CELLULAR BIOLOGY
 Evolve Website (
Evolve Website (
All body functions depend on the integrity of cells. There-fore, an understanding of cellular biology is intrinsically necessary for an understanding of disease. An overwhelming amount of information is revealing how cells behave as a multicellular “social” organism. At the heart of cellular biology is cellular communication (“cellular crosstalk”)—how messages originate and are transmitted, received, interpreted, and used by the cell. Fossil records suggest that unicellular organisms resembling bacteria were present on earth 3.5 billion years ago, yet it took another 2.5 billion years for the first multicellular organisms to appear. This delay was seemingly slow because elaborate signaling mechanisms had to evolve that would allow cells to crosstalk. This streamlined conversation between, among, and within cells maintains cellular function. Intercellular signals allow each cell to determine its position and specialized role. Cells must demonstrate a “chemical fondness” for other cells and their surrounding environment to maintain the integrity of the entire organism. When they no longer tolerate this fondness, the conversation breaks down and cells either adapt (sometimes altering function) or become vulnerable to isolation, injury, or disease.
PROKARYOTES AND EUKARYOTES
Living cells generally are divided into two major classes—eukaryotes and prokaryotes. The cells of higher animals and plants are eukaryotes, as are the single-celled organisms fungi, protozoa, and most algae. Prokaryotes include cyanobacteria (blue-green algae), bacteria, and rickettsiae. Prokaryotes traditionally were studied as core subjects of molecular biology. Current emphasis is on the eukaryotic cell; much of its structure and function has no counterpart in bacterial cells.
Eukaryotes (eu = good; karyon = nucleus) are larger and have more extensive intracellular anatomy and organization than do prokaryotes. Eukaryotic cells have a characteristic set of membrane-bound intracellular compartments, called organelles, that includes a well-defined nucleus. Prokaryotes contain no organelles, and their nuclear material is not encased by a nuclear membrane. Prokaryotic cells are characterized by lack of a distinct nucleus.
In addition to having structural differences, prokaryotic and eukaryotic cells differ in chemical composition and biochemical activity. The nuclei of prokaryotic cells carry genetic information in a single circular chromosome, and they lack a class of proteins called histones, which in eukaryotic cells bind with deoxyribonucleic acid (DNA) and are involved in the supercoiling of DNA (see Figure 1-2). Eukaryotic cells have several chromosomes. Protein production, or synthesis, in the two classes of cells also differs because of major structural differences in ribonucleic acid (RNA) protein complexes. Other distinctions include differences in mechanisms of transport across the outer cellular membrane and differences in enzyme content.
CELLULAR FUNCTIONS
Cells become specialized through the process of differentiation, or maturation, so that some cells eventually perform one kind of function and other cells perform other functions. Highly developed functions, such as movement, are often associated with the absence of some other property, such as hormone production, which is more highly developed in some other type of specialized cell. The eight chief cellular functions follow:
1. Movement. Muscle cells can generate forces that produce motion. Muscles that are attached to bones produce limb movements, whereas those that enclose hollow tubes or cavities move or empty contents when they contract. For example, the contraction of smooth muscle cells surrounding blood vessels changes the diameter of the vessels; the contraction of muscles in walls of the urinary bladder expels urine.
2. Conductivity. Conduction as a response to a stimulus is manifested by a wave of excitation, an electrical potential, that passes along the surface of the cell to reach its other parts. Conductivity is the chief function of nerve cells.
3. Metabolic absorption. All cells take in and use nutrients and other substances from their surroundings. Cells of the intestine and the kidney are specialized to carry out absorption. Cells of the kidney tubules reabsorb fluids and synthesize proteins. Intestinal epithelial cells reabsorb fluids and synthesize protein enzymes.
4. Secretion. Certain cells, such as mucous gland cells, can synthesize new substances from substances they absorb and then secrete the new substances to serve as needed elsewhere. Cells of the adrenal gland, testis, and ovary can secrete hormonal steroids.
5. Excretion. All cells can rid themselves of waste products resulting from the metabolic breakdown of nutrients. Membrane-bound sacs (lysosomes) within cells contain enzymes that break down, or digest, large molecules, turning them into waste products that are released from the cell.
6. Respiration. Cells absorb oxygen, which is used to transform nutrients into energy in the form of adenosine triphosphate (ATP). Cellular respiration, or oxidation, occurs in organelles called mitochondria.
7. Reproduction. Tissue growth occurs as cells enlarge and reproduce themselves. Even without growth, tissue maintenance requires that new cells be produced to replace cells that are lost normally through cellular death. Not all cells are capable of continuous division (see Chapter 2).
8. Communication. Communication is critical for all the other functions above that enable the survival of the society of cells. Pancreatic cells, for instance, secrete and release insulin to tell muscle cells to take up sugar from the blood for energy. Constant communication allows the maintenance of a dynamic steady state.
STRUCTURE AND FUNCTION OF CELLULAR COMPONENTS
Figure 1-1 shows a “typical” eukaryotic cell. It consists of three components: an outer membrane called the plasma membrane, or plasmalemma; a fluid filling called cytoplasm; and the “organs” of the cell-membrane–bound intracellular organelles, among them the nucleus.
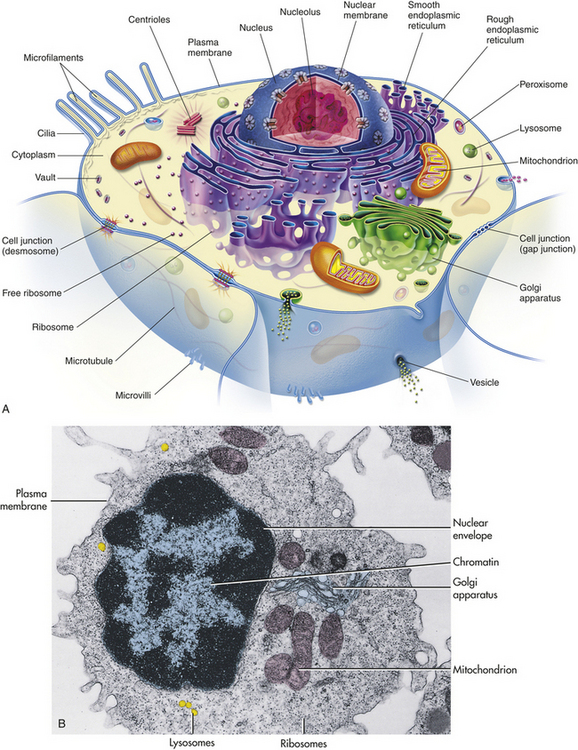
Figure 1-1 Typical or composite cell. A, Artist’s interpretation of cell structure. B, Color-enhanced electron micrograph of a cell. Both show the many mitochondria known as the “power plants of the cell.” Note, too, the innumerable dots bordering the endoplasmic reticulum. These are ribosomes, the cell’s “protein factories.” (B from Thibodeau GA, Patton KT: Anatomy & physiology, ed 5, St Louis, 2003, Mosby.)
Nucleus
The nucleus, which is surrounded by the cytoplasm and generally is located in the center of the cell, is the largest membrane-bound organelle. Two membranes comprise the nuclear envelope (Figure 1-2, A). The outer membrane is continuous with membranes of the endoplasmic reticulum. The nucleus contains the nucleolus, a small dense structure composed largely of RNA; most of the cellular DNA; and the DNA-binding proteins, the histones, that regulate its activity. The DNA chain in eukaryotic cells is so extensive that the risk of breakage is high. Therefore, the histones that bind to DNA cause the folding of DNA into chromosomes (Figure 1-2, C). The wrapping of DNA into tight packages of chromosomes is essential for cell division in eukaryotes.
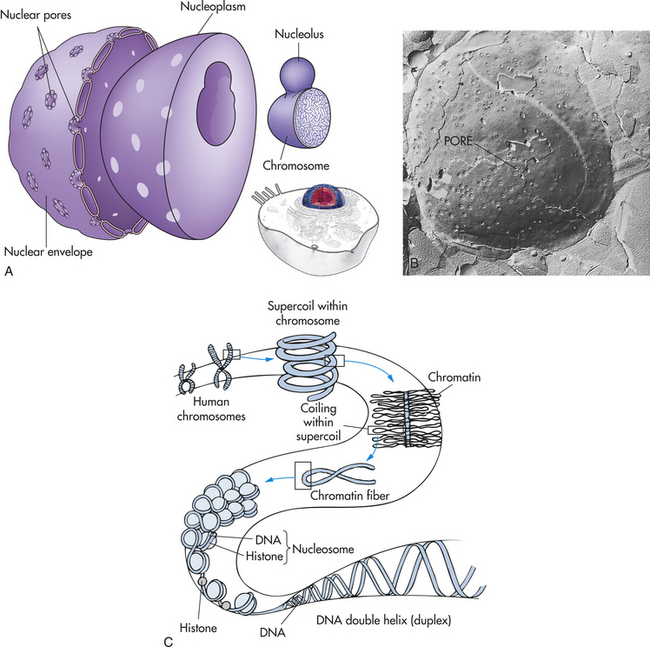
Figure 1-2 The nucleus. The nucleus is composed of a double membrane, called a nuclear envelope, that encloses the fluid-filled interior, called nucleoplasm. The chromosomes are suspended in the nucleoplasm (shown here much larger than real size to show the tightly packed DNA strands). A, Swelling at one or more points of the chromosome occurs at a nucleolus, where genes are being copied into RNA. The nuclear envelope is studded with pores. B, The pores are visible as dimples in this freeze etch of a nuclear envelope. C, How DNA is coiled within a chromosome. (B from Raven PH, Johnson GB: Biology, St Louis, 1992, Mosby.)
The primary functions of the nucleus are cell division and control of genetic information. Other functions include the replication and repair of DNA and the transcription of the information stored in DNA. Genetic information is transcribed into RNA, which can be processed into messenger, transport, and ribosomal RNA and introduced into the cytoplasm, where it directs cellular activities. Most of the processing of RNA occurs in the nucleolus. (The role of DNA and RNA in protein synthesis is discussed in Chapter 4.)
Cytoplasmic Organelles
Cytoplasm is an aqueous solution (cytosol) that fills the cytoplasmic matrix—the space between the nuclear envelope and the plasma membrane. The cytosol represents about half the volume of a eukaryotic cell. It contains thousands of enzymes involved in intermediate metabolism and is crowded with ribosomes making proteins. Newly synthesized proteins remain in the cytosol if they lack a signal for transport to a cell organelle.1 The organelles suspended in the cytoplasm are enclosed in biologic membranes, which enables them simultaneously to carry out functions that require different biochemical environments. These functions, many of which are directed by coded messages carried from the nucleus by RNA, include synthesis of proteins and hormones and their transport out of the cell, isolation and elimination of waste products from the cell, metabolic processes, breakdown and disposal of cellular debris and foreign proteins (antigens), and maintenance of cellular structure and motility. Also the cytosol functions as a storage unit for fat, carbohydrate, and secretory vesicles.
Ribosomes
Ribosomes are RNA-protein complexes (nucleoproteins) that are synthesized in the nucleolus and secreted into the cytoplasm, possibly through pores in the nuclear envelope. These tiny organelles may float free in the cytoplasm or attach themselves to the outer membranes of the endoplasmic reticulum (see Figure 1-1). Their chief function is to provide sites for cellular protein synthesis. Newly formed ribosomes synthesize a “recognition sequence,” or signal, like an address on a letter. Signal recognition particles (SRPs) in the cytosol bind to the ribosome after recognizing the SRP. Ribophorins, receiver proteins found on the rough sections of the endoplasmic reticulum (ER), act as the “address” site or binding sites. The developing protein threads its way through the ER membrane into the lumen. The SRP is removed and the new protein chain is folded into its final conformation.
Endoplasmic Reticulum
The endoplasmic reticulum (endo = within; plasma = cytoplasm; reticulum = network) is a membrane factory that specializes in the synthesis and transport of the protein and lipid components of most of the cell’s organelles. It consists of a network of tubular or saclike channels (cisternae) that extend throughout the cytoplasm and are continuous with the outer nuclear membrane (Figure 1-3). The folded membranes that form the cisternae of the endoplasmic reticulum may be rough (granular) or smooth (agranular). The rough endoplasmic reticulum is rough because ribosomes and ribonucleoprotein particles are attached to it (see Figure 1-3). Some of the proteins synthesized by these ribosomes remain in the endoplasmic reticulum, and others are used to construct membranes of other organelles (the Golgi complex, lysosomes, peroxisomes, nucleus) and of the cell itself.
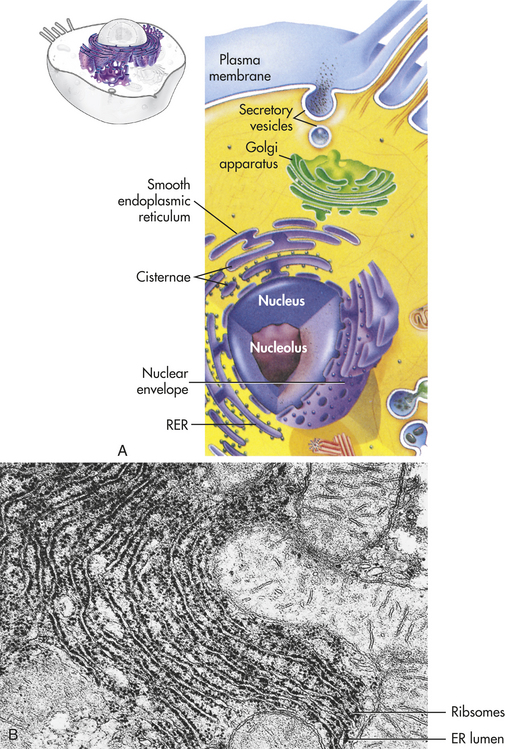
Figure 1-3 Endoplasmic reticulum (ER). A, The ER consists of rough endoplasmic reticulum (RER) arranged into ribosome-coated cisternae and vesicles of smooth endoplasmic reticulum (SER). B, Electron micrograph of rough and smooth ER. (B courtesy Kelloes C and Farmer M, Center for Advanced Ultrastructural Research, University of Georgia. From Lindsay DT: Functional human anatomy, St Louis, 1996, Mosby.)
Smooth endoplasmic reticulum does not contain ribosomes or ribonucleoprotein particles (see Figure 1-1). Rather, membranous surfaces of the smooth endoplasmic reticulum contain enzymes involved in the synthesis of steroid hormones and are responsible for a variety of reactions required to remove toxic substances from the cell. The endoplasmic reticulum communicates with the Golgi complex and interacts with other organelles, particularly lysosomes and peroxisomes.
Golgi Complex
The Golgi complex (or Golgi apparatus) is a network of flattened, smooth membranes and vesicles frequently located near the nucleus of the cell (Figure 1-4). Proteins from the endoplasmic reticulum are processed and packaged into small membrane-bound sacs or vesicles called secretory vesicles, which collect at the end of the membranous folds of the Golgi bodies—called cisternae. The secretory vesicles then break off from the Golgi complex and migrate to a variety of intracellular and extracellular destinations, including the plasma membrane. The vesicles fuse with the plasma membrane, and their contents are released from the cell. The best known vesicles are those that have coats made largely of the protein clathrin and are called clathrin-coated vesicles. They bud from the Golgi complex on the outward secretory pathway and from the plasma membrane on the inward endocytotic pathway (see p. 30). Many molecules, including lipids, proteins, glycoproteins, and enzymes of lysosomes, pass through the Golgi complex at some stage in their maturation. The Golgi complex is a refining plant and directs traffic (e.g., protein, polynucleotide, polysaccharide molecules) in the cell1 (Figure 1-5).
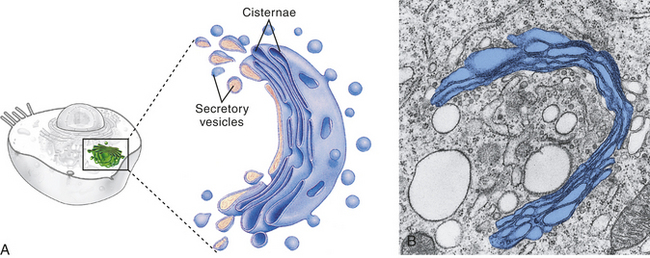
Figure 1-4 Golgi complex. A, Schematic representation of the Golgi complex showing a stack of flattened sacs, or cisternae, and numerous small membranous bubbles, or secretory vesicles. B, Transmission electron micrograph showing the Golgi complex highlighted with color. (From Thibodeau GA, Patton KT: Anatomy & physiology, ed 6, St Louis, 2007, Mosby.)
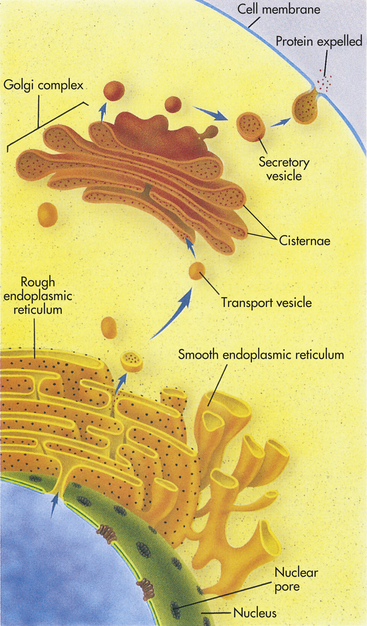
Figure 1-5 How the internal membrane system of a cell packages a protein for export. The instructions for making a protein that is destined for export from a cell, such as a digestive enzyme made by a pancreas cell, are first transcribed from DNA by RNA in the nucleus. The RNA then leaves the nucleus through a nuclear pore and proceeds to a ribosome located on the rough endoplasmic reticulum (ER). There it provides instructions for the correct sequence of amino acids for synthesizing that particular digestive enzyme. When enzyme synthesis is complete, the enzyme travels through the ER and is then encapsulated in a transport vesicle. The transport vesicle fuses with a Golgi body, releasing the enzyme. In the Golgi complex the enzyme is further modified and is then shunted to the ends of the Golgi complex, or cisternae. There the enzyme waits for a secretory vesicle, which will carry it to the perimeter of the cell, the cell membrane. The secretory vesicle membrane then fuses with the cell membrane, and the enzyme is released outside the cell. (From Raven PH, Johnson GB: Understanding biology, ed 3, Dubuque, IA, 1995, Brown.)
