CHAPTER 59
REGULATION OF BODY TEMPERATURE
John Stitt
HEAT AND TEMPERATURE: THE ADVANTAGES OF HOMEOTHERMY
Homeothermy enables an organism to maintain its activity over a wide range of environmental temperatures
The ability to regulate internal body temperature has provided higher organisms independence from the environment. Because the rates of most physical and chemical reactions depend on temperature, most physiological functions are sensitive to temperature changes. Thus, the activity levels of poikilotherms (species that do not regulate internal body temperature) generally depend on environmental temperature, whereas those of homeotherms (species that do regulate internal body temperature) are relatively stable over a broad range of ambient conditions. A lizard, for example, is capable of relatively less movement away from its lair on a cold, overcast day than on a hot, sunny day, whereas a prairie dog may be equally mobile on either day. An arctic fox acclimatizes to the extreme cold of winter by maintaining a thick, insulating coat that enables it to resist body cooling and minimizes the necessity to increase metabolic heat generation, which would require increased food intake.
The thermoregulatory system of homeotherms creates an internal environment in which reaction rates are relatively high and optimal. At the same time, an effective thermoregulatory system avoids the pathologic consequences of wide deviations in body temperature (Table 59-1). The thermoregulatory system incorporates both anticipatory controls and negative feedback controls. The components of this system are as follows: (1) thermal sensors; (2) afferent pathways; (3) an integration system in the central nervous system (CNS); (4) efferent pathways; and (5) target organs that control heat generation and transfer, such as skeletal muscle (e.g., shivering to generate heat), circulation to the skin (to dissipate heat), and the sweat glands (to dissipate heat).
Table 59-1 Consequences of Deviations in Body Temperature
Temperature (°C) |
Consequence |
40-44 |
Heat stroke with multiple organ failure and brain lesions |
38-40 |
Hyperthermia (as a result of fever or exercise) |
36-38 |
Normal range |
34-36 |
Mild hypothermia |
30-34 |
Impairment of temperature regulation |
27-29 |
Cardiac fibrillation |
The focus of this chapter is temperature regulation in homeotherms. I examine the physical aspects of heat transfer both within the body and between body and environment, as well as the physiological mechanisms involved in altering these rates of transfer. Finally, I look at the consequences of extreme challenges to the thermoregulatory mechanism, such as hyperthermia, hypothermia, and dehydration.
Body core temperature depends on time of day, physical activity, time in the menstrual cycle, and age
Temperature scales are relative scales of heat content. The centigrade scale is divided into 100 equal increments, referenced to the freezing (0°C) and boiling (100°C) points of water. The “normal” body temperature of an adult human is approximately 37°C (i.e., 98.6°F), but it may be as low as 36°C or as high as 37.5°C in active, healthy people. Body temperature usually refers to the temperature of the internal body core, measured under the tongue (sublingually), in the ear canal, or in the rectum. For clinical purposes, the most reliable (although the least practical) among these three is the last, because it is least influenced by ambient (air) temperature. Measurement devices range from traditional mercury-in-glass thermometers to electronic, digital read-out thermistors. Nearly all such instruments are accurate to 0.1°C. The least invasive approach uses an infrared thermometer to measure the radiant temperature over the temporal artery. (See Note: Body Core Temperature)
Body core temperature depends on the time of day, the stage of the menstrual cycle in women, the level of the person’s activity, and the individual’s age. All homeotherms maintain a circadian (24-hour cycle) body temperature rhythm, with variations of ~1°C. In humans, body temperature is usually lowest between 3:00 to 6:00 am, and it peaks at 3:00 to 6:00 pm. This circadian rhythmicity is inherent in the autonomic nervous system, independent of the sleep-wakefulness cycle, but it is entrained by light-dark cues to a 24-hour cycle. (See Note: Infrared Thermometry)
In many women, body temperature increases approximately 0.5°C during the postovulatory phase of their menstrual cycle (see Chapter 55). An abrupt increase in body temperature of 0.3°C to 0.5°C accompanies ovulation and may be useful as a fertility guide.
Physical activity generates excess heat as a byproduct of elevated metabolic rate. A portion of this excess heat remains in the body, causes the core temperature to rise, and triggers appropriately matching heat loss responses. Core temperature remains elevated during activity and for an extended period after exercise ceases.
Infants and older people are less able to maintain a normal body temperature than are the rest of the population, particularly in the presence of external challenges. Newborns do not readily shiver or sweat and thus behave more like poikilotherms than like homeotherms. These properties, along with a high surface-to-mass ratio, render infants more susceptible to fluctuations in core temperature when exposed to a hot or cold environment. Older people are also subject to greater fluctuations in core temperature. Aging is associated with a progressive deficit in the ability to sense heat and cold, as well as a reduced ability to generate heat (reduced metabolic rate and metabolic potential because of lower muscle mass) and to dissipate heat (reduced cardiovascular reserve and sweat gland atrophy from disuse).
The body’s rate of heat production can vary from ~70 kcal/hr at rest to 600 kcal/hr during jogging
The body’s rate of heat production is closely linked to the rate of metabolism, the rate of O2 consumption ( O2). Minor variations occur, depending on the mixture of fuels (foods) being oxidized, a process that determines the respiratory quotient (RQ; see Table 58-6). Because of their inherent inefficiency, metabolic transformations generate heat (see Chapter 58). Ultimately, all the energy contained in fuels appears as heat, mass storage or growth, or physical work done on the environment.
O2). Minor variations occur, depending on the mixture of fuels (foods) being oxidized, a process that determines the respiratory quotient (RQ; see Table 58-6). Because of their inherent inefficiency, metabolic transformations generate heat (see Chapter 58). Ultimately, all the energy contained in fuels appears as heat, mass storage or growth, or physical work done on the environment.
