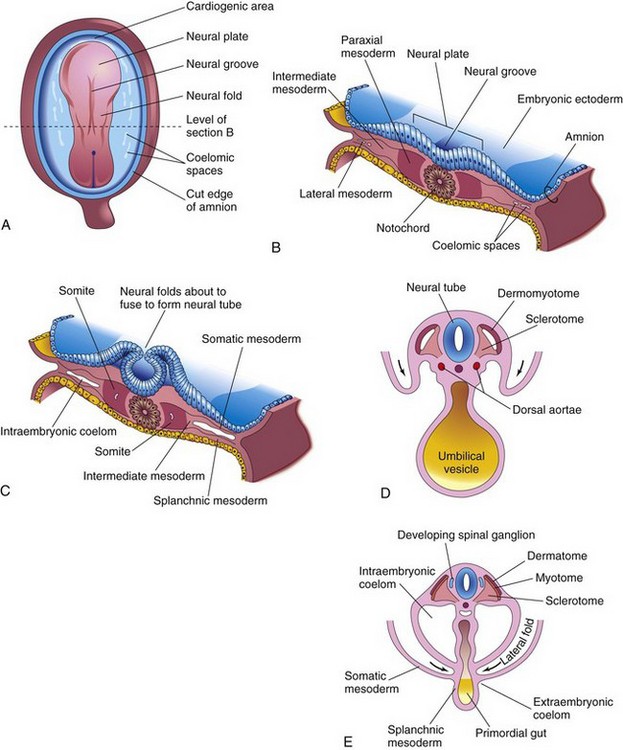
Figure 15–1 Formation and early differentiation of somites. A, Dorsal view of a presomite embryo (approximately 18 days). B, Transverse section of the embryo shown in A, illustrating the paraxial mesoderm from which the somites are derived. C, Transverse section of an embryo at approximately 22 days, at which time early somites have appeared. D, Transverse section of an embryo at approximately 24 days. The dermomyotome region of the somite gives rise to the dermatome and myotome. E, Transverse section of an embryo at approximately 26 days, showing the dermatome, myotome, and sclerotome regions of the somite. The arrows in D and E indicate movement of the lateral body folds.
The bones and connective tissue of the craniofacial structures are formed from mesenchyme in the head region that is derived from the neural crests.
Development of Cartilage and bone
Histogenesis of Cartilage
Cartilage develops from mesenchyme and first appears in embryos during the fifth week. In areas where cartilage is to develop, the mesenchyme condenses to form chondrification centers. The mesenchymal cells differentiate into chondroblasts, which secrete collagenous fibrils and extracellular matrix. Subsequently, collagenous or elastic fibers or both are deposited in the intercellular substance or matrix. Three types of cartilage are distinguishable according to the type of matrix that is formed:
Histogenesis of Bone
Bone develops primarily in two types of connective tissue—mesenchyme and cartilage—but it can also develop in other connective tissues. Most flat bones develop in mesenchyme within preexisting membranous sheaths; this type of osteogenesis is called intramembranous bone formation. Mesenchymal models of most limb bones are transformed into cartilaginous bone models, which later become ossified by endochondral bone formation. Like cartilage, bone consists of cells and an organic intercellular substance—bone matrix, which comprises collagen fibrils embedded in an amorphous component.
Studies of cellular and molecular events that occur during embryonic bone formation suggest that osteogenesis and chondrogenesis are programmed early in development and are independent processes under the influence of vascular events.
Bone morphogenetic proteins 5 and 7 and growth and differentiation factor 5, members of the tumor growth factor-b superfamily, as well as other signaling molecules, have been implicated as endogenous regulators of chondrogenesis and skeletal development.
Intramembranous Ossification
The mesenchyme condenses and becomes highly vascular; some cells differentiate into osteoblasts (bone-forming cells) and begin to deposit unmineralized matrix—osteoid (Fig. 15-2). Calcium phosphate is then deposited in the osteoid tissue as it is organized into bone. Osteoblasts are trapped in the matrix and become osteocytes. Spicules of bone soon become organized and coalesce into lamellae (layers).
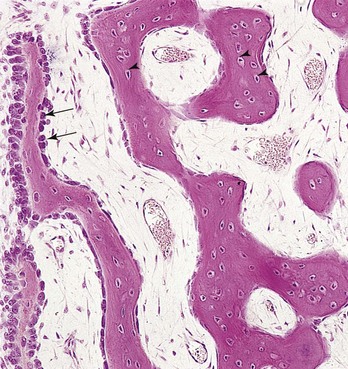
Figure 15–2 Light micrograph of intramembranous ossification (×132). The trabeculae of the bone are being formed by osteoblasts lining their surface (arrows). Observe that osteocytes are trapped in the lacunae (arrowheads) and that primordial osteons are beginning to form. The osteons (canals) contain blood capillaries.
(From Gartner LP, Hiatt JL: Color Textbook of Histology, 2nd ed. Philadelphia, WB Saunders, 2001.)
Concentric lamellae develop around blood vessels, forming osteons (Haversian systems). Some osteoblasts remain at the periphery of the bone and continue to lay down layers, forming plates of compact bone on the surfaces. Between the surface plates, the intervening bone remains spiculated, or spongy. This spongy environment is somewhat accentuated by the action of osteoclasts, which reabsorb bone. In the interstices of the spongy bone, the mesenchyme differentiates into bone marrow. During fetal and postnatal life, continuous remodeling of bone occurs by the coordinated action of osteoclasts and osteoblasts.
Endochondral Ossification
Endochondral ossification is a type of bone formation that occurs in preexisting cartilaginous models (Fig. 15-3A to E). In a long bone, the primary center of ossification appears in the diaphysis, which forms the shaft of the bone (Fig. 15-3B). Here the cartilage cells hypertrophy, the matrix becomes calcified, and the cells die. Concurrently, a thin layer of bone is deposited under the perichondrium surrounding the diaphysis; thus, the perichondrium becomes the periosteum (Fig. 15-3A and B). Invasion of the vascular connective tissue by the blood vessels surrounding the periosteum breaks up the cartilage. Some invading cells differentiate into hemopoietic cells, which are responsible for the formation of blood cells in the bone marrow. This process continues toward the epiphyses, or ends of the bone. The spicules of bone are remodeled by the action of osteoclasts and osteoblasts.
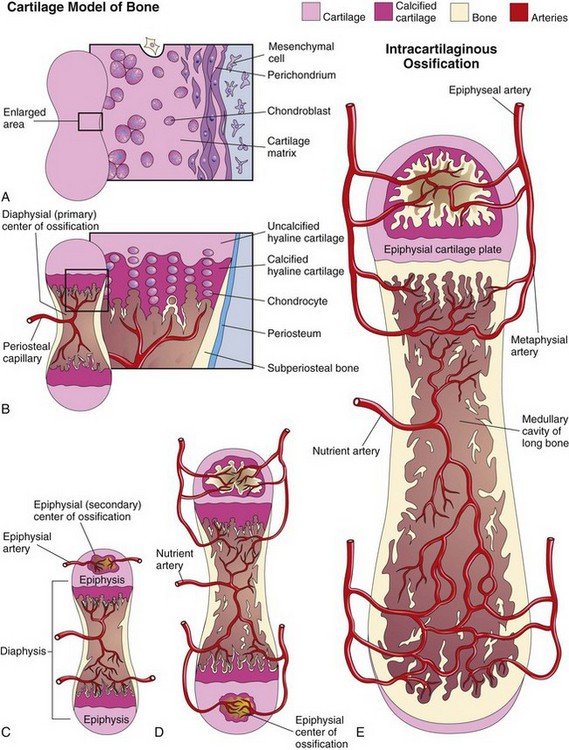
Figure 15–3 A to E, Schematic longitudinal sections, showing endochondral ossification in a developing long bone.
Lengthening of the long bones occurs at the diaphysial-epiphysial junction. The lengthening of bone depends on the epiphysial cartilage plates (growth plates), whose chondrocytes proliferate and participate in endochondral bone formation (Fig. 15-3D and E). Toward the diaphysis, the cartilage cells hypertrophy and the matrix becomes calcified. Spicules are isolated from each other by vascular invasion from the marrow or medullary cavity. Bone is deposited on these spicules by osteoblasts; resorption of this bone keeps the spongy bone masses relatively constant in length and enlarges the medullary cavity.
Rickets
Rickets is a disease that occurs in children with vitamin D deficiency. This vitamin is required for calcium absorption by the intestine. The resulting calcium deficiency causes disturbances in ossification of the epiphysial cartilage plates and orientation of cells at the metaphysis. The limbs are shortened and deformed, with severe bowing of the bones.
Ossification of the limb bones makes demands on the maternal supply of calcium and phosphorus beginning at approximately 8 weeks. At birth, the diaphyses are largely ossified but the epiphyses are still cartilaginous. Secondary ossification centers appear in the epiphyses during the first few years after birth. The epiphysial cartilage cells hypertrophy, and there is invasion by vascular connective tissue. Ossification spreads radially. The articular cartilage and the epiphysial cartilage plate remain cartilaginous. On completion of bone growth, this plate is replaced by spongy bone, the epiphyses and the diaphysis are united, and no further elongation of the bone occurs.
In most bones, the epiphyses have fused with the diaphysis by 20 years of age. Growth in the diameter of a bone results from deposition of bone at the periosteum and from resorption on the medullary surface. The rate of deposition and resorption is balanced to regulate the thickness of the compact bone and the size of the medullary cavity. The internal reorganization of bone continues throughout life.
