CHAPTER 10 Brain stem, cranial nerves, visual system, vestibular system, reticular system
Brain stem
Although the brain stem incorporates many important structures, only the structures perceived to be most relevant to the therapist in terms of the neurologically impaired patient are discussed in this section.
Basic anatomy
Anatomically, the brain stem is divided into three regions: the mid-brain, the pons and the medulla oblongata. Although the three regions have features which are common to all, they have others which allow us to distinguish between them.
Mid-brain
The mid-brain is approximately 1.5 cm long and extends from the pons to the mamillary bodies. The ventral surface connects to the two superior cerebellar peduncles (S2.12). The roof of the mid-brain is composed of four colliculi, the superior colliculi associated with the visual system and the inferior colliculi related to the auditory system. The red nucleus is also contained within the mid-brain. The periaqueductal grey, an area of grey matter within the mid-brain, is important in the descending modulation of pain (S3.29).
Pons
The pons is about 2.5 cm long and is composed of transverse fibres which form ridges across the surface of the area. The middle cerebellar peduncles attach to the pons.
Medulla oblongata
The medulla is about 3 cm long and forms the base of the brain stem, adjoining the spinal cord at the level of foramen magnum. The descending motor tracts passing through it form two pyramids on the ventral surface of the medulla between which is the anterior median fissure. This fissure is disrupted where the tracts cross over the midline, termed the ‘decussation of the pyramids’. The inferior cerebellar peduncle attaches to the medulla and behind it is the ‘olive’. The olive is a prominent oval swelling that marks the position of the inferior olivary nucleus (S2.12).
Function of the brain stem
The three regions of the brain stem contain the nuclei of the cranial nerves which control a variety of vital functions (S2.10). The ascending (S2.15) and descending tracts (S2.14) also pass through the brain stem between the spinal cord and cerebral cortex. The reticular formation (S2.10) and the cerebellum (S2.12) are integrally linked with all three regions and have specialist roles related to movement.
Cranial nerves
Basic anatomy
The names, numbers and functions of the cranial nerves can be seen in Table 10.1.
Table 10.1 Names and functions of the cranial nerves
| Cranial nerve | Motor/sensory/mixed | Function of the cranial nerves |
|---|---|---|
| Olfactory (I) | S | Smell |
| Optic (II) | S | Vision – acuity and visual fields |
| Oculomotor (III) | Mo | |
| Trochlear (IV) | Mo | Assists in turning eyeball down and laterally |
| Trigeminal (V) | Mix | |
| Ophthalmic division | S | Cornea, skin of forehead, scalp, eyelids and nose |
| Maxillary division | S | Skin over maxilla, teeth of upper jaw and palate |
| Mandibular division | Mo | Muscles of mastication and swallow. Tensor tympani (functions to reduce the amplitude of sounds) |
| S | Skin of cheek, mandible and side of head. Teeth of lower jaw and temporomandibular joint. Anterior tongue | |
| Abducens (VI) | Mo | Turns the eyeball laterally |
| Facial (VII) | Mix | |
| Mo | Muscles of face and swallow | |
| S | Taste anterior 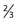 of tongue of tongue |
|
| Secretomotor | Parasympathetic control of salivary glands and lacrimal gland (tear duct) | |
| Vestibulocochlear (VIII) | S | |
| Vestibular | S | From vestibular apparatus related to position and movement of head |
| Cochlear | S | Organ of hearing |
| Glossopharyngeal (IX) | Mix | |
| Mo | Swallowing | |
| S | Taste posterior 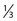 of tongue of tongue |
|
| Secretomotor | Parasympathetic control of salivary gland | |
| Vagus (X) | Mix | |
| Mo | Parasympathetic innervation of heart and large blood vessels | |
| S | General sensation to lungs, larynx, trachea and bronchi, gastrointestinal tract, colon, liver and kidneys | |
| Accessory (XI) | Mo | |
| Cranial root | Mo | Muscles of soft palate, some of the muscles of pharynx and larynx |
| Spinal root | Mo | Innervates sternocleidomastoid and trapezius |
| Hypoglossal (XII) | Mo | Muscles of tongue related to shape and movement |
Motor nuclei
Five of the cranial nerves are entirely motor. Afferent input to the motor nuclei is from the cerebral cortex via the cortico-bulbar tract (S2.14). The output from the cranial nerve nuclei to the effector muscle is via a lower motor neuron.
