1 Overview of the Circulation and Blood
1. Describe the general structure of the cardiovascular system.
2. Compare the compositions and functions of the blood vessels.
3. Compare the relationship of the vascular cross-sectional area to the velocity of blood flow in the various vascular segments.
4. Indicate the pressure changes and pathways of blood flow throughout the vasculature.
5. Describe the constituents of the blood and explain the functions of the cellular elements of blood.
6. Know the importance of blood group matching before blood transfusions.
The circulatory, endocrine, and nervous systems constitute the principal coordinating and integrating systems of the body. Whereas the nervous system is primarily concerned with communication and the endocrine glands with regulation of certain body functions, the circulatory system serves to transport and distribute essential substances to the tissues and to remove metabolic by-products. The circulatory system also shares in such homeostatic mechanisms as regulation of body temperature, humoral communication throughout the body, and adjustments of O2 and nutrient supply in different physiologic states.
The Circulatory System
The cardiovascular system accomplishes these functions with a pump (see Chapter 4), a series of distributing and collecting tubes (see Chapter 7), and an extensive system of thin vessels that permit rapid exchange between the tissues and the vascular channels (see Chapter 8). The primary purpose of this text is to discuss the function of the components of the vascular system and the control mechanisms (with their checks and balances) that are responsible for alteration of blood distribution necessary to meet the changing requirements of different tissues in response to a wide spectrum of physiological (see Chapter 9) and pathological (see Chapter 13) conditions.
Before one considers the function of the parts of the circulatory system in detail, it is useful to consider it as a whole in a purely descriptive sense (Figure 1-1). The heart consists of two pumps in series: the right ventricle to propel blood through the lungs for exchange of O2 and CO2 (the pulmonary circulation) and the left ventricle to propel blood to all other tissues of the body (the systemic circulation). The total flow of blood out of the left ventricle is known as the cardiac output (CO). The rhythmic contraction of the heart is an intrinsic property of the heart whose sino-atrial node pacemaker generates action potentials spontaneously (see Chapter 3). These action potentials are propagated in an orderly manner through the organ to trigger contraction and to produce the currents detected in the electrocardiogram (see Chapter 3).
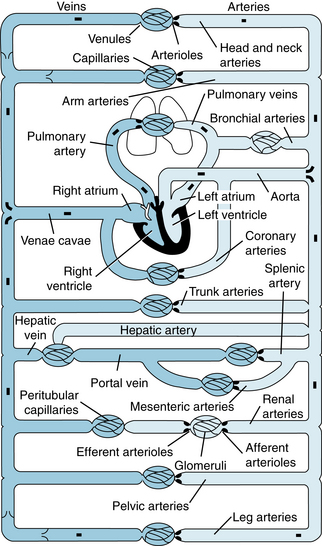
FIGURE 1-1 Schematic diagram of the parallel and series arrangement of the vessels composing the circulatory system. The capillary beds are represented by thin lines connecting the arteries (on the right) with the veins (on the left). The crescent-shaped thickenings proximal to the capillary beds represent the arterioles (resistance vessels).
(Redrawn from Green HD: In Glasser O, editor: Medical physics, vol 1, Chicago, 1944, Mosby-Year Book.)
Unidirectional flow through the heart is achieved by the appropriate arrangement of effective flap valves. Although the cardiac output is intermittent, continuous flow to the periphery occurs by distention of the aorta and its branches during ventricular contraction (systole) and elastic recoil of the walls of the large arteries that propel the blood forward during ventricular relaxation (diastole). Blood moves rapidly through the aorta and its arterial branches (see Chapter 7). The branches become narrower and their walls become thinner and change histologically toward the periphery. From the aorta, a predominantly elastic structure, the peripheral arteries become more muscular until the muscular layer predominates at the arterioles (Figure 1-2).