Development of Joints
Joints begin to develop with the appearance of the interzonal mesenchyme during the sixth week (Fig. 15-4A), and they resemble adult joints by the end of the eighth week.
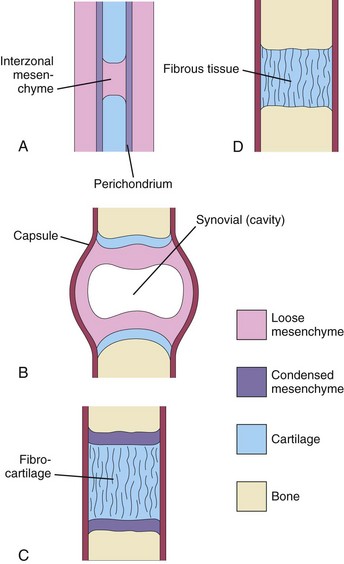
Figure 15–4 Development of joints during the sixth and seventh weeks. A, Condensed mesenchyme continues across the gap, or interzone, between the developing bones, enclosing some interzonal mesenchyme between them. This primordial joint may differentiate into a synovial joint (B), a cartilaginous joint (C), or a fibrous joint (D).
Fibrous Joints
During the development of fibrous joints, the interzonal mesenchyme between the developing bones differentiates into dense fibrous tissue (Fig. 15-4D). The sutures of the cranium are an example of fibrous joints.
Cartilaginous Joints
During the development of cartilaginous joints, the interzonal mesenchyme between the developing bones differentiates into hyaline cartilage (e.g., the costochondral joints) or fibrocartilage (Fig. 15-4C) (e.g., the pubic symphysis).
Synovial Joints
During the development of synovial joints (e.g., the knee joint), the interzonal mesenchyme between the developing bones differentiates as follows (Fig. 15-4B):
Development of Axial Skeleton
The axial skeleton is composed of the cranium (skull), vertebral column, ribs, and sternum. During the fourth week, cells in the sclerotomes surround the neural tube (primordium of spinal cord) and the notochord, the structure around which the primordia of the vertebrae develop. This positional change of the sclerotomal cells is effected by differential growth of the surrounding structures, not by the migration of sclerotomal cells.
Development of Vertebral Column
During the precartilaginous stage, mesenchymal cells from the sclerotomes are found in three main areas (Fig. 15-5A): around the notochord, surrounding the neural tube, and in the body wall.
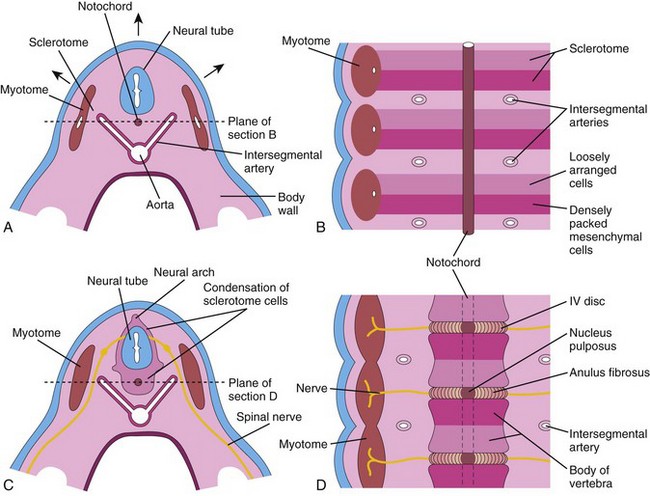
Figure 15–5 A, Transverse section through a 4-week embryo. The arrows indicate the dorsal growth of the neural tube and the simultaneous dorsolateral movement of the somite remnant, which leaves behind a trail of sclerotomal cells. B, Frontal section of the same embryo as in A, showing that the condensation of sclerotomal cells around the notochord consists of a cranial area of loosely packed cells and a caudal area of densely packed cells. C, Transverse section through a 5-week embryo. Note the condensation of sclerotomal cells around the notochord and the neural tube, which forms a mesenchymal vertebra. D, Frontal section, showing that the vertebral body forms from the cranial and caudal halves of two successive sclerotomal masses. The intersegmental arteries now cross the bodies of the vertebrae and the spinal nerves lie between the vertebrae. The notochord is degenerating, except in the region of the intervertebral disc, where it forms the nucleus pulposus.
In a frontal section of a 4-week embryo, the sclerotomes appear as paired condensations of mesenchymal cells around the notochord (Fig. 15-5B). Each sclerotome consists of loosely arranged cells cranially and densely packed cells caudally. Some densely packed cells move cranially, opposite the center of the myotome, where they form the intervertebral (IV) disc (Fig. 15-5C and D). These cells express Pax-1, a paired box gene. The remaining densely packed cells fuse with the loosely arranged cells of the immediately caudal sclerotome to form the mesenchymal centrum, the primordium of the vertebral body. Thus, each centrum develops from two adjacent sclerotomes and becomes an intersegmental structure. The spinal nerves now lie in close relationship to the IV discs, and the intersegmental arteries lie on each side of the vertebral bodies. In the thorax, the dorsal intersegmental arteries become the intercostal arteries. Studies indicate that the regional development of the vertebral column is regulated along the anterior-posterior axis by homeobox (Hox) and paired box (Pax) genes.
Where it is surrounded by the developing vertebral bodies, the notochord degenerates and disappears. Between the vertebrae, the notochord expands to form the gelatinous center of the IV disc—the nucleus pulposus (Fig. 15-5D). This nucleus is later surrounded by circularly arranged fibers that form the anulus fibrosus. The nucleus pulposus and anulus fibrosus together constitute the IV disc. The mesenchymal cells that surround the neural tube form the neural arch, the primordium of the vertebral arch (Figs. 15-5C and 15-6D). The mesenchymal cells in the body wall form the costal processes, which form the ribs in the thoracic region.
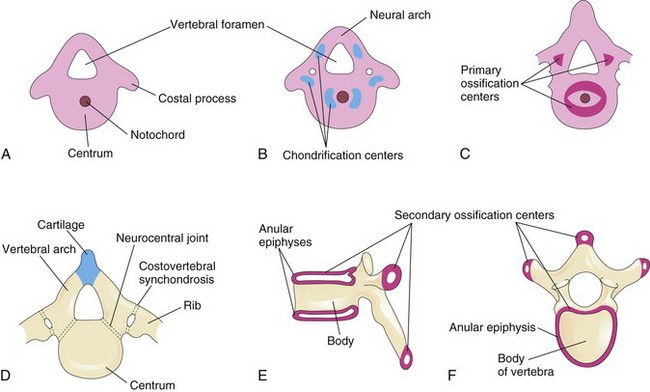
Figure 15–6 Stages of vertebral development. A, Mesenchymal vertebra at 5 weeks. B, Chondrification centers in a mesenchymal vertebra at 6 weeks. The neural arch is the primordium of the vertebral arch of the vertebra. C, Primary ossification centers in a cartilaginous vertebra at 7 weeks. D, Thoracic vertebra, consisting of three bony parts, at birth. Note the cartilage between the halves of the vertebral arch and between the arch and the centrum (neurocentral joint). E and F, Two views of a typical thoracic vertebra at puberty, showing the location of the secondary centers of ossification.
Chordoma
Remnants of the notochord may persist and give rise to a chordoma. Approximately one third of these slowly growing, malignant tumors involve the base of the cranium and extend to the nasopharynx. Chordomas infiltrate bone and are difficult to remove; few patients survive longer than 5 years. Chordomas may also develop in the lumbosacral region.
Cartilaginous Stage of Vertebral Development
During the sixth week, chondrification centers appear in each mesenchymal vertebra (Fig. 15-6A and B). At the end of the embryonic period, the two centers in each centrum fuse to form a cartilaginous centrum. Concomitantly, the centers in the neural arches fuse with each other and the centrum. The spinous and transverse processes develop from extensions of chondrification centers in the vertebral arch. Chondrification spreads until a cartilaginous vertebral column is formed.
Bony Stage of Vertebral Development
Ossification of typical vertebrae begins during the embryonic period and usually ends by the 25th year. There are primary ossification centers in the centrum—ventral and dorsal (Fig. 15-6C), which soon fuse to form one center. Primary centers are also present by the end of the embryonic period in each half of the vertebral arch.
Ossification becomes evident in the neural arches during the eighth week. At birth, each vertebra consists of three bony parts connected by cartilage (Fig. 15-6D). The bony halves of the vertebral arch usually fuse during the first 3 to 5 years of life. The arches first unite in the lumbar region, and union progresses cranially. The vertebral arch articulates with the centrum at cartilaginous neurocentral joints. These articulations permit the vertebral arches to grow as the spinal cord enlarges. These joints disappear when the vertebral arch fuses with the centrum during the third to sixth years. Five secondary ossification centers appear in the vertebrae after puberty:
• One for the tip of the spinous process
• One for the tip of each transverse process
• Two anular epiphyses, one on the superior rim and one on the inferior rim of the vertebral body (Fig. 15-6E and F)
The vertebral body is a composite of the anular epiphyses and the mass of bone between them. All secondary centers unite with the rest of the vertebrae at approximately 25 years of age. Variations in the ossification of vertebrae occur in C1 (atlas), C2 (axis), and C7 vertebrae, and in the lumbar vertebrae, sacrum, and coccyx.