Sensory nuclei
Three of the cranial nerves are entirely sensory. The afferent input is from the relevant sensory organ, e.g. the rods and cones of the retina for the optic nerve. They synapse in the brain stem in the relevant cranial nerve nuclei and finally decussate and ascend to other regions including the thalamus and ultimately the cerebral cortex.
Assessment of the cranial nerves
Traditionally, this has been carried out by the medical team on initial contact and is not covered in this text. However, it is important for the therapist to understand the findings and to identify any changes that may occur. Symptoms associated with cranial deficits may be identified by the patient themselves during the subjective assessment (S3.16) or noticed by the therapist by general observation (S3.17). In either case, any suspicions should be followed up with a referral to an experienced clinician or the medical team.
Visual system
Basic anatomy
The visual pathway
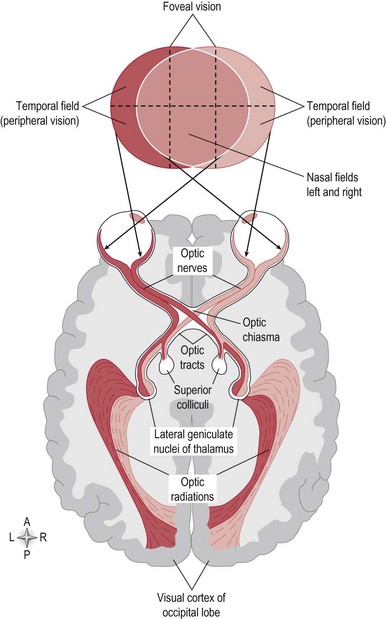
The sensory receptors of the visual system are the photosensitive rod and cone proteins of the retina. The rods and cones are most dense in the central region of the retina called the fovea. An image of the environment is focused onto the retina by alterations in the thickness of the lens. The retina converts a light pattern into a neuronal signal, which is conducted along the visual pathway to various parts of the brain to be processed (Fig. 10.1).
Function of the visual system
Binocular vision
This is defined as vision in which two eyes are used. Although there is a huge area of overlap between the visual fields of the left and right eye the eyes still view the environment slightly differently. The brain uses this slight disparity to estimate distance, allowing us to perceive 3D objects and spatial depth. If the two views are not successfully merged, the individual may develop double vision (diplopia).
Central or foveal vision
The fovea is responsible for sharp central vision necessary for any activity where visual detail is of primary importance. For example, reading and watching TV. Hence, if clarity is required, visual gaze must be centred on this region.
Peripheral vision
This aspect of vision occurs outside the centre of gaze and is weaker because the density of rods and cones outside the fovea region is much less. However, our peripheral vision is essential for fast reactions to visual stimuli in the periphery and monitoring body position relative to gravity. Our peripheral vision also allows the individual to recognize well-known structures and forms without the need to focus centrally.
Visual acuity
This is the clearness of vision and is achieved by the alteration of lens thickness and pupil size by small local muscles within the eyeball itself which focus the image at the fovea. However, acuity also relies upon coordination between head and body movements to ensure that the visual stimuli remains at fovea. This involves the ability to fix and scan.
Fixing (S3.27)
The ability to fix gaze on an object while the body is moving, e.g. when travelling on a train or in a car. The mechanism can be explained as follows. When your head turns to the right, there is a slow reflex movement of your eyes to the left, in order that they appear to remain stationary. This is achieved via the vestibulocular reflex and as the name suggests, is under the direction of the vestibular system (S2.10). The effector muscles are innervated by the cranial nerve nuclei (III and VI).
Scanning (S3.27)
The ability to scan the eyes and head in a coordinated manner to maintain the object of interest or visual stimuli in the field of vision, e.g. while reading. The relevant head/neck movements are achieved via the tectospinal and tectobulbar tracts (S2.15) and eye movements by the cranial nerve nucleui (III and VI).
Visual fields (S3.27)
Information from the temporal and nasal visual fields of both eyes reaches the occipital cortex simultaneously, via the visual pathway (Fig. 10.1).
Visual perception
Vision provides the central nervous system with information related to our body position and the environment, however how we perceive and make sense of this raw information is highly subjective. The interpretation of our surroundings is integrally linked with memory and experience and therefore is individual in nature.
Vision and movement
The visual system plays an important part in movement and balance as it provides us with:
However, we can of course balance without vision and vision can be unreliable. For example, our visual system is not always accurate when differentiating between self-motion and the movement of an external body. This may be observed when stationary in a car/bus/train and a vehicle pulls off alongside. Often, this can be perceived as self-motion and you may react accordingly, when actually it is the external object that is in motion.