The body’s metabolic rate, and thus its rate of heat production, is not constant. The resting metabolic rate (RMR; see Chapter 58) is the rate necessary to maintain the functions of resting cells; these functions include active transport as well as cardiac and respiratory muscle activity. Voluntary or involuntary (e.g., shivering) muscular activity adds to the overall metabolic heat production. Even digesting a meal increases the metabolic rate (see Chapter 58). An increase in tissue temperature itself raises the metabolic rate, according to the van’t Hoff relation (i.e., a 10°C increase in tissue temperature more than doubles the metabolic rate). Furthermore, certain hormones, notably thyroxine and epinephrine, increase the cellular metabolic rate. Because the body’s heat production rate is variable, the rate of heat loss must match it closely if the body temperature is to remain constant. At an RQ of 0.8 (see Chapter 58), the average person under sedentary (i.e., RMR) conditions has a resting O2 of 250 mL/min, which corresponds to a heat production of 72 kcal/hr (~85 watts). In other words, an adult of average size generates the heat of an 85-watt light bulb (see Chapter 58).
During physical exercise, the rate of energy production—and hence, heat generation—increases in proportion to the intensity of exercise. An average adult can comfortably sustain an energy production rate of 400 to 600 kcal/hr for extended periods (e.g., a fast walk or a modest jog). Nearly all this increased heat generation occurs in active skeletal muscle, although a portion arises from increased cardiac and respiratory muscle activity. A thermal load of this magnitude would raise core temperature by 1.0°C every 8 to 10 minutes if the extra heat could not escape the body. Physical activity would be limited to 25 to 30 minutes, at which time the effects of excessive hyperthermia (>40°C) would begin to impair body function. This impairment, of course, does not occur, primarily because of the effectiveness of the thermoregulatory system. Within a relatively short period, the increase in body temperature resulting from exercise leads to an increased rate of heat dissipation that matches heat production. Thereafter, the body maintains a new, elevated steady temperature. When exercise ceases, body temperature gradually decreases to its pre-exercise level.
MODES OF HEAT TRANSFER
Maintaining a relatively constant body temperature requires a fine balance between heat production and heat losses
Temperature homeostasis requires that increases or decreases in heat production balance increases or decreases in heat loss. Physiologists usually express this concept in terms of a whole-body heat balance equation, which, for an adult of constant mass, is as follows:

All terms in the foregoing equation have the units kcal/hr.
Several physiological processes contribute to temperature homeostasis, including modulation of metabolic heat production, physical heat transfer, and elimination of heat. These processes operate at the level of cells, tissues, and organ systems. Let us discuss in order the terms in Equation 59-1.
The rate of metabolism (M) arises from the cellular oxidation of carbohydrates, fats, and proteins. Because of their inherent inefficiency, metabolic transformations generate heat (see Chapter 58). Table 59-2 shows the fractional contributions of different body systems to total heat production under sedentary, resting conditions.
Table 59-2 Contribution of Body Systems to Resting Metabolism
System |
RMR |
Respiration and circulation |
15% |
CNS and nerves |
20% |
Musculature (at rest) |
20% |
Abdominal viscera |
45% |
RMR |
100% (~70 kcal/hr) |
Heat production by skeletal muscle can play a vital emergency role in temperature regulation in the cold. Shivering—the rhythmic, clonic activation of the major muscle masses surrounding the head, torso, and upper limbs—can increase total body heat production by as much as 400%. The body can grade the intensity of shivering and heat production to match heat loss. Physical exercise can generate even more heat than shivering. Under conditions of maximal exercise, O2max may correspond to a total energy expenditure (M in Equation 59-1) of 1300 kcal/hr for an endurance athlete. If 75% to 80% of this energy evolves as heat—so that the athlete does ~300 kcal/hr of useful work on the environment (W)—the rate of heat production (H = M − W) would be 1000 kcal/hr (~1200 watts) for a brief period of time. This change is equivalent to changing from an 85-watt light bulb to a 1200-watt space heater. Unless the body can dissipate this heat, death from hyperthermia and heat stroke (see box on Heat Stroke) will ensue rapidly.
Virtually all heat leaving the body must exit through the skin surface. In the following three sections, the three major routes of heat elimination are discussed: radiation (R), convection (C), and evaporation (E). As the heat balance equation shows, the difference between (M − W) and (R + C + E) is the rate of heat storage (S) within the body. The value of S may be positive or negative, depending on whether (M − W) > (R + C + E) or vice versa. A positive value of S results in a rise of body core temperature, and a negative value results in a fall.
Heat moves from the body core to the skin, primarily by convection
Generally, all heat production occurs within the body’s tissues, and all heat elimination occurs at the body surface. Figure 59-1 illustrates a passive system in which heat flows depend on the size, shape, and composition of the body, as well as on the laws of physics. The circulation carries excess heat away from active tissues, such as muscle, to the body core—represented by the heart, lungs, and their central circulating blood volume. How does the body prevents its core from overheating? The answer is that the core transfers this heat to a dissipating heat sink. The organ serving as the body’s greatest potential heat sink is the relatively cool skin, which is the largest organ in the body. Only a minor amount of the body’s generated heat flows directly from the underlying body core to the skin by conduction across the body tissues. Most of the generated body heat flows to the skin by convection in the blood, and blood flow to the skin can increase markedly. There, nearly all the heat transferred to the skin must flow to the environment, facilitated by the skin’s large surface area. (See Note: Heat Transfer from Muscle to Body Core)
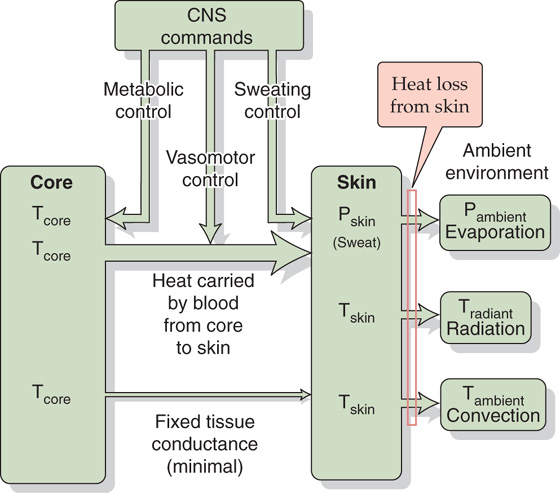
Figure 59-1 Passive or unregulated heat transfer. In the steady state, the rate of heat production by the body core must be matched by the flow of heat from the core to the skin, and from skin to environment. Various homeostatic controls—systems not directly involved in temperature regulation—can affect heat flow. Examples include sweating in response to hypoglycemia, changes in blood flow patterns in response to a fall in blood pressure, and changes in metabolism in response to alterations in thyroid metabolism.