Variations in the Number of Vertebrae
Most people have 7 cervical, 12 thoracic, 5 lumbar, and 5 sacral vertebrae. A few have one or two more vertebrae or one less. An apparent extra (or absent) vertebra in one segment of the column may be compensated for by an absent (or extra) vertebra in an adjacent segment.
Klippel-Feil Syndrome
The main features of the Klippel-Feil syndrome (Brevicollis) are a short neck, a low hairline, and restricted neck movements. In most cases, the number of cervical vertebral bodies is fewer than normal. In some cases, there is a lack of segmentation of several elements of the cervical region of the vertebral column. The number of cervical nerve roots may be normal, but they are small, as are the intervertebral foramina. Persons with this syndrome are often otherwise normal, but the association of this anomaly with other congenital anomalies is not uncommon.
Development of Ribs
The ribs develop from the mesenchymal costal processes of the thoracic vertebrae (Fig. 15-6A). They become cartilaginous during the embryonic period and ossify during the fetal period. The original site of union of the costal processes with the vertebrae is replaced by costovertebral synovial joints (Fig. 15-6D). Seven pairs of ribs (true ribs) attach by their own cartilages to the sternum. Five pairs of ribs (false ribs) attach to the sternum through the cartilage of another rib or ribs. The last two pairs of ribs (floating ribs) do not attach to the sternum.
Development of Sternum
A pair of vertical mesenchymal bands—sternal bars—develops ventrolaterally in the body wall. Chondrification occurs in these bars as they move medially. They fuse craniocaudally in the median plane to form cartilaginous models of the manubrium, sternebrae (segments of the sternal body), and xiphoid process. Centers of ossification appear craniocaudally in the sternum before birth, except the ossification center for the xiphoid process, which appears during childhood.
Development of Cranium
The cranium (skull) develops from the mesenchyme around the developing brain. The cranium consists of:
Cartilaginous Neurocranium
Endochondral ossification of the neurocranium forms the bones of the base of the cranium. The ossification pattern of these bones has a definite sequence, beginning with the occipital bone, the body of the sphenoid, and the ethmoid bone. The parachordal cartilage, or basal plate, forms around the cranial end of the notochord (Fig. 15-7A) and fuses with the cartilages derived from the sclerotome regions of the occipital somites. This cartilaginous mass contributes to the base of the occipital bone; later, extensions grow around the cranial end of the spinal cord and form the boundaries of the foramen magnum (Fig. 15-7C). The hypophysial cartilage forms around the developing pituitary gland and fuses to form the body of the sphenoid bone (Fig. 15-7B). The trabeculae cranii fuse to form the body of the ethmoid bone, and the ala orbitalis forms the lesser wing of the sphenoid bone.
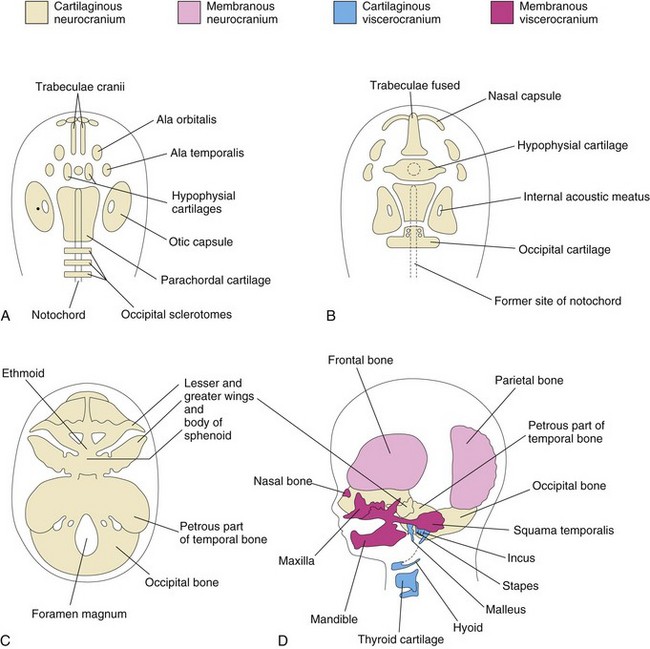
Figure 15–7 Stages in the development of the cranium. The base of the developing cranium is viewed superiorly (A to C), and laterally (D). A, At 6 weeks, showing the various cartilages that will fuse to form the chondrocranium. B, At 7 weeks, after fusion of some of the paired cartilages. C, At 12 weeks, showing the cartilaginous base of the cranium, or chondrocranium, formed by the fusion of various cartilages. D, At 20 weeks, indicating the derivation of the bones of the fetal cranium.
Otic capsules develop around the otic vesicles, the primordia of the internal ears (see Chapter 17), and form the petrous and mastoid parts of the temporal bone. Nasal capsules develop around the nasal sacs (Chapter 10) and contribute to the formation of the ethmoid bone.
Membranous Neurocranium
Intramembranous ossification occurs in the mesenchyme at the sides and top of the brain, forming the calvaria (cranial vault). During fetal life, the flat bones of the calvaria are separated by dense connective tissue membranes that form fibrous joints, that is, sutures (Fig. 15-8). Six large fibrous areas—fontanelles—are present where several sutures meet. The softness of the bones and their loose connections at the sutures enable the calvaria to undergo changes of shape (molding of fetal cranium) during birth. The frontal bones become flat, the occipital bone is drawn out, and one parietal bone slightly overrides the other one. Within a few days after birth, the shape of the calvaria returns to normal.
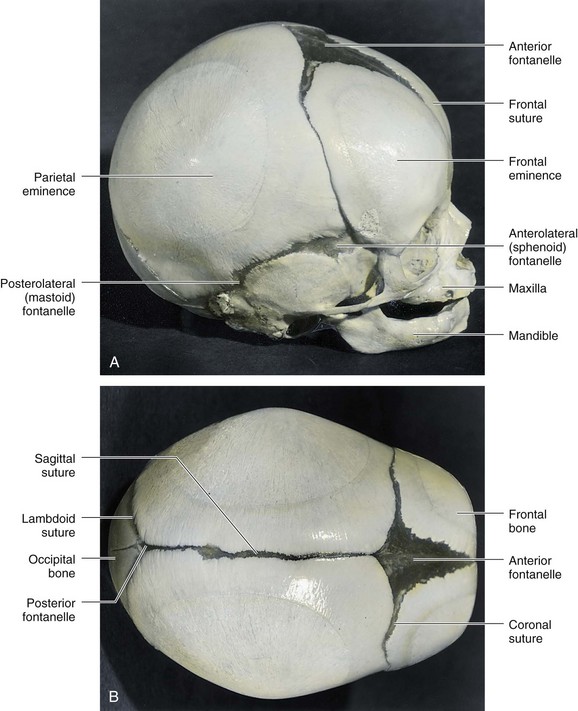
Figure 15–8 Fetal cranium (skull), showing the bones, fontanelles, and connecting sutures. A, Lateral view. B, Superior view. The posterior and anterolateral fontanelles disappear within 2 or 3 months after birth because of the growth of the surrounding bones, but they remain as sutures for several years. The posterolateral fontanelles disappear in a similar manner by the end of the first year, and the anterior fontanelle disappears by the end of the second year. The halves of the frontal bone normally begin to fuse during the second year, and the frontal suture is usually obliterated by the eighth year.
Cartilaginous Viscerocranium
The cartilaginous viscerocranium is derived from the cartilaginous skeleton of the first two pairs of pharyngeal arches (see Chapter 10).
• The dorsal end of the first pharyngeal arch cartilage forms the malleus and incus.
• The dorsal end of the second pharyngeal arch cartilage forms the stapes and styloid process of the temporal bone. Its ventral end ossifies to form the lesser horn and the superior part of the body of the hyoid bone.
• The third, fourth, and sixth pharyngeal arch cartilages form only in the ventral parts of the arches. The third pharyngeal arch cartilages give rise to the greater horns and the inferior part of the body of the hyoid bone. The fourth and sixth pharyngeal arch cartilages fuse to form the laryngeal cartilages, except for the epiglottis.
Membranous Viscerocranium
Intramembranous ossification occurs in the maxillary prominence of the first pharyngeal arch (see Chapter 10) and subsequently forms the squamous temporal, maxillary, and zygomatic bones. The squamous temporal bones become part of the neurocranium. The mandibular prominence forms the mandible. Some endochondral ossification occurs in the median plane of the chin and in the mandibular condyle.
Newborn Cranium
The cranium of a neonate is large in proportion to the rest of the skeleton, and the face is relatively small compared with the calvaria (roof of cranium). The small facial region of the cranium results from the small size of the jaws, the virtual absence of paranasal (air) sinuses, and underdevelopment of the facial bones.
Postnatal Growth of Cranium
The fibrous sutures permit the brain and calvaria to enlarge during infancy and childhood. The increase in size is greatest during the first 2 years of life, but the calvaria continues to expand to conform to brain growth until approximately 16 years, after which its size usually increases slightly for 3 to 4 years because of thickening of its bones.
There is also rapid growth of the face and jaws, coinciding with eruption of the primary (deciduous) teeth. These facial changes are more marked after the secondary (permanent) teeth erupt (see Chapter 18). There is concurrent enlargement of the frontal and facial regions, associated with the increase in the size of the paranasal sinuses (e.g., the maxillary sinuses). Growth of these sinuses is important in adding resonance to the voice.