Lysosomes
Lysosomes (lyso = dissolution; soma = body) are saclike structures that originate from the Golgi complex (see Figure 1-1). They contain more than 40 digestive enzymes called hydrolases, which catalyze bonds in proteins, lipids, nucleic acids, and carbohydrates. Lysosomes function as the intracellular digestive system (Figure 1-6). Lysosomal enzymes are capable of digesting most cellular constituents down to their basic forms, such as amino acids, fatty acids, and sugars.
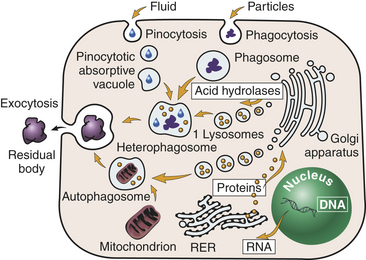
Figure 1-6 Lysosomes. Primary (1) lysosomes, which originate from the Golgi apparatus, give rise to heterophagosomes and autophagosomes. Undigested material in phagosomes is extruded from the cell or remains in the cytoplasm as lipofuscin-rich residual bodies. RER, Rough endoplasmic reticulum. (From Damjanov I: Pathology for the health-related professionals, ed 3, Philadelphia, 2006, Saunders.)
The lysosomal membrane acts as a protective shield between the powerful digestive enzymes within the lysosome and the cytoplasm, preventing their leakage into the cytoplasmic matrix. Disruption of the membrane by various treatments or cellular injury leads to a release of the lysosomal enzymes, which can then react with their specific substrates, causing cellular self-digestion. Lysosomal abnormalities are involved in a number of conditions that involve cellular injury and death.
Lysosomal storage diseases may be the result of a genetic defect or lack of one or more lysosomal enzymes. For example, the lack of lysosomal α-1,4-glucosidase leads to an accumulation of glycogen in lysosomes known as Pompe disease. Tay-Sachs disease is characterized by an accumulation of GM2 ganglioside (a lipid) in lysosomes as a result of the deficiency or absence of lysosomal hexosaminidase A. In gout, undigested uric acid accumulates within lysosomes, damaging the lysosomal membrane. Subsequent enzyme leakage results in cell death and tissue injury.
Lysosomes are necessary for normal digestion of cellular nutrients, intracellular debris, and potentially harmful extracellular substances that must be removed from the body. Extracellular substances are taken into the cell and encapsulated in a membrane-bound vesicle (see discussion on endocytosis, p. 30). Lysosomes merge with the vesicle to form a digestive vacuole. Lysosomes remain fully active by maintaining a low internal pH. They do this by pumping hydrogen ions into their interiors. The hydrolytic enzymes are only maximally active at acid pH values. Lysosomes that are not active do not maintain such an acid internal pH. Lysosomes in this “holding pattern” are called primary lysosomes. When a primary lysosome fuses with a vacuole or other organelle, its pH falls and the hydrolytic enzymes become activated. When it becomes active, it is called a secondary lysosome, or heterophagosome.
As cells complete their life span and die, lysosomes digest the resultant cellular debris. Lysosomes involved in this process, which is called autodigestion, are called autolysosomes, or autophagosomes. In living cells, cellular debris is encapsulated within a vesicle that reacts with a lysosome to complete its degradation. This process is called autophagy. Autophagy also occurs during starvation, enabling the cell to use a part of its own substance for fuel without doing itself irreparable harm.
Products of autophagy (and of phagocytosis, the ingestion of harmful foreign substances; see Chapter 6) pass out of the lysosome and are reused by the cell. Indigestible material is stored in vesicles called residual bodies, whose contents are actively expelled from the cell (see Figure 1-6). High concentrations of lipids may accumulate within the residual bodies and remain there for a long time. The lipids are eventually oxidized, and a pigmented substance containing polyunsaturated fatty acids and proteins accumulates in the cell. This pigmented substance, termed lipofuscin, is often called “age pigment” or “age spots,” and is noted in older individuals (see Chapter 2).
Peroxisomes
Peroxisomes (microbodies) are similar to lysosomes in microscopic appearance, but they are larger and oval or irregular in shape. Peroxisomes contain several oxidative enzymes, such as catalase and urate oxidase. Like mitochondria, peroxisomes are major sites of oxygen utilization. Peroxisomes are so named because they usually contain enzymes that use oxygen to remove hydrogen atoms from specific substrates in an oxidative reaction that produces hydrogen peroxide (H2O2). Hydrogen peroxide is a powerful oxidant, potentially destructive if it accumulates or escapes from peroxisomes. Catalase, an antioxidant enzyme, uses the H2O2 to oxidize a variety of other substrates—phenols, formic acid, formaldehyde, and alcohol—by the peroxidative reaction:
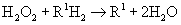
Thus the reaction of H2O2 breaks down to H2O and O2 (see discussion of free radicals in Chapter 2). Peroxisomes also have an important role in the synthesis of specialized phospholipids necessary for nerve cell myelination. Such reactions are important in detoxifying various wastes within the cell or foreign components that enter the cell, such as ethanol.
Mitochondria
Mitochondria (mito = thread; chondros = granule) are of much interest because of their role in cellular energy metabolism (see p. 21). These cytoplasmic organelles appear as spheres, rods, or filamentous bodies that are bound by a double membrane (Figure 1-7). The outer membrane is smooth and surrounds the mitochondrion itself; the inner membrane is convoluted in the mitochondrial matrix to form partitions called cristae. The inner membrane contains the enzymes of the respiratory chain—the name given to the electron transport chain. These enzymes are essential to the process of oxidative phosphorylation that generates most of the cell’s ATP. Metabolic pathways involved in the metabolism of carbohydrates, lipids, and amino acids and special pathways involving urea and heme synthesis are located in the mitochondrial matrix.
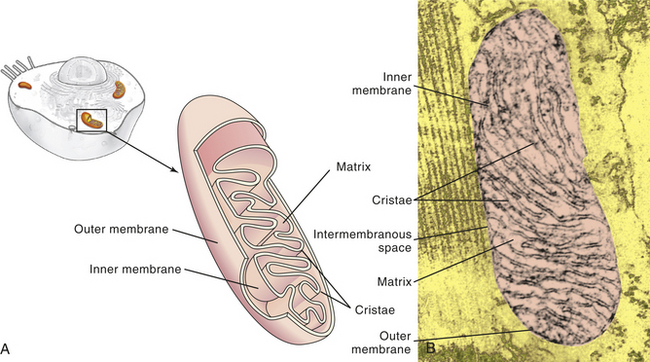
Figure 1-7 Mitochondrion. A, Cutaway sketch showing outer and inner membranes. Note the many folds (cristae) of the inner membrane. B, Transmission electron micrograph of a mitochondrion. Although some mitochondria have the capsule shape shown here, many are round or oval. (From Thibodeau GA, Patton KT: Anatomy & physiology, ed 6, St Louis, 2007, Mosby.)
The outer membrane is permeable (passable) to many substances, but the inner membrane is highly selective and contains many transmembranous transport systems. The inner membrane contains a transporter to move electrically charged calcium (calcium ions). (Membrane transport is discussed on p. 25.)
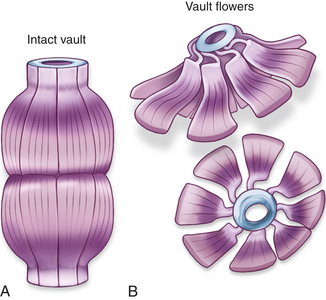
Vaults
Vaults are cytoplasmic ribonucleoproteins, much larger than ribosomes, and shaped like octagonal barrels (Figure 1-8). Their name comes from their multiple arches, which reminded their discoverers of vaulted or cathedral ceilings. A single cell can contain thousands of vaults. Vaults were identified only recently because of changes in staining techniques. The function of vaults may be related to their octagonal shape. Similarly, the pores in the membrane surrounding the nucleus (see Figure 1-2, B) are also octagonally shaped and the same size as vaults, leading to speculation that vaults may be cellular “trucks.” Further, vaults would dock at nuclear pores, pick up molecules synthesized in the nucleus, and deliver their load elsewhere in the cell. Because at any given time about 5% of the vaults are localized near the nuclear pores, it is thought that vaults may be carrying messenger RNA (mRNA) from the nucleus to the ribosomal sites of protein synthesis within the cytoplasm. Investigators suggest that vaults transport several copies of untranslated RNA and that they are transported along cytoskeletal-based cellular tracks—much like an assembly line.2 Researchers are investigating the role of vaults in cancer cells’ resistance to drug therapy. Perhaps transporting chemotherapy drugs to sites for exocytosis from the cancer cell increases the drugs’ elimination, or vaults may mediate multidrug resistance by transporting drugs away from their intracellular targets, for example, the nucleous.3 Although the normal cellular function of the vault is as yet undetermined, the structure of the vault is consistent with a role in either subcellular transport or sequestering large nuclear protein assemblies.4
Cytosol
Cytosol is the gelatinous, semiliquid portion of the cytoplasm accounting for about 55% of the total cell volume. Functions of the cytosol include intermediary metabolism involving enzymatic biochemical reactions; ribosomal protein synthesis; and storage of carbohydrates, fat, and secretory vesicles.
Intermediary metabolism refers to the intracellular chemical reactions that include synthesis, degradation, and transformation of small organic molecules (e.g., simple sugars, fatty acids, and amino acids). All intermediary metabolism occurs in the cytoplasm or that portion of the cell interior not occupied by the nucleus—with most of the metabolism being accomplished in the cytosol. These reactions enable energy to be used for cellular activities and for providing substrates to maintain cell integrity.
Ribosomal protein synthesis takes place in free ribosomes in the cytosol. Cytosolic ribosomes that synthesize identical proteins are collected together in “factories” known as polyribosomes.
Storage of excess nutrients not immediately used for ATP production is converted in the cytosol into storage forms; for example, excess glucose is stored as glycogen. These temporary masses are known as inclusions (see Chapter 2). Secretory vesicles that have been processed and packaged by the endoplasmic reticulum and Golgi complex also remain in the cytosol. By means of signaling, the vesicles transport and empty their contents to the outside.
Cytoskeleton
All eukaryotic cells contain elaborate and specialized internal structures in the cytosol that provide the “bones and muscles” of the cell—the cytoskeleton. The cytoskeleton maintains the cell’s shape and internal organization, and it permits movement of substances within the cell and movement of external projections (cilia or microvilli; flagella in sperm) outside the plasma membrane. The internal skeleton is composed of a network of protein filaments; two of the most important are microtubules and actin filaments, or microfilaments.
Microtubules are small, hollow, cylindric, unbranched tubules made of protein. When found together, microtubules exhibit rigidity, unlike the rest of the cytoplasm. Microtubules thus add strength to the cell’s structure (Figure 1-9, A). Within the cell, microtubules support and move organelles from one part of the cytoplasm to another, facilitate transport of impulses along nerve cells, and have roles in the inflammatory and immune responses and hormone secretion. Microtubules are also involved in external movement, or motility, of some cells.
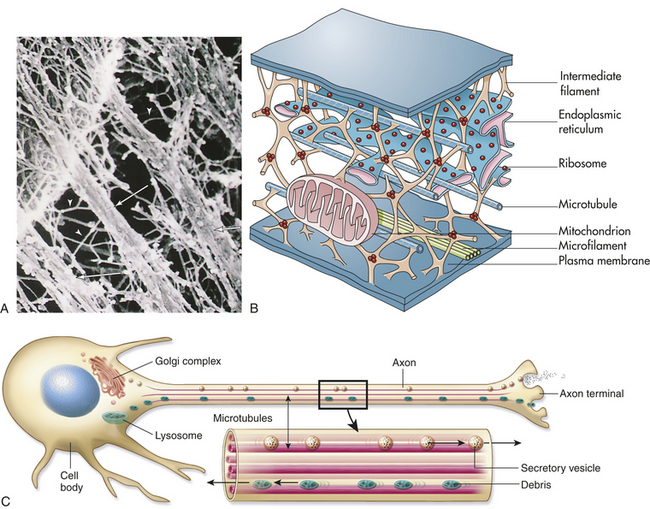
Figure 1-9 Cytoskeleton. A, Color-enhanced electron micrograph of a portion of the cell’s internal framework. Arrowheads mark the intermediate filaments, and the complete arrows mark the microtubules. B, Artist’s interpretation of the cell’s internal framework. Note that the “free” ribosomes and other organelles are not really free at all. C, Microtubules are necessary for maintaining an asymmetric cell shape, such as that of a nerve cell. In addition, specific chemicals are released from the terminal end of the axon to influence neural transmission. (A and B from Thibodeau GA, Patton KT: Anatomy & physiology, ed 6, St Louis, 2007, Mosby.)
Microtubules are arranged in the thickened base, or basal body, of a protrusion from the cell’s plasma membrane. This arrangement occurs in the basal bodies of sperm flagella and the cilia of certain other cells. The long, whiplike flagella enable sperm cells to move. Cilia usually move substances past the cell, which remains stationary. For example, cilia on cells lining the respiratory tract move together to “beat” mucus toward the throat so it can be removed by coughing.
While the cell is not in the process of division, only a few microtubules are assembled; cellular division (mitosis) or defense (phagocytosis) does, however, induce a cycle of rapid assembly and disassembly. Microtubules involved in cellular division are arranged in a centriole. Centrioles always consist of nine bundles containing three microtubules each. During division the pairs of centrioles split and migrate to opposite poles of the cell (see p. 34).
Alterations of microtubular function are implicated in disease processes. For example, alterations in actin microfilament act as a driving force for cell extension during cancer spread.5
Actin filaments (microfilaments) are smaller fibrils that generally occur in bundles rather than singly (Figure 1-9, C). Like microtubules, actin filaments are associated with cellular locomotion and maintenance of cell and tissue shape.5 In addition, microfilaments are necessary for regulating cell growth.6 Cellular locomotion depends on contractile properties that involve both microtubules and actin filaments. Anesthetic drugs can affect both structures, disrupting intracellular movement and cellular motility.
Plasma Membranes
Whether they surround the cell or enclose an intracellular organelle, membranes are exceedingly important to normal physiologic function because they control the composition of the space, or compartment, they enclose. Membranes can allow or exclude various molecules, and because of selective transport systems, they can move molecules into or out of the space (Figure 1-10). By controlling the movement of substances from one compartment to another, membranes exert a powerful influence on metabolic pathways. In addition to these functions, the plasma membrane has an important role in cell-to-cell recognition. For example, protein receptors for hormones and for other chemical signals are associated with the membrane and act as markers that identify a cell to its neighbors. Other functions of the plasma membrane include cellular mobility and the maintenance of cellular shape (Table 1-1).
Table 1-1

Modified from King DW, Fenoglio CM, Lefkowitch JH: General pathology: principles and dynamics, Philadelphia, 1983, Lea & Febiger.
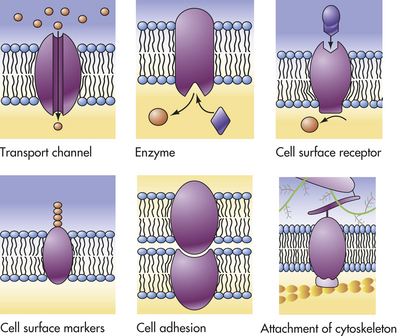
Figure 1-10 Functions of plasma membrane proteins. The plasma membrane proteins illustrated here show a variety of functions performed by the different types of plasma membranes. (From Raven PH, Johnson GB: Understanding biology, ed 3, Dubuque, IA, 1995, Brown.)
Membrane Composition
The outer surface of the plasma membrane is not smooth but dimpled with cavelike indentations known as caveolae (“tiny caves”). Caveolae were not thought to be functionally significant until the mid-1990s, when evidence suggested that they (1) serve as a repository for some receptors, (2) provide a new route for transport into the cell, and (3) act as the initiator for relaying signals from several extracellular chemical messengers into the cell’s interior7 (see p. 32).
The major chemical components of all membranes are lipids and proteins, but the percentage of each varies among different membranes. Lipid molecules are the most abundant, but the protein molecules are so large that in total mass these two constituents are roughly equal. The structure of a plasma membrane is shown in Figure 1-11. Intracellular membranes have a higher percentage of proteins than do plasma membranes, presumably because most enzymatic activity occurs within organelles. Carbohydrates are mainly associated with plasma membranes, where they are combined chemically with lipids, forming glycolipids, and with proteins, forming glycoproteins.
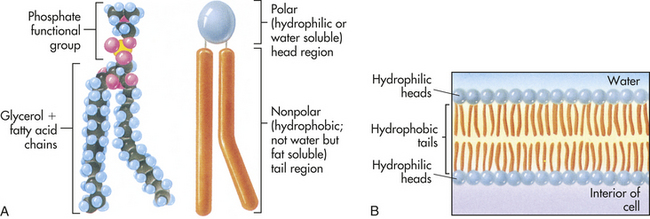
Figure 1-11 Structure of a phospholipid molecule. A, Each phospholipid molecule consists of a phosphate functional group and two fatty acid chains attached to a glycerol molecule. B, The fatty acid chains and glycerol form nonpolar, hydrophobic “tails,” and the phosphate functional group forms the polar, hydrophilic “head” of the phospholipid molecule. When placed in water, the hydrophobic tails of the molecule face inward, away from the water, and the hydrophilic head faces outward, toward the water. (From Raven PH, Johnson GB: Understanding biology, ed 3, Dubuque, IA, 1995, Brown.)
Lipids: The basic component of the plasma membrane is a bilayer of lipid molecules—phospholipids, glycolipids, and cholesterol (respective ratios 70:5:25). The lipids are responsible for the structural integrity of the membrane. Each lipid molecule is said to be polar, or amphipathic. An amphipathic molecule is one in which one part is hydrophobic (uncharged, or “water hating”) and another part is hydrophilic (charged, or “water loving”) (see Figure 1-11). The membrane spontaneously organizes itself into a bilayer because of these two incompatible solubilities. The hydrophobic region (hydrophobic tail) of each lipid molecule is protected from water, whereas the hydrophilic region (hydrophilic head) is immersed in it. The bilayer’s structure accounts for one of the essential functions of the plasma membrane: it is impermeable to most water-soluble molecules (molecules that dissolve in water) because they are insoluble in the oily core region. The bilayer serves as a barrier to the diffusion of water and hydrophilic substances while allowing lipid-soluble molecules, such as oxygen (O2) and carbon dioxide (CO2), to diffuse through it readily. Because the bilayer is fluid at temperatures above freezing, components of the cellular environment move slowly and selectively across the membrane all the time. (Components of the cellular environment are also discussed in Chapter 3.)