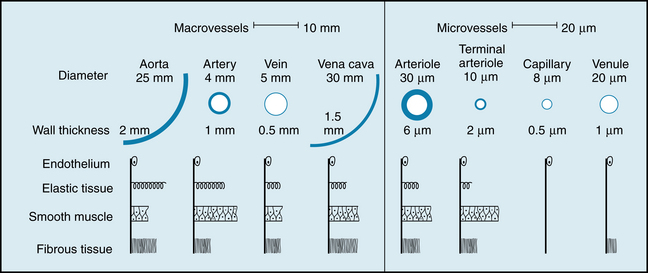
FIGURE 1-2 Internal diameter, wall thickness, and relative amounts of the principal components of the vessel walls of the various blood vessels that compose the circulatory system. Cross-sections of the vessels are not drawn to scale because of the huge range from aorta and venae cavae to capillary.
(Redrawn from Burton AC: Relation of structure to function of the tissues of the wall of blood vessels. Physiol Rev 34:619, 1954.)
In the large arteries, frictional resistance is relatively small, and mean pressure throughout the system of large arteries is only slightly less than in the aorta. The small arteries and arterioles serve to regulate flow to individual tissues by varying their resistance to flow. The small arteries offer moderate resistance to blood flow, and this resistance reaches a maximal level in the arterioles, sometimes referred to as the stopcocks of the vascular system. Hence the pressure drop is significant and is greatest in the small arteries and in the arterioles (Figure 1-3). Adjustments in the degree of contraction of the circular muscle of these small vessels permit regulation of tissue blood flow and aid in the control of arterial blood pressure (see Chapter 9).
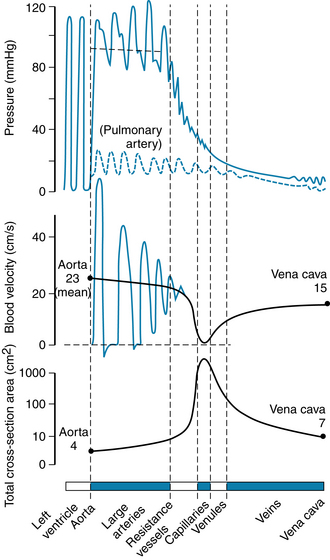
FIGURE 1-3 Phasic pressure, velocity of flow, and cross-sectional area of the systemic circulation. The important features are the major pressure drop across the small arteries and arterioles, the inverse relationship between blood flow velocity and cross-sectional area, and the maximal cross-sectional area and minimal flow rate in the capillaries.
(From Levick JR: An introduction to cardiovascular physiology, ed 5, London, 2010, Hodder Arnold.)
In addition to a sharp reduction in pressure across the arterioles, there is also a change from pulsatile to steady flow as pressure continues to decline from the arterial to the venous end of the capillaries (see Figure 1-3). The pulsatile arterial blood flow, caused by the phasic cardiac ejection, is damped at the capillaries by the combination of distensibility of the large arteries and frictional resistance in the arterioles.
In a patient with hyperthyroidism (Graves disease), the basal metabolism is elevated and is often associated with arteriolar vasodilation. This reduction in arteriolar resistance diminishes the dampening effect on the pulsatile arterial pressure and is manifested as pulsatile flow in the capillaries, as observed in the fingernail beds of patients with this ailment.
Many capillaries arise from each arteriole to form the microcirculation (see Chapter 8), so that the total cross-sectional area of the capillary bed is very large, despite the fact that the cross-sectional area of each capillary is less than that of each arteriole. As a result, blood flow velocity becomes quite slow in the capillaries (see Figure 1-3), analogous to the decrease in velocity of flow seen at the wide regions of a river. Conditions in the capillaries are ideal for the exchange of diffusible substances between blood and tissue because the capillaries are short tubes whose walls are only one cell thick and because flow velocity is low.
On its return to the heart from the capillaries, blood passes through venules and then through veins of increasing size with a progressive decrease in pressure until the blood reaches the vena cava (see Figure 1-3). As the heart is approached, the number of veins decreases, the thickness and composition of the vein walls change (see Figure 1-2), the total cross-sectional area of the venous channels diminishes, and the velocity of blood flow increases (see Figure 1-3). Note that the velocity of blood flow and the cross-sectional area at each level of the vasculature are essentially mirror images of each other (see Figure 1-3).