Accessory Ribs
Accessory ribs, usually rudimentary, result from the development of costal processes from the cervical or lumbar vertebrae (Fig. 15-6A). The most common type of accessory rib is a lumbar rib, but it is clinically insignificant. Cervical ribs occur in 0.5% to 1% of people (Fig. 15-9A). A cervical rib is attached to the seventh cervical vertebra and may be unilateral or bilateral. Pressure of a cervical rib on the brachial plexus of nerves or the subclavian artery often produces neurovascular symptoms.
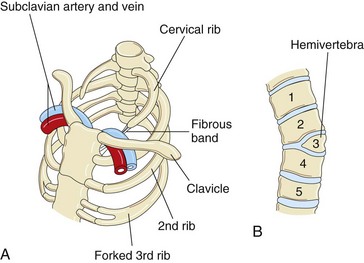
Figure 15–9 Vertebral and rib abnormalities. A, Cervical and forked ribs. Observe that the left cervical rib has a fibrous band that passes posterior to the subclavian vessels and attaches to the sternum. B, Anterior view of the vertebral column, showing a hemivertebra. The right half of the third thoracic vertebra is absent.
Hemivertebra
Developing vertebral bodies have two chondrification centers that soon unite. A hemivertebra results from failure of one of the chondrification centers to appear and subsequent failure of half of the vertebra to form (see Fig. 15-9B). These vertebral defects produce scoliosis (lateral curvature) of the vertebral column.
Rachischisis
Rachischisis (cleft vertebral column) refers to vertebral abnormalities in a complex group of anomalies (axial dysraphic disorders) that primarily affect axial structures (Fig. 15-10). In affected infants, the neural folds do not fuse, either because of faulty induction by the underlying notochord or because of a teratogenic agent.
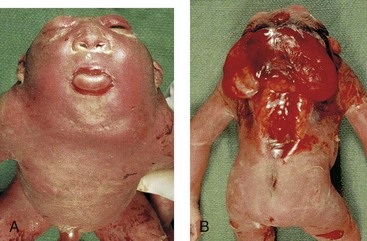
Figure 15–10 Anterior (A) and posterior (B) views of a 20-week fetus with severe birth defects, including acrania (absence of calvaria), cervical rachischisis (extensive clefts in the vertebral arches), cerebral regression (meroencephaly or anencephaly), and iniencephaly (defect in occiput—back of cranium).
(Courtesy of Dr. Marc Del Bigio, Department of Pathology [Neuropathology], University of Manitoba, Winnipeg, Manitoba, Canada.)
Acrania
In acrania, the calvaria is absent, and extensive defects of the vertebral column are often present (Fig. 15-10). Acrania is associated with meroencephaly (anencephaly). Partial absence of the brain occurs in approximately 1 in 1000 births and is incompatible with life. Meroencephaly occurs when the cranial end of the neural tube does not close during the fourth week of development, resulting in subsequent failure of the calvaria to form.
Craniosynostosis
Several cranial deformities result from premature closure of the cranial sutures. Prenatal closure results in the most severe abnormalities. The cause of craniosynostosis is unclear, but genetic factors appear to be important. Homeobox gene (MSX2 and ALX4) mutations have been implicated in cases of craniosynostosis and other cranial defects. These abnormalities are much more common in males than in females, and they are often associated with other skeletal anomalies. The type of cranial deformation produced depends on which sutures close prematurely. If the sagittal suture closes early, the cranium becomes elongated and wedge-shaped, a condition known as scaphocephaly (Fig. 15-11A). This type of cranial deformity constitutes approximately half of the cases of craniosynostosis. Another 30% of cases involve premature closure of the coronal suture, which results in a high, tower-like cranium, a condition known as brachycephaly (see Fig. 15-11B). If the coronal suture closes prematurely on one side only, the cranium is twisted and asymmetrical, resulting in a condition known as plagiocephaly. Premature closure of the metopic suture results in a deformity of the frontal bone and other anomalies, known collectively as trigonocephaly.
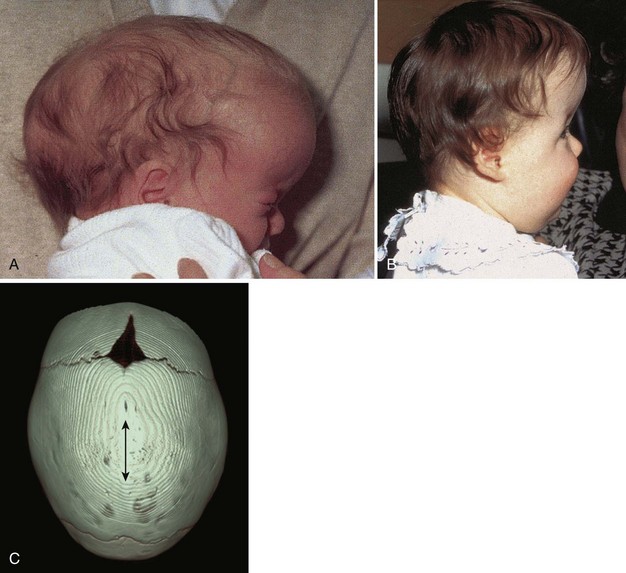
Figure 15–11 Craniosynostosis. A, An infant with scaphocephaly (long narrow head) resulting from premature closure of the sagittal suture. B, An infant with bilateral premature closure of the coronal suture—brachycephaly—resulting in a high, tower-like forehead. C, Cranium of a 9-month-old infant with scaphocephaly resulting from premature closure of the sagittal suture (sagittal synostosis; double arrow). CT reconstructed image.
(A, B, Courtesy of Dr. John A. Jane, Sr., Department of Neurological Surgery, University of Virginia Health System, Charlottesville, VA. C, Courtesy of Dr. Gerald S. Smyser, Altru Health System, Grand Forks, ND.)
Development of Appendicular Skeleton
The appendicular skeleton consists of the pectoral and pelvic girdles and the limb bones. During the sixth week, the mesenchymal bone models undergo chondrification to form hyaline cartilage bone models (Fig. 15-12). The clavicle initially develops by intramembranous ossification; later, growth cartilages form at both ends. The models of the pectoral girdle and the upper limb bones appear slightly before those of the pelvic girdle and the lower limbs; the bone models appear in a proximodistal sequence. The molecular mechanism of limb morphogenesis is regulated by specialized signaling centers along three axes of development (proximal/ distal, ventral/dorsal, and anterior/posterior). Patterning in the developing limbs is controlled by Hox and other complex signaling pathways.
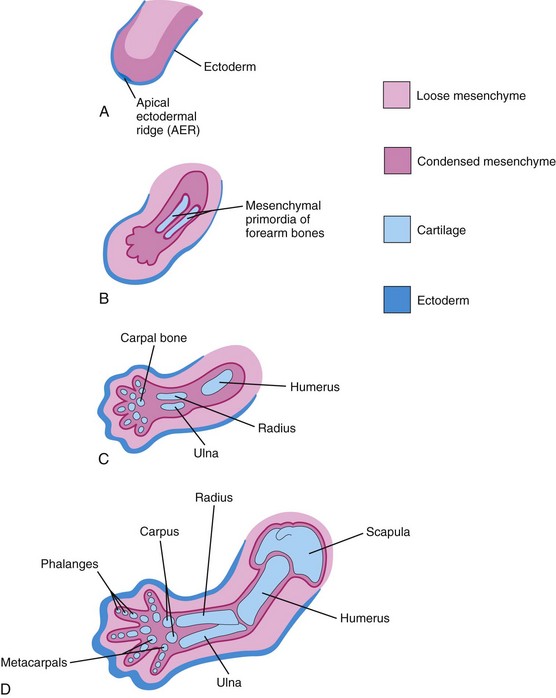
Figure 15–12 Longitudinal sections of the upper limb of a embryo, showing development of the cartilaginous bones. A, At 28 days. B, At 44 days. C, At 48 days. D, At 56 days.
Endochondral ossification begins in the long bones by the eighth week (Fig. 15-3B and C). By 12 weeks, primary ossification centers have appeared in nearly all limb bones (Fig. 15-13). The clavicles begin to ossify before any other bones in the body, followed by the femurs. Virtually all primary centers of ossification (diaphysial) are present at birth.
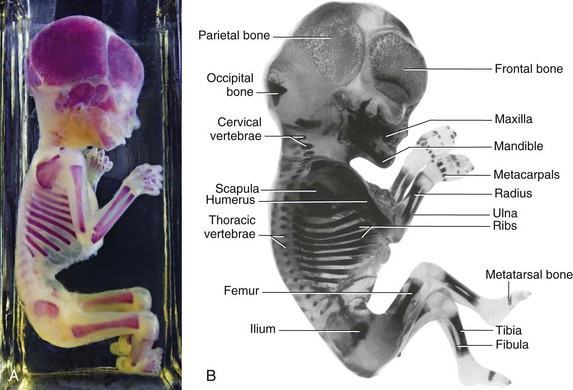
Figure 15–13 A, Alizarin-stained fetus, 20-week fetus. B, Alizarin-stained, 12-week fetus. Observe the degree of progression of ossification from the primary centers of ossification, which are endochondral in the appendicular and axial parts of the skeleton, except for most of the cranial bones. Note that the carpus and tarsus are wholly cartilaginous at this stage, as are the epiphyses of all long bones.
(A, Courtesy of Dr. David Bolender, Department of Cell Biology, Neurobiology, and Anatomy, Medical College of Wisconsin, Milwaukee, Wisconsin. B, Courtesy of Dr. Gary Geddes, Lake Oswego, Oregon.)