Proteins: Research suggests two ways to classify membrane proteins. One way is classification as peripheral or integral proteins. Integral membrane proteins are those embedded in the lipid bilayer linked to either phosphatidylinositol, a minor phospholipid, or a fatty acid chain. The integral proteins can be removed from the membrane only by detergents that solubilize (dissolve) the liquid. Peripheral membrane proteins are not embedded in the bilayer but reside at one surface or the other, bound to an integral protein.
Although the classification of membrane proteins as peripheral or integral is commonly used, it does not describe how proteins are associated with the bilayer. The second mode of classification does so by taking into account the membrane-spanning, or transmembranous, nature of membrane proteins1 (see Figure 1-13). According to this classification, proteins are associated with the lipid bilayer in four ways:
1. Some proteins, called transmembrane proteins, extend across the bilayer and are exposed to an aqueous environment on both sides of it.
2. Some intracellular proteins extend their polypeptide chain partially through the bilayer by means of a fatty acid chain.
3. Some cell-surface proteins are attached to the bilayer by a covalent linkage (i.e., a specific oligosaccharide).
4. Some proteins do not extend even partially through the bilayer but are bound to the membrane by noncovalent linkages with other membrane proteins.
Proteins exist in densely folded molecular configurations rather than straight chains, so an excess of hydrophilic units is at the surface of the molecule and an excess of hydrophobic units is inside. Although membrane structure is determined by the lipid bilayer, membrane functions are determined largely by proteins. For example, proteins facilitate transport across membranes by serving as receptors, enzymes, or transporters. Proteins act as (1) recognition and binding units (receptors) for substances moving in and out of the cell; (2) pores or transport channels for various electrically charged particles called ions or electrolytes and specific carriers for amino acids and monosaccharides; (3) specific enzymes that drive active pumps that promote concentration of certain ions, particularly potassium (K+), within the cell while keeping concentrations of other ions, for example, sodium (Na+), below concentrations found in the extracellular environment; (4) cell surface markers, such as glycoproteins (proteins attached to carbohydrates) that identify a cell to its neighbor; (5) cell adhesion molecules (CAMs) or proteins that allow cells to hook together and form attachments to the cytoskeleton for maintaining cellular shape; and (6) catalysts of chemical reactions, for example, conversion of lactose to glucose (see Figure 1-10). (Membrane transport is discussed on p. 25.)
The interaction of plasma membrane proteins with lipids is complex and is currently the subject of much research. The role of proteins in the onset and progression of disease is important because of their enzymatic, transport, and recognition-receptor functions in cellular physiology.
Proteolytic Cascades: About 500 human genes encode proteases.8 Proteases are involved in the physiologic regulation of essential processes by participating in a tightly orchestrated sequence of events termed a proteolytic cascade. Four major proteolytic cascades with disease relevance are candidates for treatment modalities including (1) cell death or caspase-mediated apoptosis, (2) blood coagulation cascade, (3) degrading membrane enzymes or matrix metalloproteinase cascade, and (4) the complement cascade. Some proteases within a proteolytic cascade act as initiators, others are involved in amplification and propagation and execution (Figure 1-12). Understanding the various steps involved is crucial for designing drug interventions. Dysregulation of proteases features prominently in many human diseases, including cancer, autoimmunity, and neurodegenerative disorders.9–11
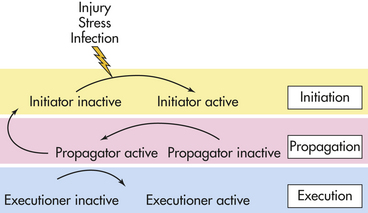
Figure 1-12 Schematic representation of a prototype proteolytic cascade. In the initiation phase, the cascade is triggered by an external stimulus, such as injury, stress, or infection. During the propagation phase, the initiator converts a downstream propagator into its active form by proteolysis. In the execution phase, the propagator will activate an executor. The process of coagulation is the best known proteolytic cascade. (Redrawn from Amour A et al: General considerations for proteolytic cascades, Biochem Soc Trans 32:15-16, 2004.)
Carbohydrates: A significant amount of carbohydrate is contained within the plasma membrane in the form of glycoprotein. Intercellular recognition, which is required for tissue formation, is an important function of membrane glycoproteins. Abnormal surface carbohydrate markers have been identified in certain tumor cells, leading investigators to claim that these markers are involved in tissue growth. Cells do not normally “trespass” their boundaries and overgrow their own territory.
Membrane Fluidity: The Fluid Mosaic Model
In the 1960s GL Nicholson and SJ Singer proposed the popular fluid mosaic model for biologic membranes (Figure 1-13). The model, which is continually being modified, presents integral proteins as pieces of a mosaic that float singly or as aggregates in the fluid lipid bilayer. The protein molecules serve to (1) transport other molecules into and out of the cell; (2) facilitate (catalyze) membrane reactions; (3) receive messages, thus acting as receptors for extracellular and intracellular signals; and (4) create structural linkages between the external and internal cellular environments. The fluid mosaic model accounts for the flexibility of cellular membranes, their self-sealing properties, and their impermeability to many substances.
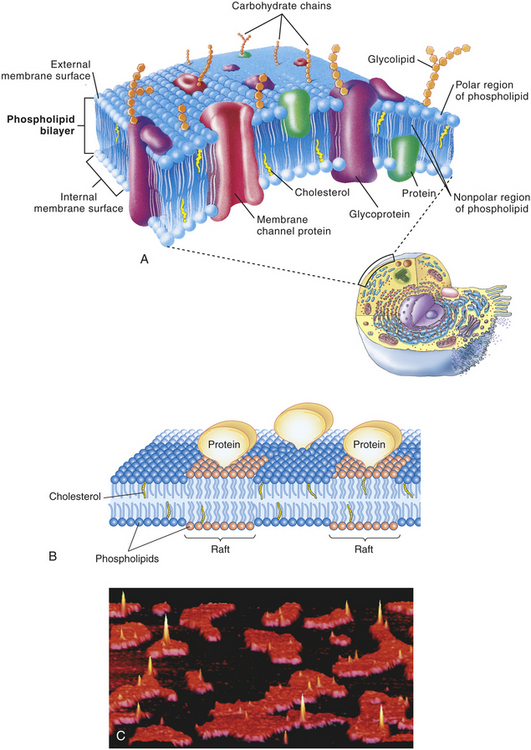
Figure 1-13 Fluid mosaic model and rafts. A, Schematic, three-dimensional view of the fluid mosaic model of membrane structure. The lipid bilayer provides the basic structure and serves as a relatively impermeable barrier to most water-soluble molecules. B, Diagram showing the basic structure of a membrane with rafts. The raft phospholipids have a richer supply of cholesterol than surrounding regions do and, along with attached proteins, form rather rigid floating platforms in the surface of the membrane. Rafts help organize functions at the surfaces of cells and organelles. C, Atomic force micrograph (AFM) in which an extremely fine-tipped needle drags over the surface of a cell membrane to reveal detailed surface features. Rafts are seen here as raised red-orange areas surrounded by black areas of less rigid phospholipid structure. (Modified from Thibodeau GA, Patton KT: Anatomy & physiology, ed 6, St Louis, 2007, Mosby.)
New revisions of the model now state that most membrane proteins do not enjoy unrestricted, lateral movement. Instead, multiple modes of diffusion and transport indicate a mix or heterogeneity in the membrane. Thus some proteins may randomly diffuse, others are confined or static, and still others are tethered to the cytoskeleton. The degree of a membrane’s fluidity depends on temperature. At lower temperatures the lipids are in a gel crystalline state, and at higher temperatures they become highly fluid. These properties are critical for cellular growth, division, and receptor function. Because some proteins are free to move within the plasma membranes (like floating icebergs), certain foreign proteins (antigens) may become buried in the bilayer, emerging at the surface only after injury and then attracting antibodies (proteins produced by the immune system), which attack host cells. Antigens and antibodies, which are the cause and effect of the immune response, are discussed in Chapter 7. The burial and reemergence of antigens may be one cause of autoimmune disease, described in Chapter 8.
In the fluid mosaic model, cellular membranes are dynamic. Not only do some lipids and proteins move laterally on the membrane, but also ions and other molecules move through it. Cells, however, do have ways of immobilizing specific membrane proteins in a specific region of the membrane. Confinement may be necessary for certain functions to occur, for example, formation of intercellular junctions by proteins. The fluid mosaic model is logical in that it describes the membrane as existing in a state of change and modulation, which allows the cell to protect itself actively against injurious agents. Hormones, bacteria, viruses, drugs, antibodies, chemicals that transmit nerve impulses (neurotransmitters), and other substances attach to the plasma membrane by means of receptor molecules on its outer layer. The number of receptors present may vary at different times, and the cell is capable of modulating the effects of injurious agents by altering receptor number and pattern.12 This aspect of the fluid mosaic model has drastically modified previously held concepts concerning the onset of disease.
The concentration of cholesterol in the plasma membrane affects membrane fluidity. Increased concentration results in less fluidity on the membrane’s hydrophilic outer surface and more fluidity at its hydrophobic core. Changes in cholesterol content are factors in some diseases. In cirrhosis of the liver, for example, the cholesterol content of the red blood cell’s plasma membrane increases. This causes an overall decrease in membrane fluidity that seriously affects the cell’s ability to transport oxygen.
Stiff groupings of membrane molecules, often cholesterol rich, form loglike rafts. Rafts are noted as raised groupings of membranes (see Figure 13, B) that help organize components of a membrane.
Cellular Receptors
Cellular receptors are protein molecules (proteins are discussed on p. 11) on the plasma membrane, in the cytoplasm, or in the nucleus that are capable of recognizing and binding with specific smaller molecules called ligands. Hormones, for example, are ligands. Recognition and binding depend on the chemical configuration of the receptor and its smaller ligand, which must fit together somewhat like pieces of a jigsaw puzzle (see Chapter 20). New data reveal that activation of a receptor also may depend on differences in movement and binding of the extracellular face of the receptor.13
Plasma membrane receptors are particularly important for cellular uptake of ligands (Table 1-2). They protrude from or are exposed at the external surface of the membrane and often are attached to integral proteins. Some of these recognition units have all the mobile properties related to membrane fluidity. The ligands that bind with membrane receptors include hormones, neurotransmitters, antigens, complement components, lipoproteins, infectious agents, drugs, and metabolites. The past several years have brought many new discoveries concerning the specific interactions of cellular receptors with their respective ligands. In many instances this information has provided a basis for understanding disease.
Table 1-2
Classes of Plasma Membrane Receptors
| Type of Receptor | Description |
| Channel linked | Also called ligand-gated channels; involve rapid synaptic signaling between electrically excitable cells. Channels open and close briefly in response to neurotransmitters changing ion permeability of plasma membrane of postsynaptic cell. |
| Catalytic | Once activated by ligands, function directly as enzymes. Composed of transmembrane proteins that function intracellularly as tyrosine-specific protein kinases. |
| G-protein linked | Indirectly activate or inactivate plasma membrane enzyme or ion channel; interaction mediated by guanosine triphosphate (GTP)–binding regulatory protein (G protein). When activated, a chain of reactions occurs that alters concentration of intracellular messengers, such as cyclic adenosine monophosphate (cAMP) and calcium, or signaling molecules. Other target proteins’ behavior also altered. May also interact with inositol phospholipids, which are significant in cell signaling, and molecules involved in the inositol-phospholipid transduction pathway. A G protein–linked receptor activates the enzyme phosphoinositide-specific phospholipase, which in turn generates two intracellular messengers: (1) inositol triphosphate (InP3) releases Ca++, and (2) diacylglycerol remains in the plasma membrane and activates protein kinase C. Protein kinase C further activates various cell proteins. Several different plasma membrane receptors are known to use the inositol-phospholipid transduction pathway. |
Data from Alberts B et al: Molecular biology of the cell, ed 4, New York, 2001, Garland.
Although the chemical nature of both ligands and the receptors to which they bind differs, receptors are classified on the basis of their location and function (see Cellular Communication and Signal Transduction, p. 18). Cellular type determines overall cellular function, but plasma membrane receptors determine which ligands a cell will bind with and how the cell will respond to binding with each. For example, the ability of a hormone or a neurotransmitter to stimulate a cell is regulated by the specificity and number of receptors present on the plasma membrane. Specific processes also control intracellular mechanisms. Hormone binding, for example, depends on special messenger molecules that regulate protein synthesis within the cell (see Chapter 20). Neurotransmitters (discussed in Chapter 14) also operate by causing special messengers to react with specific receptors.
Receptors for different drugs are found on the plasma membrane, in the cytoplasm, and in the nucleus. Membrane receptors have been found for certain anesthetics, opiates, endorphins, enkephalins, antibiotics, cancer chemotherapeutic agents, digitalis, and other drugs. Membrane receptors for endorphins, which are opiate-like peptides isolated from the pituitary gland, are found in large quantities in pain pathways of the nervous system (see Chapters 14 and 15). With binding, the endorphins (or drugs like morphine) change the cell’s permeability to ions, increase the concentration of molecules that regulate intracellular protein synthesis, and initiate molecular events that modulate pain perception.
Receptors for infectious microorganisms, or antigen receptors, bind bacteria, viruses, and parasites. Antigen receptors on white blood cells (lymphocytes, monocytes, macrophages, granulocytes) recognize and bind with antigenic microorganisms and activate the immune and inflammatory responses (see Chapters 6 and 7).
CELL-TO-CELL ADHESIONS
Cells are small and squishy, not at all like bricks. They are enclosed only by a flimsy membrane, yet the cell depends on the integrity of this membrane for its survival. How can cells be formed together strongly, with their membranes intact, to form a muscle that can lift this textbook? Plasma membranes not only serve as the outer boundaries of all cells but also allow groups of cells to be held together robustly, in cell-to-cell adhesions, to form tissues and organs. Once arranged, cells are held together by three different means: the extracellular matrix, cell adhesion molecules in the cell’s plasma membrane, and specialized cell junctions.
Extracellular Matrix
Cells can be bound together by attachment to one another or via the extracellular matrix (also including the basement membrane), which the cells secrete around themselves. The extracellular matrix is an intricate meshwork of fibrous proteins embedded in a watery, gel-like substance composed of complex carbohydrates (Figure 1-14). The matrix is like glue; however, it does provide a pathway for diffusion of nutrients, wastes, and other water-soluble traffic between the blood and tissue cells. Interwoven within the matrix are three groups of macromolecules: (1) fibrous structural proteins, including collagen and elastin; (2) a diverse group of adhesive glycoproteins, such as fibronectin; and (3) proteoglycans and hyaluronic acid.
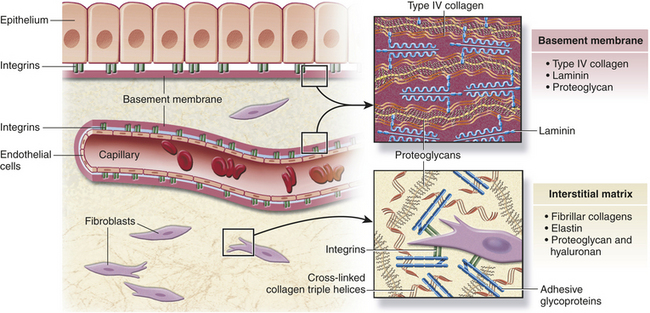
Figure 1-14 Extracellular matrix. Tissues are not just cells but also extracellular space. The extracellular space is an intricate network of macromolecules called the extracellular matrix (ECM). The macromolecules that constitute the ECM are secreted locally (by mostly fibroblasts) and assembled into a meshwork in close association with the surface of the cell that produced them. Two main classes of macromolecules include proteoglycans, which are bound to polysaccharide chains called glycosaminoglycans, and fibrous proteins (e.g., collagen, elastin, fibronectin, and laminin), which have structural and adhesive properties. Together the proteogylcan molecules form a gel-like ground substance in which the fibrous proteins are embedded. The gel permits rapid diffusion of nutrients, metabolites, and hormones between the blood and the tissue cells. Matrix proteins modulate cell-matrix interactions including normal tissue remodeling (which can become abnormal, for example, with chronic inflammation), embryogenesis, wound healing, and angiogenesis. Disruption of this balance results in serious diseases such as arthritis, tumor growth, and others. (Modified from Kumar V, Abbas A, Fausto N: Robbins and Cotran pathologic basis of disease, ed 7, Philadelphia, 2005, Saunders.)
Collagen forms cable-like fibers or sheets that provide tensile strength or resistance to longitudinal stress. Collagen breakdown, such as occurs in osteoarthritis, destroys the fibrils that give cartilage its tensile strength.
Elastin is a rubber-like protein fiber most abundant in tissue that must be capable of stretching and recoiling, such as the lungs.
Fibronectin, a large glycoprotein, promotes cell adhesion and cell anchorage. Reduced amounts have been found in certain types of cancerous cells; this allows cancer cells to travel or metastasize to other parts of the body.
All of these macromolecules occur in intercellular junctions and cell surfaces and may assemble into two different components: interstitial matrix and basement membrane (BM)14 (see Figure 1-14).
The extracellular matrix is secreted by fibroblasts (“fiber formers”), local cells that are present in the matrix. The matrix and the cells within it are known collectively as connective tissue because they connect cells together to form tissue and organs. Human connective tissues are enormously varied. They can be hard and dense, like bone; flexible, like tendons or the dermis of the skin; resilient and shock-absorbing, like cartilage; or soft and transparent, like the jelly that fills the eye. In all these examples, the majority of the tissue is composed of extracellular matrix, and the cells that produce the matrix are scattered within it like raisins in a pudding15 (see Figure 1-14).
The matrix is not just a passive scaffolding for cellular attachment; it also helps regulate the functions of the cells within which it interacts. The matrix helps regulate cell growth, movement, and differentiation.
Specialized Cell Junctions
Cells in direct physical contact with neighboring cells are often linked together at specialized regions of their plasma membranes called cell junctions. Cell junctions have two main functions: (1) to hold cells together and (2) to allow small molecules to pass from cell to cell, allowing coordination of the activities of cells that form tissues. The three main types of cell junctions are (1) desmosomes (adhering junctions, or macula adherens), (2) tight junctions (impermeable junctions, or zonula occludens), and (3) gap junctions (adhering [communicating] junctions) (Figure 1-15). Together they form the junctional complex. Desmosomes hold cells together by forming either continuous bands or belts of epithelial sheets or button-like points of contact. Desmosomes also act as a system of braces to maintain structural stability. Tight junctions serve as a barrier to diffusion, prevent the movement of substances through transport proteins in the plasma membrane, and prevent the leakage of small molecules between the plasma membranes of adjacent cells. Gap junctions are clusters of communicating tunnels, connexons, that allow small ions and molecules to pass directly from the inside of one cell to the inside of another. Connexons are joining proteins that extend outward from each of the adjacent plasma membranes. Cells connected by gap junctions are considered ionically (electrically) and metabolically coupled. Gap junctions coordinate the activities of adjacent cells. They are important, for example, in synchronizing contractions of heart muscle cells through ionic coupling and in permitting action potentials to spread rapidly from cell to cell in neural tissues. The reason that gap junctions occur in tissues that are not electrically active is unknown. Although most gap junctions are associated with junctional complexes, they sometimes exist as independent structures.