The transfer of heat from core to skin occurs by two routes:
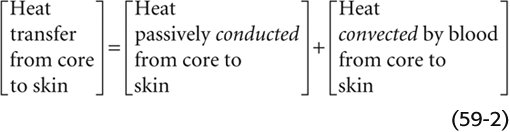
Both the conduction and convection terms in the previous equation are proportional to the temperature gradient from core to skin (Tcore − 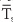 skin), where skin is the average temperature of at least four representative skin sites. The proportionality constant for passive conduction across the subcutaneous fat (the body’s insulation) is relatively fixed. However, the proportionality constant for heat convection by blood is a variable term, reflecting the variability of the blood flow to the skin. The ability to alter skin blood flow, under autonomic control, is therefore the primary determinant of heat flow from core to skin. The capacity to limit blood flow to the skin is an essential defense against body cooling in the cold (hypothermia). A side effect, however, is that skin temperature falls. Conversely, the capacity to elevate skin blood flow is an essential defense against hyperthermia. On very hot days when skin temperature may be very high and close to body core temperature, even high skin blood flow may not be adequate to transfer sufficient heat to allow body core temperature to stabilize because the temperature gradient (Tcore − skin) is too small.
skin), where skin is the average temperature of at least four representative skin sites. The proportionality constant for passive conduction across the subcutaneous fat (the body’s insulation) is relatively fixed. However, the proportionality constant for heat convection by blood is a variable term, reflecting the variability of the blood flow to the skin. The ability to alter skin blood flow, under autonomic control, is therefore the primary determinant of heat flow from core to skin. The capacity to limit blood flow to the skin is an essential defense against body cooling in the cold (hypothermia). A side effect, however, is that skin temperature falls. Conversely, the capacity to elevate skin blood flow is an essential defense against hyperthermia. On very hot days when skin temperature may be very high and close to body core temperature, even high skin blood flow may not be adequate to transfer sufficient heat to allow body core temperature to stabilize because the temperature gradient (Tcore − skin) is too small.
Although most of the heat leaving the core moves to the skin, a small amount also leaves the body core by the evaporation of water from the respiratory tract. The evaporative rate is primarily a function of the rate of ventilation (see Chapter 31), which, in turn, increases linearly with the metabolic rate over a wide range of exercise intensities.
Heat moves from the skin to the environment by radiation, conduction, convection, and evaporation
Figure 59-2 is a graphic summary of the heat balance equation (Equation 59-1) for an athlete exercising in an outdoor environment. This illustration depicts the movement of heat within the body, its delivery to the skin surface, and its subsequent elimination to the environment by radiation, convection, and evaporation.
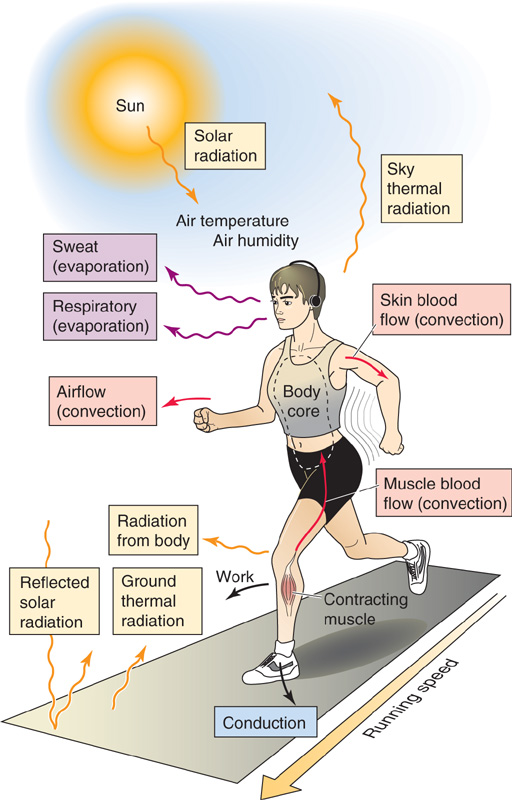
Figure 59-2 Model of energy transfer from the body to the environment.
Radiation Heat transfer by radiation occurs between the skin and solid bodies in the environment. The infrared portion of the electromagnetic energy spectrum carries this energy, which is why infrared cameras can detect warm bodies at night. The body gains or loses heat by radiation at a rate that is proportional to the temperature difference between the skin and the radiating body:

R is positive when the body loses heat and negative when it gains heat.
One may not be so aware of radiative heat fluxes to and from the body, particularly when the radiating body temperature (Tradiant) differs from the ambient environmental temperature (Tambient) that tends to dominate our attention. Indoors, Tradiant is the same as Tambient because surrounding objects thermally equilibrate with one another. Outdoors, radiating bodies may be at widely different temperatures. The radiant heat load from the sun to the body on a cloudless summer day may exceed the RMR by a considerable amount. The radiant heat load from a fire or a radiant lamp can provide substantial warming of bodies in the radiant field. Conversely, on a winter evening, the radiant heat loss from the body to a cloudless, dark sky—which has a low radiant temperature—may exceed RMR. Thus, one may feel a sudden chill when walking past an uncurtained window. This chill is caused by the sudden fall in skin temperature owing to increased radiant heat loss. Radiation of heat from the body accounts for ~60% of heat lost when the body is at rest in a neutral thermal indoor environment. A neutral thermal environment is a set of conditions (air temperature, airflow and humidity, and temperatures of surrounding radiating surfaces) in which the temperature of the body does not change when the subject is at rest (i.e., RMR) and is not shivering.
Conduction Heat transfer by conduction occurs when the body touches a solid material of different temperature. For example, lying on the hot sand causes one to gain heat by conduction. Conversely, placing an ice pack on a sore muscle causes heat loss by conduction. However, under most normal circumstances (e.g., when one is standing and wearing shoes or recumbent and wearing clothes), the heat gain or loss by conduction is minimal.
Convection Heat transfer by convection occurs when a fluid such as air or water carries the heat between the body and the environment. The convective heat loss is proportional to the difference between skin and ambient temperature:

C is positive when the body loses heat and negative when it gains heat.