The secondary ossification centers of the bones at the knee are the first to appear in utero. The secondary centers for the distal end of the femur and the proximal end of the tibia usually appear during the last month of intrauterine life (34–38 weeks). The secondary centers of the other bones appear after birth. The bone formed from the primary center in the diaphysis does not fuse at the epiphyseal plate with that formed from the secondary centers in the epiphyses until the bone grows to its adult length. This delay enables lengthening of the bone to continue until the final size is reached.
Bone Age
Bone age is a good index of general maturation. A radiologist can determine the bone age of a person by assessing the ossification centers using two criteria:
• The time of appearance of calcified material in the diaphysis, epiphysis, or both is specific for each diaphysis and epiphysis and for each bone and sex.
• The disappearance of the dark line representing the epiphysial cartilage plate indicates that the epiphysis has fused with the diaphysis.
Fusion of the diaphysial–epiphysial centers, which occurs at specific times for each epiphysis, happens 1 to 2 years earlier in females than in males. In the fetus, ultrasonography is used for the evaluation and measurement of bones.
Generalized Skeletal Malformations
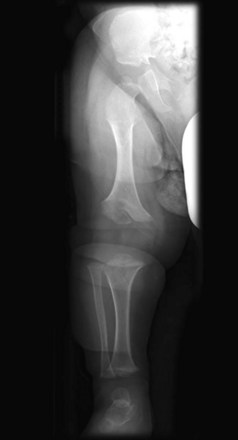
Achondroplasia is a common cause of short stature. It occurs in approximately 1 in 15,000 births. The limbs become bowed and short because of a disturbance of endochondral ossification at the epiphysial cartilage plates, particularly of the long bones, during fetal life (Fig. 15-14). The trunk is usually short, and the head is enlarged, with a bulging forehead and a “scooped-out” nose (flat nasal bone). Achondroplasia is an autosomal dominant disorder, and approximately 80% of cases arise from new mutations; the rate increases with paternal age. The majority of cases are due to a point mutation (f.1,11,12) in the FGFR3 gene, which results in magnification of the normal inhibiting effect of endochondral ossification, specifically in the zone of chondrocyte proliferation. This results in shortened bone, but does not affect growth of bone width (periosteal growth).
Hyperpituitarism
Congenital infantile hyperpituitarism, which causes abnormally rapid growth in infancy, is rare. This condition may result in gigantism (excessive height and body proportions). In adults, hyperpituitarism results in acromegaly (enlargement of the soft tissues, visceral organs, and bones of the face, hands, and feet). In acromegaly, the epiphysial and diaphysial centers of the long bones fuse, thereby preventing elongation of the bones. Both gigantism and acromegaly result from an excessive secretion of growth hormone.
Muscular System
The muscular system develops from the mesoderm, except for the muscles of the iris, which develop from the neuroectoderm. Myoblasts—embryonic muscle cells—are derived from mesenchyme.
Development of Skeletal Muscle
The myoblasts that form the skeletal muscles of the trunk are derived from the mesenchyme in the myotome regions of the somites. The limb muscles develop from myogenic precursor cells in the limb buds. Studies show that these cells originate from the ventral dermomyotome of somites in response to molecular signals from nearby tissues (Fig. 15-15). The myogenic precursor cells migrate into the limb buds, where they undergo epitheliomesenchymal transformation. The first indication of myogenesis (muscle formation) is the elongation of the nuclei and cell bodies of mesenchymal cells as they differentiate into myoblasts. These primordial muscle cells soon fuse to form elongated, multinucleated, cylindric structures—myotubes. At the molecular level, these events are preceded by gene activation and expression of the MyoD family of muscle-specific basic helix-loop-helix transcription factors (MyoD, myogenin, Myf-5, and MRF4) in the precursor myogenic cells. It has been suggested that signaling molecules from the ventral neural tube (Shh), the notochord (Shh), the dorsal neural tube (Wnt, BMP-4), and the overlying ectoderm (Wnt, BMP-4) regulate the beginning of myogenesis and the induction of the myotome.
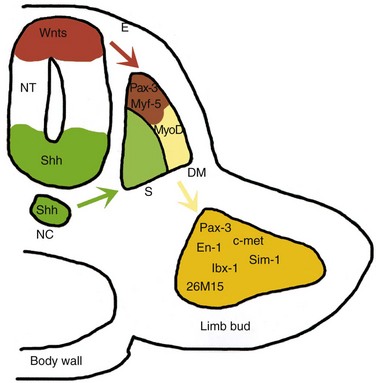
Figure 15–15 A model for molecular interactions during myogenesis. Shh and Wnt, produced by the neural tube (NT) and the notochord (NC), induce Pax-3 and Myf-5 in the somites. Either of them can activate the initiation of MyoD transcription and myogenesis. The surface ectoderm (E) is also capable of inducing Myf-5 and MyoD. Pax-3 also regulates the expression of c-met, which, in turn, is necessary for the migratory ability of the myogenic precursor cells, which also express En-1, Sim-1, lbx-1, and 26M15. DM, dermomyotome; S, sclerotome.
(From Kablar B, Rudnicki MA: Skeletal muscle development in the mouse embryo. Histol Histopathol 15:649, 2000.)
Muscle growth during development results from the ongoing fusion of myoblasts and myotubes. Myofilaments develop in the cytoplasm of the myotubes during or after fusion of the myoblasts. Soon after that, myofibrils and other organelles characteristic of striated muscle cells develop. Because muscle cells are long and narrow, they are called muscle fibers. As the myotubes differentiate, they become invested with external laminae, which segregate them from the surrounding connective tissue. Fibroblasts produce the perimysium and epimysium layers of the fibrous sheath; the endomysium is formed by the external lamina, which is derived from the muscle fiber, and by reticular fibers. Most skeletal muscle develops before birth, and almost all remaining muscles are formed by the end of the first year. The increase in the size of a muscle after the first year results from an increase in the diameter of the fibers because of the formation of more myofilaments. Muscles increase in length and width to grow with the skeleton.
Myotomes
Typically, each myotome part of a somite divides into a dorsal epaxial division and a ventral hypaxial division (Fig. 15-16). Each developing spinal nerve also divides and sends a branch to each division, with the dorsal primary ramus supplying the epaxial division and the ventral primary ramus supplying the hypaxial division. Some muscles—the intercostal muscles, for example—remain segmentally arranged like the somites, but most myoblasts migrate away from the myotome and form nonsegmented muscles.
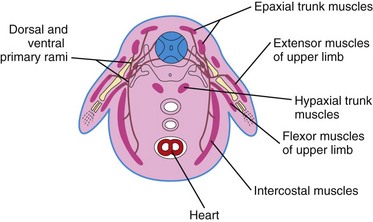
Derivatives of Epaxial Divisions of Myotomes
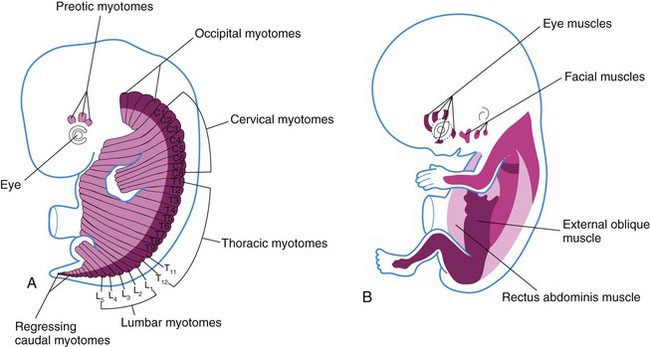
Myoblasts from the epaxial divisions of the myotomes form the segmental muscles of the main body axis, the extensor muscles of the neck and vertebral column (Fig. 15-17). The embryonic extensor muscles that are derived from the sacral and coccygeal myotomes degenerate; their adult derivatives are the dorsal sacrococcygeal ligaments.
Derivatives of Hypaxial Divisions of Myotomes
Myoblasts from the hypaxial divisions of the cervical myotomes form the scalene, prevertebral, geniohyoid, and infrahyoid muscles (Fig. 15-17A). Those from the thoracic myotomes form the lateral and ventral flexor muscles of the vertebral column, whereas the lumbar myotomes form the quadratus lumborum muscle. The muscles of the limbs, the intercostal muscles, and the abdominal muscles are also derived from the hypaxial division of the myotomes. The sacrococcygeal myotomes form the muscles of the pelvic diaphragm and probably the striated muscles of the anus and sex organs.
Pharyngeal Arch Muscles
Myoblasts from the pharyngeal arches form the muscles of mastication and facial expression as well as those of the pharynx and larynx (see Chapter 10). These muscles are innervated by the pharyngeal arch nerves.
Ocular Muscles
The mesoderm in the prechordal plate area is believed to give rise to three preotic myotomes from which myoblasts differentiate (see Fig. 15-17B). Groups of myoblasts, each supplied by its own cranial nerve (CN III, CN IV, or CN VI), form the extrinsic muscles of the eye.
Tongue Muscles
Myoblasts from the occipital (postotic) myotomes form the tongue muscles, which are innervated by the hypoglossal nerve (CN XII).
Limb Muscles
The musculature of the limbs develops from myoblasts surrounding the developing bones (Fig. 15-16). The precursor myogenic cells in the limb buds originate from the somites. These cells are first located in the ventral part of the dermomyotome, and they are epithelial (see Fig. 15-1D). After epitheliomesenchymal transformation, the cells migrate into the primordium of the limb.