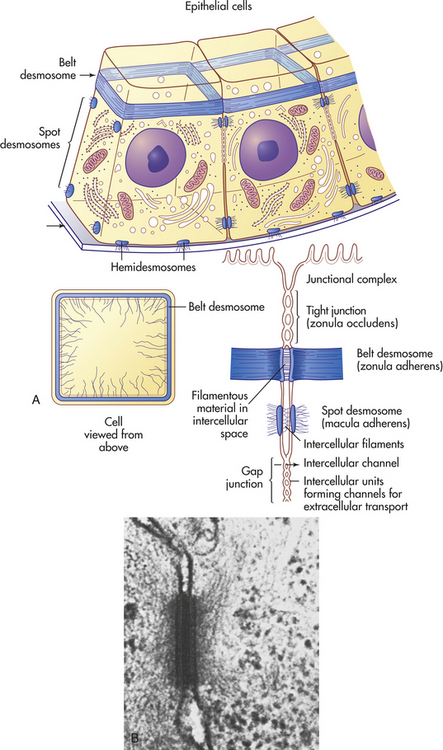
Figure 1-15 Types of cell connections. A, Schematic drawing of a belt desmosome between epithelial cells. This junction, also called zonula adherens, encircles each interacting cell. The spot desmosomes and hemidesmosomes, like the belt desmosomes, are adhering junctions. This tight junction is an impermeable junction that holds cells together but seals them in such a way that molecules cannot leak between them. The gap junction, as a communicating junction, mediates the passage of small molecules from one interacting cell to the other. B, Electron micrograph of desmosomes. (From Raven PH, Johnson GB: Biology, St Louis, 1992, Mosby.)
The junctional complex is a highly permeable part of the plasma membrane. Its permeability is controlled by a process called gating, which depends on concentrations of calcium ions in the cytoplasm. Increased cytoplasmic calcium causes decreased permeability at the junctional complex. Gating is an important cellular defense mechanism because it enables uninjured cells to seal themselves off from injured neighbors. As damaged cells release calcium, it travels through the junctional complex and increases calcium levels in neighboring cells. (The damaging effects of calcium influx are described in Chapter 2.) This decreases the permeability of the junctional complexes of the neighboring cells, which form a relatively impermeable wall around the injured area.
CELLULAR COMMUNICATION AND SIGNAL TRANSDUCTON
Cells need to communicate with each other to maintain a stable internal environment, or homeostasis; to regulate their growth and division and their development and organization into tissues; and to coordinate their functions. Cells communicate in three ways: (1) they form protein channels (gap junctions, see preceding discussion) that directly coordinate the activities of adjacent cells; (2) they display plasma membrane–bound signaling molecules (receptors) that affect the cell itself and other cells in direct physical contact; and (3) (the most common means) they secrete chemicals that signal to cells some distance away (Figure 1-16). Alterations in cellular communication affect disease onset and progression. In fact, if a cell is unable to perform gap junctional intercellular communication, it is hypothesized that normal growth control and cell differentiation are compromised, favoring cancerous tumor development (see Chapter 11). (Communication through gap junctions is discussed earlier, and contact signaling by plasma membrane–bound molecules is shown in Figure 1-16.) Secreted chemical signals involve communication at a distance. Primary modes of chemical signaling are hormonal, neurohormonal, paracrine, autocrine, and neurotransmitter (Figure 1-17).
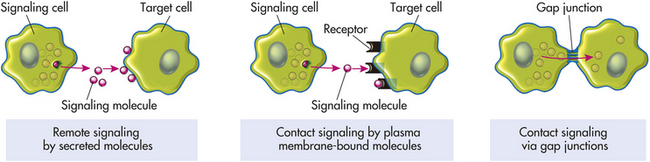
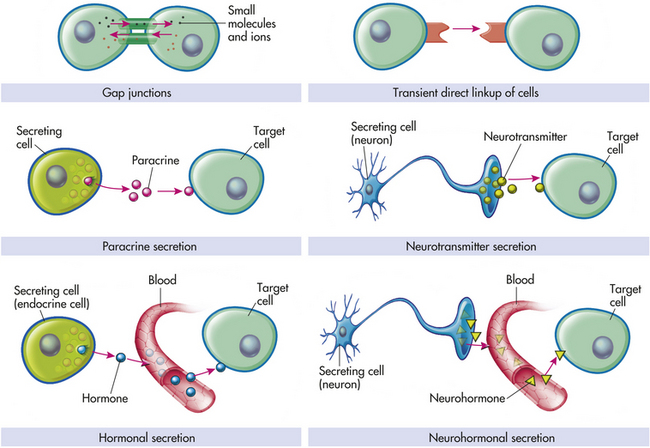
Figure 1-17 Modes of chemical signaling and cell communication. Paracrines, neurotransmitters, hormones, and neurohormones are all intercellular chemical messengers that accomplish communication between cells. Gap junctions provide the most intimate means of intercellular communication where small molecules and ions are exchanged between interacting cells without even entering the extracellular fluid. Autocrine stimulation (not illustrated) occurs when the secreting cell targets itself.
Hormonal signaling involves specialized endocrine cells that secrete hormone chemicals (e.g., thyroid-stimulating hormone) released by one set of cells and travel through the tissue and through the bloodstream to produce a response in other sets of cells (see Chapter 20). In neurohormonal signaling, hormones (e.g., angiotensin II) are released into the blood by neurosecretory neurons. Like endocrine cells, neurosecretory neurons release blood-borne chemical messengers, whereas ordinary neurons secrete short-range neurotransmitters (e.g., acetylcholine) into a small discrete space. In paracrine signaling, cells secrete local chemical mediators that are quickly taken up, destroyed, or immobilized. The mediators act only on nearby cells. In autocrine signaling, signaling molecules may act back on the cells of origin (i.e., autostimulation); autocrine circuits function as a component of normal growth-regulatory mechanisms in many adult tissue types.16,17 Neurons communicate directly with the cells they innervate by releasing chemicals or neurotransmitters at specialized junctions called chemical synapses; the neurotransmitter diffuses across the synaptic cleft and acts on the postsynaptic target cell (Figure 1-17). In each type of chemical signaling, the target cell receives the signal by first attaching to its receptors. Many of these same signaling molecules are receptors used in hormonal, neurohormonal, paracrine, and autocrine signaling. The important differences lie in the speed and selectivity with which the signals are delivered to their targets.1
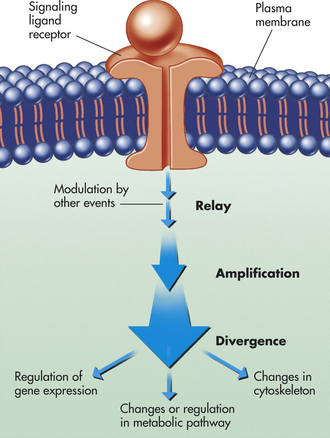
Figure 1-18 An intracellular signaling cascade. An extracellular chemical messenger (ligand) binds to a receptor protein located on the plasma membrane, where it is transduced into an intracellular signal. This process initiates a signaling cascade that relays the signal into the cell interior, amplifying and distributing it en route. Steps in the cascade can be modulated by other events in the cell.
Plasma membrane receptors belong to one of three classes that are defined by the signaling (transduction) mechanism used. Table 1-2 summarizes these receptors.
Signal Transduction
Signal transduction involves incoming signals or instructions from extracellular chemical messengers (ligands) that are conveyed to the cell’s interior for execution. Within the outer surface of the plasma membrane, specialized protein receptors bind with the selected chemical messengers. This combination of messenger with receptor triggers a cascade of cellular events important to the maintenance of homeostasis, such as membrane transport, cell division and differentiation, movement, secretion, and metabolism. Some types of altered cell behavior, such as increased cell growth and division, involve changes in gene expression and the synthesis of new proteins and therefore occur slowly. Others, such as changes in cell movement, secretion, or metabolism, do not involve the nuclear machinery and therefore occur more rapidly. If deprived of appropriate signals, most cells undergo a form of cell suicide known as programmed cell death, or apoptosis (see p. 84).
Signaling cascades, or relay chains, of intercellular signaling molecules have several important functions (see Figure 1-18):
1. They physically transfer the signal from the place at which it is received to some other part of the cell where the response is expected.
2. They amplify the signal received, making it stronger; this is caused by a multiplying effect in the pathways; for example, binding of one ligand molecule to a receptor activates a number of adenylyl cyclase molecules.
3. They distribute the signal so that it influences several processes in parallel; at any step in the pathway, the signal can diverge and be relayed to several different intracellular targets, creating branches in the flow and causing a complex response (Figure 1-19).
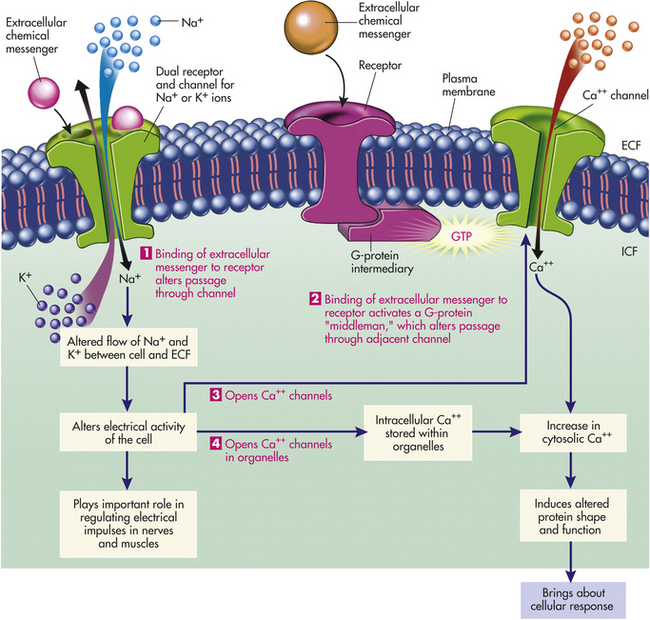
Figure 1-19 How extracellular messengers regulate channel function. Binding of an extracellular messenger to a dual receptor/channel brings about a quick opening or closing of ion channels, such as Na+ or K+ channels, which generates electrical impulses (1). A transient opening of membrane Ca++ channels occurs when binding of an extracellular messenger to a receptor activates a G-protein intermediary, which alters a nearby ion channel, such as a Ca++ channel (2). A transient opening of Ca++ channels also occurs indirectly in response to electrical impulses produced by extracellular messenger-induced changes in Na+ and K+ channels (3). Release of Ca++ from intracellular stores results when Ca++ channels in organelles open in response to electrical impulses (4). An increase in cytosolic Ca++ arising from pathways 2, 3, or 4 causes change in the shape and function of specific intracellular proteins to produce the desired cellular response. ECF, Extracellular fluid; GTP, guanosine triphosphate; ICF, intracellular fluid. (Redrawn with permission from Sherwood L: Human physiology, ed 3. © 1997 Brooks/Cole, a partof Cengage Learning, Inc. Reproduced by permission. www.cengage.com/permissions.)
4. Last, the signal can be modulated by other interfering factors prevailing inside or outside the cell.
Two general responses from binding of the extracellular chemical messenger, or first messenger, to the membrane receptors occur: (1) opening or closing specific channels in the membrane to regulate the movement of ions into or out of the cell, and (2) transferring the signal to an intracellular messenger, or second messenger, which in turn triggers a cascade of biochemical events within the cell.
Extracellular Messengers and Channel Regulation
Membrane channels, or “gates,” can open and close depending on the circumstances of the first messenger. Opening and closing occur because of conformational changes (shaping) of the proteins that form the channels—blocking the channel (closing) or permitting passage through it (opening). Channel opening and closing can be initiated in one of three ways: (1) by binding of a ligand to a specific membrane receptor that is closely associated with the channel (for example, G proteins); (2) by changes in electric current in the plasma membrane, altering flow of Na+ and K+; and (3) by stretching or other chemical deformation of the channel. Figure 1-19 summarizes ways by which extracellular messengers regulate channel function for the other two methods of controlling channels (see p. 28).
Second Messengers
Many ligands cannot enter their target cells to bring about the desired intracellular response. Instead, the first messengers, or ligands, issue orders by binding with receptors on the surface membrane, triggering a “pass it on” signal. Second messengers are generated in large numbers when the membrane-bound enzyme is activated, and they then rapidly diffuse away from their source, broadcasting the signal throughout the cell (Figure 1-20). Remember, most cell-surface receptor proteins belong to one of three large classes: ion-channel-linked receptors, G-protein-linked receptors, or enzyme-linked receptors.
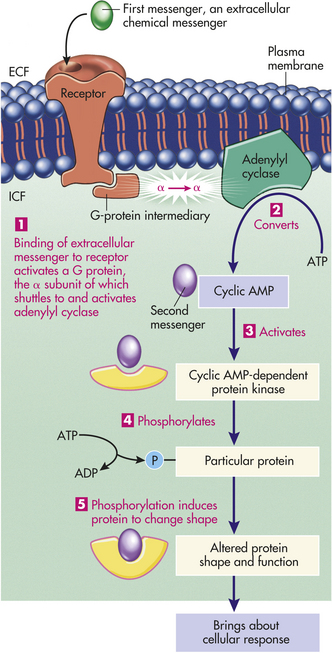
Figure 1-20 Extracellular messenger and activation of the cAMP second messenger system. The first messenger, or binding of an extracellular chemical messenger to a surface membrane receptor, activates the membrane-bound enzyme adenylyl cyclase by means of a G-protein intermediary (1), which in turn converts intracellular ATP into cAMP (2). cAMP is an intracellular second messenger, triggering the cellular response by activating the cAMP-dependent protein kinase (3), which in turn phosphorylates (4), and therefore modifies (5) a specific intracellular protein. The altered protein then directs the cellular response dictated by the extracellular messenger. ADP, adenosine diphosphate; AMP, adenosine monophosphate; ATP, adenosine triphosphate; ECF, Extracellular fluid; ICF, intracellular fluid. (Redrawn with permission from Sherwood L: Humanphysiology, ed 3. © 1997 Brooks/Cole, a part of Cengage Learning, Inc.Reproduced by permission. www.cengage.com/permissions.)
The two major second messenger pathways are cyclic adenosine monophosphate (cyclic AMP, cAMP) and Ca++. In the cAMP pathway, binding of the ligand to its surface receptor eventually activates the enzyme adenylyl cyclase on the inner surface of the membrane. A membrane-bound “middleman,” a G protein, acts as an intermediary between the receptor and adenylyl cyclase. G proteins are named because they are bound to guanine nucleotides—guanosine triphosphate (GTP) or guanosine diphosphate (GDP). An unactivated G protein consists of a complex of alpha (α), beta (β), and gamma (γ) subunits, with a GDP molecule bound to the α subunit. The cAMP pathway with G proteins is summarized in Figure 1-20.
Instead of cAMP, some cells use Ca++ as a second messenger. In this pathway, binding of the first messenger to the surface receptor eventually leads, by means of G proteins, to activation of the enzyme phospholipase C, an enzyme protein effector (an ion channel for an enzyme) that is bound to the inner side of the membrane. Figure 1-21 summarizes the Ca++ second messenger pathway. The cAMP and Ca++ pathways frequently overlap in bringing about a specific cellular response. For example, cAMP and Ca++ can influence each other. Calcium-activated calmodulin can regulate adenylyl cyclase and thus influence cAMP; conversely, cAMP-dependent kinase may phosphorylate and thereby change the activity of Ca++ channels or carriers. In some instances, both Ca++ and cAMP regulate the same intracellular protein. In a few cells, cyclic guanosine monophosphate (cyclic GMP, cGMP) serves as a second messenger similar to the cAMP pathway. For example, cGMP is the signal transduction pathway involved in vision. Some cellular responses mediated by cAMP and phospholipase C are summarized in Table 1-3. Major types of receptors and signal transduction pathways are contained in Table 1-4.
Table 1-3
Hormone-Induced Cell Responses Mediated by cAMP
| Signaling Ligands | Target Tissue | Major Response |
| Epinephrine | Heart | Increase in heart rate and force of contraction |
| Epinephrine, ACTH | Muscle | Glycogen breakdown |
| Glucagon | Fat | Fat breakdown |
| ACTH | Adrenal gland | Cortisol secretion |
| Antidiuretic hormone | Liver | Glycogen breakdown |
| Acetylcholine | Pancreas; smooth muscle | Amylase secretion; contraction |
| Antigen | Mast cells | Histamine secretion |
| Thrombin | Blood platelets | Serotonin and platelet-derived growth factor secretion; platelet aggregation |
ACTH, Adrenocorticotropic hormone; cAMP, Cyclic adenosine monophosphate.
Table 1-4
Major Types of Receptors and Signaling Transduction Pathways

ADH, Antidiuretic hormone; cAMP, cyclic adenosine monophosphate; EGF, epidermal growth factor; FGF, fibroblast growth factor; G-CSF, granulocyte colony-stimulating factor; HGF, hepatocyte growth factor; IL-2, IL-3, interleukin-2 and interleukin-3; IP3, inositol triphosphate; JAK/STAT, Janus kinase-signal transducers and activators of transcription; MAP-kinase, mitogen activated protein kinase; PDGF, platelet-derived growth factor; TGF-α, transforming growth factor-alpha; VEGF, vascular endothelial growth factor.
