STRUCTURE AND FUNCTION OF THE RENAL AND UROLOGIC SYSTEMS


The primary function of the kidney is to maintain a stable internal environment for optimal cell and tissue metabolism. The kidneys accomplish these life-sustaining tasks by balancing solute and water transport, excreting metabolic waste products, conserving nutrients, and regulating acids and bases. The kidney also has an endocrine function, secreting the hormones renin, erythropoietin, and 1,25-dihydroxyvitamin D3 for regulation of blood pressure, erythrocyte production, and calcium metabolism, respectively. In times of severe fasting the kidney also can synthesize glucose from amino acids, performing the process of gluconeogenesis. The formation of urine is achieved through the processes of filtration, reabsorption, and secretion by the glomeruli and tubules within the kidney. The bladder stores the urine that it receives from the kidney by way of the ureters. Urine is then removed from the body through the urethra.
STRUCTURES OF THE RENAL SYSTEM
The kidneys are paired organs located on the posterior abdominal wall outside the peritoneal cavity. They lie on either side of the vertebral column with their upper and lower poles extending from the twelfth thoracic to the third lumbar vertebrae (Figure 35-1). Each kidney is approximately 11 cm long, 5 to 6 cm wide, and 3 to 4 cm thick. A tightly adhering capsule (the renal capsule) surrounds each kidney, and the kidney then is embedded in a mass of fat. The capsule and fatty layer are covered with a double layer of renal fascia, fibrous tissue that attaches the kidney to the posterior abdominal wall.
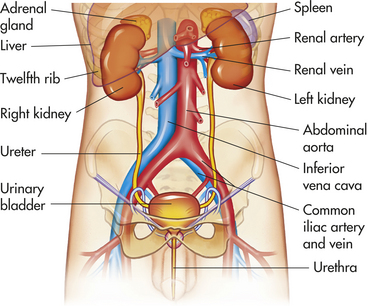
Figure 35-1 Organs of the urinary system. (From Patton KT, Thibodeau GA: Anatomy & physiology, ed 7, St Louis, 2010, Mosby.)
The cushion of fat and the position of the kidney between the abdominal organs and muscles of the back protect it from trauma. The right kidney is slightly lower than the left; it is displaced downward by the overlying liver. A medial indentation (the hilum) contains the entry and exit for the renal blood vessels, nerves, lymphatic vessels, and ureter.
The gross structure of the kidney can be identified when it is divided from top to bottom in a coronal plane (Figure 35-2). The major components are the outer renal cortex and the inner renal medulla. The cortex contains all the glomeruli and portions of the tubules. The medulla is formed by the straight segments of the proximal and distal tubules and the collecting ducts. It consists of a series of wedges, called renal pyramids, with an outer zone close to the cortex and an inner zone. Renal columns extend from the cortex down between the renal pyramids. The apexes of the pyramids project into a minor calyx (a cup-shaped cavity) that join together to form a major calyx. The calyces receive urine from the large collecting ducts. The major calyces join to form the renal pelvis, an extension of the upper end of the ureter. The walls of the calyces and ureters contain smooth muscles that contract to move urine to the bladder.
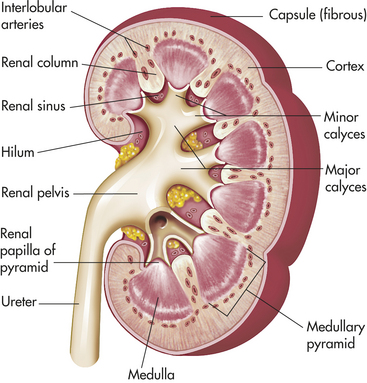
Figure 35-2 Kidney structure. (From Patton KT, Thibodeau GA: Anatomy & physiology, ed 7, St Louis, 2010, Mosby.)
The structural unit of the kidney is the lobe. Each lobe is composed of a pyramid and the overlying cortex. There are about 14 lobes in each kidney.
Nephron
The nephron is the functional unit of the kidney. Approximately 1.2 million nephrons are contained in each kidney. The nephron is a tubular structure with subunits that include the renal corpuscle, proximal convoluted tubule, loop of Henle, distal convoluted tubule, and collecting duct, all of which contribute to the formation of final urine (Figure 35-3). The different structures of the epithelial cells lining various segments of the tubule facilitate the special functions of secretion and reabsorption (Figure 35-4).
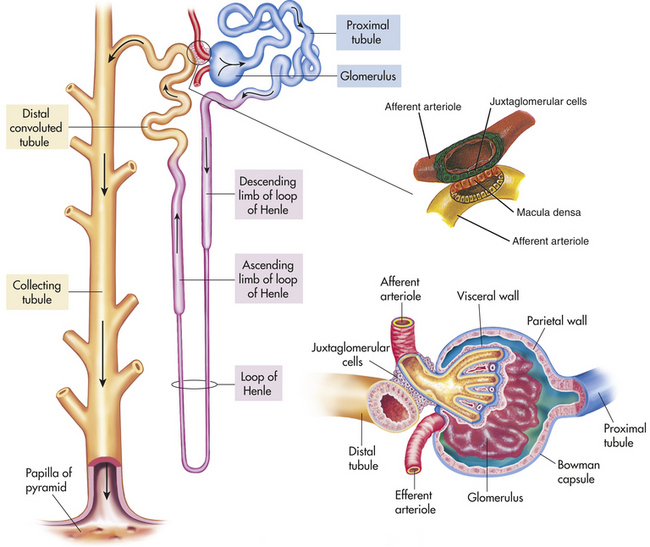
Figure 35-3 Components of the nephron. (From Patton KT, Thibodeau GA: Anatomy & physiology, ed 7, St Louis, 2010, Mosby; Damjanov: Pathology for health professions, St Louis, 2006, Mosby.)
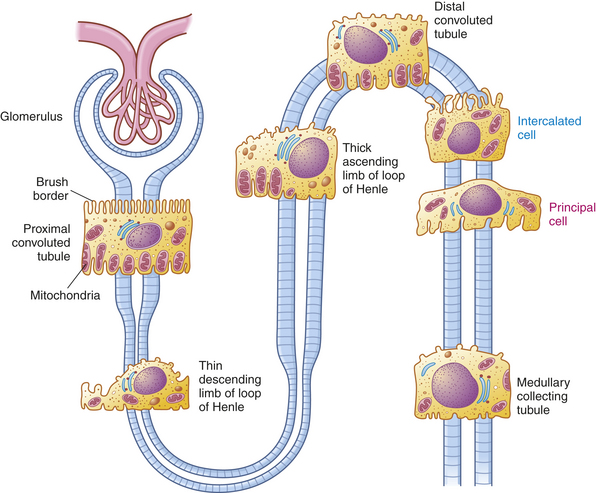
Figure 35-4 Epithelial cells of the various segments of nephron tubules. The brush border and high number of mitochondria in the cells of the proximal convoluted tubule permit reabsorption of 60% of the glomerular filtrate. Intercalated cells (blue) secrete either H+ (reabsorb 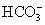 ) or and reabsorb K+Principal cells (magenta) reabsorb Na+ and water and secrete K+.
) or and reabsorb K+Principal cells (magenta) reabsorb Na+ and water and secrete K+.
The kidney has three kinds of nephrons: (1) superficial cortical nephrons (85% of all nephrons), which extend only partially into the medulla; (2) midcortical nephrons with short or long loops; and (3) juxtamedullary nephrons, which lie close to and extend deep into the medulla and are important for the process of concentrating urine (Figure 35-5). The glomerulus (Figure 35-6; see also Figure 35-3) is a tuft of capillaries, the glomerular capillaries, that loop into a circular capsule, the Bowman capsule, like fingers pushed into bread dough. Mesangial cells (shaped like smooth muscle cells) and the mesangial matrix lie between and support the glomerular capillaries.1 They have contractile and phagocytic properties, similar to monocytes, release inflammatory cytokines, and produce vasoactive substances that influence the glomerular filtration rate (GFR) by regulating glomerular capillary blood flow. The space inside the Bowman capsule is called the Bowman space. Together, the glomerulus, Bowman capsule, and mesangial cells are called the renal corpuscle.
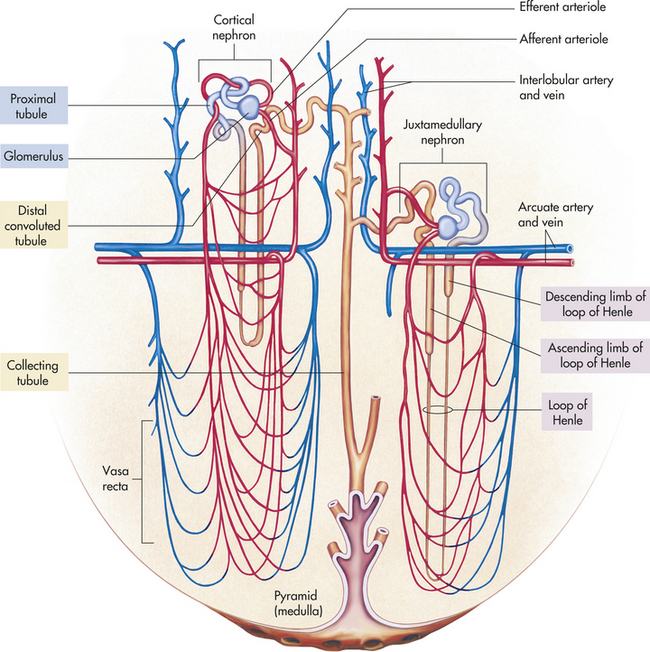
Figure 35-5 The nephron unit with its blood vessels. Blood flows through nephron vessels as follows: interlobular artery, afferent arteriole, glomerulus, efferent arteriole, peritubular capillaries (around the tubules), venules, interlobular vein. (From Patton KT, Thibodeau GA: Anatomy & physiology, ed 7, St Louis, 2010, Mosby.)