Development of Smooth Muscle
Some smooth muscle fibers differentiate from the splanchnic mesenchyme surrounding the endoderm of the primordial gut and its derivatives (see Fig. 15-1E). The smooth muscle in the walls of many blood and lymphatic vessels arises from the somatic mesoderm. The muscles of the iris (sphincter and dilator pupillae) and the myoepithelial cells in the mammary and sweat glands are believed to be derived from mesenchymal cells that originate from ectoderm.
The first sign of differentiation of smooth muscle is the development of elongated nuclei in spindle-shaped myoblasts. During early development, new myoblasts continue to differentiate from mesenchymal cells, but do not fuse; they remain mononucleated. During later development, the division of existing myoblasts gradually replaces the differentiation of new myoblasts in the production of new smooth muscle tissue. Filamentous, but nonsarcomeric, contractile elements develop in their cytoplasm, and the external surface of each differential cell acquires a surrounding external lamina. As smooth muscle fibers develop into sheets or bundles, they receive autonomic innervation; fibroblasts and muscle cells synthesize and lay down collagenous, elastic, and reticular fibers.
Development of Cardiac Muscle
The lateral splanchnic mesoderm gives rise to the mesenchyme surrounding the developing heart tube (see Chapter 14). Cardiac myoblasts are derived from this mesenchyme by differentiation and growth of single cells, unlike striated skeletal muscle fibers, which develop by the fusion of cells. The myoblasts adhere to each other as in developing skeletal muscle, but the intervening cell membranes do not disintegrate; these areas of adhesion give rise to intercalated discs. Growth of cardiac muscle fibers results from the formation of new myofilaments. Late in the embryonic period, special bundles of muscle cells develop that have relatively few myofibrils and relatively larger diameters than typical cardiac muscle fibers. These atypical cardiac muscle cells—Purkinje fibers—form the conducting system of the heart (see Chapter 14).
Anomalies of Muscles
Any muscle in the body may occasionally be absent; common examples are the sternocostal head of the pectoralis major, the palmaris longus, the trapezius, the serratus anterior, and the quadratus femoris. Absence of the pectoralis major, often its sternal part, is usually associated with syndactyly (fusion of digits). This anomaly is part of the Poland syndrome, which also includes breast and nipple aplasia or hypoplasia, deficiencies of axillary hair and subcutaneous fat, and shortened arms and fingers.
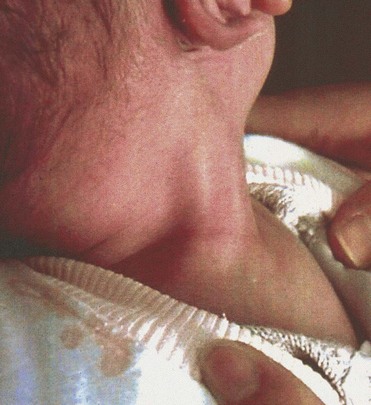
Congenital absence of the diaphragm is usually associated with pulmonary atelectasis (incomplete expansion of the lungs or part of a lung) and pneumonitis (pneumonia). The sternocleidomastoid muscle is sometimes injured at birth, resulting in congenital torticollis. There is fixed rotation and tilting of the head because of concomitant muscle fibrosis, as well as shortening of the sternocleidomastoid muscle on one side (Fig. 15-18). Although birth trauma is commonly considered a cause of congenital torticollis, this may also result from malpositioning in utero.
Development of Limbs
Early Stages of Limb Development
The limb buds first appear toward the end of the fourth week as small elevations of the ventrolateral body wall (Fig. 6-9). Limb development begins with the activation of a group of mesenchymal cells in the lateral mesoderm. The upper limb buds are visible by day 26 or 27, whereas the lower limb buds appear 1 to 2 days later. Each limb bud consists of a mass of mesenchyme covered by ectoderm (Fig. 15-12A and B). The mesenchyme is derived from the somatic layer of the lateral mesoderm. The limb buds elongate by the proliferation of the mesenchyme. Although the early stages of limb development are alike for the upper and lower limbs (Figs. 6-9 and 6-10), there are distinct differences because of their form and function. The upper limb buds develop opposite the caudal cervical segments, whereas the lower limb buds form opposite the lumbar and upper sacral segments.
At the apex of each limb bud, the ectoderm thickens to form an apical ectodermal ridge (AER) (Fig. 15-12A). The AER, a multilayered epithelial structure, interacts with the mesenchyme in the limb bud, promoting outgrowth of the bud. Retinoic acid promotes the formation of the limb bud by inhibiting fibroblast growth factor (FGF8) signaling. The AER exerts an inductive influence on the limb mesenchyme that initiates growth and development of the limbs in a proximodistal axis. Mesenchymal cells aggregate at the posterior margin of the limb bud to form a zone of polarizing activity. Fibroblast growth factors from the AER activate the zone of polarizing activity, causing expression of the sonic hedgehog gene (Shh), which controls the patterning of the limb along the anteroposterior axis. Expression of Wnt7 from the dorsal epidermis of the limb bud and engrailed-1 (En-1) from the ventral aspect is involved in specifying the dorsoventral axis. Curiously, the AER itself is maintained by inductive signals from Shh and Wnt7. The mesenchyme adjacent to the AER consists of undifferentiated, rapidly proliferating cells, whereas the mesenchymal cells proximal to it differentiate into blood vessels and cartilage bone models. The distal ends of the limb buds eventually flatten into hand- and foot-plates (Fig. 15-19).
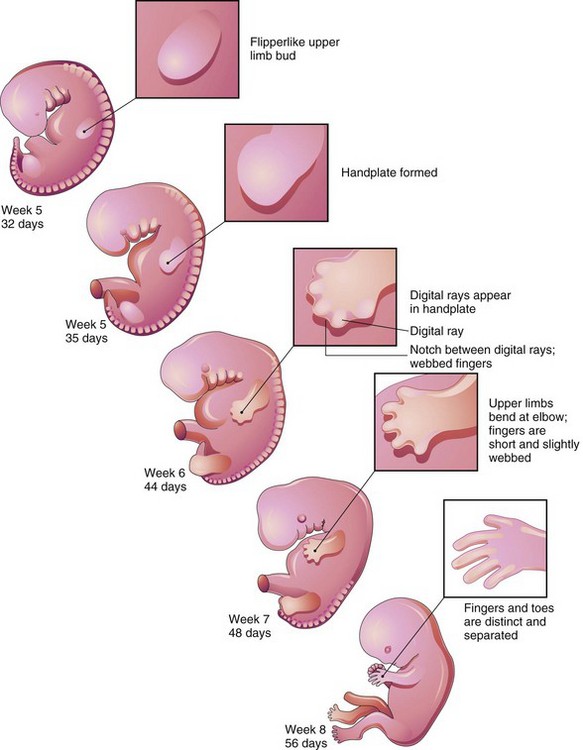
Figure 15–19 Development of the limbs (32-56 days). Note that development of the upper limbs precedes that of the lower limbs.
By the end of the sixth week of development, mesenchymal tissue in the handplates has condensed to form finger buds—digital rays—(Figs. 15-19 and 15-20A to C), which outline the pattern of the digits. During the seventh week, similar condensations of mesenchyme in foot plates form toe buds—digital rays—(Fig. 15-20G to I). At the tip of each digital ray, a part of the AER induces development of the mesenchyme into the mesenchymal primordia of the bones (phalanges) in the digits. The intervals between the digital rays are occupied by loose mesenchyme. Soon the intervening regions of mesenchyme undergo apoptosis (programmed cell death), forming notches between the digital rays (see Figs. 15-19 and 15-20D and J). As this tissue breakdown progresses, separate digits are produced by the end of the eighth week of development (see Fig. 15-19). Blocking of cellular and molecular events during this process may account for webbing, or fusion, of the fingers or toes, a condition known as syndactyly (see Fig. 15-25C and D).
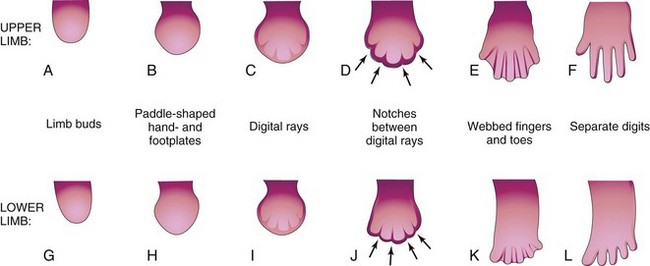
Figure 15–20 Development of the hands and feet between the fourth and eighth weeks. The early stages of limb development are similar, except that development of the hands precedes that of the feet by approximately 1 day. A, At 27 days. B, At 32 days. C, At 41 days. D, At 46 days. E, At 50 days. F, At 52 days. G, At 28 days. H, At 36 days. I, At 46 days. J, At 49 days. K, At 52 days. L, At 56 days. The arrows in D and J indicate the tissue breakdown processes that separate the fingers and toes.
Final Stages of Limb Development
The mesenchyme in the limb bud gives rise to bones, ligaments, and blood vessels (Fig. 15-12). As the limb buds elongate during the early part of the fifth week of development, mesenchymal models of the bones are formed by cellular aggregations (see Fig. 15-12A and B). Chondrification centers appear later in the fifth week. By the end of the sixth week, the entire limb skeleton is cartilaginous (see Fig. 15-12C and D).