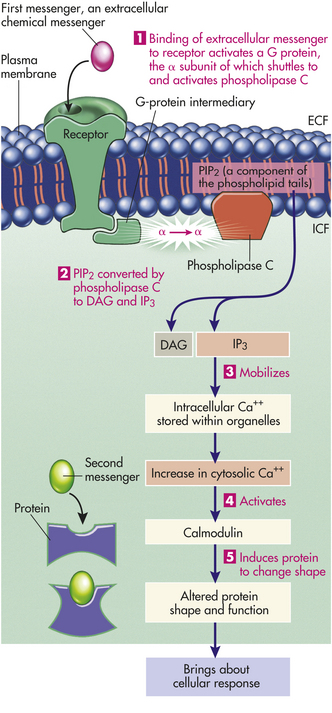
Figure 1-21 Extracellular messenger and activation of the calcium second messenger system. Binding of an extracellular messenger to a membrane receptor activates the membrane-bound enzyme phospholipase C by means of a G-protein intermediary (1). Phospholipase C converts phosphatidylinositol biphosphate (PIP2) into diacylglycerol (DAG) and inositol triphosphate (IP3) (2). IP3 then mobilizes Ca++ stored within organelles (3). Ca++, as a second messenger, activates calmodulin (4), causing a change in the shape and function of a specific intracellular protein to produce the cellular response (5). ECF, Extracellular fluid; ICF, intracellular fluid. (Redrawn withpermission from Sherwood L: Human physiology, ed 3. © 1997 Brooks/Cole,a part of Cengage Learning, Inc. Reproduced by permission. www.cengage.com/permissions.)
A large number of human disorders involve problematic signaling in cells. Cancer, for example, results from genetic mutations leading to the overactivity of proteins in signal relaying pathways that normally induce the cells to divide. Affected proteins cause cells to behave as if other cells were constantly telling them to reproduce, even when no such orders were sent.18 Signal blockers are already in use against breast cancer.
CELLULAR METABOLISM
All the chemical tasks of maintaining essential cellular functions are referred to as cellular metabolism. The energy-using process of metabolism is called anabolism (ana = upward), and the energy-releasing process is known as catabolism (cata = downward). Metabolism provides the cell with the energy it needs to synthesize (produce) cellular structures.
Dietary proteins, fats, and starches are hydrolyzed in the intestinal tract into amino acids, fatty acids, and glucose. These constituents are then absorbed, circulated, and taken up by the cell, where they may be used for various vital cellular processes, including the production of ATP. The process by which ATP is produced is one example of a series of reactions called a metabolic pathway. A metabolic pathway involves several intermediate steps whose end products are not always detectable. A key feature of cellular metabolism is the directing of biochemical reactions by protein catalysts, or enzymes. Most biochemical reactions in a pathway are catalyzed by a specific enzyme. Each enzyme has a high affinity for a substrate—a specific substance that is converted to a product of the reaction.
Role of Adenosine Triphosphate
For a cell to function it must be able to extract and use the chemical energy contained within the structure of organic molecules. When 1 mole of glucose is metabolically broken down in the presence of oxygen into carbon dioxide (CO2) and water (H2O), 686 kilocalories (kcal) of energy are released. In a test tube this energy is released as heat. Because a cell cannot transform heat into work, chemical energy, rather than heat, is created by metabolism. The chemical energy lost by one molecule is transferred to the chemical structure of another molecule by an energy-carrying or transferring molecule, such as ATP. The energy stored in ATP can be used in a variety of energy-requiring reactions and in the process is generally converted to adenosine diphosphate (ADP) and inorganic phosphate (Pi). The energy available as a result of this reaction is about 7 kcal/mol of ATP. In addition to its use in synthesis (anabolism) of organic molecules, ATP is used by the cell for muscle contraction and active transport of molecules across cellular membranes. The function of ATP is not only to store energy but also to transfer it from one molecule to another. Energy is stored by molecules of carbohydrate, lipid, and protein, which, when catabolized, transfer energy to ATP.
Food and Production of Cellular Energy
The process of catabolism of the proteins, lipids, and polysaccharides found in food can be divided into three phases (Figure 1-22). In phase 1, large molecules are broken down into their smaller subunits—proteins into amino acids, polysaccharides into simple sugars, and fats into fatty acids and glycerol. These processes are called digestion and occur outside the cell by the action of secreted enzymes.
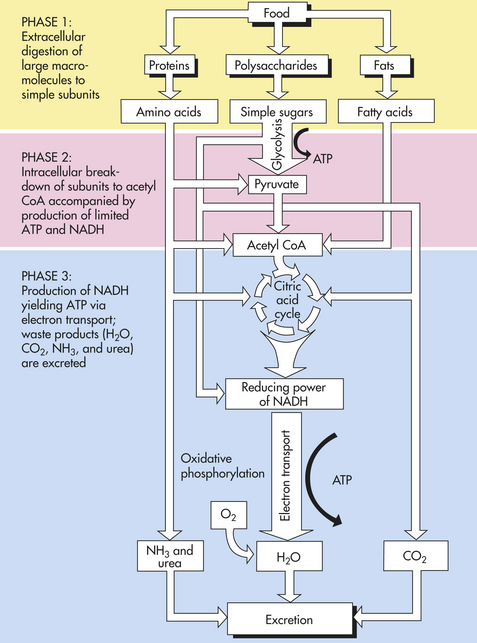
Figure 1-22 Three phases of catabolism, which leads from food to waste products. These reactions produce ATP, which is used to drive other processes in the cell.
In phase 2 the small molecules enter cells and are further broken down in the cytoplasm. Most of the sugars are converted into pyruvate. Pyruvate then enters mitochondria and is converted to the acetyl groups of acetyl coenzyme A (acetyl CoA). Acetyl CoA, like ATP, releases energy when it is hydrolyzed. The most important part of phase 2 is the lysis (splitting) of glucose, known as glycolysis (Figure 1-23). Glycolysis produces a net of two molecules of ATP per glucose molecule through the process of oxidation, or the removal and transfer of a pair of electrons. This process, often called oxidative cellular metabolism, involves 10 biochemical reactions. In reactions 1 through 5, glucose is converted to two, three-carbon aldehyde (glyceraldehyde-3-phosphate [G3P]), which requires energy in the form of ATP. The next five reactions convert G3P molecules into pyruvate molecules and generate four molecules of ATP for each two molecules of G3P. In addition, two molecules of nicotinamide adenine dinucleotide (NAD) are further oxidized to produce four more molecules of ATP. After subtracting two molecules of ATP to drive the reactions, the net yield is six ATP molecules for each molecule of glucose.
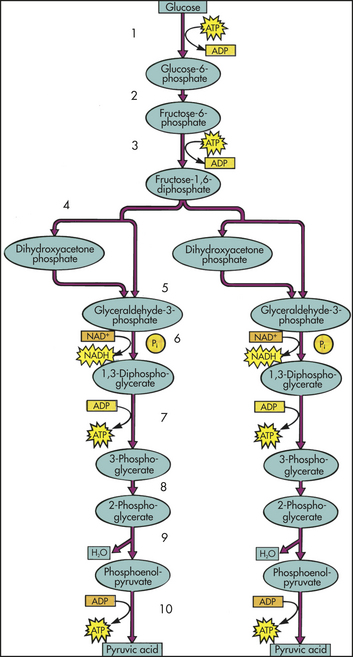
Figure 1-23 Glycolysis. Each of the numbered reactions is catalyzed by a different enzyme. At step 4, a six-carbon sugar is broken down to give two three-carbon sugars, so that the number of molecules at every step after this is doubled. Reactions 5 and 6 are the reactions responsible for the net synthesis of adenosine triphosphate (ATP) and reduced nicotinamide adenine dinucleotide (NADH) molecules. (Modified from Thibodeau GA, Patton KT: Anatomy & physiology, ed 6, St Louis, 2007, Mosby.)
Phase 3 occurs when the acetyl group of acetyl CoA is completely degraded to CO2 and H2O. It is in this final phase that most of the ATP is generated. Phase 3 begins with the citric acid cycle (also called the Krebs cycle or the tricarboxylic acid cycle) and ends with oxidative phosphorylation. The citric acid cycle accounts for approximately two thirds of the total oxidation of carbon compounds in most cells. Its major end products are CO2 and two dinucleotides, reduced NADH, and the reduced form of flavin adenine dinucleotide (FADH2), which transfer their electrons into the electron-transport chain.
Oxidative Phosphorylation
Oxidative phosphorylation occurs in the mitochondria and is the mechanism by which the energy produced from carbohydrates, fats, and proteins is transferred to ATP. During the breakdown (catabolism) of foods, many of the reactions involve the removal of electrons from various intermediates. These reactions generally require a coenzyme (a nonprotein carrier molecule), such as nicotinamide adenine dinucleotide (NAD), to transfer the electrons and thus are called transfer reactions.
In oxidative phosphorylation, molecules of NAD and flavin adenine dinucleotide (FAD) transfer electrons they have gained from the oxidation of substrates to molecular oxygen, O2. The electrons from reduced NAD and FAD, NADH and FADH2, are transferred to a series of carrier molecules (the electron-transport chain) on the inner surfaces of the mitochondria with the release of hydrogen ions. Some of the carrier molecules are a group of brightly colored iron-containing proteins known as cytochromes that accept a pair of electrons. After passing through a sequence of different cytochromes, these electrons are eventually combined with molecular oxygen. If oxygen is not available to the electron-transport chain, ATP will not be formed by the mitochondria. Instead, an anaerobic (without oxygen) metabolic pathway synthesizes ATP. This process, called substrate phosphorylation, or anaerobic glycolysis, does not take place in the mitochondria and is linked to the breakdown (glycolysis) of carbohydrate (Figure 1-24).
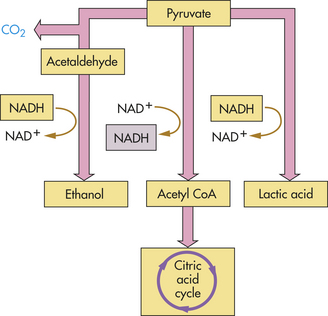
Figure 1-24 What happens to pyruvate, the product of glycolysis? In the presence of oxygen, pyruvate is oxidized to acetyl coenzyme A (CoA) and enters the citric acid cycle. In the absence of oxygen, pyruvate instead is reduced, accepting the electrons extracted during glycolysis and carried by reduced nicotinamide adenine dinucleotide (NADH). When pyruvate is reduced directly, as it is in muscle, the product is lactic acid. When CO2 is first removed from pyruvate and the remainder reduced, as it is in yeasts, the product is ethanol.
Because glycolysis occurs in the cytoplasm of the cell, it provides energy for cells that lack mitochondria. However, as noted, glycolysis also provides energy to the cell when oxygen delivery is insufficient or delayed (e.g., with strenuous exercise). The reactions in anaerobic glycolysis involve the conversion of glucose to pyruvic acid (pyruvate) with the simultaneous production of ATP. With the glycolysis of one molecule of glucose, two ATP molecules and two molecules of pyruvate are liberated. If oxygen is present, the two molecules of pyruvate move into the mitochondria, where they enter the citric acid cycle. If oxygen is absent, pyruvate is converted to lactic acid, which is released into the extracellular fluid (see Figure 1-24). The conversion of pyruvic acid to lactic acid is reversible; therefore, once oxygen is restored, lactic acid is quickly converted back to either pyruvic acid or glucose. The anaerobic generation of ATP from glucose, through the reactions of glycolysis, is not as efficient as the aerobic generation of ATP. The addition of an oxygen-requiring stage to the catabolic process (stage 3) provides cells with a much more powerful method for extracting energy from food molecules.
MEMBRANE TRANSPORT: CELLULAR INTAKE AND OUTPUT
Cells continually take in nutrients, fluids, and chemical messengers from the extracellular environment and expel metabolites or the products of metabolism and end products of lysosomal digestion. Intake and output, or transport, occur by different mechanisms, depending on the characteristics of the substance to be transported. Water and small electrically uncharged molecules move easily through pores in the plasma membrane’s lipid bilayer. This process, called passive transport, will occur naturally through any semipermeable barrier. It is driven by osmosis, hydrostatic pressure, and diffusion, all of which depend on the laws of physics and do not require life. The process is passive in that it does not require any expenditure of energy by the cell.
Other molecules cannot be driven across the plasma membrane solely by forces of diffusion, hydrostatic pressure, or osmosis because they are too large or are ligands that have bound with receptors on the cell’s plasma membrane. Some of these molecules are moved into the cell by mechanisms of active transport, which requires life, biologic activity, and the expenditure of metabolic energy by the cell. Unlike passive transport, which can be duplicated across any semipermeable barrier in a laboratory, active transport occurs only across living membranes that (1) use energy generated by cellular metabolism and (2) have receptors that are capable of recognizing and binding with the substance to be transported. Large molecules (macromolecules), along with fluids, are transported by means of endocytosis (taking in) and exocytosis (expelling). Water and electrically charged molecules are transported by protein channels embedded in the plasma membrane. Ligands enter the cell by means of receptor-mediated endocytosis.
Movement of Water and Solutes
Cellular membranes are semipermeable and generally allow passage of water and small particles of dissolved substances called solutes. The movement of solute molecules through membranes is related to their size, solubility, electrical properties, and concentration on either side of the membrane. Small lipid-soluble particles, such as oxygen, carbon dioxide, and urea, can readily pass through the lipid bilayers of the plasma membrane. Larger, water-soluble particles may pass through pores in the membranes. Although large protein molecules, such as albumin and globulin, pass through membranes by endocytosis, they influence the movement of water by exerting an osmotic effect (see p. 27).
Body fluids are composed of two types of solutes: electrolytes, which are electrically charged and dissociate into constituent ions when placed in solution; and nonelectrolytes, such as glucose, urea, and creatinine, which do not dissociate. Electrolytes account for approximately 95% of the solute molecules in body water. Electrolytes exhibit polarity by orienting themselves toward the positive or negative pole. Ions with a positive charge are known as cations and migrate toward the negative pole, or cathode, if an electrical current is passed through the electrolyte solution. Anions carry a negative charge and migrate toward the positive pole, or anode, in the presence of electrical current. Anions and cations are located in both the intracellular fluid (ICF) and extracellular fluid (ECF) compartments, although concentration of particular ions varies depending on their location. (Fluid and electrolyte balance between body compartments is discussed in Chapter 3.) For example, Na+ is the predominant extracellular cation, and K+ is the principal intracellular cation. The difference in ICF and ECF concentrations of these ions is important to the transmission of electrical impulses across the plasma membranes of nerve and muscle cells.
Electrolytes are measured in milliequivalents per liter (mEq/L) or milligrams per deciliter (mg/dl). Milliequivalents per liter indicate the number of electrical charges per unit volume of fluid. The term milliequivalent thus indicates the chemical-combining activity of an ion, which depends on the electrical charge, or valence, of its ions. In abbreviations, valence is indicated by the number of plus or minus signs. Monovalent ions, or ions with one charge, include sodium (Na+), chloride (Cl−), and potassium (K+). Divalent ions, which have two charges, include calcium (Ca++) and magnesium (Mg++). One milliequivalent of any cation can combine chemically with 1 mEq of any anion: one monovalent anion will combine with one monovalent cation. Divalent ions combine more strongly than monovalent ions. To maintain electrochemical balance, one divalent ion will combine with two monovalent ions (e.g., Ca++ + 2 Cl− = CaCl2).
Passive Transport: Diffusion, Filtration, and Osmosis
Diffusion: Diffusion is the movement of a solute molecule from an area of greater solute concentration to an area of lesser solute concentration. This difference in concentration is known as a concentration gradient. Particles in a solution move randomly in any direction. If the concentration of particles in one part of the solution is greater than in another part, the particles distribute themselves evenly throughout the solution. According to the same principle, if the concentration of particles is greater on one side of a permeable membrane than on the other side, the particles diffuse spontaneously from the area of greater concentration to the area of lesser concentration until equilibrium is reached. The higher the concentration on one side, the greater the diffusion rate. The overall effect of diffusion is the passive movement of particles “down” a concentration gradient, that is, from an area of high concentration to an area of low concentration.
The diffusion rate is influenced by differences of electrical potential across the membrane (see p. 33). Because the pores in the lipid bilayer are often linked with Ca++, other cations (e.g., Na+ and K+) diffuse slowly because they are repelled by positive charges in the pores.
The rate of diffusion of a substance depends also on its size (diffusion coefficient) and its lipid solubility (Figure 1-25). Usually the smaller the molecule and the more soluble it is in oil, the more hydrophobic or nonpolar it is and the more rapidly it will diffuse across the bilayer. Oxygen, carbon dioxide, and the steroid hormones are all examples of nonpolar molecules. Water-soluble substances, such as sugars and inorganic ions, diffuse very slowly, whereas uncharged lipophilic (“lipid-loving”) molecules, such as fatty acids and steroids, diffuse rapidly. Ions and other polar molecules generally diffuse across cellular membranes more slowly than lipid-soluble substances.
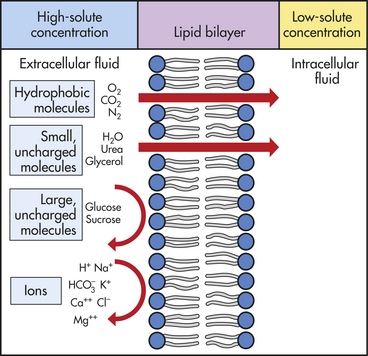
Figure 1-25 Passive diffusion of solute molecules across plasma membrane. Oxygen, nitrogen, water, urea, glycerol, and carbon dioxide can diffuse readily down the concentration gradient. Macromolecules are too large to diffuse through pores in the plasma membrane. Ions may be repelled if the pores contain substances with identical charges. If the pores are lined with cations, for example, other cations will have difficulty diffusing because the positive charges will repel one another. Diffusion can still occur, but it occurs more slowly.
Water readily diffuses through biologic membranes because water molecules are small and uncharged. Although the mechanism is not known with certainty, the dipolar structure of water allows it to cross rapidly the regions of the bilayer containing the lipid head groups. Lipid head groups constitute the two outer regions of the lipid bilayer.
Filtration: Hydrostatic Pressure: Filtration is the movement of water and solutes through a membrane because of a greater pushing pressure (force) on one side of the membrane than on the other side. Hydrostatic pressure is the mechanical force of water pushing against cellular membranes. In the vascular system, hydrostatic pressure is the blood pressure generated in vessels by the contraction of the heart. Blood reaching the capillary bed has a hydrostatic pressure of 25 to 30 mmHg, which is sufficient force to push water across the thin capillary membranes into the interstitial space. Hydrostatic pressure is partially balanced by osmotic pressure, whereby water moving out of the capillaries is partially balanced by osmotic forces that tend to pull water into the capillaries. Water that is not osmotically attracted back into the capillaries moves into the lymph system (see discussion of Starling forces in Chapter 3).
