CHAPTER 13 The spinal cord and spinal reflexes
Basic anatomy and function of the spinal cord
Gross anatomy
The spinal cord begins superiorly at the foramen magnum where it is continuous with the medulla oblongata (S2.10). It continues inferiorly within the vertebral column protected in the vertebral canal by the meninges and cerebrospinal fluid (S2.8). The lowest part of the spinal cord lies at the first lumbar vertebra (level L1/2). Along the length of the spinal cord, there are two swollen regions: the cervical and lumbar enlargements, which represent the origins of the brachial and lumbosacral plexus, respectively.
Grey and white matter
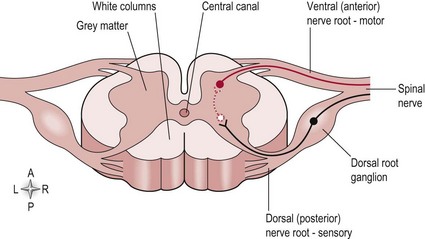
In cross-section (Fig. 13.1), the spinal cord can be seen to consist of the inner grey matter (H shaped) and the remaining area termed the white matter. These are so named because of their microscopic appearance and a consequence of the cells located within the area. The white matter consists of myelinated axons and represents the existence of the ascending and descending tracts (S2.14, 15). The grey matter is formed by the cell bodies of the same neurons and includes motor neurons and many interneurons which make connections within segments and between segments of the spinal cord. The grey matter is described in terms of the anterior/ventral horn and posterior/dorsal horn (Fig. 13.1). In the thoracic and upper lumbar region there is also a lateral horn, which represents the sympathetic trunk, part of the autonomic nervous system.
Central pattern generators (CPG)
A central pattern generator is a network of neurons within the spinal cord which underlie the production of most of our rhythmic motor output, such as walking, chewing/swallowing and breathing. These are by nature stereotyped, repetitive and complex movements but unlike reflex activity, are still under voluntary control. The network of neurons is thought to work in two opposing halves which when activated switch each other on and off to create a repetitive rhythmic action. The stereotyped movement pattern created avoids the need for descending commands from higher centres, however changes in the environment may require the CPG to be modulated and adapted to meet the changing needs. This modulation primarily comes from the higher centres but the CPGs also modulate themselves by responding to sensory feedback from the body. A small change in function may result in a long-term change to the whole CPG network as it adapts to achieve a motor goal. In terms of neurologically impaired patients, this may be positive or negative in relation to function.
Spinal nerves
At each spinal level there are a pair (left and right) of spinal nerves which exit the vertebral canal via the intervertebral foramen. There are 31 pairs of spinal nerves in total (8 cervical, 12 thoracic, 5 lumbar and 5 sacral). The spinal nerve consists of a sensory root and motor root (Fig. 13.1) from the spinal cord. The sensory root transmits information into the spinal cord and its cell body is found in the dorsal root ganglion just adjacent to the spinal cord but within the vertebral canal. The motor root carries information out of the spinal cord and its cell body is located in the anterior/ventral horn of the grey matter of the spinal cord. The sensory and motor roots at one level combine to form a spinal nerve and are part of the peripheral nervous system. Spinal nerves from several levels of the spinal cord will combine to form a peripheral nerve. For example, the median nerve consists of the spinal nerves from levels C5/6/7/8 and T1, whereas the musculocutaneous nerve includes spinal nerves from levels C5/6/7.
Cauda equinus
From lumbar level L1/2, where the spinal cord terminates the anatomy, is different. The cord itself is referred to as the ‘cauda equinus’ and represents a bundle of spinal roots from the segments above within the vertebral canal. This occurs because the vertebral column grows longer than the spinal cord during development. Therefore in the adult, the spinal cord segments do not correspond to the vertebral segment of the same level. For example, the lower lumbar and sacral spinal nerves exit at the appropriate levels (i.e. L3/4/5 and S1–5), however their spinal cord segments are found between T9 and L2 vertebral segments. This is important when considering pathologies which affect the spinal cord.
Lower motor neuron/final common pathway and upper motor neuron
Understanding the anatomy of the spinal cord and its divisions is vital for ensuring your assessment of a neurologically impaired patient is appropriate and accurate. One important division is the differentiation between an upper motor neuron and lower motor neuron pathology and their clinical presentations.
The lower motor neuron/final common pathway
The neurons indicated as nerve roots in Figure 13.1 are the starting point of the lower motor neuron division. Anything more peripheral is included in this division. In effect, damage to these neurons is interpreted as a deficit of the peripheral nervous system, for example, Guillain–Barré syndrome and motor neuron disease. However, different clinical presentations can occur within this division depending upon the location of the lesion. For example, damage to the spinal root or spinal nerve would present very differently to that of a peripheral nerve.