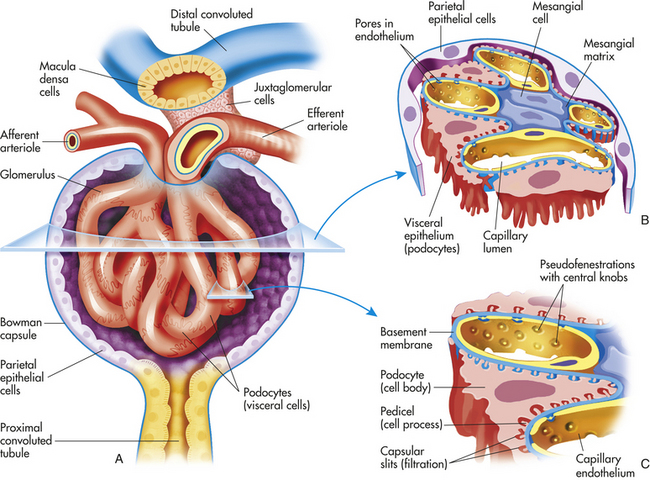
Figure 35-6 Anatomy of the glomerulus and juxtaglomerular apparatus. A, Longitudinal cross section of glomerulus and juxtaglomerular apparatus. B, Horizontal cross section of glomerulus. C, Enlargement of glomerular capillary filtration membrane.
The wall of the glomerular capillary serves as a filtration membrane (the glomerular filtration membrane) and has three layers: (1) an inner capillary endothelium, (2) a middle basement membrane, and (3) an outer layer of capillary epithelium (also called podocytes or visceral epithelium). Each layer has unique structural properties that allow all components of the blood to filter through, with the exception of blood cells and plasma proteins with a molecular weight greater than 70,000 (Figure 35-7; see also Figure 35-6). The glomerular endothelium is composed of cells in continuous contact with the basement membrane. Glomerular endothelial cells synthesize nitric oxide (a vasodilator) and endothelin-1 (a vasoconstrictor important to regulating glomerular blood flow). The glomerular endothelium is perforated by many small openings or windows, called fenestrae. The fenestrae are maintained by vascular epithelial growth factor (VEGF) produced by visceral epithelium. The middle basement membrane is a negatively charged, selectively permeable network of glycoproteins and mucopolysaccharides and may be secreted and maintained by the epithelial cells.2 The visceral epithelium, also called podocytes,3 has footlike processes that radiate and adhere to the basement membrane covering the glomerular capillaries. The visceral epithelium is reflected back at the vascular pole to become the parietal epithelium. The space between the visceral and parietal epithelia is the Bowman space, which continues to become the proximal tubule. The foot processes of one podocyte interlock with the foot processes of adjacent podocytes, forming an elaborate network of intercellular clefts. These clefts are called filtration slits (see Figure 35-7), or slit membranes, and modulate filtration. Nephrin, podocin, and CD2-associated protein are proteins that are exclusively located in the slit membrane and are required for normal filtration.4 The podocytes are endocytic, which allow molecules to enter the cell without passing through the cell membrane, and they prevent leakage of proteins into the urine.
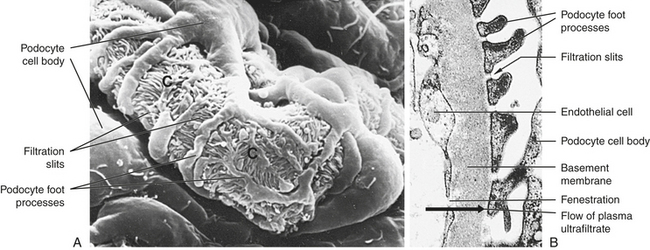
Figure 35-7 Glomerular capillary. A, Scanning electron micrograph of normal glomerular capillary (C). B, Glomerular capillary wall (× 40,000.) (From Kissane JM, editor: Anderson’s pathology, ed 9, St Louis, 1990, Mosby.)
The glomerular filtration membrane separates the blood within the glomerular capillaries from the filtered fluid in the Bowman space. The glomerular filtrate passes through the three layers of the glomerular membrane and forms the primary urine. The endothelial cells and basement membrane of the filtration membrane express negatively charged glycoproteins and form a filtration barrier to anionic proteins.
The glomerulus is supplied by the afferent arteriole and drained by the efferent arteriole. A group of specialized cells known as juxtaglomerular cells are located around the afferent arteriole where it enters the glomerulus (see Figures 35-3 and 35-6). Between the afferent and efferent arterioles is a portion of the distal convoluted tubule with specialized sodium and chloride-sensing cells known as the macula densa (see Figure 35-6). Together the juxtaglomerular cells and macula densa cells form the juxtaglomerular apparatus (JGA) (see Figure 35-6). Control of renal blood flow, glomerular filtration, and renin secretion occurs at this site.
The proximal tubule continues from the Bowman space and has an initial convoluted segment (pars convoluta) and then a straight segment (pars recta) that descends toward the medulla (see Figure 35-3). The proximal tubular lumen consists of one layer of cuboidal cells with a surface layer of microvilli that increases reabsorptive surface area. This is the only surface inside the nephron where the cells are covered with microvilli (a brush border) (see Figure 35-4). The proximal tubule joins the loop of Henle, a hairpin-shaped loop composed of thick and thin portions of a descending segment that goes into the medulla. The tube then loops and becomes the thickening ascending segment that extends toward the cortex. The thin segment is composed of thin squamous cells with no active transport function. The cells of the thick segment are cuboidal and actively transport several solutes but not water.
The more numerous cortical nephrons have glomeruli originating close to the surface of the cortex or in the midcortex, unlike the juxtamedullary nephrons, whose glomeruli are located deep in the cortex close to the medulla. The major structural difference between the glomeruli in the two types of nephrons is the length of the loop of Henle. In cortical nephrons the loop is short and may not extend into the medulla. The loop of Henle for the juxtamedullary nephrons, however, may extend the whole length of the medulla (40 mm). Juxtamedullary nephrons represent about 12% of the total number of nephrons and are important for the concentration and dilution of urine.
The distal tubule has convoluted and straight segments. It extends from the macula densa to the collecting duct. The collecting duct is a large tubule that descends down the cortex, through the renal pyramids of the inner and outer medullae, and into the minor calyx. The collecting duct is composed of two cell types: principal cells and intercalated cells (see Figure 34-4). Principal cells resorb sodium and water and secrete potassium. Intercalated cells secrete either hydrogen or bicarbonate and reabsorb potassium.
Blood Vessels
The blood vessels of the kidney closely parallel nephron structure. The renal arteries arise as the fifth branches of the abdominal aorta. At the renal hilum they divide into anterior and posterior branches and then subdivide into lobar arteries that supply blood to the lower, middle, and upper thirds of the kidney. The interlobar arteries are further subdivisions that travel down the renal columns and between the pyramids. At the cortical medullary junction, interlobar arteries branch into the arcuate arteries that arch over the base of the pyramids and run parallel to the surface of the kidney.
The interlobular arteries arise from the arcuate arteries and extend through the cortex toward the periphery and form the afferent glomerular arterioles (see Figure 35-5). The afferent arterioles subdivide into a fistlike structure of four to eight glomerular capillaries (see Figure 35-6). The glomerular capillaries empty into the efferent arteriole, which conveys blood to a second capillary bed, the peritubular capillaries. This is the only place in the body where an arteriole is positioned between two capillary beds. Increases or decreases in the resistance of the afferent and efferent arterioles increase or decrease glomerular filtration.
The peritubular capillaries surround the convoluted portions of the proximal and distal tubules and the loop of Henle (see Figure 35-5). The peritubular capillaries are adapted differently for the cortical and juxtamedullary nephrons. The peritubular capillaries surrounding the tubules of the cortical nephrons are similar to capillaries in other tissues. For the juxtamedullary nephrons a network of capillaries called the vasa recta forms loops and closely follows the loops of Henle. The capillaries of the vasa recta are the only blood supply to the medulla. They influence the osmolar concentration of the medullary extracellular fluid, which is important to the formation of a concentrated urine. All capillaries then drain into the venous system. The renal veins follow the arterial path in a reverse direction and have the same names as the arteries. The renal vein empties into the inferior vena cava. The lymphatic vessels tend also to follow the distribution of the blood vessels.
Urinary Structures
The urine formed by the nephrons flows from the distal tubules and collecting ducts through the duct of Bellini, the renal papillae (projections of the ducts), and into the calyces and is collected in the renal pelvis (see Figure 35-2). From the renal pelvis, urine is funneled into the ureters. Each adult ureter is approximately 30 cm long and is composed of long, intertwining muscle bundles. The lower ends of the ureters pass obliquely through the posterior aspect of the bladder wall. The close approximation of muscle cells permits the direct transmission of electrical stimulation, and the resulting peristaltic activity propels urine into the bladder. Peristaltic activity is affected by urine volume. When urine flow is slow, the contraction is segmented, with downward propulsion of urine. Increasing flow rates increase peristalsis. Peristalsis is maintained even when the ureter is denervated, so ureters can be transplanted.
Sensory innervation for the upper part of the ureter arises from the tenth thoracic nerve roots, with referred pain to the umbilicus. The innervation of lower segments arises from the sacral nerves with referred pain to the vulva or penis. The ureters have a rich blood supply. The primary arteries come from the kidney with contributions from the lumbar and superior vesical arteries. Contraction of the bladder during micturition (urination) compresses the lower end of the ureter, preventing reflux.