Osmosis: Osmosis is the movement of water “down” a concentration gradient, that is, across a semipermeable membrane from a region of higher water concentration to a lower water concentration. For osmosis to occur, the membrane must be more permeable to water than to solutes and the concentration of solutes must be greater so that water moves more easily. Osmosis is directly related to both hydrostatic pressure and solute concentration but not to particle size or weight. For example, particles of the plasma protein albumin are small but more concentrated in body fluids than the larger and heavier particles of globulin. Therefore, albumin exerts a greater osmotic force than globulin.
Osmolality controls distribution and movement of water between body compartments. The terms osmolality and osmolarity are often used interchangeably in reference to osmotic activity, but they define different measurements. Osmolality is a measure of the number of milliosmoles per kilogram of water, or the concentration of molecules per weight of water. Osmolarity is a measure of the number of milliosmoles per liter of solution, or the concentration of molecules per volume of solution. When solute is added to water, the volume is expanded and includes the original liter of water plus the volume occupied by the solute particles. In measuring osmolarity, the volume of water is therefore reduced by an amount equal to the volume of added solute.
In solutions that contain only dissociable substances, such as Na+ and Cl−, the difference between the two measurements is negligible. In considering all the different solutes in plasma (e.g., proteins, glucose, lipids), however, the difference between osmolality and osmolarity becomes more significant. In plasma, less of the plasma weight is water and the overall concentration of particles is therefore greater. The osmolality will be greater than the osmolarity because of the smaller proportion of water. Osmolality is thus the preferred measure of osmotic activity in clinical assessment of individuals.
The normal osmolality of body fluids is 280 to 294 mOsm/kg (milliosmoles per kilogram). The osmolality of intracellular and extracellular fluid tends to equalize and so provides a measure of body fluid concentration and thus the body’s hydration status (see Chapter 3). Hydration is also affected by hydrostatic pressure because the movement of water by osmosis can be opposed by an equal amount of hydrostatic pressure. The amount of hydrostatic pressure required to oppose the osmotic movement of water is called the osmotic pressure of the solution. Factors that determine osmotic pressure are the type and thickness of the plasma membrane, the size of the molecules, the concentration of molecules or the concentration gradient, and the solubility of molecules within the membrane. Examples of movement of water in relation to hydrostatic and osmotic forces occur in the glomerulus in the kidney (see Chapter 35) and in the capillaries of the microcirculation (see Chapter 29).
Effective osmolality is sustained osmotic activity and depends on the concentration of solutes remaining on one side of a permeable membrane. If the solutes penetrate the membrane and equilibrate with the solution on the other side of the membrane, the osmotic effect will be diminished or lost. For example, urea is a small solute that readily diffuses across cellular membranes. Solutions containing urea rapidly lose their effective osmolality because they rapidly equilibrate. Solutes too large to pass through the membrane thus sustain an effective osmolality, meaning that they enhance osmotic activity. Plasma proteins are examples of molecules that provide effective osmolality because they normally do not cross cellular membranes.
Plasma proteins also influence osmolality because they have a negative charge. The principle by which the plasma protein charge influences osmolality is known as Gibbs-Donnan equilibrium, and it affects the distribution of ions across cellular membranes. Gibbs-Donnan equilibrium occurs when fluid in one compartment contains small diffusible ions such as Na+ and Cl−, together with large, nondiffusible charged particles, such as plasma proteins. Because the body tends to maintain an electrical equilibrium, the nondiffusible protein molecules cause asymmetry in the distribution of small ions. Anions such as Cl− are thus driven out of the cell or plasma, and cations such as Na+ are attracted. The protein-containing compartment will maintain a state of electroneutrality, but the osmolality will be higher. The overall osmotic effect of colloids, such as plasma proteins, is called the oncotic pressure, or colloid osmotic pressure.
Tonicity describes the effective osmolality of a solution. (The terms osmolality and tonicity may be used interchangeably; also see Chapter 3.) Solutions, then, have relative degrees of tonicity. An isotonic solution (or isoosmotic solution) has the same osmolality or concentration of particles (285 mOsm/kg) as the ICF or ECF. Diarrhea, for example, is loss of isoosmotic fluid from the gastrointestinal tract. As a result, ECF volume decreases but there is no change in ECF osmolarity. Examples of isotonic solutions include 5% dextrose in water and normal (0.9%) saline solution. A hypotonic solution has a lower concentration and is thus more dilute than body fluids. Water is a hypotonic solution. Consequently, water is osmotically pulled into the cells, causing them to swell or burst. A hypertonic solution has a concentration of more than 285 to 294 mOsm/kg. An example of a hypertonic solution is 3% saline solution. Water can be pulled out of the cells by a hypertonic solution, so the cells shrink. The concept of tonicity is important when correcting water and solute imbalances by administering different types of replacement solutions.
Mediated and Active Transport
Mediated Transport: Mediated transport (passive and active) involves integral or transmembrane proteins with receptors having a high degree of specificity for the substance being transported. Inorganic anions and cations (e.g., Na+, K+, Ca++, Cl−, 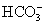 ) and charged and uncharged organic compounds (e.g., amino acids, sugars) require specific transport systems to facilitate movement through different cellular membranes. Rates at which substances are moved by mediated transport mechanisms have often been measured, yet the specific membrane proteins involved have not been identified. Mediated transport is much faster than simple diffusion.
) and charged and uncharged organic compounds (e.g., amino acids, sugars) require specific transport systems to facilitate movement through different cellular membranes. Rates at which substances are moved by mediated transport mechanisms have often been measured, yet the specific membrane proteins involved have not been identified. Mediated transport is much faster than simple diffusion.
A transport protein (carrier protein) is a transmembrane or integral protein that binds with and transfers a specific solute molecule across the lipid bilayer. (Proteins are discussed on p. 11.) Each transport protein, or transporter, has receptors for a specific solute. When the transporter is saturated—that is, when all receptor sites are occupied by solute molecules—the rate of transport is maximal. Solute binding can be blocked by competitive inhibitors that compete for the same receptor site and may or may not be transported by the transport protein. Noncompetitive inhibitors bind elsewhere but can alter the structure of the transporter.
The transporter protein is a multipass transmembrane protein; that is, its polypeptide chain crosses the lipid bilayer multiple times. This chain forms a continuous pathway enabling solutes to pass across the membrane without coming into direct contact with the hydrophobic interior of the lipid bilayer (Figure 1-26). (Transmembrane proteins are illustrated in Figure 1-13.)
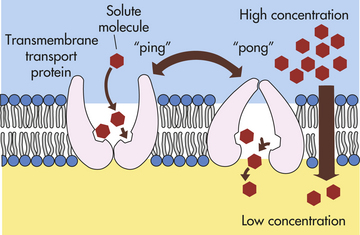
Figure 1-26 Conformational-change model of mediated transport (facilitated diffusion). The transporter protein has two states, “ping” and “pong.” In the ping state, sites for molecules of a specific solute are exposed on the outside of the bilayer. In the pong state, the sites are exposed to the inner side of the bilayer.
Another mechanism of mediated transport is the channel protein. The protein transporter creates a water-filled pore or channel across the bilayer through which specific ions can diffuse. These channels are sometimes called ion channels, and because they are permeable mainly to K+, they are also called K+ leak channels (Figure 1-27). The channel is controlled by a gate mechanism that determines which receptor-bound solutes can move into the channel that is created after receptor-solute contact. Binding stimulates conformational changes in the protein transporter that move the solute through the channel short distances at a time until it reaches the other side of the membrane. Ion channels are responsible for the electrical excitability of nerve and muscle cells and play a critical role in the membrane potential.
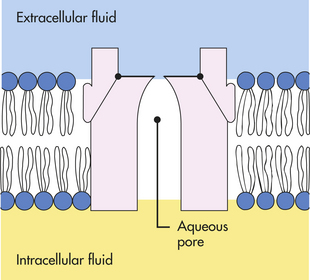
Figure 1-27 Channel mode of mediated transport (facilitated diffusion). A channel protein forms a water-filled pore across the bilayer through which specific ions can diffuse.
Mediated transport systems can move solute molecules singly or two at a time. Two molecules can be moved simultaneously in one direction (a process called symport) or in opposite directions (called antiport), or a single molecule can be moved in one direction (called uniport) (Figure 1-28).
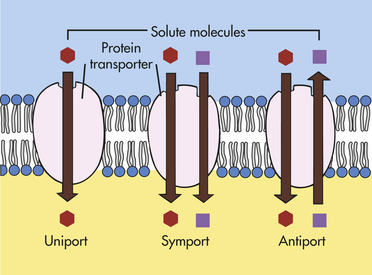
Figure 1-28 Mediated transport. Simultaneous movement of a single solute molecule in one direction (uniport), of two different solute molecules in one direction (symport), and of two different solute molecules in opposite directions (antiport).
In passive mediated transport, also called facilitated diffusion, the protein transporter moves solute molecules through cellular membranes without expending metabolic energy. The direction of movement is the same as in simple diffusion—down the concentration gradient. Perhaps the most widely referred to passive transport system is that for glucose in erythrocytes (red blood cells). Glucose is transported by a uniport mechanism and demonstrates saturation kinetics; that is, the transport system is saturated when all the glucose-specific receptors on the membrane are occupied and operating at their maximal capacity.
The anions Cl− and bicarbonate also undergo passive mediated transport in the erythrocyte. This antiport mechanism allows Cl− movement in one direction and simultaneous movement in the opposite direction. The directions of movement depend on the concentration gradients of the ions across the membrane.
In active mediated transport, also called active transport, the protein transporter moves molecules against, or up, the concentration gradient. Unlike passive mediated transport, active mediated transport requires the expenditure of energy. Many active mediated transport systems, or pumps, have ATP as their primary energy source, but not all. Some use the electrochemical gradient of Na+ across the membrane (Figure 1-29). Energy in the form of ATP, however, is required for activation of the Na+ gradient.
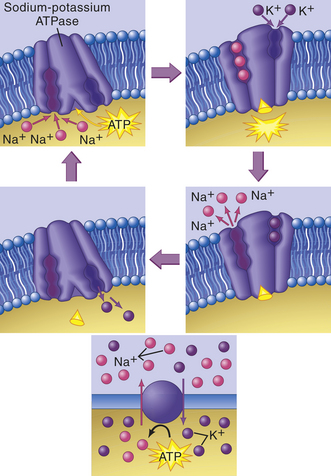
Figure 1-29 Active transport and the sodium-potassium pump. Three Na+ ions bind to sodium-binding sites on the carrier’s inner face. At the same time an energy-containing adenosine triphosphate (ATP) molecule produced by the cell’s mitochondria binds to the carrier. The ATP breaks apart, transferring its stored energy to the carrier. The carrier then changes shape, releases the three Na+ ions to the outside of the cell, and attracts two K+ ions to its potassium-binding sites. The carrier then returns to its original shape, releasing the two K+ ions and the remnant of the ATP molecule to the inside of the cell. The carrier is now ready for another pumping cycle. (From Thibodeau GA, Patton KT: Anatomy & physiology, ed 6, St Louis, 2007, Mosby.)
A “carrier” mechanism in the plasma membrane mediates the transport of ions, such as Na+, K+, H+, Cl−, and , and of nutrients, such as glucose and amino acids. Energy supplied by ATP is required to pump ions against a concentration gradient. The best-known pump is the Na+-K+–dependent ATPase pump. It continuously regulates the cells’ volume by controlling leaks through pores or protein channels and maintains the ionic concentration gradient necessary for cellular excitation and membrane conductivity (see p. 33). The maintenance of intracellular K+ concentrations is also required for enzyme activity, including that of enzymes involved in protein synthesis.
Active Transport of Na+ and K+: The active transport system for Na+ and K+ is found in virtually all mammalian cells. The Na+-K+ antiport system (Na+ moving out of and K+ moving into the cell) uses the direct energy of ATP to move these cations. The transporter protein is an enzyme, ATPase. ATPase has a requirement for Na+, K+, and Mg++ ions. The concentration of ATPase in plasma membranes is directly related to Na+-K+ transport activity. Approximately 60% to 70% of the ATP synthesized by cells, especially muscle and nerve cells, is used to maintain the Na+-K+ transport system. Excitable tissues (e.g., muscle and nerve tissues) have a high concentration of Na+-K+ ATPase, as do other tissues that transport significant amounts of Na+, for example, kidneys and salivary glands. For every ATP molecule hydrolyzed, three molecules of Na+ are transported out of the cell, whereas only two molecules of K+ move into the cell. The process leads to an electrical potential and is called electrogenic, with the inside of the cell more negative than the outside. The exact mechanism for transport of Na+ and K+ across the membrane is uncertain. One proposal is that ATPase induces the transporter protein to undergo several conformational changes, causing Na+ and K+ to move short distances (see Figure 1-29). The conformational change creates a lowering affinity for Na+ and K+ to the ATPase transporter, resulting in the release of the cations after transport.
The sarcoplasmic reticulum of heart muscle and skeletal muscle has an ATP-dependent Ca++ active transport system that regulates the Ca++ levels in the cell’s cytoplasm, which in turn regulates muscle contraction and relaxation cycles (see Chapter 29). The Ca++ transport system depends on ATPase activity and is similar to that of Na+-K+ ATPase.
The transport of sugars and amino acids across the plasma membrane depends on the simultaneous movement (symport) of Na+ or Na+-dependent transport (see Figure 1-28). Na+-dependent symport occurs primarily in the plasma membrane of epithelial cells of the kidney tubules and intestines. The transport of glucose is not directly dependent on the hydrolysis of ATP; however, the Na+ gradient is ATP dependent, and thus ATP is indirectly involved in glucose transport.
The epithelial cells that line the intestines depend on Na+ to transport various amino acids. Similarly, the uptake of Cl− by the small intestine depends on Na+ symport and antiport mechanisms for the secretion of Ca++ from the cell.
Table 1-5 summarizes the major mechanisms of transport through pores and protein transporters in the plasma membranes. Many disease states are caused or manifested by loss of these membrane transport systems.
Table 1-5
Major Transport Systems in Mammalian Cells
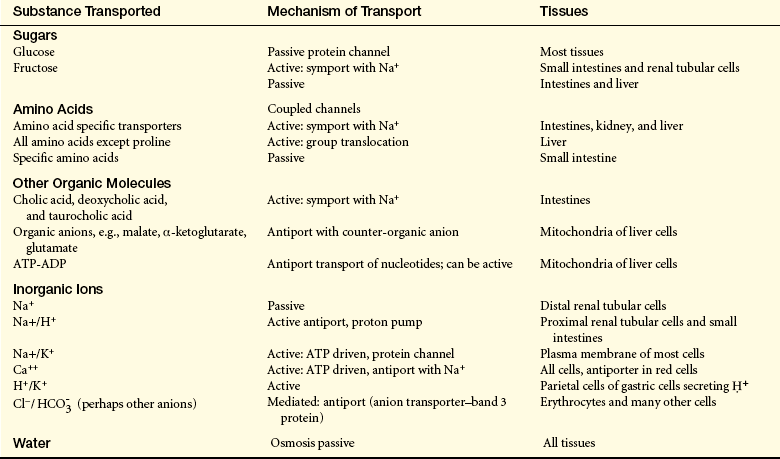
NOTE: The known transport systems are listed here; others have been proposed. Most transport systems have been studied in only a few tissues, and their sites of activity may be more limited than indicated.
ADP, Adenosine diphosphate; ATP, adenosine triphosphate.
Data from Alberts B et al: Molecular biology of the cell, ed 4, New York, 2001, Garland; Devlin TM, editor: Textbook of biochemistry: with clinical correlations, ed 3, New York, 1992, Wiley; Raven PH, Johnson GB: Understanding biology, ed 3, Dubuque, IA, 1995, Brown.
Transport by Vesicle Formation
The active transport mechanisms by which the cells move large proteins, polynucleotides, or polysaccharides (macromolecules) across the plasma membrane are very different from those that mediate small solute and ion transport. Transport of macromolecules involves the sequential formation and fusion of membrane-bound vesicles.
In endocytosis a section of the plasma membrane enfolds substances from outside the cell, invaginates (folds inward), and separates from the plasma membrane, forming a vesicle that moves into the inside of the cell (Figure 1-30, A). Two types of endocytosis are designated based on the size of the vesicle formed. Pinocytosis (cell drinking) involves the ingestion of fluids and solute molecules through formation of small vesicles, and phagocytosis (cell eating) involves the ingestion of large particles, such as bacteria, through formation of large vesicles (also called vacuoles).
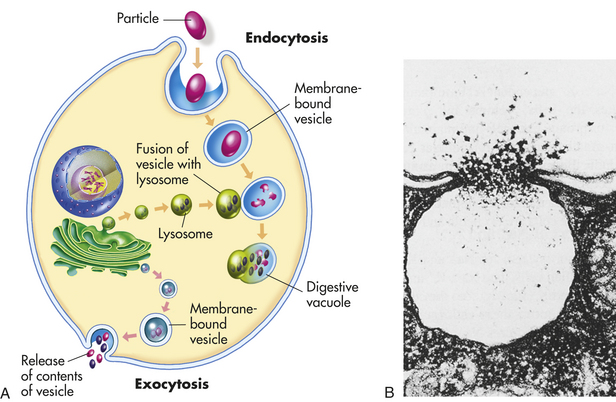
Figure 1-30 Endocytosis and exocytosis. A, Endocytosis and fusion with lysosome and exocytosis. B, Electron micrograph of exocytosis. (B from Raven PH, Johnson GB: Biology, ed 5, New York, 1999, McGraw-Hill.)
Because most cells continually ingest fluid and solutes by pinocytosis, the terms pinocytosis and endocytosis are often used interchangeably. In pinocytosis the vesicle containing fluids, solutes, or both fuses with a lysosome, and lysosomal enzymes digest them for use by the cell. In phagocytosis the large molecular substances are engulfed by the plasma membrane and enter the cell so that they can be isolated and destroyed by lysosomal enzymes (see Chapter 6). Substances that are not degraded by lysosomes are isolated in residual bodies and released by the cell by exocytosis. Both pinocytosis and phagocytosis require metabolic energy and often involve binding of the substance with plasma membrane receptors before membrane invagination and fusion with lysosomes in the cell.