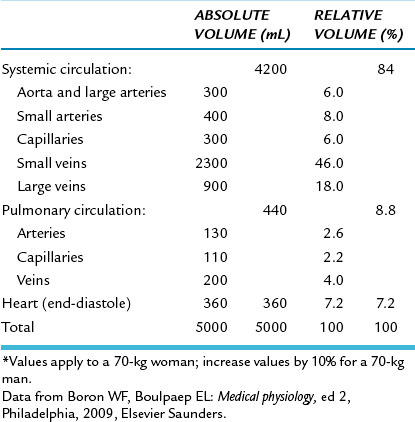
Data indicate that between the aorta and the capillaries the total cross-sectional area increases about 500-fold (see Figure 1-3). The volume of blood in the systemic vascular system (Table 1-1) is greatest in the veins and small veins (64%). Of the total blood volume, only about 6% of it is in the capillaries and 14% in the aorta, arteries, and arterioles. In contrast, blood volume in the pulmonary vascular bed is about equal between arteries and capillaries; venous vessels display a slightly larger percentage of pulmonary blood volume. The cross-sectional area of the venae cavae is larger than that of the aorta. Therefore, the velocity of flow is slower in the venae cavae than that in the aorta (see Figure 1-3).
Blood entering the right ventricle via the right atrium is pumped through the pulmonary arterial system at a mean pressure about one seventh that in the systemic arteries. The blood then passes through the lung capillaries, where CO2 is released and O2 taken up. The O2-rich blood returns via the four pulmonary veins to the left atrium and ventricle to complete the cycle. Thus, in the normal intact circulation, the total volume of blood is constant, and an increase in the volume of blood in one area must be accompanied by a decrease in another. However, the distribution of the circulating blood to the different body organs is determined by the output of the left ventricle and by the contractile state of the arterioles (resistance vessels) of these organs (see Chapters 9 and 10). In turn, the cardiac output is controlled by the rate of heartbeat, cardiac contractility, venous return, and arterial resistance. The circulatory system is composed of conduits arranged in series and in parallel (see Figure 1-1).
It is evident that the systemic and pulmonary vascular systems are composed of many blood vessels arranged in series and parallel, with respect to blood flow. The total resistance to blood flow of the systemic blood vessels is known as the total peripheral resistance (TPR), and the total resistance of the pulmonary vessels is known as the total pulmonary resistance. Total peripheral resistance and cardiac output determine the mean pressure in the large arteries, though the hydraulic resistance equation (see Chapter 7).
The main function of the circulating blood is to carry O2 and nutrients to the various tissues in the body, and to remove CO2 and waste products from those tissues. Furthermore, blood transports other substances, such as hormones, white blood cells, and platelets, from their sites of production to their sites of action. Blood also aids in the distribution of fluids, solutes, and heat. Hence, blood contributes to homeostasis, the maintenance of a constant internal environment.
A fundamental characteristic of normal operation of the cardiovascular system is the maintenance of a relatively constant mean (average) blood pressure within the large arteries. The difference between mean arterial pressure (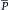 a) and the pressure in the right atrium (Pra) provides the driving force for flow through the resistance (R) of blood vessels of the individual tissues. Thus, when the circulatory system is in steady-state, total flow of blood from the heart (cardiac output, CO) equals total flow of blood returning to the heart. The relation among these variables is described in the following hydraulic equation:
a) and the pressure in the right atrium (Pra) provides the driving force for flow through the resistance (R) of blood vessels of the individual tissues. Thus, when the circulatory system is in steady-state, total flow of blood from the heart (cardiac output, CO) equals total flow of blood returning to the heart. The relation among these variables is described in the following hydraulic equation:
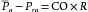
The cardiovascular system, together with neural, renal, and endocrine systems, maintains a at a relatively constant level, despite the large variations in cardiac output and peripheral resistance that are required in daily life. If the a is maintained at its normal level under all circumstances, then each individual tissue will be able to obtain the necessary blood flow required to sustain its functions. Because blood flow to the brain and the heart cannot be interrupted for even a few seconds without endangering life, maintenance of the a is a critical function of the cardiovascular system.
Blood
Blood consists of red blood cells, white blood cells, and platelets suspended in a complex solution (plasma) of various salts, proteins, carbohydrates, lipids, and gases. The circulating blood volume accounts for about 7% of the body weight. Approximately 55% of the blood is plasma; the protein content is 7 g/dL (about 4 g/dL of albumin and 3 g/dL of plasma globulins).