Whereas the radiative heat transfer coefficient (hradiative) is constant, the convective coefficient (hconvective) is variable and can increase up to 5-fold when air velocity is high. Thus, even when (skin − Tambient) is fixed, convective heat loss increases markedly as wind speed increases. In the absence of air movement, the air immediately overlying the skin warms as heat leaves the skin. As this warmer and lighter air rises off the skin, cooler ambient air replaces it and, in turn, is warmed by the skin. This is the process of natural convection. However, with forced air movement, such as by wind or a fan, the cooler “ambient” air replaces the warmer air overlying the skin more rapidly. This change increases the effective convective heat transfer from the skin, even though the temperature of the ambient air is unchanged. This is a process of forced convection, which underlies the wind chill factor.
Evaporation Humans can dissipate nearly all the heat produced during exercise by evaporating sweat from the skin surface. The evaporative rate is independent of the temperature gradient between skin and environment. Instead, it is proportional to the water vapor pressure gradient between skin and environment:

E is positive when the body loses heat by evaporation and negative when it gains heat by condensation.
The evaporation of 1 g of water removes ~0.58 kcal from the body. Because the body’s sweat glands can deliver up to 30 g fluid/min or 1.8 L/hr to the skin surface, evaporation can remove 0.58 × 1800 g or ~1000 kcal/hr. Thus, under ideal conditions (i.e., when ambient humidity is sufficiently low to allow efficient evaporation), evaporation could theoretically remove nearly all the heat produced during heavy exercise. As with convection, increased air velocity over the skin increases the effective vapor pressure gradient between skin and the overlying air because of the faster movement of water vapor away from the skin.
The efficiency of heat transfer from the skin to the environment depends on both physiological and environmental factors. If ambient humidity is high, the gradient of water vapor pressure between skin and air will be low, thus slowing evaporation and increasing the body’s tendency to accumulate excess heat produced during exercise. This phenomenon underlies the temperature humidity index (heat index). Conversely, if ambient humidity is low, as in the desert, net heat loss from the body by evaporation will occur readily, even when ambient temperature exceeds skin temperature and the body is gaining heat by radiation and convection.
When the body is immersed in water, nearly all heat exchange occurs by convection, because essentially no exchanges can occur by radiation or evaporation. Because of the high conductivity and thermal capacity of water, the heat transfer coefficient (hconvective) is ~100 times greater than that of air. Thus, rate of heat exchange underwater is much greater than it is in air. It is therefore not surprising that nearly all the deaths in the Titanic shipwreck disaster resulted from hypothermia in the cold Atlantic waters, rather than from drowning.
When heat gain exceeds heat loss, body core temperature rises
With a knowledge of the transfer coefficients—hradiative (Equation 59-3), hconvective (Equation 59-4), and hevaporative (Equation 59-5)—and the gradients of temperature and water vapor pressure between the skin and environment, we can calculate the body heat fluxes (R, C, and E). Knowing M (computed from O2 by indirect calorimetry; see Chapter 49) and W (if any), we can use the heat balance equation (Equation 59-1) to calculate the heat storage (S). From this value, we can predict the rate of change in mean body temperature:
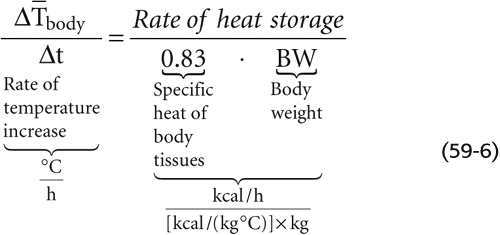
We can verify the accuracy of this predicted rate of change in body by comparing it to the body measured by direct thermometry, using a weighted average of the measured Tcore and average Tskin. (See Note: Mean Body Temperature)
The body has to deal with two types of heat loads that tend to make its temperature rise. In the heat balance equation (Equation 59-1), the term (M – W) constitutes an internal heat load. In contrast, the term (R + C + E)—normally representing a net heat loss—can represent an external heat load if either the radiation or convection terms are heat gains rather than heat losses. Thus, if we stand in the sun and Tradiant exceeds skin (Equation 59-3), we experience a radiant heat load. If we stand in a hot sauna and Tambient exceeds skin (Equation 59-4), we experience a convective heat load. Clearly, both internal and external heat loads can result in net heat storage and thus a rise in body temperature. Changes in environmental temperature (Tradiant and Tambient) exert their influence from the outside, through the body surface. If, starting from relatively low values, Tradiant or Tambient rises, at first the rate at which heat leaves the body decreases, so core temperature tends to rise. Further increases in environmental temperature produce a frank heat load rather than a loss.
Conversely, metabolism produces heat inside the body. For the athlete, all the terms of the heat balance equation are important because dissipating the thermal load is essential for prolonging exercise. The clinician must understand these principles to treat thermally related illnesses. For example, excessive heat exposure can lead to heat exhaustion, in which core temperature rises to as high as 39°C because the body cannot dissipate the heat load. The causes are dehydration (which reduces sweating) and hypovolemia (which reduces blood flow from muscle to core to skin). Heat exhaustion is the most common temperature-related abnormality in athletes. In more severe cases, excessive heat can lead to heat stroke (see the box on Heat Stroke), in which core temperature rises to 41°C or more, as a result of impaired thermoregulatory mechanisms.
Clothing insulates the body from the environment and limits heat transfer from the body to the environment
Placing one or more layers of clothing between the skin and the environment insulates the body and retards heat transfer between the core and the environment. In the presence of clothing, heat transfer from a warmer body to a cooler environment occurs by the same means as without clothing (i.e., radiation, conduction, convection, and evaporation), but from the clothing surface rather than from the skin surface. The insulating effect of clothing is described by the clo unit. By definition, one clo is the insulation necessary to maintain a resting person at a thermal steady state in comfort at 21°C with minimal air movement. Obviously, clo units increase with a greater area of skin coverage by clothing or with thicker clothing.
ACTIVE REGULATION OF HEAT TRANSFER
The body actively regulates its temperature by a feedback system that includes temperature sensors, afferent nerve fibers that carry sensory information to the brain, a hypothalamic control center, efferent nerve fibers that are principally part of the autonomic nervous system, and thermal effectors that either control heat transfer between the body and environment or regulate the body’s rate of heat production. This active system contrasts with the passive system.