In eukaryotic cells, secretion of macromolecules almost always occurs by exocytosis (see Figure 1-30, B). Exocytosis is the discharge or secretion of material from the intracellular vesicles at the cell surface. For example, to secrete macromolecules of insulin across plasma membranes, insulin-producing cells store and package insulin molecules in intracellular vesicles, which fuse with the plasma membrane and open to the extracellular space, or matrix, releasing the insulin. Not all secreted substances are secreted into the extracellular matrix. Some adhere to the plasma membrane and are thought to replace segments of the membrane lost through endocytosis or diffuse into the blood to nourish or signal other cells. Recent findings suggest membrane lipids may be a regulator of exocytosis.19 Exocytosis has two main functions: (1) replacement of portions of the plasma membrane that have been removed by endocytosis, and (2) release of molecules synthesized by the cells into the extracellular matrix.
Receptor-Mediated Endocytosis
Ligand binding to some plasma membrane receptors leads to clustering, aggregation, and immobilization of the receptors in specialized areas of the membrane called coated pits (Figure 1-31). The pits, which are coated with bristle-like structures (clathrin), deepen and enfold (invaginate), internalizing ligand-receptor complexes and forming a coated vesicle. The clathrin coat or bristles are thought to be responsible for trapping membrane receptors in coated pits. This internalization process, called receptor-mediated endocytosis (ligand internalization), is rapid and enables the cell to ingest large amounts of specific ligands without ingesting large volumes of extracellular fluid. Inside the cell, the ingested material is processed by lysosomal enzymes.
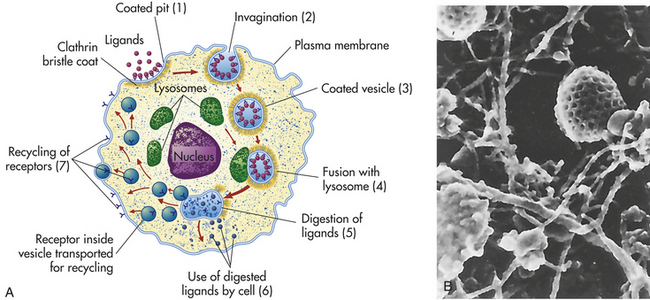
Figure 1-31 Ligand internalization by means of receptor-mediated endocytosis. A, The ligand attaches to its surface receptor (through the bristle coat or clathrin coat) and, through receptor-mediated endocytosis, enters the cell (1-3). The ingested material fuses with a lysosome (4) and is processed by hydrolytic lysosomal enzymes (5). Processed molecules can then be transferred to other cellular components (6 or recycled 7). B, Electron micrograph of a coated pit showing different sizes of filaments of the (×382,000). (B from Erlandsen SL, Magney JE: Color atlas of histology, St Louis, 1992, Mosby.)
The cellular uptake of cholesterol, for example, depends on receptor-mediated endocytosis. Cholesterol (a ligand) is carried primarily in blood plasma attached to an acceptor protein. This cholesterol-protein complex is called low-density lipoprotein (LDL). LDL receptors, which bind LDL to the plasma membrane, control the rate at which cholesterol is transferred into the cell (see Chapter 30).
Caveolae
The outer surface of the plasma membrane is dimpled with tiny flask-shaped pits (cavelike) called caveolae. Caveolae are also called microdomains. Caveolae are cholesterol-rich domains where protein caveolin are involved in several processes, including clathrin-independent endocytosis, the regulation and transport of cellular cholesterol, and cell communication.20 Many proteins, including a variety of receptors, cluster in these tiny chambers. Some of these receptors appear to be important in a new form of cellular uptake of small molecules and ions, for example, the cellular uptake of the B vitamin folic acid. When folic acid binds with its receptors, which are concentrated in the caveolae, the extracellular openings of these tiny caves close off. Closure of the caveolar indentation facilitates the movement of this vitamin across the caveolar membrane into the cytoplasm. Cellular uptake through the opening and closing of caveolae is called potocytosis. Potocytosis is thought to be an uptake mechanism for a variety of small molecules and ions, in contrast to receptor-mediated endocytosis, which transports selected large molecules into the cell. In potocytosis the caveolae are thought to remain attached to the plasma membrane and not form a membrane-enclosed vesicle such as occurs with endocytosis.
Caveolae not only function as uptake vesicles but also are important sites for signal transduction, a tedious process in which extracellular chemical messages or signals are communicated to the cell’s interior for execution (see p. 18). For example, strong evidence exists that plasma membrane estrogen receptors localize in caveolae and crosstalk with estradiol causing several intracellular functions, including cell growth and survival, migration, and new blood vessel formation.21–23
Movement of Electrical Impulses: Membrane Potentials
All body cells are electrically polarized, with the inside of the cell more negatively charged than the outside. The difference in electrical charge, or voltage, is known as the resting membrane potential and is about −70 to −85 millivolts. The difference in voltage across the plasma membrane is a result of the differences in ionic composition of ICF and ECF. Sodium ions have a greater concentration in the ECF, and potassium ions have a greater concentration in the ICF. The concentration difference is maintained by the active transport of Na+ and K+ (the sodium-potassium pump), which transports sodium outward and potassium inward (Figure 1-32). Because the resting plasma membrane is more permeable to K+ than to Na+, K+ can diffuse easily from its area of higher concentration in the ICF to its area of lower concentration in the ECF. Because Na+ and K+ are both cations, the net result is an excess of anions inside the cell, resulting in the resting membrane potential.
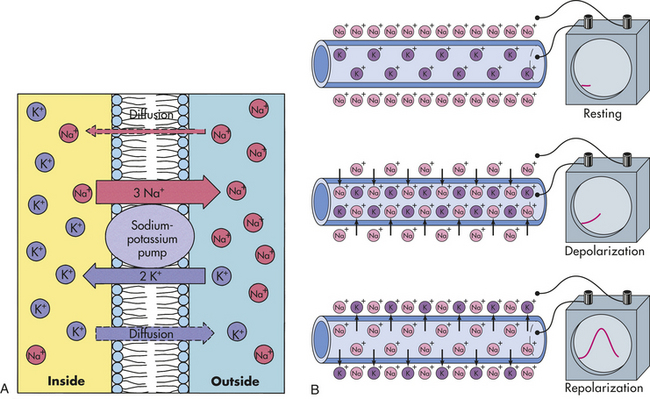
Figure 1-32 Sodium-potassium pump and propagation of an action potential. A, Concentration difference of Na+ and K+ intracellularly and extracellularly. The direction of active transport by the sodium-potassium pump is also shown. B, Top diagram represents the polarized state of a neuronal membrane when at rest. The lower diagrams represent changes in sodium and potassium membrane permeabilities with depolarization and repolarization. (From Thibodeau GA, Patton KT: Anatomy & physiology, ed 6, St Louis, 2007, Mosby.)
Nerve and muscle cells are excitable and can change their resting membrane potential in response to electrochemical stimuli. Changes in resting membrane potential convey messages from cell to cell. When a nerve or muscle cell receives a stimulus that exceeds the membrane threshold value, there is a rapid change in the resting membrane potential known as the action potential. The action potential carries signals along the nerve or muscle cell and conveys information from one cell to another. (Nerve impulses are described in Chapter 14.) When a resting cell is stimulated through voltage-regulated channels, the cell membranes become more permeable to sodium. There is a net movement of sodium into the cell, and the membrane potential decreases, or “moves forward,” from a negative value (in millivolts) to zero. This decrease is known as depolarization. The depolarized cell is more positively charged, and its polarity is neutralized.
To generate an action potential and the resulting depolarization, a critical value known as the threshold potential must be reached. Generally this occurs when the cell has depolarized by 15 to 20 millivolts. When the threshold is reached, the cell will continue to depolarize with no further stimulation. The sodium gates open, and sodium rushes into the cell, causing the membrane potential to reduce to zero and then become positive (depolarization). The rapid reversal in polarity results in the action potential.
During repolarization the negative polarity of the resting membrane potential is reestablished. As the voltage-gated sodium channels begin to close, voltage-gated potassium channels open. Membrane permeability to sodium decreases, and potassium permeability increases, with an outward movement of potassium ions. The sodium gates close, and with the outward movement of potassium, the membrane potential becomes more negative. The Na+-K+ pump then returns the membrane to the resting potential by pumping potassium back into the cell and sodium out of the cell.
During most of the action potential, the plasma membrane cannot respond to an additional stimulus. This time is known as the absolute refractory period and is related to changes in permeability to sodium. During the latter phase of the action potential, when permeability to potassium increases, a stronger-than-normal stimulus can evoke an action potential known as the relative refractory period.
When the membrane potential is more negative than normal, the cell is in a hyperpolarized (less excitable) state. A larger-than-normal stimulus is then required to reach the threshold potential and generate an action potential. When the membrane potential is more positive than normal, the cell is in a hypopolarized (more excitable than normal) state, and a smaller-than-normal stimulus is required to reach the threshold potential. Changes in the intracellular and extracellular concentration of ions or a change in membrane permeability can cause these alterations in membrane excitability.
CELLULAR REPRODUCTION: THE CELL CYCLE
Human cells are subject to wear and tear, and most do not last for the lifetime of the individual. In almost all tissues, new cells are created as fast as old ones die. Cellular reproduction is therefore necessary for the maintenance of life. Reproduction of gametes (sperm and egg cells) occurs through a process called meiosis, described in Chapter 4. The reproduction, or division, of other body cells (somatic cells) involves two sequential phases: mitosis, or nuclear division, and cytokinesis, or cytoplasmic division. These two phases occur in close succession, with cytokinesis beginning toward the end of mitosis. Before a cell can divide, however, it must double its mass and duplicate all its contents. Most of the work of preparing for division occurs during the growth phase, called interphase. The alternation between mitosis and interphase in all tissues with cellular turnover is known as the cell cycle.
Most of the early work on the cell cycle was limited to microscopic observation of mitosis and cytokinesis. Interphase was considered the “resting stage” of the cell. With recent technologic advances a considerable amount has been learned about the interphase part of the cell cycle. During interphase many important processes are taking place as the cell produces DNA, RNA, protein, lipids, and other substances, and each pair of chromosomes (paired organelles that carry genetic information) makes exact copies of themselves.
The four designated phases of the cell cycle are (1) the G1 phase (G = gap), which is the period between the M phase and the start of DNA synthesis; (2) the S phase (S = synthesis), in which DNA is synthesized in the cell nucleus; (3) the G2 phase, in which RNA and protein synthesis occurs, the period between the completion of DNA synthesis and the next phase (M); and (4) the M phase (M = mitosis), which includes both nuclear and cytoplasmic division (Figure 1-33).
Figure 1-33 The cell cycle. The cell cycle consists of gap 1 (or G1) (presynthesis), S (DNA synthesis), G2 (premitotic) and M (mitotic) phases. Quiescent cells (quiet or resting state) are in the G0 phase; however, most mature tissues have a combination of continuously dividing cells, terminally differentiated cells, stems cells, and some quiescent cells that infrequently enter the cell cycle. Continuously dividing cells replace those that are destroyed (e.g., epithelia of the oral cavity, skin). Quiescent or stable tissues exhibit a low level of replication; however, these cells can undergo rapid division in response to stimuli such as growth factors (e.g., EGF, TGF-α). Cyclins increase and activate cyclin-dependent protein kinase (CDK) complexes at the G1/S restriction point, causing phosphorylation (addition of phosphate group) of the molecular ON-OFF switch, the retinoblastoma susceptibility protein (RB). In its hypophosphorylated state, RB prevents cells from replicating by forming a tight inactive complex with the transcription factor E2F. Phosphorylation of RB eliminates the “brakes” to cell cycle progression and promotes cell replication. The orderly progression of cells through the phases of the cell cycle is regulated by cyclins, CDKs, and their inhibitors. Cyclin levels rise and fall (thus the name cyclin) during the cell cycle, periodically activating CDKs. Unless CDKs are bound to cyclins, they have no protein kinase activity. Cyclin-CDK complexes trigger cell cycle events. Each complex phosphorylates a different set of proteins that then promote advancement to the next phase (G1, S, G2, M, G0). After completion of the task, cyclin levels decline rapidly. The activity of cyclin-CDK complexes is regulated by CDK inhibitors including Cip/Kip and the 7NK4/ARF.
Phases of Mitosis and Cytokinesis
Interphase (the G1, S, and G2 phases) is the longest phase of the cell cycle. During interphase the chromatin consists of very long, slender rods that are jumbled together in the nucleus. Late in interphase, strands of chromatin (the substance that gives the nucleus its granular appearance) begin to coil, causing them to shorten and thicken.
The M phase of the cell cycle, mitosis and cytokinesis, begins with prophase, the first appearance of chromosomes. As the phase proceeds, each chromosome is seen as two identical halves called chromatids, which lie together and are attached at some point by a spindle attachment site called a centromere. (The two chromatids of each chromosome, which are genetically identical, are sometimes called sister chromatids.) The nuclear membrane, which surrounds the nucleus, disappears. Spindle fibers are microtubules formed in the cytoplasm. Spindle fibers radiate from two centrioles located at opposite poles of the cell. The role of the spindle fibers is to pull the chromosomes to opposite sides of the cell.
During metaphase, the next phase of mitosis and cytokinesis, the spindle fibers begin to pull the centromeres of the chromosomes. The centromeres become aligned in the middle of the spindle, which is called the equatorial plate (or metaphase plate) of the cell. In this stage chromosomes are easiest to observe microscopically because they are highly condensed and arranged in a relatively organized fashion in the two-dimensional equatorial plate.
Anaphase begins when the centromeres split and the sister chromatids are pulled apart. The spindle fibers shorten, causing the sister chromatids to be pulled, centromere first, toward opposite sides of the cell. When the sister chromatids are separated, each is considered to be a chromosome. Thus the cell has 92 chromosomes during this stage. By the end of anaphase, 46 chromosomes are lying at each side of the cell. Barring mitotic errors, each of the two groups of 46 chromosomes is identical to the original 46 chromosomes present at the start of the cell cycle.
During telophase, the final stage, a new nuclear membrane is formed around each group of 46 chromosomes, the spindle fibers disappear, and the chromosomes begin to uncoil. Cytokinesis causes the cytoplasm to divide into roughly equal parts during this phase. At the end of telophase, two identical diploid cells, called daughter cells, have been formed from the original cell.
Rates of Cellular Division
Although the complete cell cycle lasts 12 to 24 hours, about 1 hour is generally required for the four stages of mitosis and cytokinesis. All types of cells undergo mitosis during formation of the embryo, but many adult cells, such as nerve cells, lens cells of the eye, and muscle cells, lose their ability to replicate and divide. The cells of other tissues, particularly epithelial cells (e.g., of the intestine, lung, skin), divide continuously and rapidly, completing the entire cell cycle in less than 10 hours.
The difference between cells that divide slowly and cells that divide rapidly is the length of time spent in the G1 phase of the cell cycle. Some cells that divide very slowly remain in the G1 phase for days or even years. Once the S phase begins, however, progression through mitosis takes a relatively constant amount of time. Once a cell has progressed out of the G1 phase, there is no turning back; it is committed to completing the S, G2, and M phases. Times associated with the four successive phases differ.
The mechanisms that control cell division depend on “social control genes” and protein growth factors. Individual cells are members of a complex cellular society in which survival of the entire organism is key and not survival or proliferation of just the individual cells. To grow and divide, a cell must receive specific positive signals from other cells. Many of these signals are protein growth factors that act by overriding intracellular negative controls that block progress of the cell cycle.1
When a need arises for new cells, as in repair of injured cells, previously nondividing cells must be rapidly triggered to reenter the cell cycle. With continual wear and tear, the cell birth rate and the cell death rate must be kept in balance. Therefore, cell-division controls must govern this balance. Protein growth factors governing the proliferation of different cell types and genes involved in the social control of cell division are currently being identified.1
The best model for understanding disruption of cell division and study of these so-called social control genes is tumor biology. Current emphasis in locating and identifying these genes is to study tumor cells that have presumably originated because of mutations to these genes, or proto-oncogenes. Proto-oncogenes are thought to encode key components of the normal system of social controls of cell division1; that is, the mechanisms by which signals from a cell’s neighbors can impel it to divide, differentiate, or die. Some proto-oncogenes code for growth factors, some for growth factor receptors, some for intracellular regulatory proteins that are involved in cell adhesion, and some for proteins that help relay signals for cell division to the cell nucleus.1 Although more than 50 proto-oncogenes have been identified, many more are yet to be discovered (see Chapter 11).
Growth Factors
Growth factors, also called cytokines, are peptides that transmit signals within and between cells. They have a major role in the regulation of tissue growth and development (Table 1-6). Having nutrients is not enough for a cell to proliferate; it must also receive stimulatory chemical signals (growth factors) from other cells, usually its neighbors. These signals act to overcome intracellular braking mechanisms that tend to restrain cell growth and block progress through the cell cycle.
Table 1-6
Examples of Growth Factors and Their Actions
| Growth Factor | Physiologic Actions |
| Platelet-derived growth factor (PDGF) | Stimulates proliferation of connective tissue cells and neuroglial cells |
| Epidermal growth factor (EGF) | Stimulates proliferation of epidermal cells and other cell types |
| Insulin-like growth factor I (IGF-I) | Collaborates with PDGF and EGF; stimulates proliferation of fat cells and connective tissue cells |
| Insulin-like growth factor II (IGF-II) | Collaborates with PDGF and EGF; stimulates proliferation of fat cells and connective tissue cells |
| Transforming growth factor β (TGF-β) | Stimulates or inhibits response of most cells to other growth factors; regulates differentiation of some cell types (e.g., cartilage) |
| Fibroblast growth factor (FGF) | Stimulates proliferation of fibroblasts, endothelial cells, myoblasts, and other cell types |
| Interleukin-2 (IL-2) | Stimulates proliferation of T lymphocytes |
| Nerve growth factor (NGF) | Promotes axon growth and survival of sympathetic and some sensory and CNS neurons |
| Hematopoietic cell growth factors (IL-3, GM-CSF, M-CSF, G-CSF, erythropoietin) | See Chapter 25 |
CNS, Central nervous system; CSF, colony-stimulating factor; G, granulocyte; GM, granulocyte-macrophage; M, macrophage.
Different types of cells require different factors; for example, platelet-derived growth factor (PDGF) stimulates the production of connective tissue cells. Table 1-6 summarizes the most significant growth factors. Cells that respond to a particular growth factor have specific receptors for the growth factor in their plasma membrane. Recent evidence shows that some growth factors are also regulators of other cell processes, such as cellular differentiation. In addition to growth factors that stimulate cellular processes, there are factors that inhibit functions; these factors are not well understood. Cells that are starved of growth factors come to a halt after mitosis and enter the arrested, or G0, state of the cell cycle1 (see p. 33 for cell cycle).