Vestibular system
Basic anatomy
The vestibular nuclei of the 8th cranial nerve are found within the pons and are the integrating centre for the vestibular system.
Input to the vestibular system
The sensory receptor of the vestibular system is the vestibular apparatus housed in the labyrinth of the inner ear – the labyrinth being made up of an outer bony labyrinth and an inner membraneous labyrinth full of endolymph. The labyrinth is divided into three sections, all of which function by translating movement of sensory hairs into electrical impulses:
Vestibule
The vestibule consists of two organs, the utricle and saccule. In the wall of the utricle and saccule is a thickened region of specialized epithelium, called a ‘macula’ (Fig. 10.2).
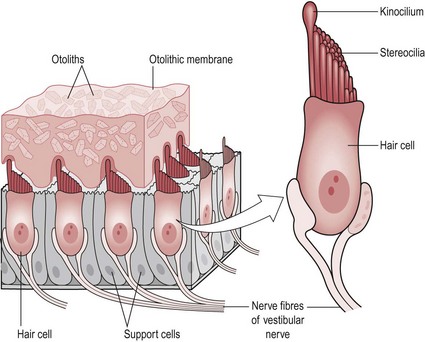
The macula consists of two types of cells:
The macula is divided into two halves with all of the hair bundles within one-half of the macula orientated in the same way. This ensures that a body movement, which leads to a graded potential in one hair bundle cell is likely to do the same in all the other cells and hence, guarantee an action potential in the sensory neuron. The hair bundles in the other half of the macula are all orientated in the opposite direction producing the opposite response and allowing the whole organ to be responsive to all relevant directions of movement. The macula orientation in the other side of the brain is the same, the relevance of this can be seen later in this section.
Semicircular canals
There are three semicircular canals which lie at 90° to each other. At the base of each canal is a thickened region termed the ‘ampulla’. Inside the ampulla is an elevation of specialized cells called the ‘crista’ (Fig. 10.3).
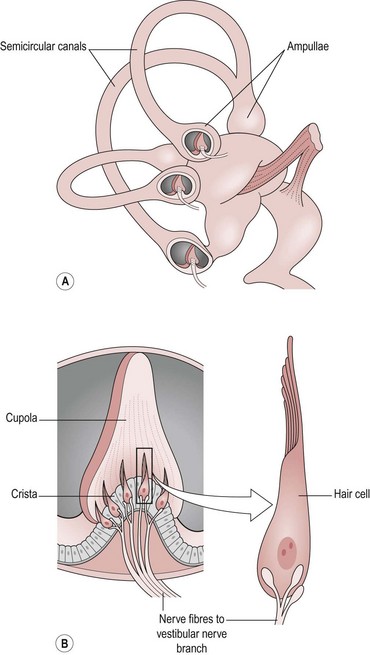
The crista consists of two types of cells:
Output from the vestibular system
The vestibular system primarily works subcortically and has few synapses, therefore it responds very quickly, which is vital in terms of balance. From the vestibular apparatus, information passes along the vestibular nerve (VIII) and is processed at the vestibular nucleus in the pons. From here efferent connections go to the:
Function of the vestibular system
Vestibule
The static labyrinth (utricle and saccule) detects linear movement by way of its sensory organ, the macula. The utricle detects linear movement in a frontal plane and the saccule in a sagittal plane.
The linear movement may be a result of gravity, e.g. head tilting or bending over to pick up an object from the floor or linear translation such as the acceleration and deceleration experienced when walking or travelling by car. In either case, the linear movement causes changes in the macula structure and consequently, a muscle action in terms of a balance response.
For example, during a forward head tilt (cervical spine flexion), the macula of the saccule will respond. The forward tilt causes the heavy mass of the otoliths and the otolithic membrane to ‘fall’ forwards moving the stereocilia in relation to the kinocilium and producing a graded potential in the sensory neuron. As the orientation of hair cells is opposite within each half of the saccule, the result will be depolarization (excitation) in one-half and hyperpolarization (inhibition) in the other (S2.6).
The action potential produced travels along the vestibular nerve (VIII) and recruits or inhibits the appropriate muscles via the vestibulospinal tract (S2.15). In this case, the forward head tilt has moved the centre of gravity forwards making the body less stable. The balance reaction required will attempt to bring the centre of gravity backwards, by recruiting extensor muscles around the pelvis/hip or trunk and simultaneously reduce the activity of the flexor muscles. This is important so that the centre of gravity is not moved further forwards and balance becomes seriously compromised. Remember also that the orientation of hair bundles in one-half of the saccule is the same in both the left and right sides of the brain and therefore the response will be bilateral. This makes sense, as the response to a linear movement needs to be symmetrical.