Thermal sensors in the skin and in the body core (mainly the hypothalamus) respond to changes in theirlocal temperature
The body has specialized sensory neurons that provide the CNS with information about the body’s thermal condition. These thermosensitive elements are free nerve endings that are distributed over the entire skin surface. These elements are also present within the body core, at particularly high densities in the preoptic area and anterior hypothalamus. (See Note: Preoptic Area and Anterior Hypothalamus)
Skin receptors, although ideal for sensing changes in environmental temperature, do not serve well during exercise because internal temperatures would rise to intolerably high levels before the skin temperature rose to detect this excess heat. Body core thermoreceptors, in contrast, although ideal for detecting changes in core temperature, are inadequate for sensing changes in the environmental temperature. Because of the thermal inertia of the body’s mass, the lag time in using body core sensors to detect externally induced changes in temperature would be too great to achieve effective regulation. Not surprisingly, then, the body is endowed with both peripheral and central thermoreceptors that are integrated within the CNS, to permit a rapid and effective balance of heat loss and heat production while maintaining body core temperature within relatively narrow limits.
Skin Thermoreceptors The entire surface of the body has a network of sensory nerve endings that serve as thermoreceptors. Peripheral thermoreceptors fall into two categories—warmth receptors and cold receptors (Fig. 59-3). Each type is anatomically distinct, and each innervates definable warm-sensitive or cold-sensitive spots on the skin surface (see Chapter 15). Thermal discrimination varies over the surface of the body; it is coarsest on the body trunk and limbs and finest on the face, lips, and fingers. Increasing local temperature, up to 44°C to 46°C, causes warmth receptors to increase their steady firing rate (Fig. 59-3). Cold receptors characteristically increase their steady firing rate as local temperature decreases from ~40°C to 24°C to 28°C. In either case, a sustained temperature change (see Fig. 15-28) may cause a stable change in the sensor’s firing rate (i.e., tonic or static response) or a temporary change (i.e., phasic or dynamic response).
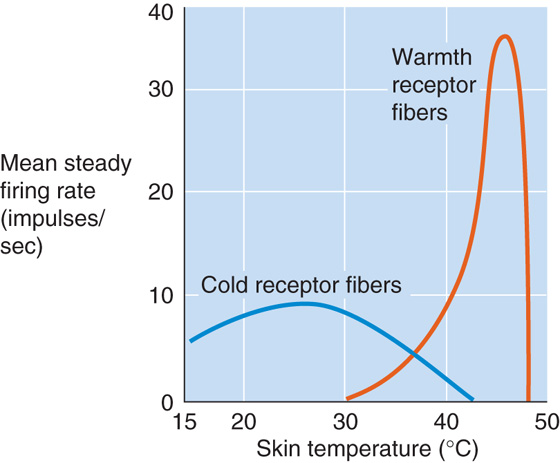
Figure 59-3 Response of warmth and cold receptors to temperature change.
Because of their location, skin thermoreceptors primarily provide the hypothalamic thermoregulatory center with information about ambient temperature (Fig. 59-4). As discussed later, they provide an anticipatory signal in conditions of rapidly changing ambient temperature and allow the autonomic nervous system to exert reflex thermoregulation. Information from skin thermoreceptors also travels through thalamic pathways to the cerebral cortex, thus providing the basis for conscious perception of the thermal environment and appreciation of thermal comfort. We can use this information, for example, to move from the sun to the shade when we sense that we are too hot.
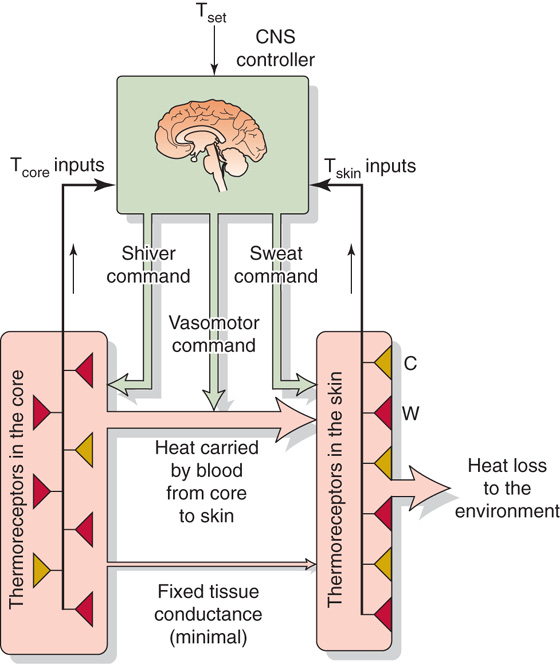
Figure 59-4 Model of negative feedback in temperature regulation.
Body Core Thermoreceptors Thermoreceptors are present in the brain, in the spinal cord, and perhaps in the muscles and major blood vessels. However, the hypothalamus clearly plays the major role in detecting changes in deep body temperature (Fig. 59-4). In the preoptic area and anterior hypothalamus (see Chapter 14), ~10% of neurons will show a positive temperature coefficient when local temperature is cycled over a range of 2°C to 4°C about the mean. Body core thermoreceptors are especially important during exercise, which is one of the few conditions in which the body’s heat production and dissipation rates can differ dramatically and can lead to rapid changes in core temperature. (See Note: Preoptic Area and Anterior Hypothalamus)
The hypothalamic center integrates thermal information and directs changes in efferent activity to modify heat transfer rates
Cooling or warming of the skin alters both the tonic and the phasic components of the activity of cold or warmth receptors (Fig. 59-4). The neural activity of these skin thermoreceptors travels through the spinal cord to the hypothalamus, which integrates thermal information from other parts of the body, including the hypothalamus, compares the prevailing thermal status with an idealized set of thermal conditions, and directs efferent commands to alter the rate of heat generation and to modify heat transfer rates within and from the body.