TISSUES
The body is made up of four levels of organization: cells, tissues, organs, and systems. Cells of common structure and function are organized into tissues, of which there are four primary types: muscle, neural, epithelial, and connective tissue.
Tissue Formation
To form tissues cells must exhibit intercellular recognition and adhesion. Specialized cells are thought to form a tissue in one of two ways. First and simplest is mitosis of one or more founder cells (the most basic precursor cell). Founder cells are prevented from “wandering away” by macromolecules in the extracellular matrix and by adherence to one another at specialized junctions on their plasma membranes. Mitosis of founder cells forms, for example, epithelial cell sheets (Figure 1-34).
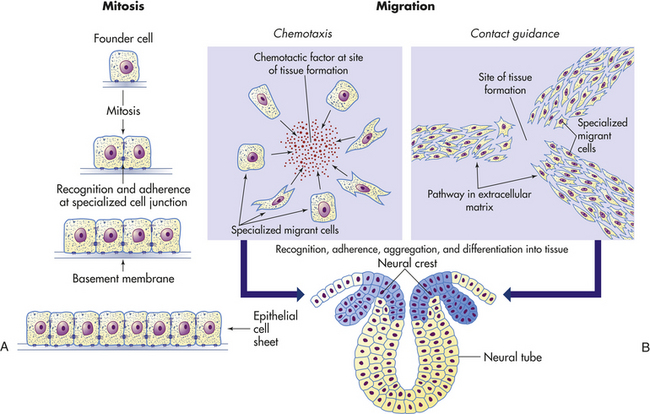
Figure 1-34 Tissue formation by mitosis and migration. A, Tissue formation by mitosis. Founder cells are kept in place by extracellular matrix and recognition and adherence at cell junctions. B, Tissue formation by migration. Specialized cells are attracted to the site of tissue formation by chemotaxis or contact guidance; then they aggregate and differentiate into organized tissue.
The second way in which specialized cells form tissues involves their migration to and subsequent assembly at the site of tissue formation. During embryonic development, for example, cells from the neural crest migrate to several different regions, where they differentiate and assemble into a variety of tissues, including those of the peripheral nervous system. Migrant cells are thought to arrive at the site of tissue formation through chemotaxis or contact guidance. Chemotaxis is movement along a chemical gradient caused by chemical attraction (see Chapter 6). Cells at the migrant cells’ destination secrete a chemical, called chemotactic factor, that attracts specific migrant cells. Contact guidance is movement along a pathway, or “pavement,” in the extracellular matrix.1
Tissues are not randomly arranged into organs. No matter how tissue is formed, staying together in groups means that cells must recognize each other and remain distinct from the cells of surrounding tissues. Little is known about the mechanisms involved in these processes.
Types of Tissues
Epithelial tissue covers most internal and external surfaces of the body. Epithelial cells are closely joined and are attached to a basement membrane or lamina (extracellular matrix), which provides a supporting layer and separates the epithelium from underlying connective tissue (see Figure 1-14). Because of its variety of locations, epithelial tissue has several diverse functions, including protection, absorption, secretion, and excretion. For example, the epidermis provides a protective barrier between the host and the outside environment, and the linings of the internal body organs help absorb substances into the body, excrete waste products, and secrete substances into body cavities.
Epithelial cell surfaces differ according to their location and function. Epithelial cells that line body cavities and blood vessels are smooth, whereas other epithelial cells have tiny cytoplasmic projections called microvilli on their free surfaces. Microvilli considerably increase a cell’s surface area and are found on cells whose main functions are absorption and secretion, such as the epithelial cells lining the digestive tract. Cilia, which are hairlike projections that propel mucus, pus, and dust particles out of the body, characterize cells lining the respiratory passages.
Epithelial tissue is classified in two ways: (1) according to the number and arrangement of cell layers, and (2) according to cell shape. Epithelium that is formed by a single layer of cells, all of which are in contact with the basement membrane, is called simple epithelium. Stratified epithelium has two or more layers of cells, and only the deepest layer is in contact with the basement membrane. Tissue that appears to consist of several cellular layers but is actually a single layer with all cells contacting the basement membrane is called pseudostratified epithelium.
Three basic cell shapes are found in epithelium: squamous, cuboidal, and columnar. Squamous cells are flat and thin; cuboidal cells are as high as they are wide and thus appear square in vertical sections; and columnar cells are taller than they are wide and appear rectangular in vertical sections. Overall classifications of epithelial tissue, which take into account both the number of cell layers and cell shape, are summarized in Table 1-7.
Connective Tissue
Connective tissue varies considerably in structure and function but is most common as the framework on which epithelial cells cluster to form organs. Other functions include binding various tissues and organs together, supporting them in their locations, and serving as storage sites for excess nutrients.
In contrast to epithelial tissue, connective tissue is characterized by an abundant extracellular matrix that surrounds few cells. The extracellular matrix is composed of ground substance and fibers. Ground substance is a homogeneous mass that varies in consistency from fluid to semisolid gel. Fibers are produced by connective tissue cells (fibroblasts) found within the ground substance. The three types of fibers are collagenous (white), elastic (yellow), and reticular. Collagenous fibers are formed of bundles of smaller fibers appearing as wavy bands under the microscope. These fibers are composed of the protein collagen and are strong and inelastic. (Collagen synthesis by fibroblasts is described with respect to tissue repair in Chapter 6.) Elastic fibers are long, branching fibers composed of a protein called elastin that enables the fibers to return to their original length after stretching. Elastin occurs not only as fibers but also as membranes, particularly the membranes of blood vessels. Reticular fibers are thin, short, branching fibers that form an inelastic network made from a collagen-like protein called reticulum. Reticular fibers form the internal framework (stroma) to which the epithelial cells of glands are attached. They are found in loose connective tissue, generally in bone marrow and in the parenchyma (i.e., the essential substance of an organ rather than its framework) of the liver, spleen, and lymph nodes.
Connective tissues are classified according to the consistency (e.g., loose, dense) of the ground substance and the type and organization of the fibers within it. Table 1-8 summarizes the characteristics of connective tissues.
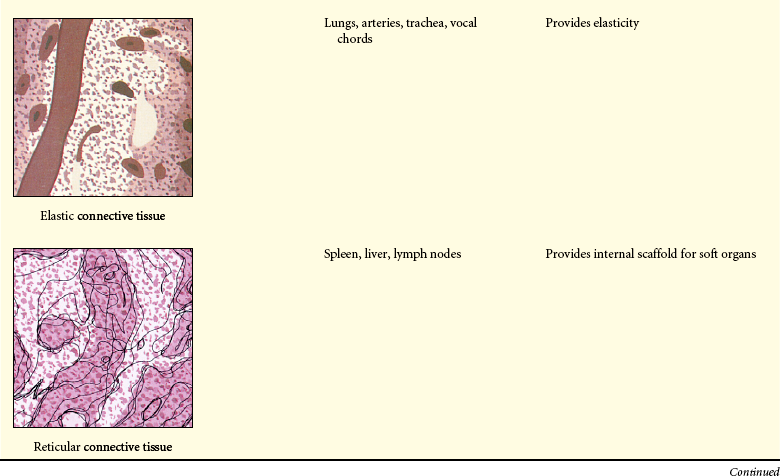
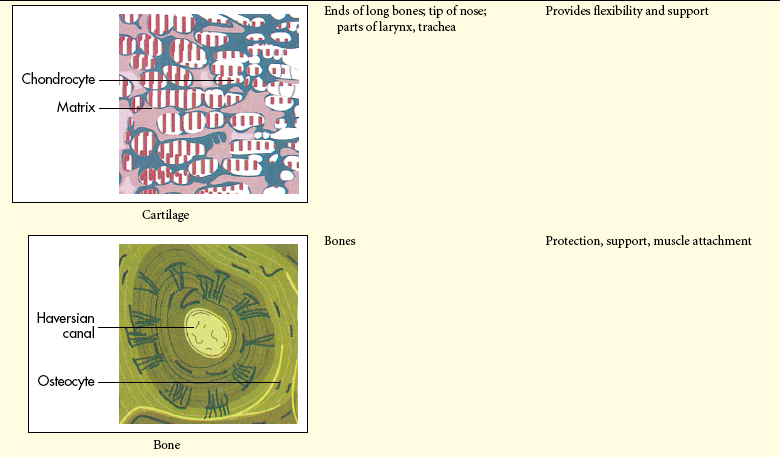
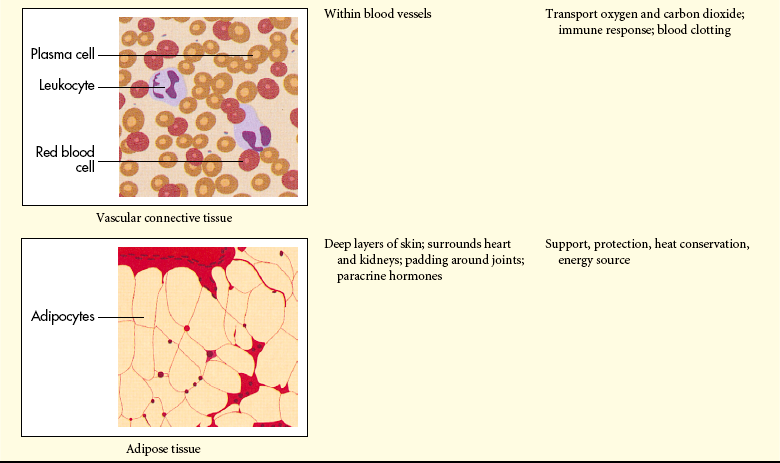
Muscle Tissue
Muscle tissue is composed of long, thin cells or fibers called myocytes. Myocytes are highly contractile. The three types of muscle tissues are skeletal, cardiac, and smooth (Table 1-9). (Muscles are discussed in detail in Chapter 41.)
Neural Tissue
Neural tissue is composed of highly specialized cells called neurons, which receive and transmit electrical impulses very rapidly across junctions called synapses. Synapses are points of functional contact between neurons. At synapses, impulses pass from neuron to neuron or from a neuron to a muscle cell as chemical messengers called neurotransmitters are released (see Chapter 14). The total number of neurons is fixed at birth, and replacement is impossible thereafter.
Different types of neurons have special characteristics that depend on their distribution and function within the nervous system. All neurons, however, are composed of the following parts: (1) a cell body, (2) a single axon, and (3) one or more dendrites (see Figure 14-1 on p. 444). The cell body contains special cytoplasmic structures, as well as microtubules, actin filaments, Golgi complex, lysosomes, and lipofuscin. The axons and dendrites can be very long. Generally, the axon conducts nerve impulses away from the cell body, and dendrites conduct nerve impulses toward the cell body. (Neuronal transmission is discussed in Chapter 14.)
Absolute refractory period 33
Actin filament (microfilament) 9
Action potential 32
Active mediated transport (active transport) 26, 29
Active transport 29
Amphipathic molecule 11
Anabolism 21
Anaerobic glycolysis 24
Anaphase 35
Anions 26
Antiport 29
Arrested (G0) state 35
Autocrine signaling 18
Autodigestion 6
Autolysosome (autophagosome) 6
Autophagy 6
Basement membrane 15
Catabolism 21
Cation 26
Caveolae 32
Cell adhesion molecule (CAM) 12
Cell cycle 34
Cell junction 16
Cell-to-cell adhesion 15
Cellular metabolism 21
Cellular receptor 13
Centriole 9
Centromere 34
Chemical synapse 18
Chemotaxis 36
Chromatid 34
Chromatin 34
Chromosome 34
Cilia 36
Cisternae 5
Citric acid cycle (Krebs cycle, tricarboxylic acid cycle) 23
Clathrin 5
Coated pit 31
Collagen 16
Collagenous fiber 36
Columnar cell 36
Competitive inhibitor 28
Concentration gradient 26
Connexon 18
Contact guidance 36
Cristae 7
Cuboidal cell 36
Cyclic adenosine monophosphate (cyclic AMP, cAMP) 20
Cyclic guanosine monophosphate (cyclic GMP, cGMP) 20
Cytochrome 24
Cytokinesis 33
Cytoplasm 2
Cytoplasmic matrix 4
Cytoskeleton 8
Cytosol 4
Depolarization 33
Desmosome 16
Differentiation 2
Diffusion 26
Digestion 23
Effective osmolality 27
Elastic fiber 38
Elastin 16
Electrolyte 26
Electron-transport chain 24
Endocytosis 30
Endoplasmic reticulum 5
Equatorial plate (metaphase plate) 35
Eukaryote 2
Exocytosis 31
Extracellular matrix (basement membrane) 15
Fibroblast 16
Fibronectin 16
Filtration 26
First messenger 20
Founder cell 36
G protein 20
Gap junction 18
Gating 18
Glycolysis 23
Glycoprotein 12
Golgi complex (Golgi apparatus) 5
Ground substance 36
Growth factor 35
Guanosine diphosphate (GDP) 20
Guanosine triphosphate (GTP) 20
Homeostasis 18
Hormonal signaling 18
Hydrolases 5
Hydrophilic 11
Hydrophobic 11
Hydrostatic pressure 26
Hypertonic solution 28
Inner membrane 7
Integral membrane protein 11
Intermediary metabolism 8
Interphase 34
Ion 26
Isotonic solution 28
Junctional complex 16
Ligand 13
Lysosome 5
Macromolecule 15
Mediated transport 28
Metabolic pathway 21
Metaphase 35
Microdomain 32
Microtubule 9
Microvilli 36
Mitochondria 7
Mitosis 33
Neurohormonal signaling 18
Neurotransmitter 18
Nuclear envelope 2
Nucleolus 2
Nucleus 2
Oncotic pressure (colloid osmotic pressure) 28
Osmolality 27
Osmolarity 27
Osmosis 27
Osmotic pressure 27
Outer membrane 7
Oxidation 23
Oxidative cellular metabolism 23
Oxidative phosphorylation 24
Paracrine signaling 18
Parenchyma 38
Passive mediated transport (facilitated diffusion) 29
Passive transport 25
Peripheral membrane protein 11
Peroxisome (microbody) 7
Phagocytosis 31
Pinocytosis 31
Plasma membrane receptor 15
Platelet-derived growth factor (PDGF) 35
Polarity 26
Polyribosome 8
Potocytosis 32
Primary lysosome 5
Prokaryote 2
Prophase 34
Proteolytic cascade 12
Pseudostratified epithelium 36
Raft 13
Receptor-mediated endocytosis (ligand internalization) 31
Relative refractory period 33
Repolarization 33
Residual body 6
Resting membrane potential 32
Reticular fiber 38
Ribosomal protein synthesis 8
Ribosome 5
Rough endoplasmic reticulum 5
Second messenger 20
Secondary lysosome (heterophagosome) 6
Secretory vesicle 5
Signal transduction 19
Simple epithelium 36
Smooth endoplasmic reticulum 5
Solute 26
Spindle fiber 35
Squamous cell 36
Stratified epithelium 36
Substrate 21
Symport 29
Synapses 41
Telophase 35
Threshold potential 33
Tight junction 18
Tissue 35
Tonicity 28
Transfer reaction 24
Transmembrane protein 11
Transport protein 28
Uniport 29
Vault 7
REFERENCES
1. Alberts, B., et al. Molecular biology of the cell, ed 5. New York: Garland, 2008.
2. van Zon, A., et al. Vault mobility depends in part on microtubules and vaults can be recruited to the nuclear envelope. Exp Cell Res. 2006;312(3):245–255.
3. Steiner, E., et al. Cellular functions of vaults and their involvement in multidrug resistance. Cult Drug Targets. 2006;7(8):923–934. [review].
4. Anderson, D.H., et al. Draft crystal structure of the vault shell at 9-A resolution. Plos Biol. 2007;5(11):e318.
5. Mooney, D.J., Mikos, A.G. Growing new organs. Sci Am. 1999;280(4):60–65.
6. Fasshauer, M., Iwig, M., Glaesser, D. Synthesis of proto-oncogene proteins and cyclins depends on intact microfilaments. Eur J Cell Biol. 1998;77(3):188–195.
7. Lofthouse, R.A., et al. Identification of caveolae and detection of caveolin in normal human osteoblasts. J Bone Joint Surg Br. 2001;83(1):124–129.
8. Southan, C. Drug discovery. Today. 2001;6:681–688.
9. Amour, A., et al. General considerations for proteolytic cascades. Biochem Soc Trans. 2004;32(Pt 1):15–16.
10. Logue, S.E., Martin, S.J. Caspase activation cascades in apoptosis. Biochem Soc Trans. 2008;36(Pt 1):1–9.
11. Chang, H.Y., Yang, X. Proteases for cell suicide: functions and regulations of caspases. Microbiol Mol Biol Rev. 2000;64(4):821–846.
12. Catt, K.J., et al. Hormonal regulation of peptide receptors and target cell responses. Nature. 1979;280(5718):109–116.
13. LaPorte, S.L., et al. Molecular and structural basis of cytokine receptor pleiotropy in the interleukin-4/13 system. Cell. 2008;132:259–272.
14. Kumar, V., Abbas, A., Fausto, N. Robbins and Cotran pathologic basis of disease, ed 7. Philadelphia: Saunders; 2005.
15. Alberts, B., et al. Essential cell biology, ed 5. New York: Garland, 1998.
16. Baserga, R., Morrione, A. Differentiation and malignant transformation: two roads diverge in a wood. J Cell Biochem Suppl. 1999;32-33:68–75.
17. Singh, A.B., Harris, R.C. Autocrine, paracrine, and juxtacrine signaling by EGFR ligands. Cell Signal. 2005;17(10):1183–1193.
18. Scott, J.D., Pawson, T. Cell communication: the inside story. Sci Am. 2000;282(6):72–79.
19. He, B., et al. Exo 70 interacts with phospholipids and mediates the targeting of the exocyst to the plasma membrane. EMBO. 2007;26(18):4053–4065.
20. Schwencke, C., et al. Caveolae and cavolin in transmembrane signaling: implications for human disease. Cardiovasc Res. 2006;70(1):42–49.
21. Levin, E.R., Pietras, R.J. Estrogen receptors outside the nucleus in breast cancer. Breast Cancer Res Treat. 2008;108(3):351–361.
22. Li, L., Haynes, M.P., Bender, J.R. Plasma membrane localization and function of the estrogen receptor alpha variant (ER46) in human endothelial cells. Proc Natl Acad Sci U S A. 2003;100(8):4807–4812.
23. Mineo, C., Shaul, P.W. Circulating cardiovascular disease risk factors and signaling in endothelial cell caveolae. Cardiovasc Res. 2006;70(1):31–41.