Bladder and Urethra
The bladder is a bag composed of a basket weave of smooth muscle fibers that forms the detrusor muscle and its smooth lining of transitional epithelium (uroepithelium). As the bladder fills with urine, it distends and the layers of transitional epithelium slide past each other and become thinner as the volume of the bladder increases. The uroepithelium maintains an important barrier function to prevent movement of water and solutes between the urine and blood.5 The trigone is a smooth triangular area lying between the openings of the two ureters and the urethra (Figure 35-8). The position of the bladder varies with age and gender. In infants and young children the bladder rises above the symphysis pubis, providing easy access for percutaneous aspiration. In adults it lies in the true pelvis, in front of the rectum and in front of the uterus in women. Inferiorly, the bladder sits on the prostate in men and on the anterior vagina in women. The bladder has a profuse blood supply, accounting for the bleeding that readily occurs with trauma, surgery, or inflammation.
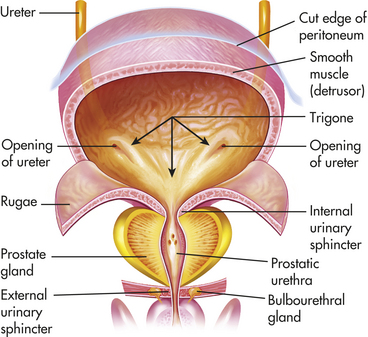
Figure 35-8 Structure and location of the urinary bladder. Frontal view of a dissected urinary bladder (male) in a fully distended position. (From Patton KT, Thibodeau GA: Anatomy & physiology, ed 7, St Louis, 2010, Mosby.)
The urethra extends from the inferior side of the bladder to the outside of the body. Two muscles called sphincters control excretion of urine from the bladder through the urethra. A ring of smooth muscle forms the internal urethral sphincter at the junction of the urethra and bladder. The external urethral sphincter is composed of striated muscles and is under voluntary control. The entire urethra is lined with mucus-secreting glands. The female urethra is short (3 to 4 cm). The male urethra is long (18 to 20 cm) and has three segments: prostatic, membranous, and cavernous. The prostatic urethra is closest to the bladder. It passes through the prostate gland and contains the openings of the ejaculatory ducts. The membranous urethra is the segment that passes through the floor of the pelvis. The cavernous segment forms the remainder of the tube. The cavernous segment is surrounded by erectile tissue and contains the openings of the bulbourethral mucous glands.
The innervation of the bladder and internal urethral sphincter is supplied by parasympathetic fibers of the autonomic nervous system. They primarily pass with the arteries to and from the sacral levels of the spinal cord. Sensory fibers may extend as high as the T6 portion of the spinal cord. Motor fibers from the pudendal nerve supply the external urethral sphincter. The reflex arc required for micturition is stimulated by mechanoreceptors that respond to stretching of tissue. The mechanoreceptors sense bladder fullness and send impulses to the sacral level of the cord with bladder filling. When the bladder accumulates 250 to 300 ml of urine, the bladder contracts and the internal urethral sphincter relaxes through activation of the spinal reflex arc (known as the micturition reflex). At this time a person feels the urge to void. In older children and adults, the reflex can be inhibited or facilitated by impulses coming from the brain, resulting in voluntary control of micturition.
RENAL BLOOD FLOW
The kidneys are highly vascular organs and usually receive 1000 to 1200 ml of blood per minute, or about 20% to 25% of the cardiac output. With a normal hematocrit of 45%, about 600 to 700 ml of blood flowing through the kidney per minute is plasma. From the renal plasma flow (RPF), 20% (approximately 120 to 140 ml/minute) is filtered at the glomerulus and passes into the Bowman capsule. The filtration of the plasma per unit of time is known as the glomerular filtration rate (GFR), which is directly related to the perfusion pressure in the glomerular capillaries.
The remaining 80% (about 480 ml) of plasma flows through the efferent arterioles to the peritubular capillaries. The ratio of glomerular filtrate to RPF per minute (120/600 = 0.20) is called the filtration fraction. Normally all but 1 to 2 ml of the glomerular filtrate is reabsorbed and returned to the circulation by the peritubular capillaries.
The GFR is directly related to renal blood flow (RBF), which is regulated by intrinsic autoregulatory mechanisms, neural regulation, and hormonal regulation. In general, blood flow to any organ is determined by the arteriovenous pressure differences across the vascular bed. If mean arterial pressure decreases or vascular resistance increases, RBF decreases.
Autoregulation
In the kidney a local mechanism of autoregulation tends to keep the rate of glomerular blood flow and therefore the GFR fairly constant over a range of arterial pressures between 80 and 180 mmHg (Figure 35-9). This means that changes in afferent arteriolar resistance and arteriolar pressure occur in the same direction. For example, as systemic blood pressure increases, the afferent arterioles constrict, preventing an increase in glomerular blood flow and filtration pressure. Opposite processes occur with a decrease in systemic blood pressure. Therefore, RBF and GFR are relatively constant. This “constant” state is maintained by intrinsic autoregulatory mechanism mediating the arteriolar resistance changes. The purpose of renal autoregulation is to prevent wide fluctuations in systemic arterial pressure from being transmitted to the glomerular capillaries. In this way, large fluctuations in GFR are prevented and solute and water excretion is constantly maintained when arterial pressure changes.6 Autoregulation may also protect the kidney from damage by hypertension.7
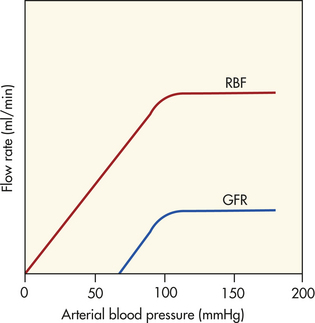
Figure 35-9 Renal autoregulation. Blood flow and glomerular filtration rate are stabilized in the face of changes in perfusion pressure. (From Berne RM, Levy MN, editors: Principles of physiology, ed 3, St Louis, 2000, Mosby.)
One mechanism responsible for the autoregulatory response in the kidney is probably a myogenic mechanism. As arterial pressure declines, the stretch on the afferent arteriolar smooth muscle decreases and the arteriole relaxes, with an increase in RBF; an increase in arteriolar pressure causes the arteriole smooth muscle to contract and decreases RBF. Tubuloglomerular feedback is a second mechanism for autoregulation of RBF and GFR. The macula densa cells of the distal tubule in the JGA sense changes in flow rate and sodium chloride content of the lumenal fluid. This information initiates a signal causing compensatory changes in afferent arteriolar resistance and GFR.8
Neural Regulation
The blood vessels of the kidney are innervated by the sympathetic noradrenergic fibers that cause arteriolar vasoconstriction and reduce renal blood flow. The innervation of the kidney comes primarily from the celiac ganglion and greater splanchnic nerve (see Figure 14-24). The afferent and efferent arterioles are richly innervated, but nerves have not been observed in the glomerular capillaries.
The RBF is reflexively related to the systemic arterial pressure. When systemic arterial pressure decreases, increased renal sympathetic nerve activity is mediated reflexively through the carotid sinus and the baroreceptors of the aortic arch. This stimulates renal arteriolar vasoconstriction and decreases RBF and GFR. Thus RBF still changes when systemic arterial pressure is significantly reduced, although autoregulatory processes dampen the response. The decreased RBF decreases the GFR and diminishes excretion of sodium and water, promoting an increase in blood volume and thus an increase in systemic pressure. The afferent and efferent arterioles are innervated by sympathetic nerves. Norepinephrine causes vasoconstriction by activation of α1-adrenoreceptors on afferent arterioles. The nerves are stimulated by decreased blood volume and cause vasoconstriction and decreased glomerular filtration.
Exercise, body position, and hypoxia also influence RBF. Exercise and change of body position activate renal sympathetic neurons and cause mild vasoconstriction. Severe hypoxia stimulates the chemoreceptors of the carotid and aortic bodies and decreases RBF by means of sympathetic stimulation. Hemorrhage induces intense sympathetic stimulation and vasoconstriction, and GFR and blood flow are reduced.
Hormones and Other Factors
Hormonal factors and many mediators can alter the resistance of the renal vasculature by stimulating vasodilation or vasoconstriction. A major hormonal regulator of RBF is the renin-angiotensin-aldosterone system, which can increase systemic arterial pressure, change RBF and increase sodium reabsorption. Renin is an enzyme formed and stored in granular cells of the afferent arterioles of the JGA (see Figure 35-3). Several complex physiologic mechanisms stimulate the release of renin. These mechanisms are principally decreased blood pressure in the afferent arterioles, which reduces stretch of the juxtaglomerular cells; decreased sodium chloride concentration in the distal convoluted tubule; sympathetic nerve stimulation of β-adrenergic receptors on the juxtaglomerular cells and prostaglandins.9
When renin is released, it cleaves an α-globulin (angiotensinogen produced by liver hepatocytes) in the plasma to form angiotensin I, which is physiologically inactive. In the presence of angiotensin-converting enzyme (ACE) produced from pulmonary and renal endothelium, angiotensin I is converted to angiotensin II. Angiotensin II stimulates secretion of aldosterone by the adrenal cortex (see Chapter 20), is a potent vasopressor, inhibits renin release, and stimulates antidiuretic hormone (ADH) secretion and thirst. Vitamin D3 is a potent negative endocrine regulator of renin gene expression.10 Numerous physiologic effects of the renin-angiotensin-aldosterone system serve the purpose of stabilizing systemic blood pressure and preserving the extracellular fluid volume during hypotension or hypovolemia, including sodium reabsorption, potassium excretion, systemic vasoconstriction, sympathetic nerve stimulation, thirst stimulation, and drinking. (The combined effects of the renin-angiotensin-aldosterone system are summarized in Figure 35-10.) ACE inhibitors are drugs that decrease blood pressure.