The skin receptors provide information mainly about environmental temperature, which affects the body’s heat loss rate and could ultimately cause core temperature to change, if the body does not initiate the appropriate thermoregulatory responses to skin cooling or warming. Thus, reflex responses to changes in the average skin temperature may be thought of as anticipatory rather than negative feedback in nature. Moreover, it is impossible to regulate skin temperature because of the skin’s exposure to the ambient environment. However, these anticipatory reflexes are essential elements for an effective thermoregulatory system because the body’s thermal inertia is too great to rely on central receptors alone. For example, low skin temperature—enhanced by cutaneous vasoconstriction in the cold—ensures a rapid and continuous cold signal that maintains a drive for shivering and thus thermogenesis. Conversely, thermoregulatory responses to changes in core (i.e., hypothalamic) temperature, such as those that occur during exercise, exhibit negative feedback, inasmuch as they modify heat transfer rates that maintain the core temperature at its regulated level.
How do skin and core thermoreceptor inputs interact and how does the CNS integrate these inputs to produce appropriate thermoregulatory responses to both external and internal heat loads? The most plausible explanation is that signals from skin thermoreceptors change the sensitivity of the response to signals from core thermoreceptors. For example, in Figure 59-5 a decrease in core temperature produces an effector response (i.e., increased metabolic rate) that depends upon the level of input from skin cold thermoreceptors. Thus, the “gain” of the centrally induced metabolic response increases as skin temperature falls.
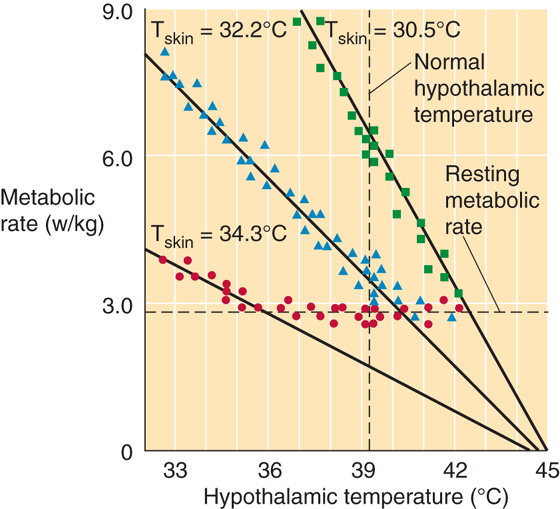
Figure 59-5 Thermoeffector responses. In these experiments on rabbits, the investigators implanted water-perfused thermodes to control the temperature of the preoptic/anterior hypothalamic area (x-axis) at three different skin temperatures (Tskin). (See Note: Effect of Skin Temperature on the Response to Hypothalamic Drive; Preoptic Area and Anterior Hypothalamus)
Thermal effectors include the cutaneous circulation, sweat glands, and skeletal muscles that are responsible for shivering
Figure 59-4 summarizes the three effectors of the thermoregulatory system. Adjusting the smooth muscle tone of cutaneous arterioles controls blood flow, and therefore heat flow, from the core to the skin surface, the primary site of heat dissipation to the environment. Over most of the skin, the autonomic nervous system controls cutaneous blood flow (see Chapter 24). When it is necessary to increase heat dissipation, active vasodilation can increase cutaneous blood flow up to 10-fold above the resting level. Conversely, when it is necessary to conserve heat in a cold environment, cutaneous vasoconstriction—mediated by sympathetic nerves—can elicit a relatively minor reduction in cutaneous blood flow, to half the resting rate. This vasoconstriction occurs at the expense of allowing skin temperature to drop closer to ambient temperature. Even with maximal vasoconstriction in effect, heat losses to a very cold environment do not fall to zero of the minimum tissue conductance.
With a moderate heat load, the autonomic response primarily increases the heat transfer rate from core to skin by increasing cutaneous blood flow. However, when the heat load is sufficiently great, the autonomic nervous system also activates the eccrine sweat glands (see Chapter 60), which secrete sweat onto the skin surface, thus elevating the partial pressure of water vapor there and promoting increased evaporation. The innervation of the secretory segment of the sweat gland is sympathetic, but it is unusual in that acetylcholine is the neurotransmitter (see Chapter 14).
When a cold stress is sufficiently great, the physiological response includes increasing heat production by involuntary, clonic, rhythmic contractions and relaxations of skeletal muscle. This shivering can double the metabolic rate for extended periods (hours) before fatigue occurs; for brief intervals, shivering can triple or quadruple the metabolic rate. Nonshivering thermogenesis in newborn infants and hibernating animals can also produce substantial amounts of heat, primarily in brown fat cells (see Chapter 57).
HYPERTHERMIA, HYPOTHERMIA, AND FEVER
Exercise raises heat production, followed by a matching rise in heat loss, but at the cost of a steady-state hyperthermia of exercise
At the onset of muscular exercise, the rate of heat production increases in proportion to the exercise intensity and exceeds the current rate of heat dissipation, thus causing heat storage and a rise in core temperature (Fig. 59-6). Hypothalamic thermoreceptors sense this increase in core temperature. The hypothalamic integrator compares this temperature signal with a reference signal, detects an error between the two, and directs neural output that activates heat dissipation (Fig. 59-4). As a result, skin blood flow and sweating increase as core temperature rises. These processes thus promote an increase in the rate of heat transfer from core to environment and slow the rate of temperature rise. At some point, the rising rate of heat dissipation equals the rate of heat production, and the rate of heat storage falls to zero. However, the now elevated steady-state core temperature persists as long as exercise continues.
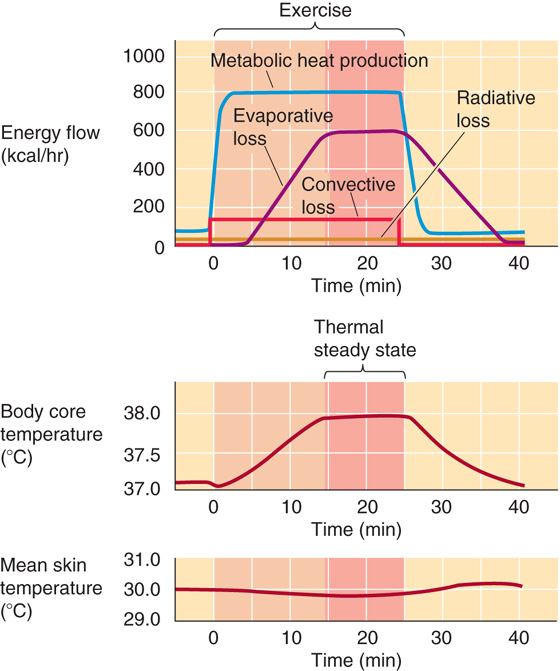
Figure 59-6 Whole-body heat balance during exercise.