Erythrocytes
The erythrocytes (red blood cells) are flexible, biconcave disks that transport oxygen to the body tissues (Figure 1-4). Mammalian erythrocytes are unusual in that they lack a nucleus. The average erythrocyte is 7 µm in diameter, and these cells arise from pluripotential stem cells in the bone marrow. All of the cells in the circulating blood are derived from these stem cells. Most of these immature cells develop into various forms of mature cells, such as erythrocytes, monocytes, megakaryocytes, and lymphocytes. The erythrocytes lose their nuclei before they enter the circulation, and their average life span is 120 days. Approximately 5 million erythrocytes are present per microliter of blood. However, a small fraction of the pluripotential stem cells remains in the undifferentiated state.
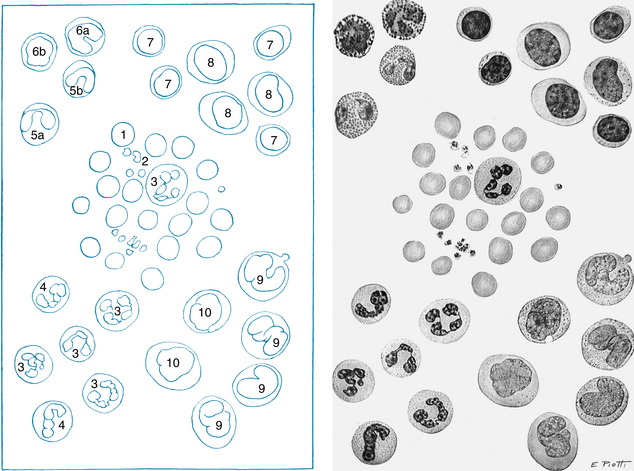
FIGURE 1-4 The morphology of blood cells. 1, Normal red blood cell; 2, platelet; 3, neutrophil; 4, neutrophil, band form; 5a, eosinophil, two lobes; 5b, eosinophil, band form; 6a, basophil, band form; 6b, metamyelocyte, basophilic; 7, lymphocyte, small; 8, lymphocyte, large; 9, monocyte, mature; 10, monocyte, young.
(From Daland GA: A color atlas of morphologic hematology, Cambridge, MA, 1951, Harvard University Press.)
Hemoglobin (about 15 g/dL of blood) is the main protein in the erythrocytes. Hemoglobin consists of heme, an iron-containing tetrapyrrole. Heme is linked to globin, a protein composed of four polypeptide chains (two α and two β chains in the normal adult). The iron moiety of hemoglobin binds loosely and reversibly to O2 to form oxyhemoglobin. The affinity of hemoglobin for O2 is a steep function of the partial pressure of O2 (Po2) at Po2 less than 60 mm Hg (Figure 1-5). This allows ready diffusion of O2 from hemoglobin to tissue. The binding of O2 to hemoglobin is affected by pH, temperature, and 2,3-diphosphoglycerate concentration. These factors affect O2 transport particularly at Po2 less than 60 mm Hg.
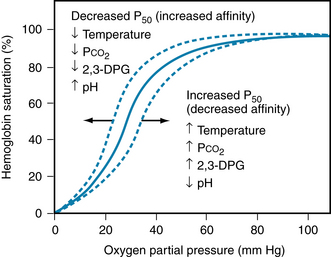
FIGURE 1-5 Oxyhemoglobin dissociation curve showing the saturation of hemoglobin as a function of the partial pressure of O2 (Po2) in the blood. Oxygenation of hemoglobin at a given Po2 is affected by temperature and the blood concentration of metabolites, CO2, 2,3-diphosphoglyerate (2,3-DPG) and H+. P50, the partial pressure where hemoglobin is 50% saturated with O2.
(From Koeppen BM, Stanton BA: Berne and Levy physiology, ed 6, Philadelphia, 2008, Mosby Elsevier.)