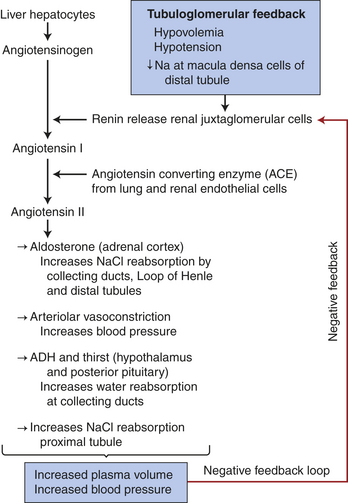
Figure 35-10 Renin-angiotensin-aldosterone system. Activation of tubuloglomerular feedback mechanisms stimulates the release of renin with activation of the renin-angiotensin-aldosterone system. Plasma volume and blood pressure are increased with the reabsorption of sodium chloride and water from the renal tubules. The restoration of plasma volume and blood pressure then decrease the release of renin, forming a negative feedback loop.
Natriuretic peptides are a group of peptide hormones including atrial natriuretic peptide (ANP) secreted from cells in the right atrium, brain natriuretic peptide (BNP) secreted from the cardiac ventricles, C-type natriuretic peptide from vascular endothelium, and urodilantin secreted by the distal tubules and collecting ducts.11 When the heart dilates during volume expansion or heart failure, ANP and BNP inhibit secretion of renin, inhibit angiotensin-induced secretion of aldosterone, vasodilate the afferent and constrict the efferent glomerular arterioles, and inhibit sodium and water absorption by kidney tubules. C-type natriuretic peptide is a vasodilator, and urodilantin promotes renal sodium chloride excretion. The result is increased urine formation and decreased blood volume and blood pressure.12 Other hormones and mediators that influence renal blood flow are summarized in Table 35-1.
Table 35-1
Hormones, Mediators, and Renal Blood Flow
| Hormone or Mediator | Effect on Renal Blood Flow |
| Adenosine | Produced within kidney; causes vasoconstriction of afferent arteriole; decreases RBF and GFR |
| Angiotensin II | Produced systemically and within kidneys; constricts afferent and efferent arterioles; decreases RBF and GFR |
| Atrial and brain natriuretic peptides | Produced by atria and ventricles of the heart with hypertension and increased blood volume; causes vasodilation of afferent arteriole and vasoconstriction of efferent arteriole; modest increase in GFR with little change in RBF |
| Bradykinin | Produced in kidney from kininogen and causes vasodilation by release of nitric oxide and prostaglandins; increases RBF and GFR |
| Dopamine | Produced by the proximal tubule; increases RBF; inhibits renin secretion |
| Endothelin | Produced by renal vessel endothelial cells, mesangial cells, and distal tubule cells in response to bradykinin, angiotensin II, epinephrine, and stretch; most active with renal disease; profound vasoconstriction of afferent and efferent arterioles; decreases RBF and GFR |
| Histamine | Produced locally within the kidney; modulates RBF in basal state and during inflammation; increases RBF by decreasing afferent and efferent arteriolar resistance and does not decrease GFR |
| Nitric oxide | Produced by renal vessel endothelial cells with increased stretch and by stimulation of acetylcholine, histamine, bradykinin, ATP; increases vasodilation of afferent and efferent arterioles |
| Prostaglandins, PGI2, PGE2 | Produced locally within kidney with decreased RBF; dampen vasoconstriction caused by sympathetic nerves and angiotensin II; prevent harmful vasoconstriction and renal ischemia |
| Urodilantin (a natriuretic peptide) | Produced by distal tubule and collecting duct when there is increased circulating volume and increased blood pressure; inhibits sodium and water reabsorption from medullary part of collecting duct, thereby producing diuresis |
ATP, Adenosine triphosphate; GFR, glomerular filtration rate; RBF, renal blood flow.
KIDNEY FUNCTION
The nephron can perform many functions simultaneously. It filters the plasma at the glomerulus and reabsorbs and secretes different substances at various parts of its tubular structure (Figure 35-11). The function of the nephron is to form a filtrate of protein-free plasma. This process, known as ultrafiltration, occurs across the glomerular capillaries. The nephron then regulates the filtrate to maintain body fluid volume, electrolyte composition, and pH within narrow limits.
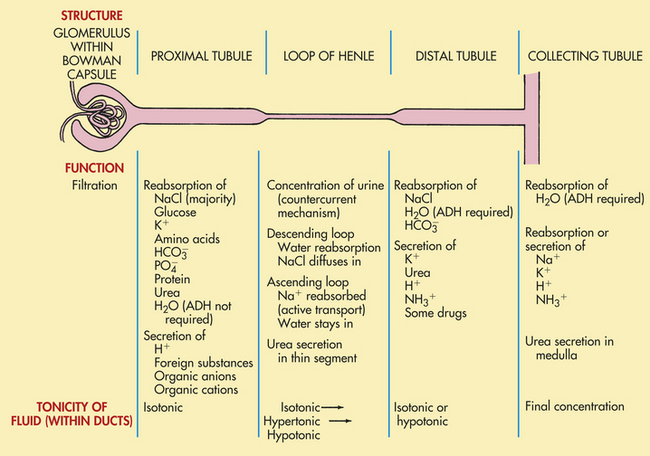
Figure 35-11 Major functions of nephron segments. ADH, Antidiuretic hormone. (Modified from Hockenberry MJ: Wong’s nursing care of infants and children, ed 8, St Louis, 2007, Mosby.)
Regulation of the filtrate occurs through two processes: tubular reabsorption and tubular secretion. Tubular reabsorption is the movement of fluids and solutes from the tubular lumen to the peritubular capillary plasma. Transfer of substances from the plasma of the peritubular capillary to the tubular lumen is tubular secretion. The transport mechanisms are active as well as passive (processes defined in Chapter 1). The elimination of a substance in the final urine is known as excretion (Figure 35-12).
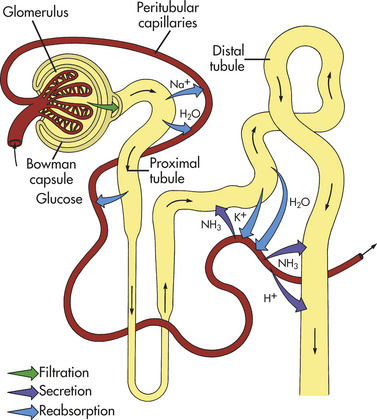
Figure 35-12 Urine formation: glomerular filtration, tubular reabsorption, and tubular secretion. The three processes by which the kidneys excrete urine. From proximal convoluted tubules, sodium and glucose are reabsorbed into peritubular capillaries by active transport. Water reabsorption by osmosis follows. From distal convoluted tubules sodium is reabsorbed by active transport. Osmotic reabsorption of water from them occurs when ADH is present. Secretion of ammonia and hydrogen occurs from peritubular capillaries into distal tubules by active transport. (From Patton KT, Thibodeau GA: Anatomy & physiology, ed 7, St Louis, 2010, Mosby.)
Glomerular Filtration
The fluid filtered by the glomerular capillary filtration membrane is protein free but contains electrolytes such as sodium, chloride, and potassium and organic molecules such as creatinine, urea, and glucose in the same concentrations as in plasma. Like other capillary membranes, the glomerulus is freely permeable to water and relatively impermeable to large colloids such as plasma proteins. The size of the molecules and their electrical charge are important factors affecting the permeability of substances crossing the glomerulus. The small size of the filtration slits or pores in the membrane restricts the passage of proteins and other macromolecules. The negative charge along the filtration membrane further impedes the passage of negatively charged macromolecules (because like forces repel each other). Positively charged macromolecules therefore permeate the membrane more readily than neutrally charged particles.
Capillary pressure, as well as electrical charge, has an effect on glomerular filtration. The hydrostatic pressure within the capillary is the major force for inducing water and solutes across the filtration membrane and into the Bowman capsule. This pressure is determined indirectly by the efficiency of cardiac contraction and directly by the systemic arterial pressure and the resistances to blood flow in the afferent and efferent arterioles. Two forces oppose the filtration effects of the glomerular capillary hydrostatic pressure (PGC): (1) the hydrostatic pressure in the Bowman space (PBC) and (2) the effective oncotic pressure of the glomerular capillary blood (πGC). (As explained in Chapter 3, hydrostatic pressure is a pushing force in relation to water, and oncotic pressure is a pulling force.) Because the fluid in the Bowman space normally contains only minute amounts of protein, it normally does not have an oncotic influence on the plasma of the glomerular capillary (Figure 35-13).
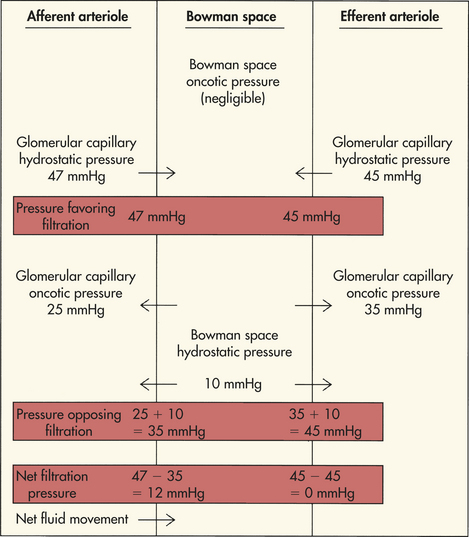
The combined effect of forces favoring and forces opposing filtration determines the filtration pressure. The net filtration pressure (NFP) is the sum of forces favoring and opposing filtration and is expressed by the following equation:
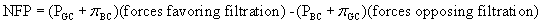
The estimated values contributing to the forces of net filtration are presented in Table 35-2.