The steady-state core temperature during exercise is not “regulated” at the elevated level; rather, the hyperthermia of exercise is the consequence of the initial imbalance between rates of heat production and dissipation. This imbalance is unavoidable because temperature must increase to provide the error signal that culminates in increased heat dissipation and because the response is not instantaneous. In Figure 59-6, metabolic heat production rises rapidly to its maximal level. However, evaporative heat loss increases only after a delay and then rises slowly to its maximal level, driven by increasing body temperature. The result is net storage during the first 15 minutes. The slight initial drop in core temperature at the onset of exercise is caused by flushing out of blood from the cooler peripheral circulation when the muscle and skin beds vasodilate in response to the onset of exercise. In addition, mean skin temperature decreases during exercise because of the increased evaporative cooling of the skin caused by sweating. (See Note: The Effect of Training on the Hyperthermia of Exercise)
Hyperthermia or hypothermia occurs when heat transfer from or to the environment overwhelms the body’s regulatory capacity
Although the body’s temperature regulating machinery is impressive, its capabilities are not limitless. Any factor that causes sufficiently large shifts—either positive or negative—in the rate of heat storage (Equation 59-1) could result in progressive hyperthermia or hypothermia (Equation 59-6). Because humans must operate within a fairly narrow core temperature range, such temperature changes could become life-threatening.
The most common environmental condition that results in excessive hyperthermia is prolonged exposure to heat and high ambient humidity, particularly when accompanied by physical activity (i.e., elevated heat production rate). The ability to dissipate heat by radiation falls as the radiant temperature of nearby objects increases (Equation 59-3), and the ability to dissipate heat by convection falls as ambient temperature increases (Equation 59-4). When ambient temperature reaches the mid-30s (°C), evaporation becomes the only effective avenue for heat dissipation. However, high ambient humidity reduces the skin-to-environment gradient for water vapor pressure and thus reduces evaporation (Equation 59-5). The combined reduction of heat loss by these three pathways can markedly increase the rate of heat storage (Equation 59-6) and can cause progressive hyperthermia.
It is uncommon for radiative or convective heat gain to cause hyperthermia under conditions of low ambient humidity, because the body has a high capacity for dissipating the absorbed heat by evaporation. Radiative heat gain can be excessively high during full exposure to the desert sun or during exposure to heat sources such as large furnaces. The most obvious protections against radiative hyperthermia are avoiding radiant sources (e.g., sitting in the shade) and covering the skin with loose clothing. Loose clothing screens the radiation while allowing air circulation and thereby maintaining evaporative and convective losses.
The most common environmental condition causing excessive hypothermia is prolonged immersion in cold water. Water has a specific heat per unit volume that is approximately 4000 times that of air and a thermal conductivity that is approximately 25 times that of air. Both properties contribute to a convective heat transfer coefficient (hconvective in Equation 59-4) that is approximately 100-fold greater in water than it is in air. The hconvective is ~200 kcal/(m2/hr/°C) at rest in still water but ~500 kcal/(m2/hr/°C) while swimming. The body’s physiological defenses against hypothermia include peripheral vasoconstriction (increasing insulation) and shivering (increasing heat production), but even these measures do not prevent hypothermia during prolonged exposure because of water’s high thermal conductivity. A thick layer of insulating fat retards heat loss to the water and postpones or even prevents hypothermia during prolonged exposures. Endurance swimmers used this knowledge to protect themselves when they applied a thick layer of grease to the skin surface (now, more commonly, they don a wet suit) before an event. Herman Melville noted this principle in 1851, when he referred to the low thermal conductivity of fat:
For the whale is indeed wrapt up in his blubber as in a real blanket…. It is by reason of this cozy blanketing that the whale is enabled to keep himself comfortable in all seas…. this great monster, to whom corporeal warmth is as indispensable as it is to man….
— Moby Dick
Like blubber, clothing adds insulation between skin and environment and thus reduces heat loss during exposure to the cold. The more skin one covers, the more one reduces the surface area for direct heat loss from skin to environment by convection and radiation. Adding layers of clothing increases the resistance of heat flow by trapping air, which is an excellent insulator. During heat exposure, the major avenue for heat loss is evaporation of sweat. Because evaporation also depends on the surface area available, the amount of clothing should be minimized. Wetting the clothing increases the rate of heat loss from the skin because water is a better conductor than air. Water also can evaporate from the clothing surface, thereby removing heat from the outer layers and increasing the temperature gradient (and rate of heat loss) from skin to clothing.
Heat Stroke
As body core temperature rises, excessive cutaneous vasodilation can lead to a fall in arterial pressure (see Chapter 25) and therefore to a decrease in brain perfusion. As core temperature approaches 41°C, confusion and, ultimately, loss of consciousness occur. Excessive hyperthermia (>41°C) leads to the clinical condition known as heat stroke. High temperature can cause fibrinolysis and consumption of clotting factors and thus disseminated intravascular coagulation (DIC), which results in uncontrolled vascular thrombosis and hemorrhage. Heat-induced damage to the cell membranes of skeletal and myocardial muscle leads to rhabdomyolysis (in which disrupted muscle cells release their intracellular contents, including myoglobin, into the circulation) and myocardial necrosis. Cell damage may also cause acute hepatic insufficiency and pancreatitis. Renal function, already compromised by low renal blood flow, may be further disrupted by the high plasma levels of myoglobin. Ultimately, the CNS is affected by the combination of high brain temperature, DIC, and metabolic disturbances.
Fever, unlike other types of hyperthermia, reflects an increase in the set point for temperature regulation
Fever is a regulated elevation of core temperature resulting from effects associated with infection or disease. Fever is caused by the action of circulating cytokines called pyrogens, which are low-molecular-weight polypeptides produced by cells of the immune system. As for the hyperthermia of exercise, fever begins when heat production temporarily exceeds heat dissipation. However, fever differs from other hyperthermias in that the hypothalamus actively regulates core temperature to an elevated set point.