Osteogenesis of the long bones begins in the seventh week from primary ossification centers in the diaphyses of the long bones. Primary ossification centers are present in all long bones by the 12th week. Primary ossification of the carpal (wrist) bones begins during the first year after birth.
From the dermomyotome regions of the somites, myogenic precursor cells also migrate into the limb bud and later differentiate into myoblasts, the precursors of muscle cells. As the long bones form, myoblasts aggregate and form a large muscle mass in each limb bud (Fig. 15-16). In general, this muscle mass separates into dorsal (extensor) and ventral (flexor) components.
Early in the seventh week, the limbs extend ventrally and the preaxial and postaxial borders are cranial and caudal, respectively (see Fig. 15-22A and D). The upper limbs rotate laterally through 90 degrees on their longitudinal axes; thus, the future elbows point dorsally and the extensor muscles lie on the lateral and posterior aspects of the limb. The lower limbs rotate medially through almost 90 degrees; thus, the future knees face ventrally and the extensor muscles lie on the anterior aspect of the lower limb (Fig. 15-21A to D).
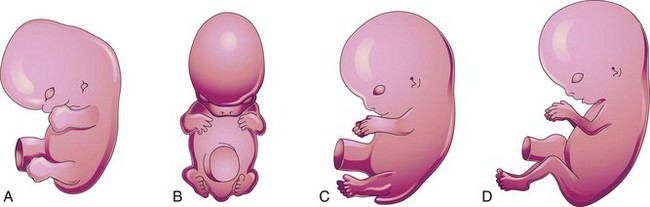
Figure 15–21 Positional changes of the developing limbs of human embryos. A, At approximately 48 days, showing the limbs extending ventrally and the handplates and footplates facing each other. B, At approximately 51 days, showing the upper limbs bent at the elbows and the hands curved over the thorax. C, At approximately 54 days, showing the soles of the feet facing medially. D, At approximately 56 days. Note that the elbows now point caudally and the knees, cranially.
The radius and the tibia are homologous bones, as are the ulna and the fibula, just as the thumb and the great toe are homologous digits. Synovial joints appear at the beginning of the fetal period, coinciding with functional differentiation of the limb muscles and their innervation.
Cutaneous Innervation of Limbs
Motor axons arising from the spinal cord enter the limb buds during the fifth week of development and grow into the dorsal and ventral muscle masses. Sensory axons enter the limb buds after the motor axons and use them for guidance. Neural crest cells, the precursors of Schwann cells, surround the motor and sensory nerve fibers in the limbs and form the neurolemmal and myelin sheaths (see Chapter 16).
A dermatome is the area of skin supplied by a single spinal nerve and its spinal ganglion. During the fifth week of development, the peripheral nerves grow from the developing limb (brachial and lumbosacral) plexuses into the mesenchyme of the limb buds (Fig. 15-22A and B). The spinal nerves are distributed in segmental bands, supplying both the dorsal and the ventral surfaces of the limb buds. As the limbs elongate, the cutaneous distribution of the spinal nerves migrates along the limbs and no longer reaches the surface in the distal part of the limbs. Although the original dermatomal pattern changes during growth of the limbs, an orderly sequence of distribution can still be recognized in the adult (Fig. 15-22C and F). In the upper limb, the areas supplied by C5 and C6 adjoin the areas supplied by T2, T1, and C8, but the overlap between them is minimal at the ventral axial line.
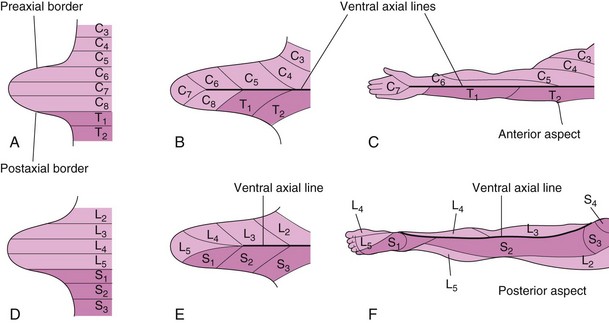
Figure 15–22 Development of the dermatomal patterns of the limbs. The axial lines indicate where there is no sensory overlap. A and D, Ventral aspect of the limb buds early in the fifth week. At this stage, the dermatomal patterns show the primordial segmental arrangement. B and E, Similar views later in the fifth week, showing the modified arrangement of dermatomes. C and F, The dermatomal patterns in the adult upper and lower limbs. The primordial dermatomal pattern has disappeared, but an orderly sequence of dermatomes can still be recognized. In F, note that most of the original ventral surface of the lower limb lies on the back of the adult limb. This results from the medial rotation of the lower limb that occurs toward the end of the embryonic period. In the upper limb, the ventral axial line extends along the anterior surface of the arm and forearm. In the lower limb, the ventral axial line extends along the medial side of the thigh and knee, to the posteromedial aspect of the leg to the heel.
Because there is overlapping of dermatomes, a particular area of skin is not exclusively innervated by a single segmental nerve. The limb dermatomes may be traced progressively down the lateral aspect of the upper limb and back up its medial aspect. A comparable distribution of dermatomes occurs in the lower limbs and may be traced down the ventral aspect and then up the dorsal aspect of the lower limb. When the limbs extend and rotate, they carry their nerves with them; this explains the oblique course of the nerves arising from the brachial and lumbosacral plexuses.
Blood Supply to Limbs
The limb buds are supplied by branches of the intersegmental arteries (Fig. 15-23A), which arise from the dorsal aorta and form a fine capillary network throughout the mesenchyme. The primordial vascular pattern consists of a primary axial artery and its branches (Fig. 15-23B and C), which drain into a peripheral marginal sinus. Blood in the sinus drains into a peripheral vein.
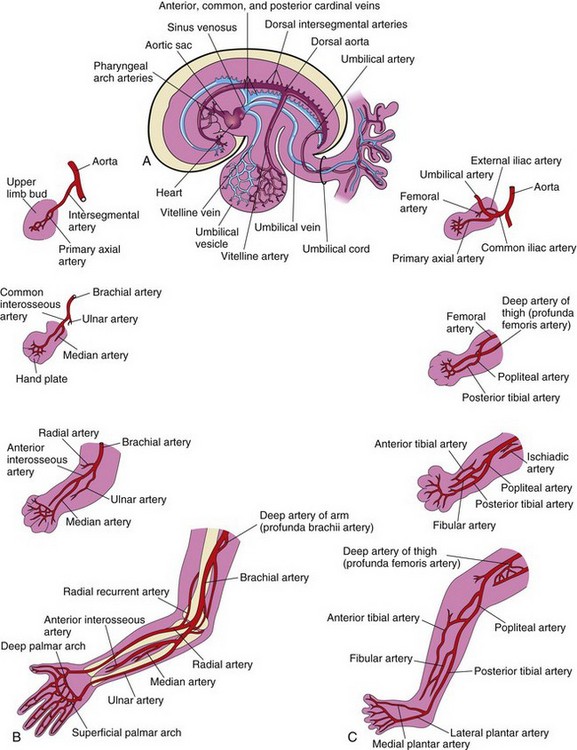
Figure 15–23 Development of the limb arteries. A and B, Development of the arteries in the upper limb. C, Development of the arteries in the lower limb.
The vascular pattern changes as the limbs develop, chiefly as a result of vessels sprouting from existing vessels (angiogenesis). The new vessels coalesce with other sprouts to form new vessels. The primary axial artery becomes the brachial artery in the arm and the ulnar and radial arteries in the forearm, its terminal branches of the brachial artery (see Fig. 15-23B). As the digits form, the marginal sinus breaks up and the final venous pattern, represented by the basilic and cephalic veins and their tributaries, develops. In the thigh, the primary axial artery is represented by the deep artery of the thigh (profunda femoris artery). In the leg, the primary axial artery is represented by the anterior and posterior tibial arteries (Fig. 15-23C).
Cleft Hand and Cleft Foot
In the rare cleft hand or cleft foot abnormalities, one or more central digits are absent—ectrodactylyl—resulting from failure of one or more digital rays to develop (Fig. 15-24A and B). The hand or foot is divided into two parts that oppose each other. The remaining digits are partially or completely fused (syndactyly).
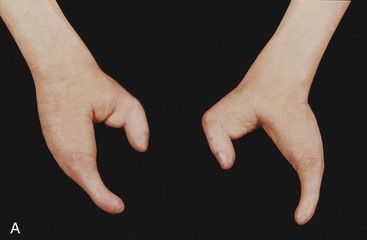
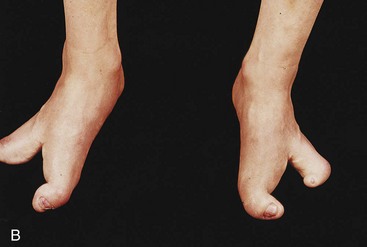
Figure 15–24 Anomalies of the hands and feet. A, Ectrodactyly in a child. Note the absence of the central digits of the hands, resulting in split hands. B, A similar type of defect involving the feet. These limb defects can be inherited in an autosomal dominant pattern.
(Courtesy of A.E. Chudley, M.D., Section of Genetics and Metabolism, Department of Pediatrics and Child Health, University of Manitoba, Children’s Hospital, Winnipeg, Manitoba, Canada.)