Semicircular canals
The kinetic labyrinth (semicircular canals) detects the movement of rotation in all planes, e.g. head rotation to look over your shoulder, or a somersault on a trampoline.
As with the static labyrinth, the movement occurring results in a muscle action response as a result of nerve cells being depolarized or hyperpolarized. However, the mechanism by which the sensory organ achieves this is slightly different.
As highlighted in the anatomy section, the cupola is the full width of the ampulla and this stops the free flow of endolymph within each canal. When a rotation movement is experienced, movement of the endolymph is blocked by the cupola producing a force upon the gel structure. The response of the hair bundles embedded in the cupola is as for the macula in terms of production of an action potential in the sensory nerve.
The hair bundles within one canal are all orientated the same way so that the response is sufficient to guarantee an action potential. In the semicircular canals, the orientation of one canal is different on the contralateral side of the brain and this is important because rotation is asymmetrical in nature and therefore requires a different motor response from each side of the body for balance to be maintained.
Reticular system
Basic anatomy
The reticular formation is a complicated network of circuits which extend from the spinal cord through the medulla, pons, mid-brain, subthalamus, hypothalamus and thalamus. Although previously considered anatomically ill-defined, more recent scientific investigation has shown the reticular formation to be well organized and functionally discrete. Anatomically it can be conveniently divided into three longitudinal columns:
Function of the reticular formation
Cardiovascular and respiratory control
The cardiorespiratory centres are now considered part of the reticular formation. In terms of respiratory function, many of the neurons from the reticular formation synapse on motor neurons that innervate the muscles of inspiration and expiration. The control of cardiovascular function is via regulation of the autonomic system (a) parasympathetic output from the vagus nerve and (b) sympathetic output from the sympathetic trunk levels T1–5 of the spinal cord.
Modulation of muscle tone
The appropriate level of resting muscle tone is regulated by modulation of the stretch reflex (S2.13) by the descending tracts. The reticulospinal tracts in combination with the vestibulospinal and reticulobulbar tracts influence muscle tone at the level of the alpha motor neuron. The two reticulospinal tracts specifically influence extensor muscle tone. The pontine/lateral reticulospinal tract terminates at the ventral horn directly on motor neurons and enhances extensor muscle tone, whereas the medullary/medial reticulospinal tract (S2.14) terminates indirectly on motor neurons and exerts an inhibitory influence on extensor muscle tone. The antagonistic effect of these tracts allows modulation and grading to set the level of extensor muscle tone appropriately for the functional need. However, the reticulospinal tracts also work in cooperation with the vestibulospinal tracts (S2.14) to specifically maintain tone of the antigravity muscles.
The reticular formation also has a role in the motor control of facial expression related to our emotional behaviour (S2.9) via its links to the limbic system and coordination of the timing and direction of eye movements via the superior colliculus and cranial nerves.
Regulation of the sleep/wake cycle
Our normal sleep/wake cycle shows a definite recognizable daily pattern, with the hypothalamus being ultimately involved in its control. However connections to the hypothalamus allow the reticular formation to influence these biological rhythms. The role of the reticular formation in this function is not wholly understood, however the reticular activating system (RAS), which lies near the junction of the pons and mid-brain, appears to be the primary cause of wakefulness and rapid eye movement (REM) sleep. REM sleep is characterized by an increase in brain activity and an inactive body. On the other hand, inactivity of the RAS region, via inhibition from the hypothalamus is important for producing non-REM sleep, a stage of deep sleep characterized by an active body and reduced brain activity.
References and Further Reading
Hunziker H-W. In the eye of the reader: foveal and peripheral perception – from letter recognition to the joy of reading. Zürich: Transmedia Stäubli Verlag; 2006.
Palmer SM, Rosa MG. A distinct anatomical network of cortical areas for analysis of motion in far peripheral vision. European Journal of Neuroscience. 2006;248:2389-2405.
Purves D, Augustine GJ, Fitzpatrick D, et al. Neuroscience, ed 4. Sunderland: Sinauer Associates; 2008.
Snell RS. Clinical neuroanatomy, ed 6. Philadelphia: Lippincott Williams and Wilkins; 2006.