Figure 59-7 illustrates the basic differences between the events leading to exercise hyperthermia and those leading to fever. During the genesis of exercise hyperthermia (Fig. 59-7A), the rate of heat production increases to more than the rate of heat dissipation for a period and causes net heat storage. Moreover, the temperature set point (Tset) is unchanged, and thus the error signal gradually increases to a new, sustained level. During the genesis of a fever (Fig. 59-7B), Tset suddenly increases to a value higher than the normal temperature, so the integrator interprets the normal temperature as being lower than the new Tset. The fever is an appropriate response to this condition and develops as the heat loss rate from the body falls or the heat production rate rises until such time as core temperature increases to the new “regulated” level. Thus, the error signal is initially large but becomes smaller as the fever develops. In the new steady state, core temperature remains elevated until the signals responsible for the fever (i.e., pyrogens) subside and Tset returns to normal.
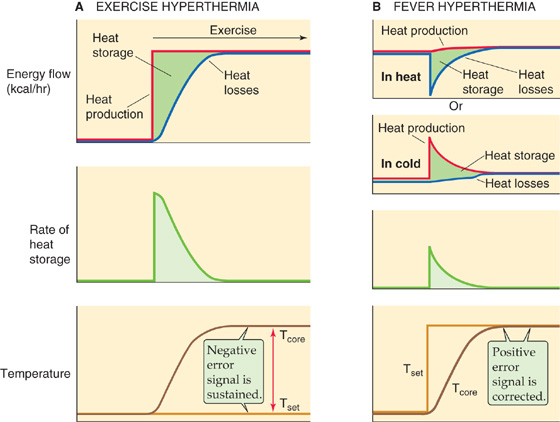
Figure 59-7 Exercise hyperthermia versus fever. A, The top panel shows how, during exercise, heat production temporarily exceeds heat loss, thus resulting in net heat storage. The middle panel shows that the rate of heat storage is highest initially and falls to zero in the new steady state. Finally, the bottom panel shows that as body core temperature rises away from the set-point, the error signal gradually increases. In the new steady state, the error signal is maximal and sustained. B, The top two panels show how, during fever, net heat storage can occur because of either reduced heat loss or increased heat production. The third panel from the top shows that, as in exercise, the rate of heat storage is highest initially. The bottom panel shows that as body core temperature rises, it approaches the new elevated set-point. Thus, the error signal is initially maximal and gradually decreases to zero in the new steady state.
The subjective assessments of thermal comfort support this description. During exercise, one perceives the rise in core temperature as body heating and may choose to remove clothing to cool the body. During the onset of a fever, however, the individual feels cold and may choose to put on additional clothing and warm the body. If fever strikes when the patient is in a warm environment in which the cutaneous vessels are dilated (Fig. 59-7B, top panel), the response to the Tset increase will be to vasoconstrict, which decreases heat loss. In contrast, if the patient is in a cold environment in which the cutaneous vessels are already constricted (Fig. 59-7B, second panel), the response will be to shiver.
Figure 59-8 summarizes the responses to fever-producing stimuli. Macrophages and, to a lesser extent, lymphocytes release cytokines into the circulation in response to a variety of infectious and inflammatory stimuli. Cytokines, the messenger molecules of the immune system, are a diverse group of proteins involved in numerous tasks in the host defense response. The first is the immune response to foreign substances including stimulation of T-lymphocyte proliferation, natural killer cells, and antibody production. The second is the acute-phase response to foreign substances, a diffuse collection of nonspecific host reactions to infection or trauma. Finally, cytokines may act as endogenous pyrogens (Table 59-3). However, no one cytokine, administered experimentally, can fully mimic the temperature increase that occurs during fever. Fever production may occur through a cascade that is initiated when interleukin (IL-1β), for example, interacts with the endothelial cells in a leaky portion of the blood-brain barrier (see Chapter 11) located in the capillary bed of the organum vasculosum laminae terminalis (OVLT). The OVLT is highly vascular tissue that lies in the wall of the third ventricle (above the optic chiasm) in the brain. IL-1β triggers endothelial cells within the OVLT to release prostaglandin E2 (see Chapter 3), which then diffuses into the adjacent hypothalamus and—in a manner not yet understood—elevates Tset and initiates the febrile response.
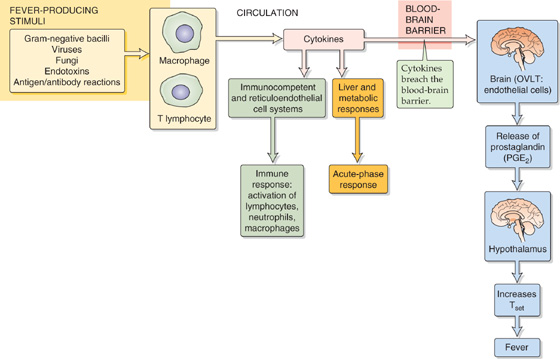
Figure 59-8 Host defense response.
Table 59-3 Endogenous Pyrogens
Pyrogen |
Symbol |
Interleukin 1α |
IL-1α |
Interleukin 1β |
IL-1β |
Interleukin 6 |
IL-6 |
Interleukin 8 |
IL-8 |
Tumor necrosis factor α |
TNF-α |
Tumor necrosis factor β |
TNF-β |
Macrophage inflammatory protein 1α |
MIP-1α |
Macrophage inflammatory protein 1β |
MIP-1β |
Interferon α |
INF-α |
Interferon β |
INF-β |
Interferon γ |
INF-γ |
The value of fever in fighting infection is still debated. A popular hypothesis is that the elevated temperature enhances the host’s response to infection. This view is supported by the in vitro observation that the rate of T-lymphocyte proliferation in response to interleukins is many-fold higher at 39°C than it is at 37°C.
REFERENCES
Books and Reviews
Blatteis CM, Sehic E: Fever: How may circulating pyrogens signal the brain? News Physiol Sci 1997; 12:1-9.
Block BA: Thermogenesis in muscle. Annu Rev Physiol 1994; 56:535-577.
Horowitz M: Do cellular heat acclimation responses modulate central thermoregulatory activity? News Physiol Sci 1998; 13:218-225.
Lee-Chiong TL, Stitt JT Jr: Disorders of temperature regulation. Compr Ther 1995; 21:697-704.
Simon HB: Current concepts: Hyperthermia. N Engl J Med 1993; 329:483-487.