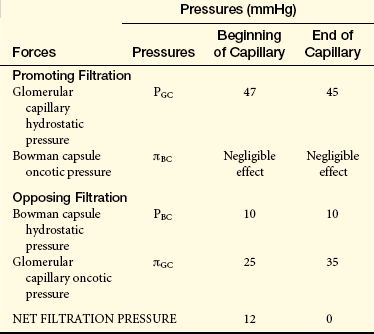
As the protein-free fluid is filtered into the Bowman capsule, the plasma oncotic pressure increases and the hydrostatic pressure decreases. The increase in glomerular capillary oncotic pressure is great enough to reduce the net filtration pressure to zero at the efferent end of the capillary and to stop the filtration process effectively. The low hydrostatic pressure and increased oncotic pressure in the efferent arteriole then are transferred to the peritubular capillaries and facilitate reabsorption of fluid from the proximal tubules.
Filtration Rate: The total volume of fluid filtered by the glomeruli averages 180 L/day, or approximately 120 ml/minute, a phenomenal amount considering the size of the kidneys. Because only 1 to 2 L of urine is excreted per day, 99% of the filtrate is reabsorbed into the peritubular capillaries and returned to the blood. The factors determining the GFR are directly related to the pressures that favor or oppose filtration. Any changes in afferent or efferent arteriolar resistance will alter glomerular capillary hydrostatic pressure and GFR. Vasoconstriction of one or the other of these two arterioles produces opposite effects on glomerular pressure. For example, if the afferent arteriole constricts, blood flow decreases, with a corresponding drop in glomerular pressure. The GFR then decreases, and body fluids are conserved. Conversely, constriction of the efferent arteriole increases the net filtration pressure, and the GFR increases. When both afferent and efferent arterioles constrict, little change occurs in filtration pressure, but RBF is reduced and so is the GFR.
Obstruction to the outflow of urine (caused by strictures, stones, or tumors along the urinary tract) can cause a retrograde increase in pressure at the Bowman capsule and a decrease in GFR. Excessive loss of protein-free fluid from vomiting, diarrhea, use of diuretics, or excessive sweating can increase glomerular capillary oncotic pressure and decrease the GFR. Renal disease also can cause changes in pressure relationships by altering capillary permeability and the surface area available for filtration (see Chapter 36).
Tubular Transport: By the time fluid reaches the end of the proximal tubule, approximately 60% to 70% of filtered sodium and water and about 50% of urea have been reabsorbed, along with 90% or more of potassium, glucose, bicarbonate, calcium, phosphate, amino acids, and uric acid. All this occurs by active transport. Chloride, water, and urea are reabsorbed passively but are linked to the active transport of sodium (cotransport). Active transport in the renal tubules can be limited as the carrier molecules become saturated, a phenomenon known as transport maximum (Tm). Transport maximums exist for most substances actively transported by the tubular epithelium. The reabsorption of glucose is a significant example. Glucose is coupled to sodium transport and is almost completely reabsorbed in the proximal tubule. Like other actively transported substances, glucose has a maximal transport capacity, or renal threshold. This means that when the carrier molecules for glucose become saturated, the excess will be excreted in the urine. Normally the plasma level and filtered glucose load are not high enough to saturate the carrier mechanism. When the plasma glucose reaches 180 mg/dl, however, as occurs in the individual with uncontrolled diabetes mellitus, the threshold for glucose is achieved. Any further increase in the plasma level causes loss of glucose in the urine.
Proximal Tubule: Active reabsorption of sodium is the primary function of the proximal tubule. Water, most electrolytes, and organic substances are cotransported with sodium. The osmotic force generated by active sodium transport promotes the passive diffusion of water out of the tubular lumen and into the peritubular capillaries. Passive transport of water is further enhanced by the elevated oncotic pressure of the blood in the peritubular capillaries. The reabsorption of water leaves an increased concentration of urea within the tubular lumen, creating a gradient for its passive diffusion to the peritubular plasma.
As the positively charged sodium ions leave the tubular lumen, negatively charged chloride ions passively follow to maintain electroneutrality. Because the luminal membrane (the inside of the tubule) of the proximal tubular cell has a limited permeability to chloride, however, chloride reabsorption lags behind sodium.
Hydrogen ions are actively exchanged for sodium ions in the tubular lumen. The hydrogen ions (H+) then combine with bicarbonate () in the tubular lumen to form carbonic acid (H2CO3). The carbonic acid rapidly breaks down, or dissociates, to carbon dioxide (CO2) and water (H2O) then diffuse into the tubular cell, where carbonic anhydrase catalyzes the CO2 and H2O to form and H+. combines with sodium and is transported (reabsorbed) to the peritubular capillary blood. The H+ is not excreted but is again exchanged for sodium and reenters the lumen to recombine with .Thus, in the proximal tubule, for every H+ ion secreted into the tubular lumen, a ion enters the blood (see Figure 3-10, page 117).
Bicarbonate is freely filtered at the glomerulus, but is not highly permeable at the peritubular capillary membrane. As described above it combines with H+ and is reabsorbed as CO2 and H2O, which are readily diffusible. One of the unusual aspects of this process is that the bicarbonate molecule filtered at the glomerulus is not the same molecule that is reabsorbed (because it dissociates) and the hydrogen ion secreted by the proximal tubule is not excreted in the urine. Bicarbonate is thus conserved, and in this exchange, bicarbonate and hydrogen normally do not contribute to the urinary excretion of acid or the addition of acid to the blood. Approximately 90% of bicarbonate is reabsorbed by the proximal tubules.
In addition to the proximal tubular secretion of hydrogen ions, secretory transport mechanisms exist for creatinine, other organic bases, and endogenous and exogenous organic acids, including para-aminohippurate (PAH) and penicillin (Box 35-1). These secretory mechanisms are important for eliminating drugs and other exogenous chemical products from the body. Frequently, exogenous substances are conjugated with sulfate and glucuronic acid by the liver and then actively secreted by the renal tubules. This has important clinical implications because many drugs and their metabolites are eliminated from the body in this way. When the renal tubules are damaged, metabolic byproducts and drugs may accumulate, causing toxic levels in the body.
Loop of Henle and Distal Tubule: The filtrate entering and leaving the proximal tubule is essentially isoosmotic with the plasma and has a concentration of about 285 mOsm. Although approximately 65% of salt and water is reabsorbed along the proximal tubule, they are reabsorbed in equal amounts, causing only minor changes in the osmotic and electrolyte concentrations of the fluid flowing into the loop of Henle. Therefore, any concentration or dilution of urine occurs at more distal sites of the nephron, principally in the medullary loop of Henle and collecting ducts. Near the top of the renal pyramids, the interstitial osmolality reaches 1200 mOsm/L.
These quantitative changes taking place in the loop of Henle are related to the length of the loop and its depth of penetration into the medulla. The structural features of the medullary hairpin loops provide the kidney with the ability to concentrate urine and conserve water for the body. The transition of the filtrate into urine is a function of the concentrating ability of the loops and final adjustments in urine composition made by the distal tubule and collecting duct.
The primary function of the loop of Henle is to establish a hyperosmotic state within the medullary interstitial fluid. This is achieved by reabsorbing more solute than water into the interstitium. The fluid leaving the ascending limb of the loop is therefore hypoosmotic, or more dilute than the fluid that entered. This dilution allows the distal tubule and collecting duct to make final adjustments in the concentration or dilution of the excreted urine according to body needs. The vasa recta act to maintain the high osmotic gradient established by the loop of Henle.
Different transport or permeability functions of the loop of Henle are important for dilution and concentration of urine. The thin, descending segment of the loop of Henle is highly permeable to water and moderately permeable to sodium, urea, and other solutes. The thin, ascending segment is more permeable to solutes and almost impermeable to water. The thick portion of the ascending segment is highly permeable to sodium, potassium, and chloride and significantly less permeable to water and urea. Tamm-Horsfall glycoprotein, also known as uromedulin, is formed on the epithelial surface of the thick ascending segment and the first segment of the distal tubule. It is the most abundant urinary protein and protects against bacterial adhesion and urolithiasis, and is a ligand for lymphokines.13
The convoluted portion of the distal tubule is poorly permeable to water but readily absorbs ions and contributes to the dilution of the tubular fluid. The later, straight segment of the distal tubule and the collecting duct are permeable to water as controlled by ADH. Sodium is readily absorbed by the later segment of the distal tubule and collecting duct under the regulation of the hormone aldosterone (see Chapter 20). Potassium is actively secreted by principal cells and is reabsorbed in lesser amounts by intercalated cells in these segments. Potassium also is controlled by aldosterone and other factors related to the concentration of potassium in body fluids.14
Hydrogen is also secreted by the distal tubule and combines with nonbicarbonate buffers (ammonium and phosphate) for the elimination of excess acids in the urine. The distal tubule thus contributes to the regulation of acid-base balance by excreting hydrogen ions into the urine and by adding new bicarbonate to the plasma (see Figure 3-10, page 117). The mechanism is similar to the conservation of bicarbonate by the proximal tubule, except that the hydrogen ion is excreted in the urine. (The specific mechanisms of acid-base balance and acid excretion are described in Chapter 3.)