Spinal root or spinal nerve
A single spinal root or nerve could be damaged with a very specific injury such as an intervertebral disc prolapse or in association with other trauma. The injury itself could relate to:
Therefore the clinical presentation resulting from damage to a single spinal root or spinal nerve will be very localized in terms of reduced sensation and/or a weakness in a specific muscle. Total paralysis of a muscle is unlikely as a single spinal root level innervates only a fraction of the muscle. For example, the triceps brachii muscle is supplied by the radial nerve (C5/6/7/8 and T1), so damage to root level C5 would present as mild weakness and not total paralysis because the muscle is still innervated by the remaining undamaged spinal root levels.
Peripheral nerve
As already stated, the peripheral nerve consists of both sensory and motor spinal nerves from various spinal levels and in different combinations. Therefore a complete severance of a peripheral nerve will present with more widespread sensory and motor loss and even the complete paralysis of a muscle. The clinical presentation will be more diffuse, dictated by the individual peripheral nerve involved and whether the severance is complete or partial. For example, a complete lesion of the musculocutaneous nerve (C5/6/7) in the upper arm would lead to complete paralysis of coracobrachialis, biceps brachii and brachialis muscles and loss of sensation in the area of skin supplied by its cutaneous branch.
The upper motor neuron
The upper motor neuron division includes any neuron that is confined entirely to the central nervous system. It includes any neuron between the brain and the peripheral nervous system, incorporating neurons within the spinal cord, the ascending and descending tracts and higher centres, such as the brain stem and cerebral cortex. Damage to the upper motor neuron pathway occurs in pathologies such as spinal cord injury, cerebrovascular accident, multiple sclerosis and Parkinson’s disease. The central nervous system is highly complex and therefore, damage within this division could result in a wide range of diverse symptoms.
Basic anatomy and function of spinal reflexes
A reflex is an involuntary, almost instantaneous movement, in response to a stimulus that does not require the higher centres of the brain. However, information received from both the periphery and higher centres is used to modulate the reflex activity at the level of the spinal cord.
The neural pathway involved in a simple reflex arc includes:
The most important reflex arcs in relation to movement control are:
Stretch reflex
The stretch reflex has both a phasic and a tonic component. The phasic component adapts rapidly to a stimulus and responds to a change in muscle length. The tonic component which is continually active is more responsive to a constant stretch.
Phasic stretch reflex
This component of the stretch reflex is primarily monosynaptic:
Function
The phasic component is evident in the intact nervous system at rest and is the component that is assessed during reflex testing (S3.22). An imposed stretch on the muscle by the clinician gives information on nerve integrity and the reflexic properties of muscle. This technique is referred to as a tendon jerk.
Tonic stretch reflex (TSR)
This component is primarily polysynaptic:
Function
The tonic component is not apparent in an intact nervous system unless the muscle is already contracting. The TSR serves to set resting muscle tone (S3.21), the base level of muscle activity. The reflex arc also responds to changes in the environment adjusting the level of activity as appropriate to functional needs, e.g. lowering muscle activity when we lie down and raising it in preparation to stand up against gravity.
Golgi tendon organ (GTO)
This reflex arc is polysynaptic:
Function
This reflex is protective in terms of reducing a muscle contraction in circumstances where excessive tension is being produced and injury is likely. The GTO works in opposition to the TSR arc. Therefore, the balance of the two outputs at the alpha motor neuron ensures the level of muscle activity/tone is appropriate at rest and during movement without the risk of excessive tension and potential damage occurring. The GTO and TSR also provide the central nervous system with continuous information about the mechanical state of the muscle.
Flexor withdrawal and crossed extensor reflex
This reflex is polysynaptic and initiated from the flexor withdrawal arc:
Function
The primary function of the flexor withdrawal reflex is protection in response to a painful stimulus. The crossed extensor reflex is supportive to this role as it simultaneously activates the extensor muscles of the contralateral lower limb for weight bearing, so that the ipsilateral lower limb can be flexed without the body falling over. There is some evidence to support the theory that this reflex also underlies our walking pattern, with alternating phases of stance (crossed extensor) and swing (flexor withdrawal).
Additional neuronal activity associated with the reflex arc
For the reflex arcs described above, additional circuitry exists to ensure the efficiency of the response and allow modification as necessary.
First, the afferent neuron within the circuit includes the following branches which synapse upon:
Second, the efferent neuron output from the reflex arc can be modulated by neurons from higher centres. This occurs via the descending tracts (S2.14), which synapse upon the alpha motor neuron directly or indirectly via interneurons. This descending control involves altering the level of output from the AMN to the muscle by enhancing either the inhibitory or excitatory synapses as required.
References and Further Reading
Hooper SL. Central pattern generators. www.elsnet/elsonline/figpage/10000202html, 1999.
Kandel ER, Schwartz JH, Jessell TM. Principles of neural science, ed 4. New York: McGraw-Hill Health Professions Division; 2000.
Kiehn O, Butt SJ. Physiological anatomical and genetic identification of CPG neurons in the developing mammalian spinal cord. Progress in Neurobiology. 2003;70:347-361.