Congenital Absence of Radius
In some individuals, the radius is partially or completely absent. The hand deviates laterally (radially), and the ulna bows with the concavity on the lateral side of the forearm. This anomaly results from failure of the mesenchymal primordium of the radius to form during the fifth week of development. Absence of the radius is usually caused by genetic factors.
Polydactyly
Supernumerary digits are common (Fig. 15-25A and B). Often, the extra digit is incompletely formed and lacks proper muscular development, rendering it useless. If the hand is affected, the extra digit is most commonly medial or lateral rather than central. In the foot, the extra toe is usually on the lateral side. Polydactyly is inherited as a dominant trait.
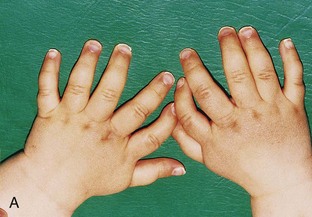
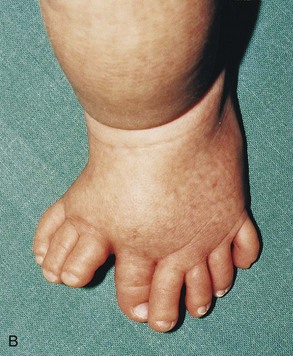
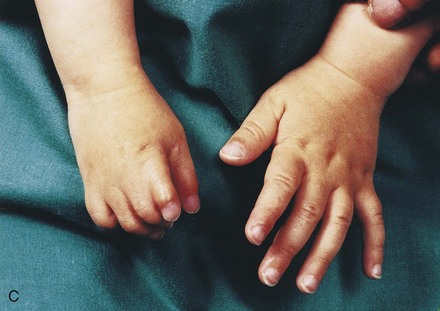
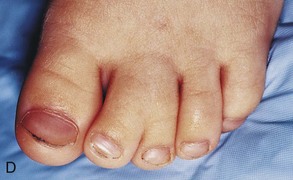
Figure 15–25 Various types of digital defects. A, Polydactyly of the hands. B, Polydactyly of the foot. This condition results from the formation of one or more extra digital rays during the embryonic period. C and D, Various forms of syndactyly involving the fingers and toes. Cutaneous syndactyly (C) is probably caused by incomplete apoptosis in the tissues between the digital rays during embryonic life. Syndactyly of the second and third toes is shown in D. In osseous syndactyly, the digital rays merge as a result of lack of apoptosis, causing fusion of the bones.
(Courtesy of A.E. Chudley, M.D., Section of Genetics and Metabolism, Department of Pediatrics and Child Health, University of Manitoba, Children’s Hospital, Winnipeg, Manitoba, Canada.)
Syndactyly
Syndactyly occurs with a frequency of approximately 1 in 2200 births. Cutaneous syndactyly (simple webbing of the digits) is the most common limb anomaly (see Fig. 15-25C). It occurs more frequently in the foot than in the hand (see Fig. 15-25C and D). Syndactyly is most frequently observed between the third and fourth fingers and between the second and third toes (see Fig. 15-25D). It is inherited as a simple dominant or simple recessive trait. Cutaneous syndactyly results from failure of the webs to degenerate between two or more digits. In some cases, there is fusion of the bones (synostosis). Osseous syndactyly occurs when the notches between the digital rays do not develop during the seventh week; as a result, separation of the digits does not occur.
Arthrogryposis
Arthrogryposis multiplex congenita refers to a heterogeneous group of musculoskeletal disorders characterized by multiple contractures and immobility of two or more joints from birth. The incidence of arthrogryposis multiplex congenita is 1 in 3000 live births and males are more affected in sex-linked cases. The causes may be both neurological (central and peripheral nervous system abnormalities) and nonneurological (cartilaginous abnormalities and restricted movement in utero).
Congenital Talipes (Clubfoot)
Talipes occurs at a rate of approximately 1 in 1000 births. Talipes equinovarus, the most common type (Fig. 15-26), occurs approximately twice as frequently in males as in females. The sole of the foot is turned medially, and the foot is inverted. There is much uncertainty about the cause of talipes. Hereditary factors are involved in some cases, and it appears that environmental factors are involved in most cases. Talipes appears to follow a multifactorial pattern of inheritance; hence, any intrauterine position that results in abnormal positioning of the feet may cause talipes if the fetus is genetically predisposed to this deformity.
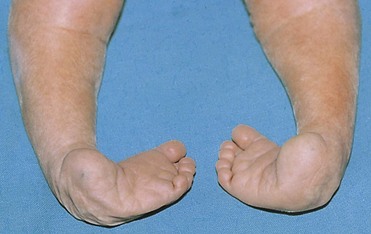
Figure 15–26 Neonate with bilateral talipes equinovarus deformities (club feet), illustrating the classic type of this anomaly, characterized by inversion and medial rotation of the soles of the feet.
(Courtesy of A.E. Chudley, M.D., Section of Genetics and Metabolism, Department of Pediatrics and Child Health, University of Manitoba, Children’s Hospital, Winnipeg, Manitoba, Canada.)
Limb Anomalies
There are two main types of limb anomalies:
Terms such as hemimelia, peromelia, ectromelia, and phocomelia are also used.
Anomalies of the limbs originate at different stages of development. Suppression of limb bud development during the early part of the fourth week results in amelia (Fig. 15-27A). Arrest or disturbance of the differentiation or growth of the limbs during the fifth week results in meromelia (Fig. 15-27B and C). Some limb defects are caused by the following:
• Genetic factors, such as chromosomal abnormalities associated with trisomy 18 (Chapter 19).
• Mutant genes, as in brachydactyly (shortness of digits) or osteogenesis imperfecta (connective tissue disorders). Molecular studies have implicated gene mutation (Hox gene, BMP, Shh, Wnt7, En-1, and others) in some cases of limb anomalies.
• Environmental factors, such as teratogens (e.g., thalidomide).
• A combination of genetic and environmental factors (multifactorial inheritance), as in congenital dislocation of the hip.
• Vascular disruption and ischemia, as in limb reduction defects.
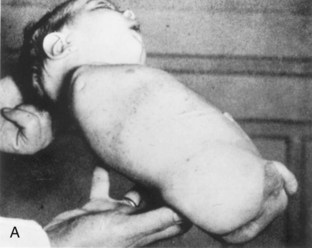
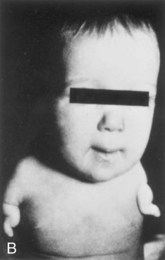
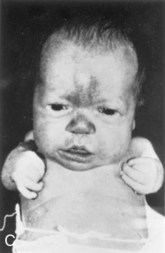
Figure 15–27 Limb anomalies due to thalidomide. A, Quadruple amelia (absence of the upper and lower limbs). B, Meromelia (partial absence) of the upper limbs. The limbs are represented by rudimentary stumps. C, Meromelia in which the rudimentary upper limbs are attached directly to the trunk.
(From Lenz W, Knapp K: Foetal malformations due to thalidomide. Ger Med Mon 7:253, 1962.)
Clinically Oriented Questions
1. Occasionally, accessory ribs are associated with the seventh cervical vertebra and the first lumbar vertebra. Are these accessory ribs of clinical importance? What is the embryologic basis of accessory ribs?
2. What vertebral defect can produce scoliosis? Define this condition. What is the embryologic basis of the vertebral defect?
3. What is meant by the term craniosynostosis? What results from this developmental abnormality? Give a common example and describe it.
4. A child presented with characteristics of Klippel-Feil syndrome. What are the main features of this condition? What vertebral anomalies are usually present?
5. A newborn infant was born with prune-belly syndrome caused by failure of the abdominal musculature to develop normally. What do you think would cause this congenital anomaly? What urinary anomaly results from abnormal development of the anterior abdominal wall?
6. A boy asked his mother why one of his nipples was much lower than the other one. She was unable to explain this anomaly. How would you explain the abnormally low position of the nipple?
7. An 8-year-old girl asked her doctor why the muscle on one side of her neck was so prominent. What would you tell her? What would happen if this muscle were not treated?
8. After strenuous exercise, a young athlete complained of pain on the posteromedial aspect of his ankle. He was told that he had an accessory calf muscle. Is this possible? If so, what is the embryologic basis of this anomaly?
9. An infant had short limbs. His trunk was normally proportioned, but his head was slightly larger than normal. Both parents had normal limbs, and these problems had never occurred in either of their families. Could the mother’s ingestion of drugs during pregnancy have caused these abnormalities? If not, what would be the probable cause of these skeletal disorders? Could they occur again if the couple had more children?
10. A woman is interested in marrying a man who happens to have very short fingers (brachydactyly). He says that two of his relatives have short fingers, but none of his brothers or sisters has them. The woman has normal digits, and so does everyone else in her family. Clearly, heredity is involved, but what are the chances that the couple’s children would have brachydactyly if they were to marry?
11. Approximately 1 year ago, a woman was reported to have given birth to a child with no right hand. She had taken a drug that contained doxylamine and dicyclomine to alleviate nausea during the 10th week of her pregnancy (8 weeks after fertilization). The woman is instituting legal proceedings against the company that makes the drug. Does this drug cause limb defects? If it does, could it have caused failure of the child’s hand to develop?
12. An infant had syndactyly of the left hand and absence of the left sternal head of the pectoralis major muscle. The infant seemed normal, except that the nipple on the left side was approximately 2 inches lower than the other one. What is the cause of these anomalies? Can they be corrected?
13. What is the most common type of clubfoot? How common is it? What is the appearance of the feet of infants born with this anomaly?