Changes in the polypeptide subunits of globin affect the affinity of hemoglobin for O2. For example, fetal hemoglobin has two γ chains instead of two β chains. This substitution increases its affinity for O2. Changes in the polypeptide subunits of globin may induce certain serious diseases, such as sickle cell anemia and erythroblastosis fetalis (Figure 1-6). Sickle cell anemia is a disorder associated with the presence of hemoglobin S, which is an abnormal form of hemoglobin in the erythrocytes. Many of the erythrocytes in the bloodstream of patients with sickle cell anemia have a sickle-like shape (Figure 1-6). Consequently, many of the abnormal cells cannot pass through the capillaries and, therefore, cannot deliver adequate O2 and nutrients to the local tissues. Thalassemia is also a genetic disorder of the globin genes; α and β forms exist. In either case, the disorder leads ultimately to a microcytic (small cell), hypochromic (inadequate quantity of hemoglobin) anemia (upper central panel of Figure 1-6).
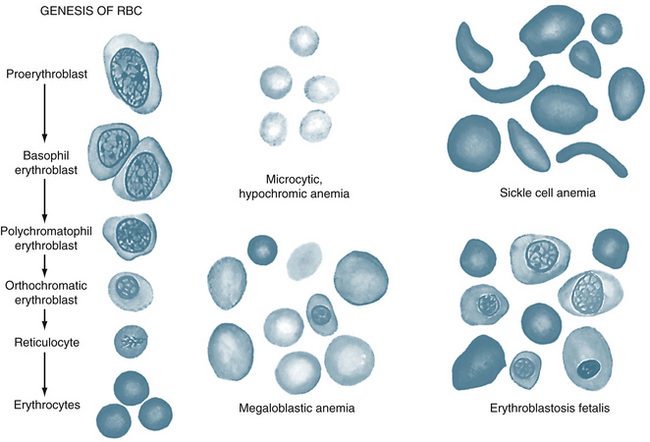
FIGURE 1-6 Genesis of red blood cells (RBCs), and red blood cells in different types of anemias.
(From Guyton AC, Hall JE: Textbook of medical physiology, ed 10, Philadelphia, 2006, WB Saunders.)
The number of circulating red cells normally remains fairly constant. The production of erythrocytes (erythropoiesis) is regulated by the glycoprotein erythropoietin, which is secreted mainly by the kidneys. Erythropoietin enhances erythrocyte production by accelerating the differentiation of stem cells in the bone marrow. This substance is often used clinically to increase red blood cell production in anemic patients.
Leukocytes
There are normally 4000 to 10,000 leukocytes (white blood cells) per microliter of blood. Leukocytes include granulocytes (65%), lymphocytes (30%), and monocytes (5%). Of the granulocytes, about 95% are neutrophils, 4% are eosinophils, and 1% are basophils. White blood cells originate from the primitive stem cells in the bone marrow. After birth, granulocytes and monocytes in humans continue to originate in the bone marrow, whereas lymphocytes originate in the lymph nodes, spleen, and thymus.
Anemia and chronic hypoxia are prevalent in people who live at high altitudes, and such conditions tend to stimulate erythrocyte production and can produce polycythemia (an increased number of red blood cells). When the hypoxic stimulus is removed in subjects with altitude polycythemia, the high erythrocyte concentration in the blood inhibits erythropoiesis. The red blood cell count is also greatly increased in polycythemia vera, a disease of unknown cause. The elevated erythrocyte concentration increases blood viscosity, often enough that blood flow to vital tissues becomes impaired.
Granulocytes and monocytes are motile, nucleated cells that contain lysosomes that have enzymes capable of digesting foreign material such as microorganisms, damaged cells, and cellular debris. Thus leukocytes constitute a major defense mechanism against infections. Microorganisms or the products of cell destruction release chemotactic substances that attract granulocytes and monocytes. When migrating leukocytes reach the foreign agents, they engulf them (phagocytosis) and then destroy them through the action of enzymes that form O2-derived free radicals and hydrogen peroxide.
Lymphocytes
Lymphocytes vary in size and have large nuclei. Most lymphocytes lack cytoplasmic granules (see Figure 1-5). The two main types of lymphocytes are B lymphocytes, which are responsible for humoral immunity, and T lymphocytes, which are responsible for cell-mediated immunity. When lymphocytes are stimulated by an antigen (a foreign protein on the surface of a microorganism or allergen), the B lymphocytes are transformed into plasma cells, which synthesize and release antibodies (gamma globulins). Antibodies are carried by the bloodstream to a site of infection, where they “tag” foreign invaders for destruction by other components of the immune system.