Glomerulotubular Balance: To regulate body fluid balance, the kidney must not reabsorb or excrete too much sodium or water. Normally 99% of the glomerular filtrate is reabsorbed. When the GFR spontaneously decreases or increases, the renal tubules, primarily the proximal tubules, automatically adjust their rate of reabsorption of sodium and water to balance the change in GFR. Thus a constant fraction of filtered sodium and water is reabsorbed from the proximal tubule. This prevents wide fluctuations in sodium and water excretion into the urine and maintains sodium and water balance.15
Concentration and Dilution of Urine
The production of a concentrated urine involves a countercurrent exchange system, in which fluid flows in opposite directions through parallel tubes. A concentration gradient causes fluid to be exchanged across the parallel pathways. In the nephron the fluid moves up and down the parallel sides of the hairpin loop of Henle in the medulla. The longer the loop, the greater the concentration gradient because the concentration gradient increases from the cortex to the tip of the medulla. The loops of Henle serve as multipliers of the concentration gradient, and the vasa recta act as a countercurrent exchanger for maintaining the gradient.15
Water, Sodium, and Chloride
The process is initiated in the thick ascending limb of the loop of Henle with the active transport of chloride and sodium out of the tubular lumen and into the medullary interstitium (Figure 35-14). Because the lumen of the ascending limb is impermeable to water, water cannot follow the sodium/chloride transport. This lack of luminal permeability causes the ascending tubular fluid to become hypoosmotic and the medullary interstitium to become hyperosmotic. The descending limb of the loop, which receives fluid from the proximal tubule, is highly permeable to water, but it is the only place in the nephron that does not actively transport either sodium or chloride. Sodium and chloride may, however, diffuse into the descending tubule from the interstitium. The hyperosmotic interstitium causes water to move out of the descending limb, and the remaining fluid in the descending tubule becomes increasingly concentrated as it flows toward the tip of the medulla. As the tubular fluid rounds the loop and enters the ascending limb, sodium and chloride are removed and water is retained. The fluid then becomes more and more dilute as it encounters the distal tubule.
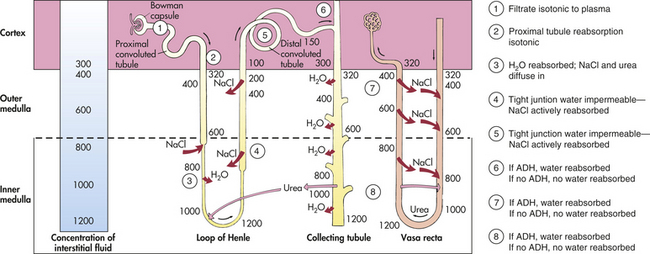
Figure 35-14 Countercurrent mechanism for concentrating and diluting urine. ADH, Antidiuretic hormone. (NOTE: Numbers on illustration represent milliosmoles [mOsm].)
The slow rate of blood flow and the hairpin structure of the vasa recta allow blood to flow through the medullary tissue without disturbing the osmotic gradient. As blood flows into the descending limb of the vasa recta, it encounters the increasing osmotic concentration gradient of the medullary interstitium. Water moves out, and sodium and chloride diffuse into the descending vasa recta. The plasma becomes increasingly concentrated as it flows toward the tip of the medulla.
As the blood flow passes into the ascending limb and back toward the cortex, the surrounding interstitial fluid becomes comparatively more dilute. Water then moves back into the vasa recta, and sodium and chloride diffuse out. The net result is a preservation of the medullary osmotic gradient. If blood were to flow rapidly through the vasa recta, as occurs in some renal diseases, the medullary concentration gradient would be washed away and the ability to concentrate urine and conserve water would be lost. The efficiency of water conservation is related to the length of the loops: the longer the loops, the greater the ability to concentrate the urine. Many desert animals have very long loops and can reabsorb water so efficiently that they rarely need to drink.
Urea
Urea is an end product of protein metabolism and is the major constituent of urine along with water. The glomerulus freely filters urea, and tubular reabsorption of urea depends on urine flow rate with less reabsorption at higher flow rates. Approximately 50% of urea is excreted in the urine, and 50% is recycled within the kidney. The recycling of urea from the tubules and collecting ducts contributes to the osmotic gradient within the medulla and is necessary for the concentration and dilution of urine. Because urea is an end product of protein metabolism, individuals with protein deprivation cannot maximally concentrate their urine.
Catecholamines
With hemorrhage or extracellular fluid depletion, sympathetic nerves are activated to release norepinephrine and dopamine, and the adrenal medulla releases epinephrine. Norepinephrine and epinephrine promote afferent arteriolar vasoconstriction and decrease GFR and RBF.15 Renalase is a hormone produced by the kidney that degrades catecholamines and may be important in blood pressure regulation.16
Antidiuretic Hormone
The distal tubule in the cortex receives the hypoosmotic urine from the ascending limb of the loop of Henle. The concentration of the final urine is controlled by antidiuretic hormone (ADH), which is secreted from the posterior pituitary, or neurohypophysis. ADH increases water permeability in the last segment of the distal tubule and along the entire length of the collecting ducts, which pass through the inner and outer zones of the medulla.
In the presence of ADH, water reabsorption is high. Most of the water is reabsorbed in the medullary collecting ducts because of the high osmotic gradient in the medullary interstitium. The water diffuses into the ascending limb of the vasa recta and returns to the systemic circulation. The excreted urine can have a high osmotic concentration, up to 1400 mOsm. The volume is normally reduced to about 1% of what was filtered at the glomerulus.
ADH secretion is therefore one cause of oliguria, or diminished excretion of urine, that is, less than 400 ml/day or 30 ml/hr. Fluid imbalance may be related to the syndrome of inappropriate secretion of ADH, which is a cause of water excess (see Chapters 3 and 21). Inadequate secretion of ADH occurs in diabetes insipidus, causing the excretion of a large volume of dilute urine.
In the absence of ADH, water diuresis, an increase in excretion of a highly dilute urine, takes place. The distal tubules and collecting ducts become impermeable to water. Water remains in the tubular lumen and is excreted as a dilute and large volume of urine. Because ADH has no effect on sodium reabsorption, it continues to be actively transported from the distal tubule. (The mechanism for the regulation of ADH and plasma osmolality is described in Chapter 3.)
Natriuretic Peptides
The natriuretic peptides (urodilantin, ANP, and BNP) promote diuresis and were described on p. 1352.
Diuretics as a Factor in Urine Flow
A diuretic is any agent that enhances the flow of urine. Clinically, diuretics interfere with renal sodium reabsorption and reduce extracellular fluid volume. Diuretics are commonly used to treat hypertension and edema caused by heart failure, cirrhosis, and nephrotic syndrome.
Different diuretics affect different sites of tubular function and may produce side effects that alter acid-base and electrolyte balance. Therefore, health professionals need to understand their indications for use, mechanisms of action, and toxic side effects. Diuretics are divided into five general categories: (1) osmotic diuretics, (2) carbonic anhydrase inhibitors (inhibitors of urinary acidification), (3) inhibitors of loop sodium or chloride transport, (4) aldosterone antagonists, and (5) aquaretics. (The physiologic mechanism related to each category is summarized in Table 35-3.)

Renal Hormones
Certain hormones are either activated or synthesized by the kidney. These hormones have significant systemic effects and include the active form of vitamin D, erythropoietin; renin-angiotensin and aldosterone and natriuretic hormones (see p. 1352).
Vitamin D
Vitamin D is a hormone that can be obtained in the diet or synthesized by the action of ultraviolet radiation on cholesterol in the skin. These forms of vitamin D (cholecalciferol) are inactive and require two hydroxylations to establish a metabolically active form. The first step occurs in the liver with hydroxylation at the 25th carbon (calcifediol), and the second hydroxylation occurs at the first carbon position in the kidneys and is stimulated by parathyroid hormone. The end product is 1,25-dihydroxycholecalciferol, or 1,25-dihydroxyvitamin D3 (1,25-OH2D3) (calcitriol), the active form of vitamin D.17
Calcitriol (vitamin D3) is necessary for the absorption of calcium and phosphate by the small intestine. A decreased plasma calcium level (less than 10 mg/dl) stimulates the secretion of parathyroid hormone. Parathyroid hormone then stimulates a sequence of events that help restore plasma calcium back toward normal including:
• Calcium mobilization from bone
• Absorption of calcium and phosphate from the intestine by stimulating renal activation of vitamin D
• Increased renal calcium and phosphate reabsorption and decreased secretion
Serum phosphate fluctuations also influence the renal hydroxylation of vitamin D. Decreased levels stimulate active 1,25-OH2D3 formation, and increased levels inhibit formation. This results in compensatory changes in phosphate absorption from bone and the intestine. The clinical significance of the role of the kidney in calcium and phosphate metabolism
is evident in renal disease. Patients with renal disease have a deficiency of 1,25-OH2D3 and manifest symptoms of disturbed calcium and phosphate balance (see Chapter 36).
Erythropoietin
Erythropoietin (Epo) is produced by the fetal liver and in the adult kidney and is essential for normal erythropoiesis. Epo stimulates the bone marrow to produce red blood cells in response to tissue hypoxia (see Chapter 25). The stimulus for erythropoietin release is decreased oxygen delivery in the kidneys. The anemia of chronic renal failure and cancer chemotherapy is treated with recombinant human erythropoietin (rH-Epo). Epo also affects endothelium and promotes angiogenesis, mitogenesis, and antiapoptosis.18
TESTS OF RENAL FUNCTION
A number of specific renal functions can be measured by renal clearance. Renal clearance techniques determine how much of a substance can be cleared from the blood by the kidneys per given unit of time. The application of this principle permits an indirect measure of GFR, tubular secretion, tubular reabsorption, and renal blood flow.