Blood Is Divided into Groups by Antigens Located on Erythrocytes
Four principal blood groups, designated O, A, B, and AB, prevail in human subjects. Each group is identified by the type of antigen that is present on the erythrocyte. People with type A blood have A antigens; those with type B blood have B antigens; those with type AB have both A and B antigens, and those with type O have neither antigen. The plasma of group O blood contains antibodies to A, B, and AB.
Group A plasma contains antibodies to B antigens, and group B plasma contains antibodies to A antigens. Group AB plasma has no antibodies to O, A, or B antigens. In blood transfusions, crossmatching is necessary to prevent agglutination of donor red cells by antibodies in the plasma of the recipient. Because plasma of groups A, B, and AB has no antibodies to group O erythrocytes, people with group O blood are called universal donors. Conversely, persons with AB blood are called universal recipients, because their plasma has no antibodies to the antigens of the other three groups. In addition to the ABO blood grouping, there are Rh (Rhesus factor)–positive and Rh-negative groups
An Rh-negative person can develop antibodies to Rh-positive red blood cells if exposed to Rh-positive blood. This can occur during pregnancy if the mother is Rh-negative and the fetus is Rh-positive (inherited from the father). In this case, Rh-positive red blood cells from the fetus enter the maternal bloodstream at the time of placental separation and induce Rh-positive antibodies in the mother’s plasma. The Rh-positive antibodies from the mother can also reach the fetus via the placenta and agglutinate and hemolyze fetal red blood cells (erythroblastosis fetalis, a hemolytic disease of the newborn). Red blood cell destruction can also occur in Rh-negative individuals who have previously had transfusions of Rh-positive blood and have developed Rh antibodies. If these individuals are given a subsequent transfusion of Rh-positive blood, the transfused red blood cells will be destroyed by the Rh antibodies in their plasma.
Summary
 The cardiovascular system is composed of a heart, which pumps blood, and blood vessels (arteries, capillaries, veins) that distribute the blood to all organs.
The cardiovascular system is composed of a heart, which pumps blood, and blood vessels (arteries, capillaries, veins) that distribute the blood to all organs.
The greatest resistance to blood flow, and hence the greatest pressure drop, in the arterial system occurs at the level of the small arteries and the arterioles.
Pulsatile pressure is progressively damped by the elasticity of the arteriolar walls and the functional resistance of the arterioles, so that capillary blood flow is essentially nonpulsatile.
Velocity of blood flow is inversely related to the cross-sectional area at any point along the vascular system.
Most of the blood volume in the systemic vascular bed is located in the venous side of the circulation.
Blood consists of red blood cells (erythrocytes), white blood cells (leukocytes and lymphocytes), and platelets, all suspended in a solution containing salts, proteins, carbohydrates, and lipids.
There are four major blood groups: O, A, B, and AB. Type O blood can be given to people with any of the blood groups because the plasma of all of the blood groups lacks antibodies to type O red cells. Hence people with type O blood are referred to as universal donors. By the same token, people with AB blood are referred to as universal recipients because their plasma lacks antibodies to red cells of all of the blood groups. In addition to O, A, B, and AB blood groups, there are Rh-positive and Rh-negative blood groups.
Adams R.H. Molecular control of arterial-venous blood vessel identity. J Anat. 2003;202:105.
Christensen K.L., Mulvany M.J. Location of resistance arteries. J Vasc Res. 2001;38:1.
Conway E.M., Collen D., Carmeliet P. Molecular mechanisms of blood vessel growth. Cardiovasc Res. 2001;49:507.
Pugsley M.K., Tabrizchi R. The vascular system. An overview of structure and function. J Pharmacol Toxicol Methods. 2000;44:333.
Secomb T.W., Pries A.R. The microcirculation: physiology at the mesoscale. J Physiol. 2011;589:1047.
Reid M.E., Lomas-Francis C. Molecular approaches to blood group identification. Curr Opin Hematol. 2002;9:152.
Urbaniak S.J., Greiss M.A. RhD haemolytic disease of the fetus and the newborn. Blood Rev. 2000;14:44.