Clearance and Glomerular Filtration Rate
The GFR provides the best estimate of functioning renal tissue. Loss or damage to nephrons leads to a corresponding decrease in GFR. The measurement of GFR requires use of a substance that has a stable plasma concentration; is not protein bound; is freely filtered at the glomerulus; does not influence GFR; and is not secreted, reabsorbed, or metabolized by the tubules. Inulin (a fructose polysaccharide) is one substance that meets the criteria for measurement of GFR.
The kidney “clears” inulin from the plasma by filtering it at the glomerulus, reabsorbing nearly all of the fluid, and excreting the inulin left behind in the urine. The amount of inulin filtered is equal to the volume of plasma filtered (GFR) multiplied by the plasma concentration of inulin (PIN). The amount of inulin in the urine is equal to a volume of urine per unit of time (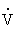 ) (usually 24 hours) multiplied by the inulin concentration of urine (UIN). Because all the inulin filtered is excreted in the urine,
) (usually 24 hours) multiplied by the inulin concentration of urine (UIN). Because all the inulin filtered is excreted in the urine,
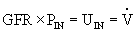
GFR can be calculated by rearranging the formula:
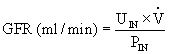
The accurate determination of inulin clearance requires constant infusion to maintain a stable plasma level. This is time consuming, inconvenient, and at risk for error. Therefore, the clearance of creatinine, a natural substance produced by muscle and released into the blood at a relatively constant rate, is commonly used clinically. It is freely filtered at the glomerulus, but a small amount is secreted by the renal tubules. Therefore, creatinine clearance overestimates the GFR but within tolerable limits. Creatinine clearance provides a good measure of GFR because only one blood sample is required in addition to a 24-hour volume of urine. The GFR estimated by creatinine clearance is calculated as follows:
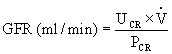
Similar calculations can be made for all solutes excreted in the urine per unit of time. Substances freely filtered at the glomerulus but with a clearance less than inulin or creatinine have been reabsorbed along the tubules. For example, glucose is completely reabsorbed and has a clearance rate of nearly zero. Conversely, substances secreted by the tubules have a clearance rate greater than inulin or creatinine (i.e., greater than 1). Numerous formulas have been developed for estimating GFR using creatine and other indicators.19
Clearance and Renal Blood Flow
The standard clearance formula also can be used to estimate RPF and RBF. The substance used for this evaluation is para-aminohippuric acid (PAH). Some PAH is filtered at the glomerulus, and most of the remainder is secreted into the tubules in one circulation through the kidney. If all the PAH were removed from the plasma during a single pass through the kidney, total RPF could be determined. Because the supporting and nonsecreting structures of the kidney receive 10% to 15% of effective renal blood flow (ERBF), clearance of PAH measures only what is known as the effective renal plasma flow (ERPF), which is 85% to 90% of the true renal plasma flow:
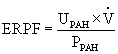
where CPAH = renal clearance of PAH, UPAH = PAH in urine, and PPAH = PAH in plasma.
The estimation of ERBF can then be calculated by considering the hematocrit in the following formula
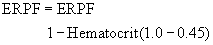
Blood Tests
Plasma Creatinine Concentration
A long-term decline in GFR over weeks or months is reflected in the plasma creatinine (PCR) concentration (normal value = 0.7 to 1.2 mg/dl). The PCR concentration has a stable value when the GFR is stable because creatinine has a constant rate of production as a product of muscle metabolism. The amount filtered is approximately equal to the amount excreted, and a small amount is secreted by kidney tubules. When the GFR declines, the PCR increases proportionately. Thus the GFR and PCR are inversely related. If the GFR were to decrease by 50%, the filtration and excretion of creatinine would be reduced by 50% and creatinine would accumulate in plasma to twice the normal value. Therefore, elevated PCR values represent decreasing GFR. In the new steady state, however, the total amount of creatinine excreted in the urine would remain the same because of the proportionate decrease in GFR and increase in PCR.
The application of this principle is simple and useful for monitoring progressive changes in renal function. The test is most valuable for monitoring the progress of chronic rather than acute renal disease because it takes 7 to 10 days for the plasma creatinine level to stabilize when GFR declines. Serial measures can be obtained over a long time and plotted as a curve of glomerular function. The PCR also becomes elevated during trauma or breakdown of muscle tissue. In such instances the value is then not useful for estimating GFR.
Plasma Cystatin C Concentration
Serum concentrations of cystatin C have been proposed for estimations of GFR, particularly in children.20 The reciprocal of the serum concentrations of cystatin C can be used as estimates of changes in GFR similar to measures of plasma creatinine concentration. The National Kidney Foundation publishes guidelines for estimating GFR for monitoring renal failure.21
Blood Urea Nitrogen
The concentration of urea nitrogen in the blood reflects glomerular filtration and urine-concentrating capacity. Because urea is filtered at the glomerulus, blood urea nitrogen (BUN) levels increase as glomerular filtration drops. Because urea is reabsorbed by the blood through the permeable tubules, the BUN rises in states of dehydration and acute and chronic renal failure when passage of fluid through the tubules is slowed. BUN also varies as a result of altered protein intake and protein catabolism and therefore is a poor measure of GFR. The normal range for BUN in the adult is 10 to 20 mg/dl of blood.
Urinalysis
Urinalysis is a noninvasive and relatively inexpensive diagnostic procedure. The best results are obtained from a fresh, cleanly voided specimen because decay permits changes in the composition of urine. Urinalysis includes evaluation of color, turbidity, protein, pH, specific gravity, sediment, and supernatant.
Urine color is normally a clear, light yellow because of urochrome and other pigments. When formed substances (crystals, blood cells, or casts) are in the urine, it appears turbid. Protein in the urine creates marked foaming when shaken, and the foam is yellow or orange when the urine contains bile pigments. Urine does not normally contain protein or bile.
Urine pH normally ranges between 5 and 6.5, but it may vary from 4.5 to 8. Urine is more alkaline after eating and then becomes less alkaline before the next meal. Because sleep is accompanied by intermittent hypoventilation, urine is more acidic on awakening.
Specific gravity is an estimated measure of the solute concentration of the urine. Specific gravity of any solution is measured by comparing the weight of the solution with an equal volume of distilled water. Hence specific gravity is not a true measure of the number or concentration of particles, but it correlates well with osmolality and is a useful clinical tool. Specific gravity usually is measured with a hydrometer in a cylinder of urine; the normal value is 1.016 to 1.022. Dipstick evaluations may be falsely high when urine pH is less than 6 and falsely low when the pH is more than 7.
The final urine osmolality is primarily a function of ADH, which controls water reabsorption in the collecting ducts. If the kidney is unable to concentrate or dilute urine, given a stimulus, the cause is usually a malfunction of the renal tubules or inappropriate ADH secretion by the posterior pituitary gland. The state of hydration also affects the urine specific gravity, so hydration status should be evaluated before making a diagnosis. This determination is helpful for differentiating oliguria caused by intrinsic renal disease from hypovolemia as a result of dehydration.
Urine Sediment
The urine sediment is examined microscopically and may contain cells, casts, crystals, and bacteria. Epithelial cells may be seen in the microscopic field because they are shed naturally throughout the urinary tract.
Red Blood Cells: Normal urine contains few or no red blood cells. If a large number of red cells are present, this is known as hematuria, and the sediment may be red. An alkaline or hypotonic urine causes lysis of red cells, however, so that the cells will not be seen. Urine then will be positive for hemoglobin, and the specific gravity will be elevated. Hematuria can occur with the administration of anticoagulants and with several renal diseases.
Casts: Casts (accumulations of cellular precipitates) originate in the renal tubules, from which they take their shape. They are cylindrical with distinct borders. All casts have a precipitated microprotein matrix and arise primarily from the ascending limb of the distal tubule. Red cell casts indicate bleeding into the tubules; white cell casts are associated with an inflammatory process. Epithelial cell casts indicate degeneration of the tubular lumen or necrosis of the renal tubules. The type of cast identified suggests the disease process occurring in the kidney.
Crystals: Numerous kinds of crystals can be observed in the urine. They may be composed of cystine, uric acid, calcium oxalate, or phosphate. They may not be initially observable, but as the urine cools, crystals will form. Crystals tend to form in a concentrated acidic or alkaline urine. Generally they are not clinically significant. Crystal formation is diagnostically significant, usually indicating inflammation, infection, or a metabolic disorder.
White Blood Cells: White blood cells (WBCs) in the urine (a condition termed pyuria) are indicative of urinary tract infection, particularly when bacteria are present. Glomerulonephritis and nephrotic syndrome also may demonstrate pyuria but usually in combination with proteinuria, red cells, and casts. The finding of WBC casts reflects a kidney infection because these casts are not formed in the bladder or prostate. If WBCs are present in the urine, a culture should be done for specific identification of bacteria and sensitivity of bacteria to antibiotics.
Other Measures
Dipsticks and reagent strips are available for detecting other substances in the urine, including glucose, bilirubin, urobilinogen, leukocyte esterase and nitrates, ketones, proteins, hemoglobin, and myoglobin.2
Aging and Renal Function
Throughout life the kidney responds to an increased workload by compensatory hypertrophy. This hypertrophy is marked in individuals who have donated a kidney for transplant or have lost functioning nephrons from trauma or disease. The glomeruli increase in diameter, and the tubules enlarge effectively to maintain the regulatory functions of the kidney. Hypertrophy occurs more rapidly and with a larger size increase in younger individuals and in those with high protein intake.
Changes in the kidneys occur throughout life, with decrease in size and a linear decrease in renal blood flow and GFR.22,23 With aging the number of nephrons decreases and may be related to oxidative stress and inflammation.24 The primary mechanism appears to be a change in the renal vasculature and perfusion pattern, which leads to a reduction in numbers of nephrons. The rate of nephron loss accelerates between 40 and 80 years of age. By 75 years of age the nephron population is reduced by 30% to 50%, with loss of renal mass occurring primarily in the cortex.2 Degenerative changes within nephrons also occur with aging. The glomerular capillaries atrophy, with a reduction in the branching vessels. The glomeruli then may disappear completely. The arcuate and interlobular arteries become tortuous, contributing to ischemia. The loss of the glomerular tuft may cause a shunt between the afferent and efferent arterioles. Although loss of juxtaglomerular nephrons still allows the vasa recta to be perfused, the combination of events contributes to a decreasing ability to excrete a concentrated urine. Thus the specific gravity of the urine in older individuals tends to be on the low side of normal.
Tubular transport changes with aging, although under normal conditions the tubules function adequately. Adaptation to stressful conditions is more difficult. Glucose, bicarbonate, and sodium are not as efficiently reabsorbed, and hyperkalemia is more common because of decreased secretion. Response to acid or base loads is delayed and prolonged. Sudden or large changes in pH or fluid load may lead to serious imbalances with increased risk of hypervolemia or hypovolemia. Acute losses or chronic fluid deficits can lead to renal insufficiency in the older adult. Administration of drugs eliminated by renal processes may require dose modifications and more astute observations for toxic side effects.25 The Tm for glucose reabsorption decreases with age, contributing to a greater amount of glucose in the urine. This is an important consideration when glycosuria is used for screening or monitoring the process of diabetes mellitus in older adults. These changes occur independently of disease, however, indicating a normal process of aging. An age-related decline in renal activation of vitamin D decreases intestinal absorption of calcium, and older adults need more vitamin D to overcome diminishing renal function.26 Previous or concurrent renal disease or urinary tract obstruction may amplify age-related changes in function.
Bladder symptoms are common among older adults and include frequency, urgency, and nocturia. Neurogenic and myogenic changes in bladder structure and function may contribute to some symptoms as well as influences outside the urinary tract. Changes in neurotransmission influence the micturition reflex and may lead to overactive bladder.27 Obstruction related to prostate hypertrophy may lead to urine retention with frequency, urgency, nocturia, and slow or intermittent urinary stream.
Angiotensin II 1352
Angiotensin converting enzyme (ACE) 1352
Antidiuretic hormone (ADH) 1358
Arcuate arteries 1348
Atrial natriuretic peptide (ANP) 1352
Autoregulation 1351
Bladder 1350
Blood urea nitrogen (BUN) 1361
Bowman capsule 1345
Bowman space 1345
Brain natriuretic peptide (BNP) 1352
Calcitriol 1359
Cast 1361
Collecting duct 1347
Countercurrent exchange system 1357
Creatinine clearance 1360
Crystal 1361
Cystatin C 1361
C-type natriuretic peptide 1352
Detrusor muscle 1350
Distal tubule 1347
Diuretic 1358
Effective renal plasma flow (ERPF) 1360
Effective renal blood flow (ERBF) 1360
Excretion 1352
External urethral sphincter 1350
Filtration slit 1346
Glomerular capillary 1348
Glomerular endothelium 1345
Glomerular filtration membrane 1345
Glomerular filtration rate (GFR) 1351
Glomerulus 1345
Hematuria 1361
Hilum 1345
Intercalated cell 1347
Interlobar artery 1347
Interlobular arteries 1348
Internal urethral sphincter 1350
Inulin clearance 1360
Juxtaglomerular apparatus (JGA) 1347
Juxtaglomerular cell 1346
Juxtamedullary nephron, 1345 1346
Kidney 1344
Loop of Henle 1347
Macula densa 1346
Major calyx 1345
Mesangial cell 1345
Mesangial matrix 1345
Micturition 1350
Midcortical nephron 1345
Minor calyx 1345
Myogenic mechanism 1351
Natriuretic peptide 1352
Nephron 1345
Net filtration pressure (NFP) 1354
Oliguria 1358
Parietal epithelium 1346
Peritubular capillary 1348
Plasma creatinine (PCR) concentration 1360
Podocyte 1345
Principal cell 1347
Proximal tubule 1347
Pyuria 1362
Renal artery 1347
Renalase 1358
Renal capsule 1344
Renal column 1345
Renal corpuscle 1345
Renal cortex 1345
Renal fascia 1344
Renal medulla 1345
Renal papillae 1348
Renal pelvis 1345
Renal pyramid 1345
Renin-angiotensin-aldosterone system 1352
Specific gravity 1361
Superficial cortical nephron 1345
Transport maximum (Tm) 1355
Trigone 1348
Tubular reabsorption 1352
Tubular secretion 1352
Tubuloglomerular feedback 1351
Ultrafiltration 1352
Urea 1358
Ureter 1348
Urethra 1350
Urinalysis 1361
Urine color 1361
Urine pH 1361
Urodilantin 1352
Vasa recta 1350
Vitamin D 1358
Visceral epithelium 1346
Water diuresis 1358
REFERENCES
1. Vaughan, M.R., Quaggin, S.E. How do mesangial and endothelial cells form the glomerular tuft? J Am Soc Nephrol. 2008;19(1):24–33.
2. Brenner, B.M. Brenner and Rector’s the kidney, ed 8. Philadelphia: Saunders; 2008.
3. Faul C et al: Actin up: regulation of podocyte structure and function by components of the actin cytoskeleton, Trends Cell Biol 17(9):438-437.
4. Marshall, S.M. The podocyte: a potential therapeutic target in diabetic nephropathy? Curr Pharm Des. 2007;13(26):2713–2720.
5. Wein, A.J. Role of the urothelium in bladder function. J Urol. 2005;173(6):2199–2200.
6. Persson, P.B. Renal blood flow autoregulation in blood pressure control. Curr Opin Nephrol Hypertens. 2002;11(1):67–72.
7. Loutzenhiser, R., et al. Renal autoregulation: new perspectives regarding the protective and regulatory roles of the underlying mechanisms. Am J Physiol Regul Integr Comp Physiol. 2006;290(5):R1153–R1167.
8. Castrop, H. Mediators of tubuloglomerular feedback regulation of glomerular filtration: ATP and adenosine. Acta Physiol (Oxf). 2007;189(1):3–14.
9. Schweda, et al. Renin release, Physiology (Bethesda). 2007;22:310–319. [review].
10. Yuan, et al. 1,25-dihydroxyvitamin D3 suppresses renin gene transcription by blocking the activity of the cyclic AMP response element in the renin gene promoter. J Biol Chem. 2007;282(41):29821–29830.
11. Martinez-Rumayor, A., et al. Biology of the natriuretic peptides. Am J Cardiol. 2008;101(3A):3–8.
12. Cea, L.B. Natriuretic peptide family: new aspects. Curr Med Chem Cardiovasc Hematol Agents. 2005;3(2):87–98.
13. Zasloff, M. Antimicrobial peptides, innate immunity, and the normally sterile urinary tract. J Am Soc Nephrol. 2007;18(11):2810–2816.
14. Goodfriend, T.L. Aldosterone—a hormone of cardiovascular adaptation and maladaptation. J Clin Hypertens (Greenwich). 2006;8(2):133–139.
15. Koeppen, B.M., Stanton, B.A. Renal physiology. St Louis: Mosby; 2007. [pp 81-83,].
16. Li, G., et al. Catecholamines regulate the activity, secretion, and synthesis of renalase. Circulation. 2008;117(10):1277–1282.
17. Lips, R. Vitamin D physiology. Prog Biophys Mol Biol. 2006;92(1):4–8.
18. Johnson, D.W., Forman, C., Vesey, D.A. Novel renoprotective actions of erythropoietin: new uses for an old hormone. Nephrology (Carlton). 2006;11(4):306–312.
19. Israni, A.K., Kasiske, B.L. Laboratory assessment of kidney disease: clearance, urinalysis, and kidney biopsy. In Brenner B.M., ed.: Brenner & Rector’s the kidney, ed 8, Philadelphia: Saunders, 2008.
20. Zaffanello, M., Franchini, M., Fanos, V. Is serum cystatin-C a suitable marker of renal function in children? Ann Clin Lab Sci. 2007;37(3):233–240.
21. Prigent, A. Monitoring renal function and limitations of renal function tests. Semin Nucl Med. 2008;38(1):32–46.
22. Fehrman-Ekholm, I., Skeppholm, L. Renal function in the elderly (>70 years old) measured by means of iohexol clearance, serum creatinine, serum urea and estimated clearance. Scand J Urol Nephrol. 2004;38(1):73–77.
23. Martin, J.E., Sheaff, M.T. Renal aging. J Pathol. 2007;211(2):198–205.
24. Csiszar, A., et al. The aging kidney: role of endothelial oxidative stress and inflammation. Acta Physiol Hung. 2007;94(1-2):107–115.
25. El Desoky, E.S. Pharmacokinetic-pharmacodynamic crisis in the elderly. Am J Ther. 2007;14(5):488–498.
26. Vieth, R., Ladak, Y., Walfish, P.G. Age-related changes in the 25-hydroxyvitamin D versus parathyroid hormone relationship suggest a different reason why older adults require more vitamin D. J Clin Endocrinol Metab. 2003;88(1):185–191.
27. Chu, F.M., Dmochowski, R. Pathophysiology of overactive bladder. Am J Med. 2006;119(3 Suppl 1):3–8.